The Dual Roles of Protein-Bound Solutes as Toxins and Signaling Molecules in Uremia


{kind=link}
{kind=link}
{kind=link}
Abstract
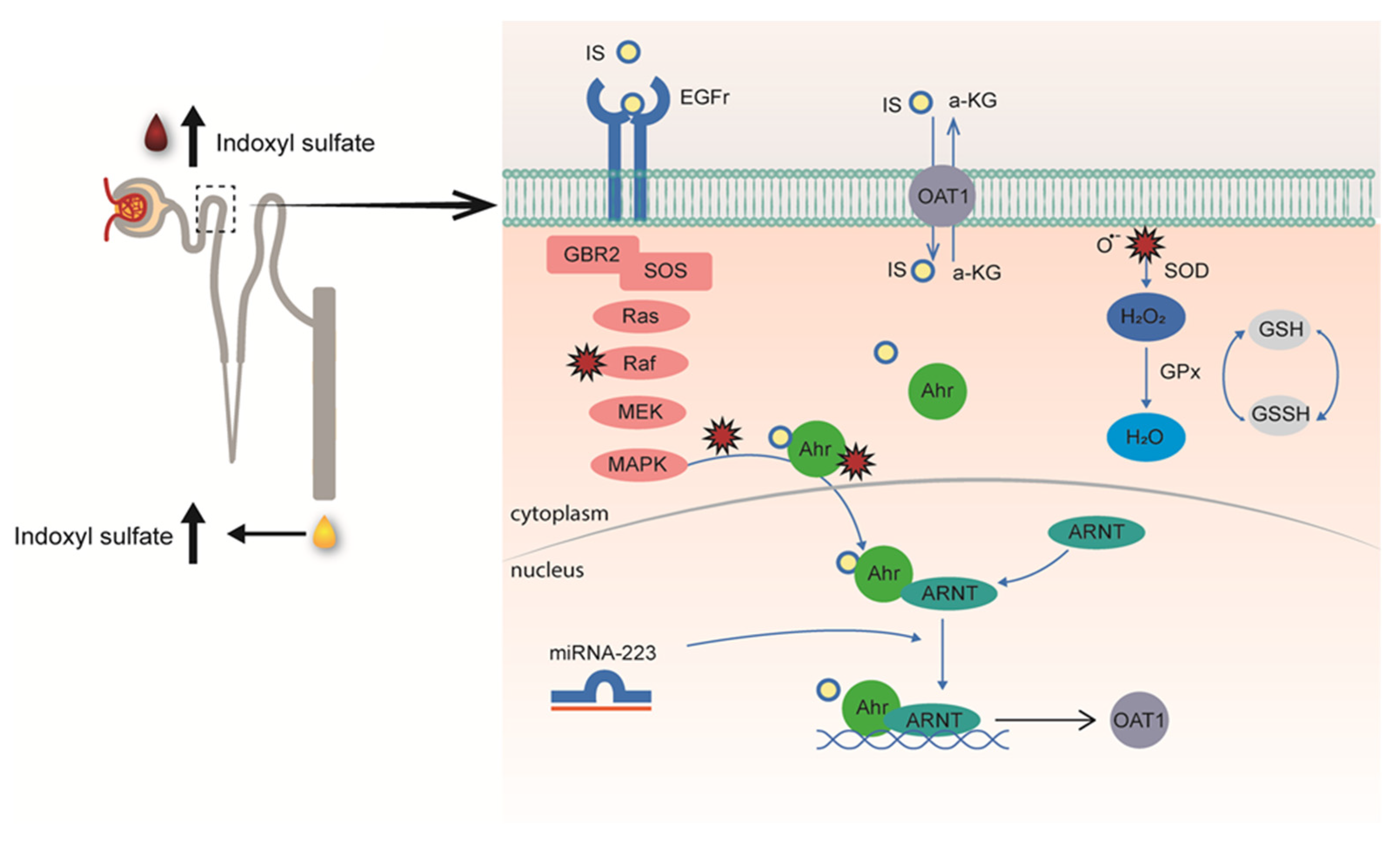
:1. Introduction
2. Dialysis Technology Advancements to Enhance Toxin Removal
3. Interventions to Reduce Plasma Levels of Uremic Toxins
4. Biohybrid Solutions
5. Remote Sensing and Signaling: Linking the Kidney to the Gut
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liyanage, T.; Ninomiya, T.; Jha, V.; Neal, B.; Patrice, H.M.; Okpechi, I.; Zhao, M.-h.; Lv, J.; Garg, A.X.; Knight, J.; et al. Worldwide access to treatment for end-stage kidney disease: A systematic review. Lancet 2015, 385, 1975–1982. [Google Scholar] [CrossRef]
- Webster, A.C.; Nagler, E.V.; Morton, R.L.; Masson, P. Chronic Kidney Disease. Lancet 2017, 389, 1238–1252. [Google Scholar] [CrossRef]
- Smith, H.W. The Kidney: Structure and Function in Health and Disease; Oxford University Press: Oxford, UK, 1951. [Google Scholar]
- Yen, S.-C.; Liu, Z.-W.; Juang, R.-S.; Sahoo, S.; Huang, C.-H.; Chen, P.; Hsiao, Y.-S.; Fang, J.-T. Carbon Nanotube/Conducting Polymer Hybrid Nanofibers as Novel Organic Bioelectronic Interfaces for Efficient Removal of Protein-Bound Uremic Toxins. ACS Appl. Mater. Interfaces 2019, 11, 43843–43856. [Google Scholar] [CrossRef]
- Krieter, D.H.; Devine, E.; Körner, T.; Rüth, M.; Wanner, C.; Raine, M.; Jankowski, J.; Lemke, H. Haemodiafiltration at increased plasma ionic strength for improved protein-bound toxin removal. Acta Physiol. 2016, 219, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Madero, M.; Cano, K.B.; Campos, I.; Tao, X.; Maheshwari, V.; Brown, J.; Cornejo, B.; Handelman, G.; Thijssen, S.; Kotanko, P. Removal of Protein-Bound Uremic Toxins during Hemodialysis Using a Binding Competitor. Clin. J. Am. Soc. Nephrol. 2019, 14, 394–402. [Google Scholar] [CrossRef]
- Etinger, A.; Kumar, S.R.; Ackley, W.; Soiefer, L.; Chun, J.; Singh, P.; Grossman, E.; Matalon, A.; Holzman, R.S.; Meijers, B.; et al. The effect of isohydric hemodialysis on the binding and removal of uremic retention solutes. PLoS ONE 2018, 13, e0192770. [Google Scholar] [CrossRef]
- Van Gelder, M.K.; Middel, I.R.; Vernooij, R.W.M.; Bots, M.L.; Verhaar, M.C.; Masereeuw, R.; Grooteman, M.P.; Nubé, M.J.; Dorpel, M.A.V.D.; Blankestijn, P.J.; et al. Protein-Bound Uremic Toxins in Hemodialysis Patients Relate to Residual Kidney Function, Are Not Influenced by Convective Transport, and Do Not Relate to Outcome. Toxins 2020, 12, 234. [Google Scholar] [CrossRef]
- Hill, K.; Walker, S.N.; Salminen, A.; Chung, H.L.; Li, X.; Ezzat, B.; Miller, J.J.; DesOrmeaux, J.S.; Zhang, J.; Hayden, A.; et al. Second Generation Nanoporous Silicon Nitride Membranes for High Toxin Clearance and Small Format Hemodialysis. Adv. Healthc. Mater. 2020, 9, e1900750. [Google Scholar] [CrossRef]
- Pavlenko, D.; Giasafaki, D.; Charalambopoulou, G.; Van Geffen, E.; Gerritsen, K.G.F.; Steriotis, T.; Stamatialis, D. Carbon Adsorbents With Dual Porosity for Efficient Removal of Uremic Toxins and Cytokines from Human Plasma. Sci. Rep. 2017, 7, 14914. [Google Scholar] [CrossRef]
- Geremia, I.; Stamatialis, D. Innovations in dialysis membranes for improved kidney replacement therapy. Nat. Rev. Nephrol. 2020, 16, 550–551. [Google Scholar] [CrossRef]
- De Pascale, M.; De Angelis, M.G.; Boi, C. Mixed Matrix Membranes Adsorbers (MMMAs) for the Removal of Uremic Toxins from Dialysate. Membranes 2022, 12, 203. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-C.; Wu, M.-Y.; Hu, P.-J.; Chen, T.-T.; Shen, W.-C.; Chang, W.-C.; Wu, M.-S. Effects and Safety of an Oral Adsorbent on Chronic Kidney Disease Progression: A Systematic Review and Meta-Analysis. J. Clin. Med. 2019, 8, 1718. [Google Scholar] [CrossRef]
- Nazzal, L.; Soiefer, L.; Chang, M.; Tamizuddin, F.; Schatoff, D.; Cofer, L.; Aguero-Rosenfeld, M.E.; Matalon, A.; Meijers, B.; Holzman, R.; et al. Effect of Vancomycin on the Gut Microbiome and Plasma Concentrations of Gut-Derived Uremic Solutes. Kidney Int. Rep. 2021, 6, 2122–2133. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.U.; Kim, H.W.; Kim, W. Effects of Probiotics, Prebiotics, and Synbiotics on Uremic Toxins, Inflammation, and Oxidative Stress in Hemodialysis Patients: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. J. Clin. Med. 2021, 10, 4456. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zelnick, L.R.; Wang, K.; Hoofnagle, A.N.; Becker, J.O.; Hsu, C.-Y.; Feldman, H.I.; Mehta, R.C.; Lash, J.P.; Waikar, S.S.; et al. Kidney Clearance of Secretory Solutes Is Associated with Progression of CKD: The CRIC Study. J. Am. Soc. Nephrol. 2020, 31, 817–827. [Google Scholar] [CrossRef] [PubMed]
- Legallais, C.; Kim, D.; Mihaila, S.M.; Mihajlovic, M.; Figliuzzi, M.; Bonandrini, B.; Salerno, S.; Yengej, F.A.Y.; Rookmaaker, M.B.; Romero, N.S.; et al. Bioengineering Organs for Blood Detoxification. Adv. Healc. Mater. 2018, 7, e1800430. [Google Scholar] [CrossRef]
- Chevtchik, N.V.; Mihajlovic, M.; Fedecostante, M.; Bolhuis-Versteeg, L.; Toraño, J.S.; Masereeuw, R.; Stamatialis, D. A bioartificial kidney device with polarized secretion of immune modulators. J. Tissue Eng. Regen. Med. 2018, 12, 1670–1678. [Google Scholar] [CrossRef]
- Jansen, J.; Fedecostante, M.; Wilmer, M.J.; Peters, J.G.; Kreuser, U.M.; van den Broek, P.H.; Mensink, R.; Boltje, T.J.; Stamatialis, D.; Wetzels, J.F.; et al. Bioengineered kidney tubules efficiently excrete uremic toxins. Sci. Rep. 2016, 6, 26715. [Google Scholar] [CrossRef]
- Marquez, I.O.; Tambra, S.; Luo, F.Y.; Li, Y.; Plummer, N.S.; Hostetter, T.H.; Meyer, T.W. Contribution of Residual Function to Removal of Protein-Bound Solutes in Hemodialysis. Clin. J. Am. Soc. Nephrol. 2010, 6, 290–296. [Google Scholar] [CrossRef]
- Masereeuw, R.; Mutsaers, H.A.; Toyohara, T.; Abe, T.; Jhawar, S.; Sweet, D.H.; Lowenstein, J. The Kidney and Uremic Toxin Removal: Glomerulus or Tubule? Semin. Nephrol. 2014, 34, 191–208. [Google Scholar] [CrossRef]
- Humes, H.; MacKay, S.M.; Funke, A.J.; Buffington, D.A. The bioartificial renal tubule assist device to enhance CRRT in acute renal failure. Am. J. Kidney Dis. 1997, 30, S28–S31. [Google Scholar] [CrossRef]
- Deguchi, T.; Kusuhara, H.; Takadate, A.; Endou, H.; Otagiri, M.; Sugiyama, Y. Characterization of uremic toxin transport by organic anion transporters in the kidney. Kidney Int. 2004, 65, 162–174. [Google Scholar] [CrossRef]
- Mutsaers, H.A.M.; Heuvel, L.P.V.D.; Ringens, L.H.J.; Dankers, A.C.A.; Russel, F.G.M.; Wetzels, J.F.M.; Hoenderop, J.G.; Masereeuw, R. Uremic Toxins Inhibit Transport by Breast Cancer Resistance Protein and Multidrug Resistance Protein 4 at Clinically Relevant Concentrations. PLoS ONE 2011, 6, e18438. [Google Scholar] [CrossRef] [PubMed]
- Jansen, J.; Jankowski, J.; Gajjala, P.R.; Wetzels, J.F.; Masereeuw, R. Disposition and clinical implications of protein-bound uremic toxins. Clin. Sci. 2017, 131, 1631–1647. [Google Scholar] [CrossRef]
- Nigam, S.K.; Bush, K.T. Uraemic syndrome of chronic kidney disease: Altered remote sensing and signalling. Nat. Rev. Nephrol. 2019, 15, 301–316. [Google Scholar] [CrossRef] [PubMed]
- Nigam, S.K.; Bush, K.T.; Bhatnagar, V.; Poloyac, S.M.; Momper, J.D. The Systems Biology of Drug Metabolizing Enzymes and Transporters: Relevance to Quantitative Systems Pharmacology. Clin. Pharmacol. Ther. 2020, 108, 40–53. [Google Scholar] [CrossRef]
- Jansen, J.; Jansen, K.; Neven, E.; Poesen, R.; Othman, A.; van Mil, A.; Sluijter, J.; Toraño, J.S.; Zaal, E.A.; Berkers, C.R.; et al. Remote sensing and signaling in kidney proximal tubules stimulates gut microbiome-derived organic anion secretion. Proc. Natl. Acad. Sci. USA 2019, 116, 16105–16110. [Google Scholar] [CrossRef]
- Poesen, R.; Mutsaers, H.; Windey, K.; Broek, P.H.V.D.; Verweij, V.; Augustijns, P.; Kuypers, D.R.; Jansen, J.; Evenepoel, P.; Verbeke, K.; et al. The Influence of Dietary Protein Intake on Mammalian Tryptophan and Phenolic Metabolites. PLoS ONE 2015, 10, e0140820. [Google Scholar] [CrossRef]
- Machado, T.S.; Poitevin, S.; Paul, P.; McKay, N.; Jourde-Chiche, N.; Legris, T.; Mouly-Bandini, A.; Dignat-George, F.; Brunet, P.; Masereeuw, R.; et al. Indoxyl Sulfate Upregulates Liver P-Glycoprotein Expression and Activity through Aryl Hydrocarbon Receptor Signaling. J. Am. Soc. Nephrol. 2017, 9, 906–918. [Google Scholar] [CrossRef]
- Caetano-Pinto, P.; Jamalpoor, A.; Ham, J.; Goumenou, A.; Mommersteeg, M.; Pijnenburg, D.; Ruijtenbeek, R.; Sanchez-Romero, N.; van Zelst, B.; Heil, S.G.; et al. Cetuximab Prevents Methotrexate-Induced Cytotoxicity in Vitro through Epidermal Growth Factor Dependent Regulation of Renal Drug Transporters. Mol. Pharm. 2017, 14, 2147–2157. [Google Scholar] [CrossRef]
- Fourdinier, O.; Schepers, E.; Meuth, V.M.-L.; Glorieux, G.; Liabeuf, S.; Verbeke, F.; Vanholder, R.; Brigant, B.; Pletinck, A.; Diouf, M.; et al. Serum levels of miR-126 and miR-223 and outcomes in chronic kidney disease patients. Sci. Rep. 2019, 9, 4477. [Google Scholar] [CrossRef] [PubMed]
- Naud, J.; Michaud, J.; Beauchemin, S.; Hébert, M.-J.; Roger, M.; Lefrancois, S.; Leblond, F.A.; Pichette, V. Effects of Chronic Renal Failure on Kidney Drug Transporters and Cytochrome P450 in Rats. Drug Metab. Dispos. 2011, 39, 1363–1369. [Google Scholar] [CrossRef] [PubMed]
- Lowenstein, J.; Nigam, S.K. Uremic Toxins in Organ Crosstalk. Front. Med. 2021, 8, 592602. [Google Scholar] [CrossRef]
- Kimura, H.; Tanaka, K.; Saito, H.; Iwasaki, T.; Oda, A.; Watanabe, S.; Kanno, M.; Shimabukuro, M.; Asahi, K.; Watanabe, T.; et al. Association of Polypharmacy with Kidney Disease Progression in Adults with CKD. Clin. J. Am. Soc. Nephrol. 2021, 16, 1797–1804. [Google Scholar] [CrossRef]
- Mihaila, S.M.; Faria, J.; Stefens, M.F.J.; Stamatialis, D.; Verhaar, M.C.; Gerritsen, K.G.F.; Masereeuw, R. Drugs Commonly Applied to Kidney Patients May Compromise Renal Tubular Uremic Toxins Excretion. Toxins 2020, 12, 391. [Google Scholar] [CrossRef]
- Van Biesen, W.; Eloot, S. Enhanced Removal of Protein-Bound Uremic Toxins Using Displacers. Clin. J. Am. Soc. Nephrol. 2019, 14, 324–326. [Google Scholar] [CrossRef]
- Meijers, B.; Evenepoel, P.; Anders, H.-J. Intestinal microbiome and fitness in kidney disease. Nat. Rev. Nephrol. 2019, 15, 531–545. [Google Scholar] [CrossRef]
- Rukavina Mikusic, N.L.; Kouyoumdzian, N.M.; Choi, M.R. Gut microbiota and chronic kidney disease: Evidences and mechanisms that mediate a new communication in the gastrointestinal-renal axis. Pflug. Arch. Eur. J. Physiol. 2020, 472, 303–320. [Google Scholar] [CrossRef]
- De Andrade, L.S.; Dalboni, M.A.; De Carvalho, J.T.G.; Grabulosa, C.C.; Pereira, N.B.F.; Aoike, D.T.; Cuppari, L. In vitro effect of uremic serum on barrier function and inflammation in human colonocytes. J. Bras. Nefrol. 2018, 40, 217–224. [Google Scholar] [CrossRef]
- Ramezani, A.; Massy, Z.A.; Meijers, B.; Evenepoel, P.; Vanholder, R.; Raj, D.S. Role of the Gut Microbiome in Uremia: A Potential Therapeutic Target. Am. J. Kidney Dis. 2016, 67, 483–498. [Google Scholar] [CrossRef]
- Giordano, L.; Mihaila, S.M.; Amirabadi, H.E.; Masereeuw, R. Microphysiological Systems to Recapitulate the Gut–Kidney Axis. Trends Biotechnol. 2021, 39, 811–823. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masereeuw, R. The Dual Roles of Protein-Bound Solutes as Toxins and Signaling Molecules in Uremia. Toxins 2022, 14, 402. https://doi.org/10.3390/toxins14060402
Masereeuw R. The Dual Roles of Protein-Bound Solutes as Toxins and Signaling Molecules in Uremia. Toxins. 2022; 14(6):402. https://doi.org/10.3390/toxins14060402
Chicago/Turabian StyleMasereeuw, Rosalinde. 2022. "The Dual Roles of Protein-Bound Solutes as Toxins and Signaling Molecules in Uremia" Toxins 14, no. 6: 402. https://doi.org/10.3390/toxins14060402
APA StyleMasereeuw, R. (2022). The Dual Roles of Protein-Bound Solutes as Toxins and Signaling Molecules in Uremia. Toxins, 14(6), 402. https://doi.org/10.3390/toxins14060402