Implications of Crop Rotation and Fungicide on Fusarium and Mycotoxin Spectra in Manitoba Barley, 2017–2019


Abstract
:1. Introduction
2. Results
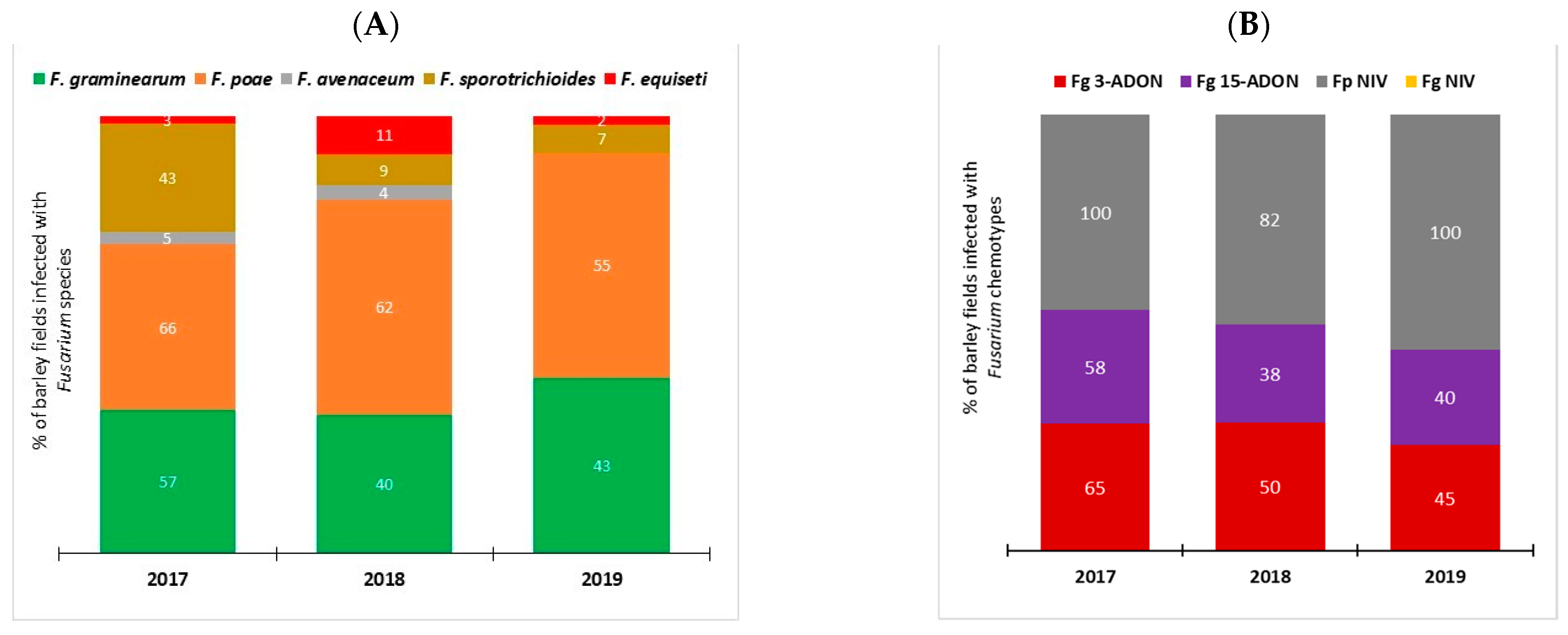
2.1. Fusarium Species Spectra and Chemotype Composition
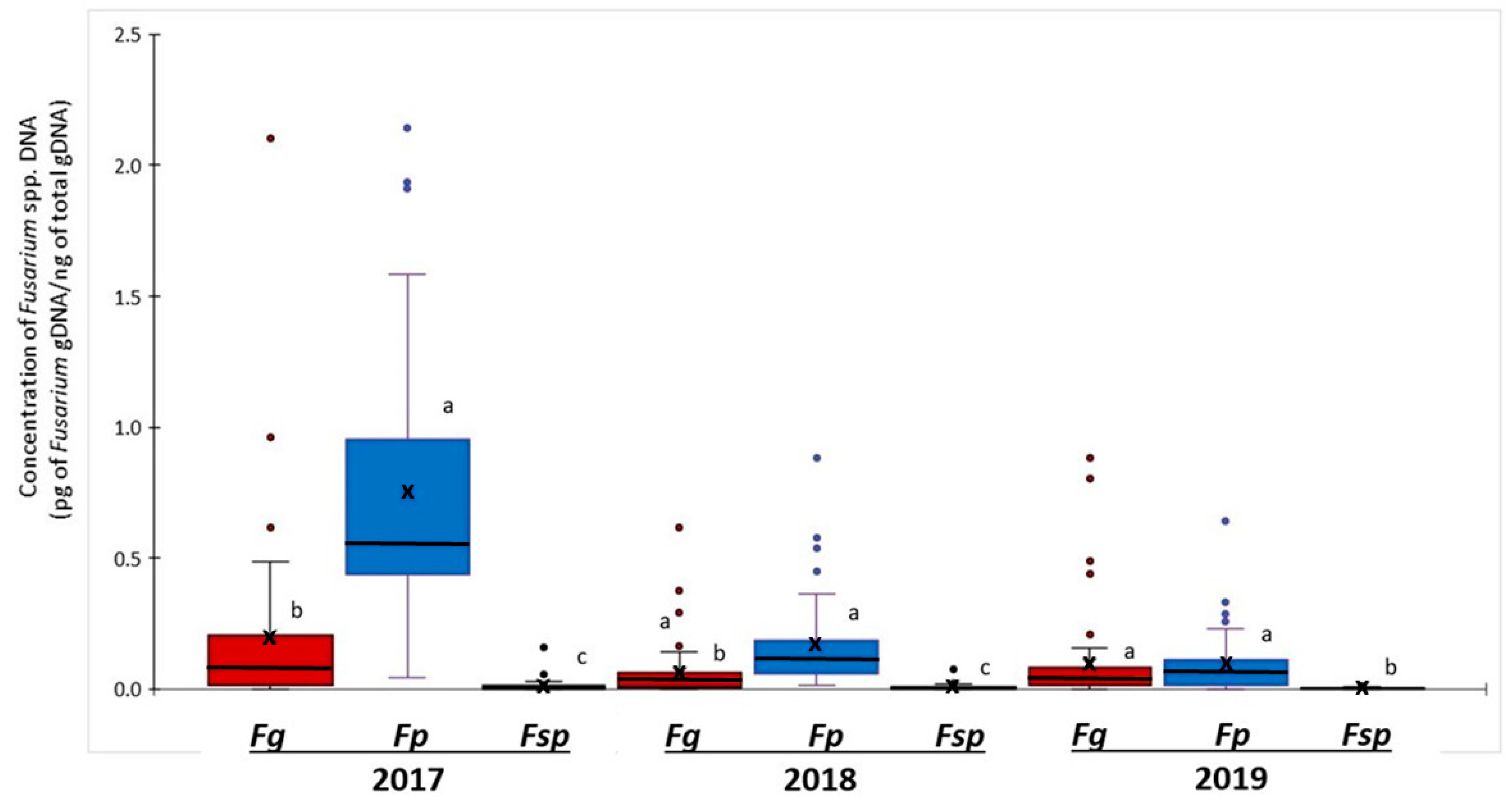
2.2. Abundance of Fusarium DNA
2.3. Fusarium Mycotoxins
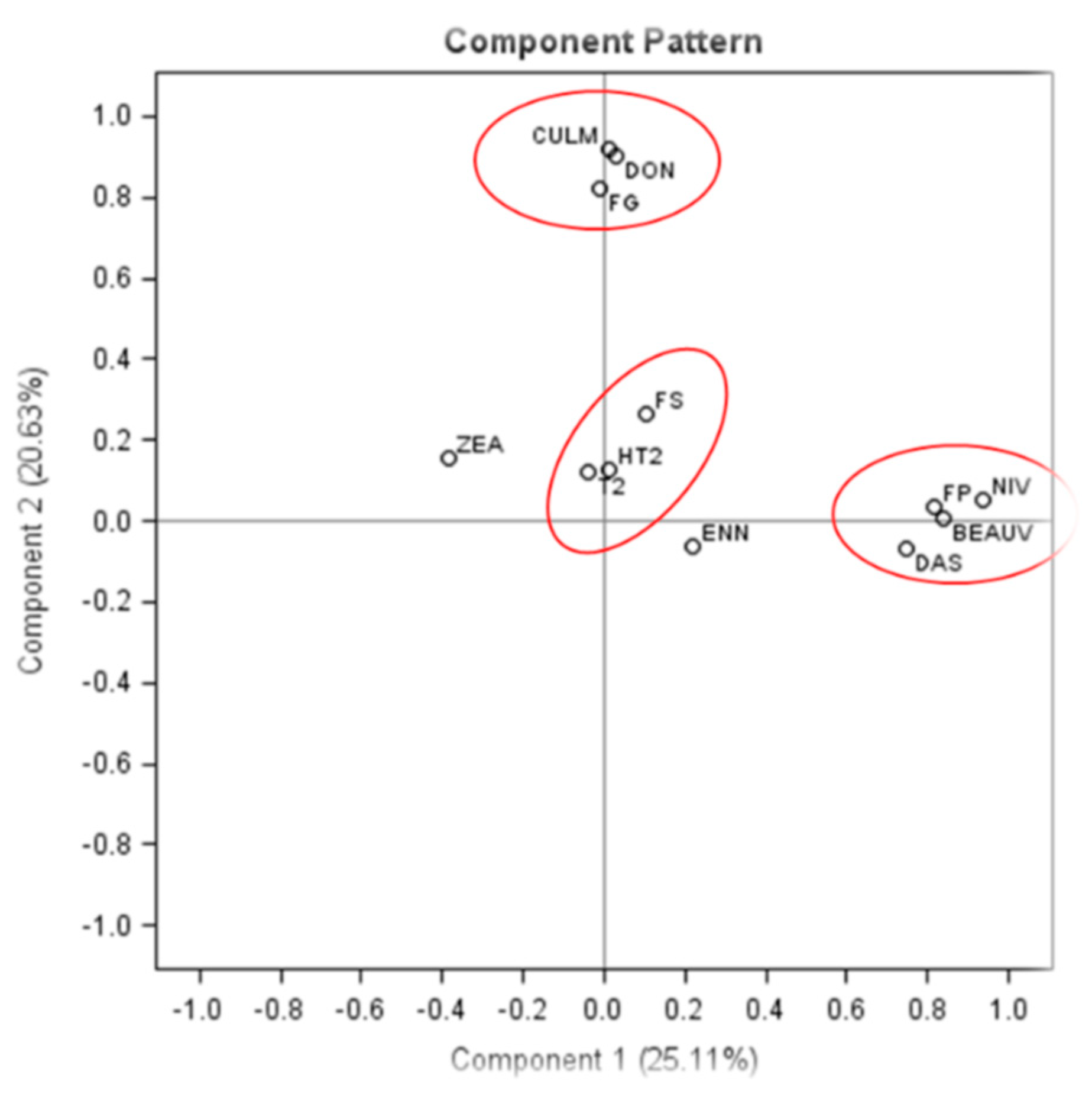
2.4. PCA of the Concentrations of Fusarium gDNA and Mycotoxins in Barley Grains
2.5. Sensitivity of F. poae and F. graminearum Strains to Triazole Fungicides
2.6. Effects of Preceding Crop and Year on Fusarium Mycotoxins in Barley Grains
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Barley Sample Collection
5.2. DNA Extraction and PCR Analysis of Fusarium Genomic DNA
5.3. Detection and Quantification of Fusarium Mycotoxins
5.4. Sensitivity of F. poae and F. graminearum to Triazole Fungicides
5.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ames, N.P.; Rhymer, C.R. Issues surrounding health claims for barley. J. Nutr. 2008, 138, 1237S–1243S. [Google Scholar] [CrossRef] [Green Version]
- Fernando, W.D.; Oghenekaro, A.O.; Tucker, J.R.; Badea, A. Building on a foundation: Advances in epidemiology, resistance breeding, and forecasting research for reducing the impact of Fusarium head blight in wheat and barley. Can. J. Plant Pathol. 2021, 43, 495–526. [Google Scholar] [CrossRef]
- He, X.; Osman, M.; Helm, J.; Capettini, F.; Singh, P.K. Evaluation of Canadian barley breeding lines for Fusarium head blight resistance. Can. J. Plant Sci. 2015, 95, 923–929. [Google Scholar] [CrossRef] [Green Version]
- Tekauz, A.; McCallum, B.; Gilbert, J. Review: Fusarium head blight of barley in western Canada. Can. J. Plant Pathol. 2009, 2000, 9–16. [Google Scholar] [CrossRef]
- Salas, B.; Steffenson, B.J.; Casper, H.H.; Tacke, B.; Prom, L.K.; Fetch, T.G., Jr.; Schwarz, P.B. Fusarium species pathogenic to barley and their associated mycotoxins. Plant Dis. 1999, 83, 667–674. [Google Scholar] [CrossRef] [Green Version]
- Foroud, N.A.; Baines, D.; Gagkaeva, T.Y.; Thakor, N.; Badea, A.; Steiner, B.; Bürstmayr, M.; Bürstmayr, H. Trichothecenes in cereal grains—An update. Toxins 2019, 11, 634. [Google Scholar] [CrossRef] [Green Version]
- Drakopoulos, D.; Sulyok, M.; Krska, R.; Logrieco, A.F.; Vogelgsang, S. Raised concerns about the safety of barley grains and straw: A Swiss survey reveals a high diversity of mycotoxins and other fungal metabolites. Food Control 2021, 125, 107919. [Google Scholar] [CrossRef]
- D’Mello, J.P.F.; Placinta, C.M.; Macdonald, A.M.C. Fusarium mycotoxins: A review of global implications for animal health, welfare and productivity. Anim. Feed. Sci. Technol. 1999, 80, 183–205. [Google Scholar] [CrossRef] [Green Version]
- Pascari, X.; Marin, S.; Ramos, A.J.; Sanchis, V. Relevant Fusarium mycotoxins in malt and beer. Foods 2022, 11, 246. [Google Scholar] [CrossRef]
- Martínez, M.; Ramirez Albuquerque, L.D.; Dinolfo, M.I.; Biganzoli, F.; F Pinto, V.; Stenglein, S.A. Effects of Fusarium graminearum and Fusarium poae on disease parameters, grain quality and mycotoxin contamination in barley (part II). J. Sci. Food Agric. 2020, 100, 3182–3191. [Google Scholar] [CrossRef]
- Sarlin, T.; Kivioja, T.; Kalkkinen, N.; Linder, M.B.; Nakari-Setälä, T. Identification and characterization of gushing-active hydrophobins from Fusarium graminearum and related species. J. Basic Microbiol. 2012, 52, 184–194. [Google Scholar] [CrossRef]
- Becher, R.; Miedaner, T.; Wirsel, S.G.R. 8 Biology, diversity, and management of FHB-causing Fusarium species in small grain cereals. In Agricultural Applications; Kempken, F., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 199–241. [Google Scholar]
- Wilson, W.W.; Dahl, B.; Nganje, W.E. Economic costs of Fusarium Head Blight, scab and deoxynivalenol. World Mycotoxin J. 2018, 11, 291–302. [Google Scholar] [CrossRef]
- Castañares, E.; Albuquerque, D.R.; Dinolfo, M.I.; Pinto, V.F.; Patriarca, A.; Stenglein, S.A. Trichothecene genotypes and production profiles of Fusarium graminearum isolates obtained from barley cultivated in Argentina. Int. J. Food Microbiol. 2014, 179, 57–63. [Google Scholar] [CrossRef]
- Spolti, P.; Del Ponte, E.M.; Cummings, J.A.; Dong, Y.; Bergstrom, G.C. Fitness attributes of Fusarium graminearum isolates from wheat in New York possessing a 3-ADON or 15-ADON trichothecene genotype. Phytopathology 2014, 104, 513–519. [Google Scholar] [CrossRef] [Green Version]
- Valverde-Bogantes, E.; Bianchini, A.; Herr, J.R.; Rose, D.J.; Wegulo, S.N.; Hallen-Adams, H.E. Recent population changes of Fusarium head blight pathogens: Drivers and implications. Can. J. Plant Pathol. 2020, 42, 315–329. [Google Scholar] [CrossRef]
- Liang, J.M.; Xayamongkhon, H.; Broz, K.; Dong, Y.; McCormick, S.P.; Abramova, S.; Ward, T.J.; Ma, Z.H.; Kistler, H.C. Temporal dynamics and population genetic structure of Fusarium graminearum in the upper Midwestern United States. Fungal Genet. Biol. 2014, 73, 83–92. [Google Scholar] [CrossRef]
- Tanaka, T.; Hasegawa, A.; Yamamoto, S.; Lee, U.S.; Sugiura, Y.; Ueno, Y. Worldwide contamination of cereals by the Fusarium mycotoxins nivalenol, deoxynivalenol, and zearalenone. 1. Survey of 19 countries. J. Agric. Food Chem. 1988, 36, 979–983. [Google Scholar] [CrossRef]
- Amarasinghe, C.C.; Tittlemier, S.A.; Fernando, W.G.D. Nivalenol-producing Fusarium cerealis associated with fusarium head blight in winter wheat in Manitoba, Canada. Plant Pathol. 2015, 64, 988–995. [Google Scholar] [CrossRef]
- Banik, M.; Beyene, M.; Wang, X. Fusarium head blight of barley in Manitoba—2015. Can. Plant Dis. Surv. 2016, 96, 93–94. [Google Scholar]
- Banik, M.; Beyene, M.; Wang, X. Fusarium head blight of barley in Manitoba—2016. Can. Plant Dis. Surv. 2017, 97, 100–101. [Google Scholar]
- Nielsen, L.K.; Cook, D.J.; Edwards, S.G.; Ray, R.V. The prevalence and impact of Fusarium head blight pathogens and mycotoxins on malting barley quality in UK. Int. J. Food Microbiol. 2014, 179, 38–49. [Google Scholar] [CrossRef] [Green Version]
- Sumíková, T.; Chrpová, J.; Džuman, Z.; Salava, J.; Štěrbová, L.; Palicová, J.; Slavíková, P.; Stránská-Zachariášová, M.; Hajšlová, J. Mycotoxins content and its association with changing patterns of Fusarium pathogens in wheat in the Czech Republic. World Mycotoxin J. 2017, 10, 143–151. [Google Scholar] [CrossRef]
- Nogueira, M.S.; Decundo, J.; Martinez, M.; Dieguez, S.N.; Moreyra, F.; Moreno, M.V.; Stenglein, S.A. Natural contamination with mycotoxins produced by Fusarium graminearum and Fusarium poae in malting barley in Argentina. Toxins 2018, 10, 78. [Google Scholar] [CrossRef] [Green Version]
- Uhlig, S.; Torp, M.; Heier, B.T. Beauvericin and enniatins A, A1, B and B1 in Norwegian grain: A survey. Food Chem. 2006, 94, 193–201. [Google Scholar] [CrossRef]
- Liu, K.; Howell, D.N.; Perfect, J.R.; Schell, W.A. Morphologic criteria for the preliminary identification of Fusarium, Paecilomyces, and Acremonium species by histopathology. Am. J. Clin. Pathol. 1998, 109, 45–54. [Google Scholar] [CrossRef] [Green Version]
- Brennan, J.M.; Fagan, B.; van Maanen, A.; Cooke, B.M.; Doohan, F.M. Studies on in vitro growth and pathogenicity of European Fusarium fungi. Eur. J. Plant Pathol. 2004, 109, 577–587. [Google Scholar] [CrossRef]
- Stack, R.W.; McMullen, M.P. Head blighting potential of Fusarium species associated with spring wheat heads. Can. J. Plant Pathol. 1985, 7, 79–82. [Google Scholar] [CrossRef]
- Xue, A.G.; Chen, Y.H.; Seifert, K.; Guo, W.; Blackwell, B.A.; Harris, L.J.; Overy, D.P. Prevalence of Fusarium species causing head blight of spring wheat, barley and oat in Ontario during 2001–2017. Can. J. Plant Pathol. 2019, 41, 392–402. [Google Scholar] [CrossRef]
- González, H.H.; Moltó, G.A.; Pacin, A.; Resnik, S.L.; Zelaya, M.J.; Masana, M.; Martínez, E.J. Trichothecenes and mycoflora in wheat harvested in nine locations in Buenos Aires province, Argentina. Mycopathologia 2008, 165, 105–114. [Google Scholar] [CrossRef]
- Oghenekaro, A.O.; Oviedo-Ludena, M.A.; Serajazari, M.; Wang, X.; Henriquez, M.A.; Wenner, N.G.; Kuldau, G.A.; Navabi, A.; Kutcher, H.R.; Fernando, W.G.D. Population genetic structure and chemotype diversity of Fusarium graminearum populations from wheat in Canada and North Eastern United States. Toxins 2021, 13, 180. [Google Scholar] [CrossRef]
- Kelly, A.C.; Clear, R.M.; O’Donnell, K.; McCormick, S.; Turkington, T.K.; Tekauz, A.; Gilbert, J.; Kistler, H.C.; Busman, M.; Ward, T.J. Diversity of Fusarium head blight populations and trichothecene toxin types reveals regional differences in pathogen composition and temporal dynamics. Fungal Genet. Biol. 2015, 82, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Lozowicka, B.; Iwaniuk, P.; Konecki, R.; Kaczynski, P.; Kuldybayev, N.; Dutbayev, Y. Impact of diversified chemical and biostimulator protection on yield, health Status, mycotoxin level, and economic profitability in spring wheat (Triticum aestivum L.) cultivation. Agronomy 2022, 12, 258. [Google Scholar] [CrossRef]
- Browne, R.A.; Cooke, B.M. Resistance of wheat to Fusarium spp. in an in vitro seed germination assay and preliminary investigations into the relationship with Fusarium head blight resistance. Euphytica 2005, 141, 23–32. [Google Scholar] [CrossRef]
- Xue, A.G.; Chen, Y.H.; Marchand, G.; Guo, W.; Ren, C.Z.; Savard, M.; McElroy, R.B. Timing of inoculation and Fusarium species affect the severity of Fusarium head blight on oat. Can. J. Plant Sci. 2015, 95, 517–524. [Google Scholar] [CrossRef]
- Tan, J.; Ameye, M.; Landschoot, S.; De Zutter, N.; De Saeger, S.; De Boevre, M.; Abdallah, M.F.; Van der Lee, T.; Waalwijk, C.; Audenaert, K. At the scene of the crime: New insights into the role of weakly pathogenic members of the Fusarium head blight disease complex. Mol. Plant Pathol. 2020, 21, 1559–1572. [Google Scholar] [CrossRef]
- Islam, M.N.; Tabassum, M.; Banik, M.; Daayf, F.; Fernando, W.; Harris, L.J.; Sura, S.; Wang, X. Naturally occurring Fusarium species and mycotoxins in oat grains from Manitoba, Canada. Toxins 2021, 13, 670. [Google Scholar] [CrossRef]
- Vogelgsang, S.; Beyer, M.; Pasquali, M.; Jenny, E.; Musa, T.; Bucheli, T.D.; Wettstein, F.E.; Forrer, H.-R. An eight-year survey of wheat shows distinctive effects of cropping factors on different Fusarium species and associated mycotoxins. Eur. J. Agron. 2019, 105, 62–77. [Google Scholar] [CrossRef]
- Gilbert, J.; Tekauz, A. Review: Recent developments in research on Fusarium head blight of wheat in Canada. Can. J. Plant Pathol. 2000, 22, 1–8. [Google Scholar] [CrossRef]
- Schöneberg, T.; Kibler, K.; Wettstein, F.E.; Bucheli, T.D.; Forrer, H.R.; Musa, T.; Mascher, F.; Bertossa, M.; Keller, B.; Vogelgsang, S. Influence of temperature, humidity duration and growth stage on the infection and mycotoxin production by Fusarium langsethiae and Fusarium poae in oats. Plant Pathol. 2019, 68, 173–184. [Google Scholar] [CrossRef] [Green Version]
- Nazari, L.; Pattori, E.; Manstretta, V.; Terzi, V.; Morcia, C.; Somma, S.; Moretti, A.; Ritieni, A.; Rossi, V. Effect of temperature on growth, wheat head infection, and nivalenol production by Fusarium poae. Food Microbiol. 2018, 76, 83–90. [Google Scholar] [CrossRef]
- Yli-Mattila, T.; Paavanen-Huhtala, S.; Jestoi, M.; Parikka, P.; Hietaniemi, V.; Gagkaeva, T.; Sarlin, T.; Haikara, A.; Laaksonen, S.; Rizzo, A. Real-time PCR detection and quantification of Fusarium poae, F. graminearum, F. sporotrichioides and F. langsethiae in cereal grains in Finland and Russia. Arch. Phytopathol. Plant Prot. 2008, 41, 243–260. [Google Scholar] [CrossRef]
- Tucker, J.R.; Badea, A.; Blagden, R.; Pleskach, K.; Tittlemier, S.A.; Fernando, W.G.D. Deoxynivalenol-3-glucoside content is highly associated with deoxynivalenol levels in two-row barley genotypes of importance to Canadian barley breeding programs. Toxins 2019, 11, 319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polišenská, I.; Jirsa, O.; Vaculová, K.; Pospíchalová, M.; Wawroszova, S.; Frydrych, J. Fusarium mycotoxins in two hulless oat and barley cultivars used for food purposes. Foods 2020, 9, 1037. [Google Scholar] [CrossRef] [PubMed]
- Tekauz, A.; Mitchell Fetch, J.; Rossnagel, B.; Savard, M. Progress in assessing the impact of Fusarium head blight on oat in western Canada and screening of avena germplasm for resistance. Cereal Res. Commun. 2008, 36, 49–56. [Google Scholar] [CrossRef]
- Gilbert, J.; Clear, R.M.; Ward, T.J.; Gaba, D.; Tekauz, A.; Turkington, T.K.; Woods, S.M.; Nowicki, T.; O’Donnell, K. Relative aggressiveness and production of 3- or 15-acetyl deoxynivalenol and deoxynivalenol by Fusarium graminearum in spring wheat. Can. J. Plant Pathol. 2010, 32, 146–152. [Google Scholar] [CrossRef]
- Guo, X.W.; Fernando, W.G.D.; Seow-Brock, H.Y. Population structure, chemotype diversity, and potential chemotype shifting of Fusarium graminearum in wheat fields of Manitoba. Plant Dis. 2008, 92, 756–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lattanzio, V.M.; Ciasca, B.; Terzi, V.; Ghizzoni, R.; McCormick, S.P.; Pascale, M. Study of the natural occurrence of T-2 and HT-2 toxins and their glucosyl derivatives from field barley to malt by high-resolution Orbitrap mass spectrometry. Food Addit. Contam. Part A 2015, 32, 1647–1655. [Google Scholar] [CrossRef]
- Kiš, M.; Vulić, A.; Kudumija, N.; Šarkanj, B.; Jaki Tkalec, V.; Aladić, K.; Škrivanko, M.; Furmeg, S.; Pleadin, J. A two-year occurrence of Fusarium T-2 and HT-2 toxin in Croatian cereals relative of the regional weather. Toxins 2021, 13, 39. [Google Scholar] [CrossRef]
- Clear, R.M.; Patrick, S.K.; Platford, R.G.; Desjardins, M. Occurrence and distribution of Fusarium species in barley and oat seed from Manitoba in 1993 and 1994. Can. J. Plant Pathol. 1996, 18, 409–414. [Google Scholar] [CrossRef]
- Tini, F.; Beccari, G.; Onofri, A.; Ciavatta, E.; Gardiner, D.M.; Covarelli, L. Fungicides may have differential efficacies towards the main causal agents of Fusarium head blight of wheat. Pest Manag. Sci. 2020, 76, 3738–3748. [Google Scholar] [CrossRef]
- Audenaert, K.; Landschoot, S.; Vanheule, A.; Waegeman, W.; De Baets, B.; Haesaert, G. Impact of fungicide timing on the composition of the Fusarium head blight disease complex and the presence of deoxynivalenol (DON) in wheat. In Fungicides—Beneficial and Harmful Aspects; Thajuddin, N., Ed.; IntechOpen: Rijeka, Croatia, 2011; pp. 79–98. [Google Scholar]
- Schöneberg, T.; Martin, C.; Wettstein, F.E.; Bucheli, T.D.; Mascher, F.; Bertossa, M.; Musa, T.; Keller, B.; Vogelgsang, S. Fusarium and mycotoxin spectra in Swiss barley are affected by various cropping techniques. Food Addit. Contam. Part A 2016, 33, 1608–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pageau, D.; Lafond, J.; Lajeunesse, J.; Savard, M. Impact of previous cultivation and nitrogen fertilization on deoxynivalenol content in barley. Can. J. Plant Pathol. 2008, 30, 397–403. [Google Scholar] [CrossRef]
- Booth, C. The present status of Fusarium taxonomy. Annu. Rev. Phytopathol. 1975, 13, 83–93. [Google Scholar] [CrossRef]
- Parry, D.W.; Jenkinson, P.; McLeod, L. Fusarium ear blight (SCAB) in small-grain cereals—A review. Plant Pathol. 2007, 44, 207–238. [Google Scholar] [CrossRef]
- Stenglein, S. Fusarium poae: A pathogen that needs more attention. J. Plant Pathol. 2009, 91, 25–36. [Google Scholar] [CrossRef]
- Ameye, M.; Audenaert, K.; De Zutter, N.; Steppe, K.; Vanhaecke, L.; De Vleesschauwer, D.; Haesaert, G.; Smagghe, G. Priming of wheat with the green leaf volatile Z-3-hexenyl acetate enhances defense against Fusarium graminearum but boosts deoxynivalenol production. Plant Physiol. 2015, 167, 1671–1684. [Google Scholar] [CrossRef] [Green Version]
- Walkowiak, S.; Bonner, C.T.; Wang, L.; Blackwell, B.; Rowland, O.; Subramaniam, R. Intraspecies Interaction of Fusarium graminearum contributes to reduced toxin production and virulence. Mol. Plant Microbe Interact. 2015, 28, 1256–1267. [Google Scholar] [CrossRef] [Green Version]
- Cheat, S.; Pinton, P.; Cossalter, A.; Cognie, J.; Vilariño, M.; Callu, P.; Raymond-Letron, I.; Oswald, I.P.; Kolf-Clauw, M. The mycotoxins deoxynivalenol and nivalenol show in vivo synergism on jejunum enterocytes apoptosis. Food Chem. Toxicol. 2016, 87, 45–54. [Google Scholar] [CrossRef]
- Demeke, T.; Clear, R.M.; Patrick, S.K.; Gaba, D. Species-specific PCR-based assays for the detection of Fusarium species and a comparison with the whole seed agar plate method and trichothecene analysis. Int. J. Food Microbiol. 2005, 103, 271–284. [Google Scholar] [CrossRef]
- Nicolaisen, M.; Supronienė, S.; Nielsen, L.K.; Lazzaro, I.; Spliid, N.H.; Justesen, A.F. Real-time PCR for quantification of eleven individual Fusarium species in cereals. J. Microbiol. Methods 2009, 76, 234–240. [Google Scholar] [CrossRef]
- Ward, T.J.; Clear, R.M.; Rooney, A.P.; O’Donnell, K.; Gaba, D.; Patrick, S.; Starkey, D.E.; Gilbert, J.; Geiser, D.M.; Nowicki, T.W. An adaptive evolutionary shift in Fusarium head blight pathogen populations is driving the rapid spread of more toxigenic Fusarium graminearum in North America. Fungal Genet. Biol. 2008, 45, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Dinolfo, M.I.; Barros, G.G.; Stenglein, S.A. Development of a PCR assay to detect the potential production of nivalenol in Fusarium poae. FEMS Microbiol. Lett. 2012, 332, 99–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Year | 2017 | 2018 | 2019 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Samples | 44 | 47 | 58 | |||||||||
| Mycotoxins | Mean (µg kg−1) | Maximum (µg kg−1) | % of Samples above LOD | % of Samples above 500 (µg kg−1) | Mean (µg kg−1) | Maximum (µg kg−1) | % of Samples above LOD | % of samples above 500 (µg kg−1) | Mean (µg kg−1) | Maximum (µg kg−1) | % of Samples above LOD | % of Samples above 500 (µg kg−1) |
| NIV | 597.7 | 4136.6 | 95.5 | 29.5 | 219.1 | 3252.1 | 91.1 | 16.7 | 412.4 | 2245.4 | 71.2 | 22.4 |
| DON | 264.7 | 8921.6 | 54.5 | 6.8 | 56.7 | 1488.8 | 31.1 | 4.5 | 65.3 | 2051.1 | 25.4 | 3.4 |
| DON-3G | 87.6 | 3117.4 | 53.5 | 2.3 | 3.9 | 20.5 | 20.5 | - | 18.9 | 443.3 | 23.7 | - |
| 3-ADON | 2.8 | 79.9 | 13.9 | - | 3.2 | 98.8 | 6.8 | - | 2.3 | 87.2 | 3 | - |
| 15-ADON | 0.9 | 32.8 | 2.3 | - | 1.1 | 44.7 | 2.3 | - | 2.1 | 111.8 | 1.7 | - |
| BEAUV | 9.3 | 86.8 | 74.7 | - | 10.4 | 211.4 | 59.1 | - | 14.5 | 122.5 | 47.5 | - |
| HT-2 | 9.9 | 120.7 | 11.4 | - | 0.3 | 51.5 | 4.3 | - | 5.1 | 278.1 | 6.7 | - |
| T-2 | 2.8 | 108.8 | 6.8 | - | 0.3 | 45.5 | 4.3 | - | 2.7 | 137.1 | 6.7 | - |
| ENNs | 21.7 | 371.8 | 21.5 | - | 14.7 | 266.1 | 18.2 | - | 27.4 | 1046.5 | 5.1 | 3.4 |
| CULM | 10.7 | 298.3 | 27.3 | - | 2.5 | 65.1 | 11.4 | - | 15.8 | 385.2 | 27.1 | - |
| DAS | 0.9 | 25.7 | 4.5 | - | 0.3 | 12.7 | 4.3 | - | 0.3 | 10.7 | 3.4 | - |
| ZEA | 2.4 | 93.5 | 9.1 | - | 0.3 | 15.2 | 6.3 | - | 0.3 | 2.5 | 3.4 | - |
| % Inhibition of Mycelial Growth of Fusarium Species | ||||||||
|---|---|---|---|---|---|---|---|---|
| Fungicides | Caramba (metconazole) | Prosaro (prothioconazole + tebuconazole) | Folicur (tebuconazole) | Proline (prothioconazole) | ||||
| Dose (mg ai/L) | 0.01 | 0.1 | 0.01 | 0.1 | 0.01 | 0.1 | 0.01 | 0.1 |
| Fusarium strains | Mean | Mean | Mean | Mean | Mean | Mean | Mean | Mean |
| F. graminearum | 43 ± 5.1 a* | 55 ± 4.6 a | 39 ± 2.6 a** | 51 ± 3.3 a** | 2.5 ± 1.1 a* | 38 ± 3.2 a* | 28 ± 1.9 a* | 42 ± 2.2 a** |
| F. poae | 34 ± 3.2 b | 59 ± 6.2 a | 25 ± 1.9 b | 29 ± 1.8 b | 14 ± 0.5 b | 28 ± 2.1 b | 21 ± 1.4 b | 29 ± 2.1 b |
| Fixed Effects | p-Value | |||
| DON | NIV | |||
| Previous Crop (PC) | 0.01 * | 0.34 | ||
| Year (2017–2019) | 0.10 | 0.15 | ||
| PC * Year | 0.02 * | 0.29 | ||
| Previous Crop | DON (mg kg−1) | NIV (mg kg−1) | ||
| Mean | SE | Mean | SE | |
| Canola (n = 73) | 0.06 b | 0.02 | 0.41 a | 0.07 |
| Cereal (n = 28) | 0.23 a | 0.07 | 0.52 a | 0.10 |
| Flax (n = 17) | 0.14 ab | 0.04 | 0.29 a | 0.08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, M.N.; Banik, M.; Sura, S.; Tucker, J.R.; Wang, X. Implications of Crop Rotation and Fungicide on Fusarium and Mycotoxin Spectra in Manitoba Barley, 2017–2019. Toxins 2022, 14, 463. https://doi.org/10.3390/toxins14070463
Islam MN, Banik M, Sura S, Tucker JR, Wang X. Implications of Crop Rotation and Fungicide on Fusarium and Mycotoxin Spectra in Manitoba Barley, 2017–2019. Toxins. 2022; 14(7):463. https://doi.org/10.3390/toxins14070463
Chicago/Turabian StyleIslam, M. Nazrul, Mitali Banik, Srinivas Sura, James R. Tucker, and Xiben Wang. 2022. "Implications of Crop Rotation and Fungicide on Fusarium and Mycotoxin Spectra in Manitoba Barley, 2017–2019" Toxins 14, no. 7: 463. https://doi.org/10.3390/toxins14070463
APA StyleIslam, M. N., Banik, M., Sura, S., Tucker, J. R., & Wang, X. (2022). Implications of Crop Rotation and Fungicide on Fusarium and Mycotoxin Spectra in Manitoba Barley, 2017–2019. Toxins, 14(7), 463. https://doi.org/10.3390/toxins14070463