Deoxynivalenol Biosynthesis in Fusarium pseudograminearum Significantly Repressed by a Megabirnavirus

Abstract
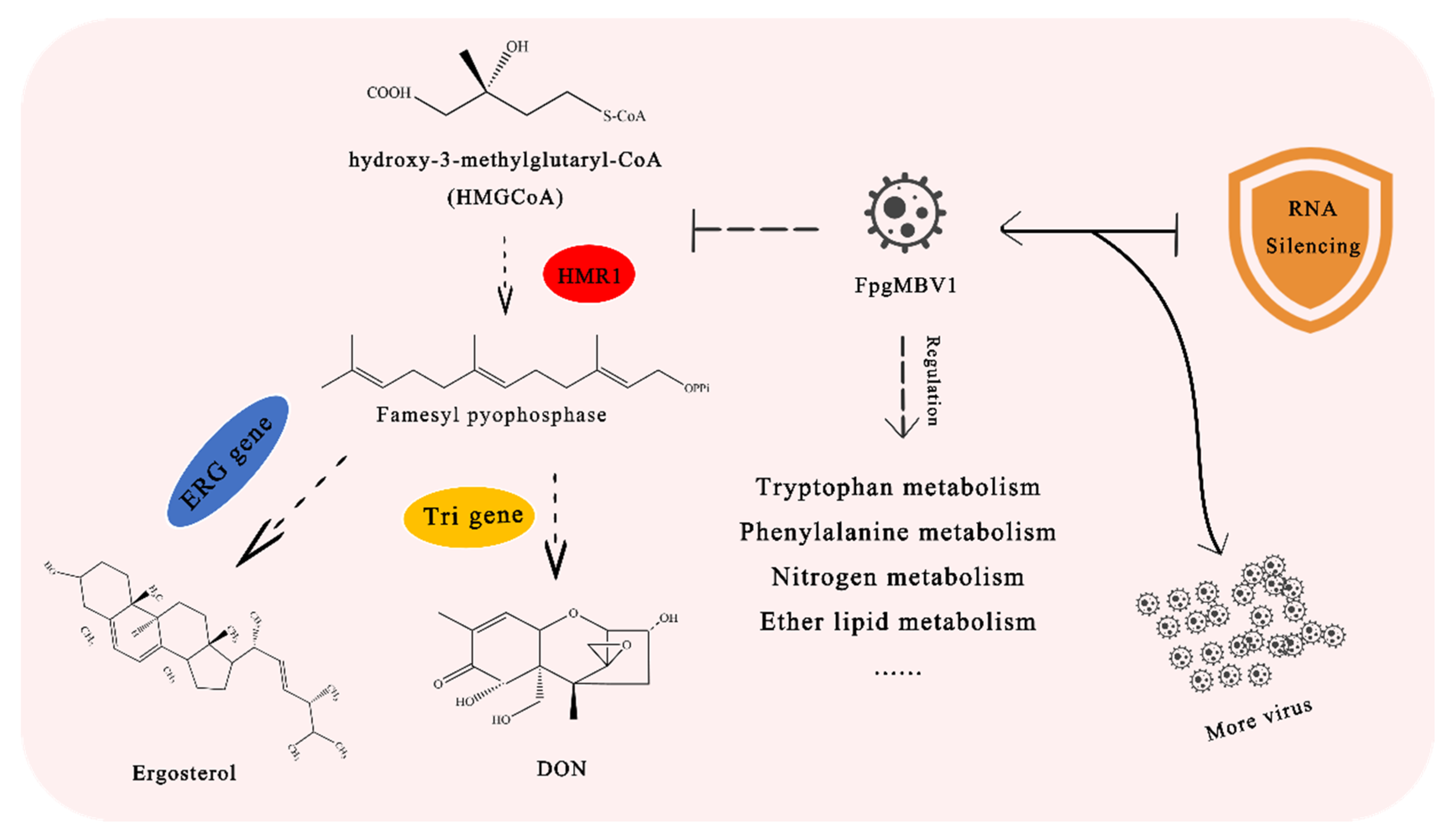
:1. Introduction
2. Results
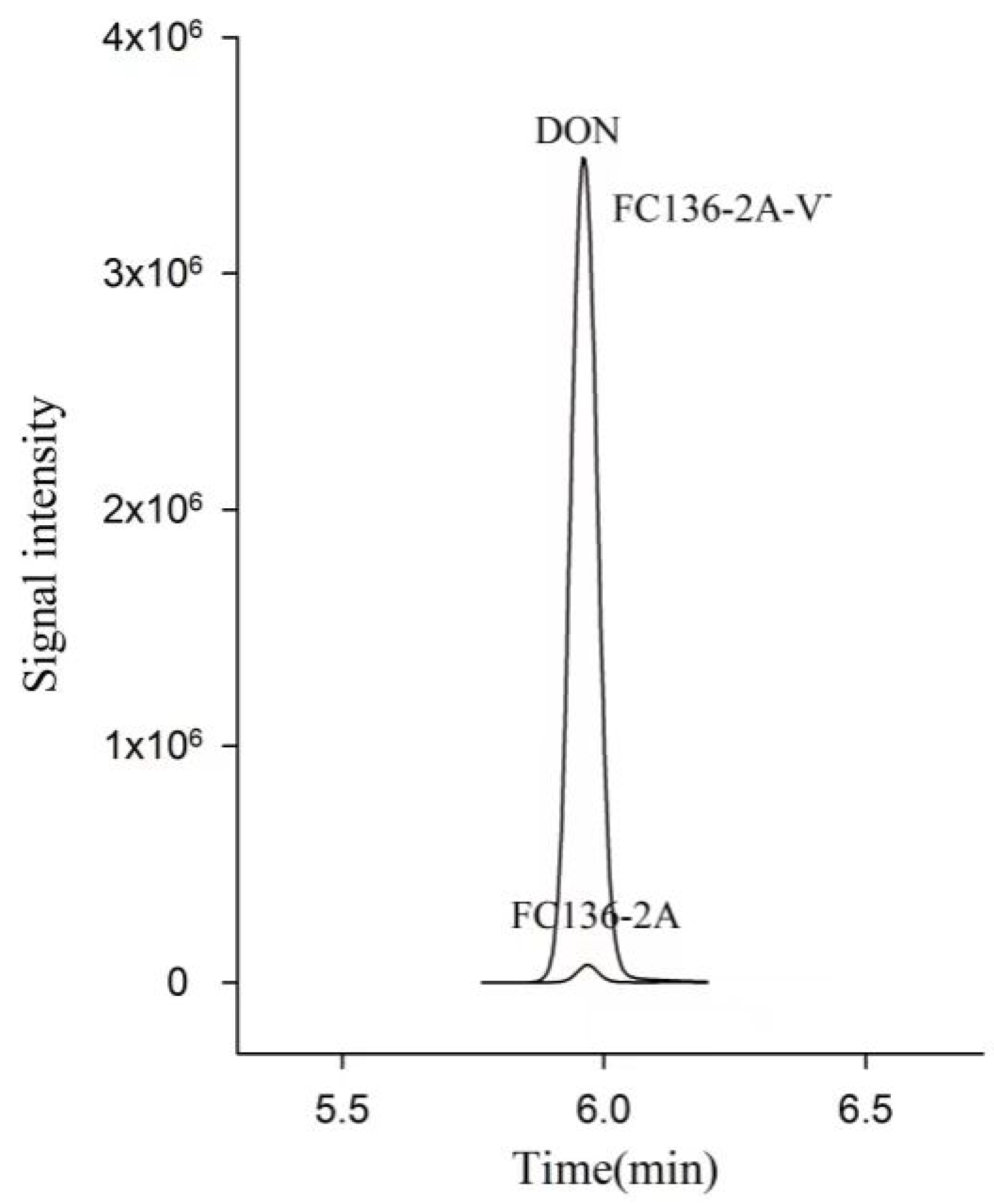
2.1. DON Synthesis Was Inhibited by Mycovirus FpgMBV1
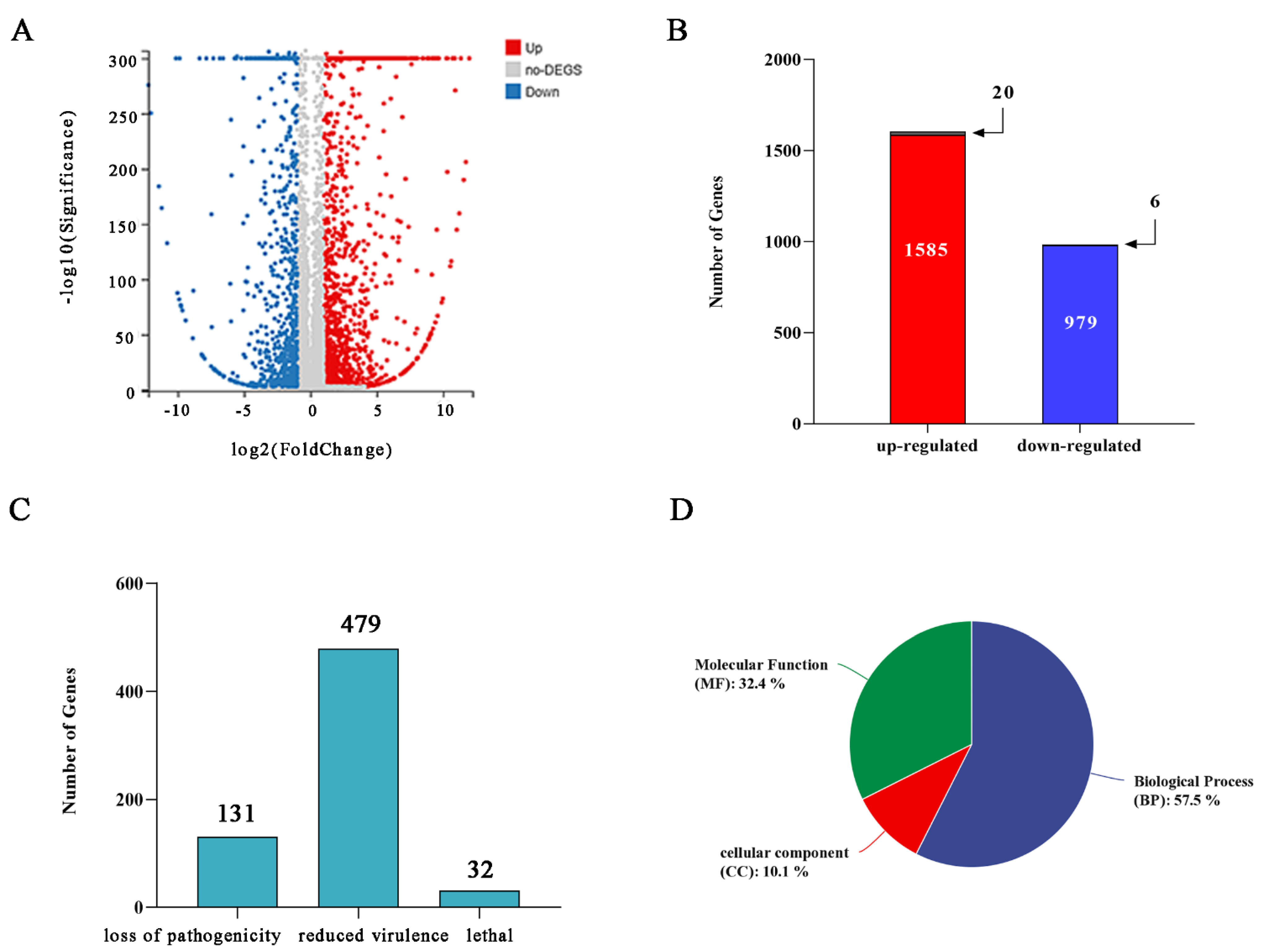
2.2. Overview of RNA-Seq Data for FC136-2A and FC136-2A-V-
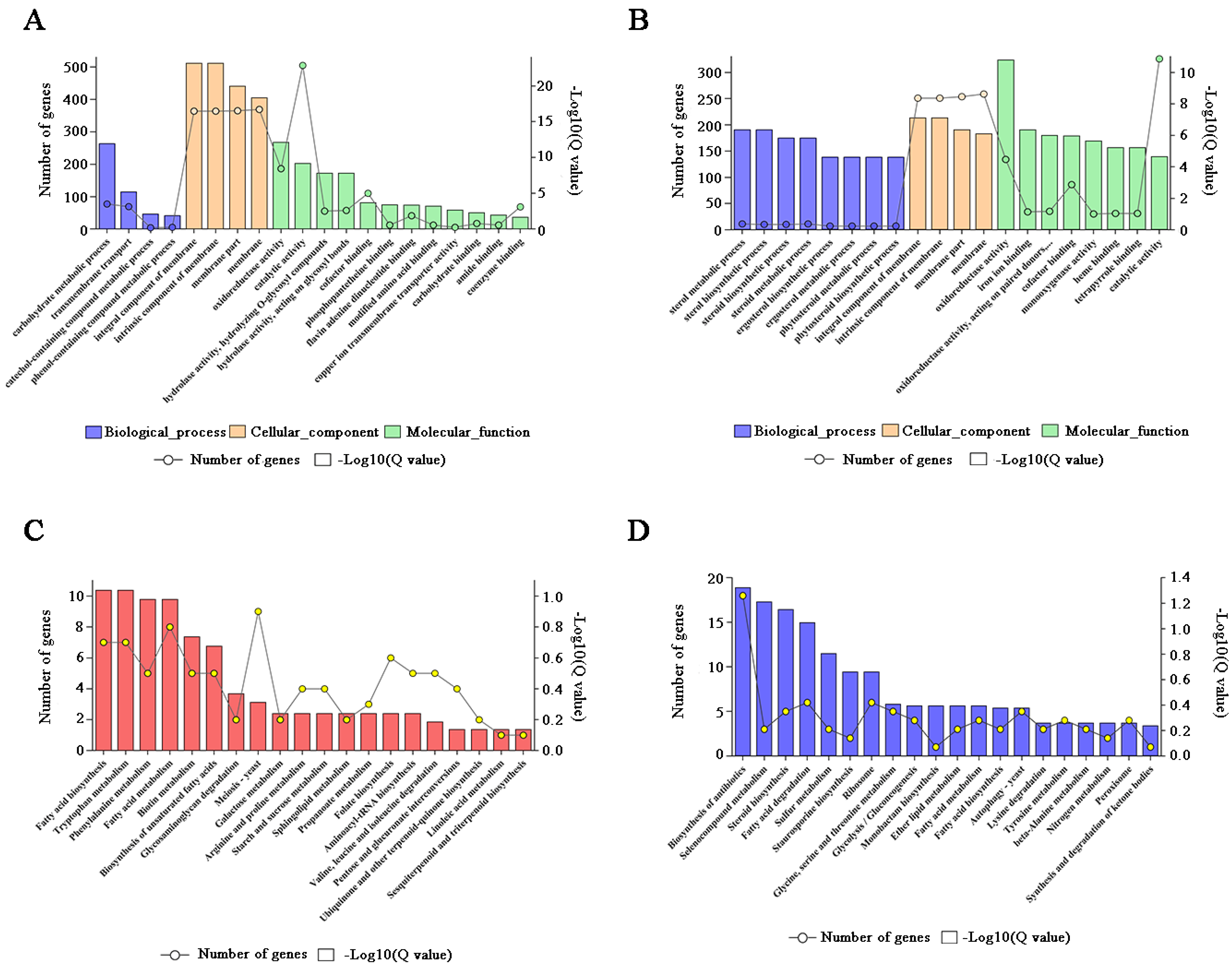
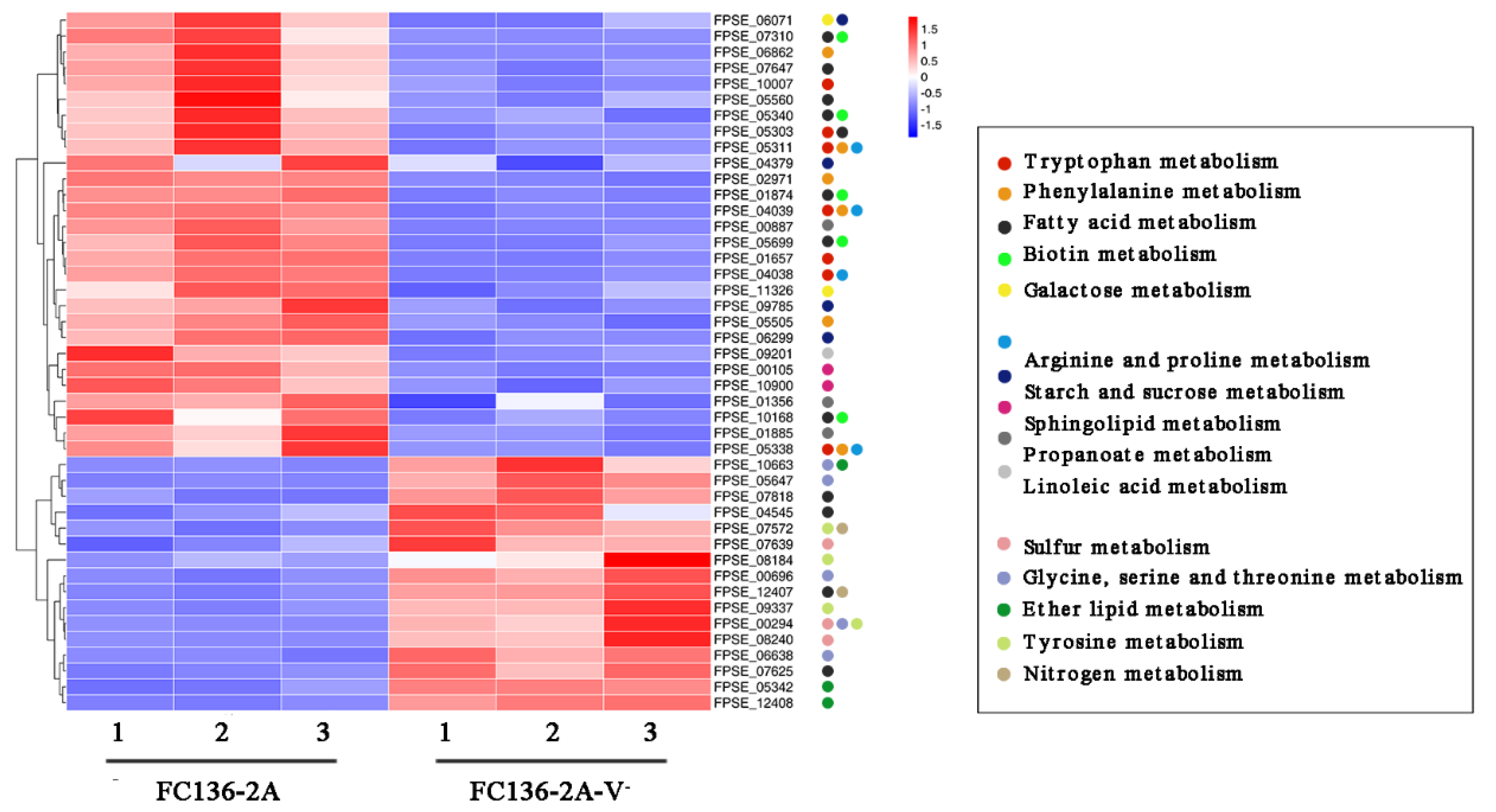
2.3. The Metabolic Balance in F. pseudograminearum Was Disturbed by FpgMBV1
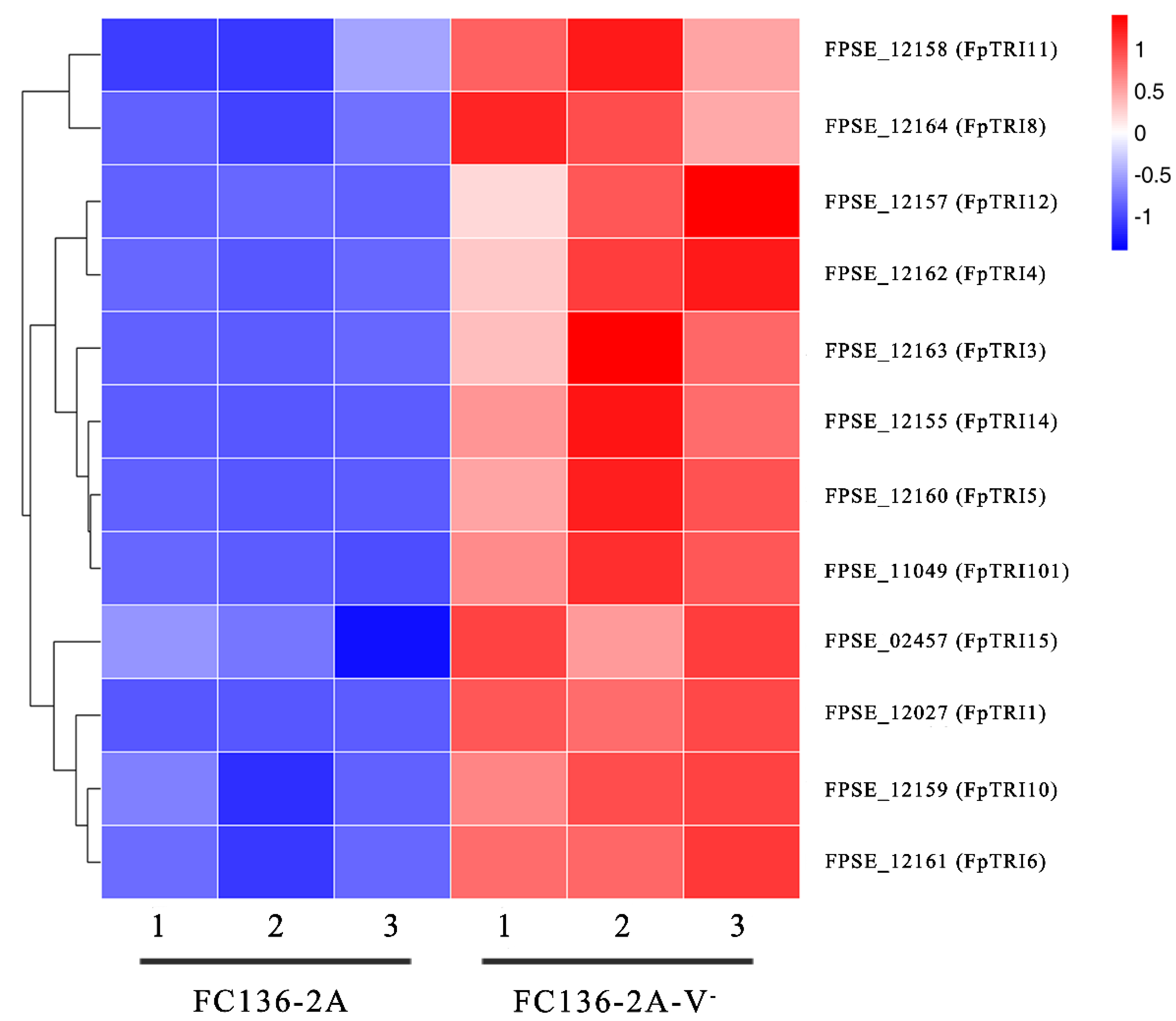
2.4. TRI Genes Regulating DON Biosynthesis Were Down-Regulated by FpgMBV1
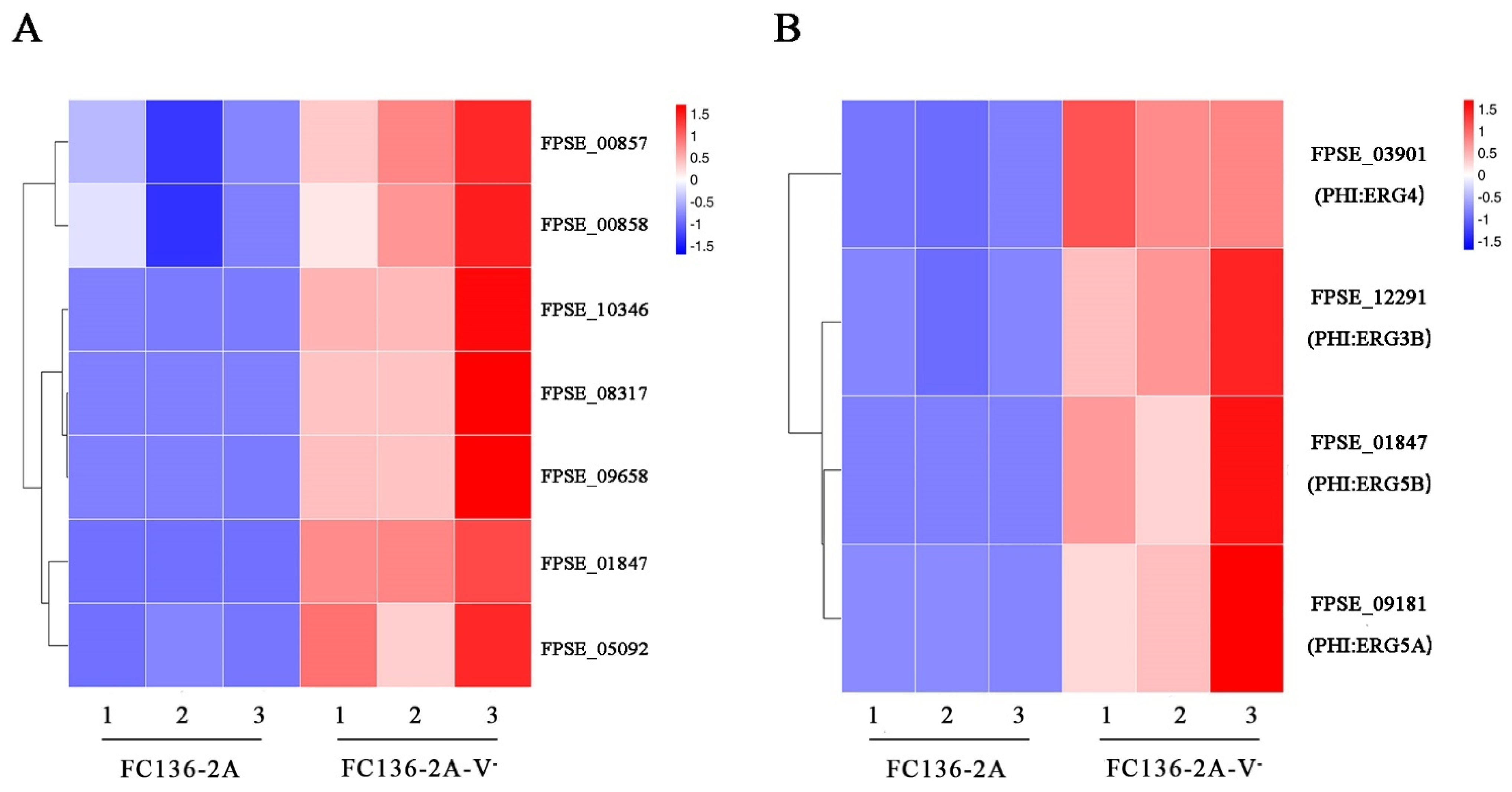
2.5. Ergosterol Biosynthesis and Metabolism Were Inhibited by FpgMBV1
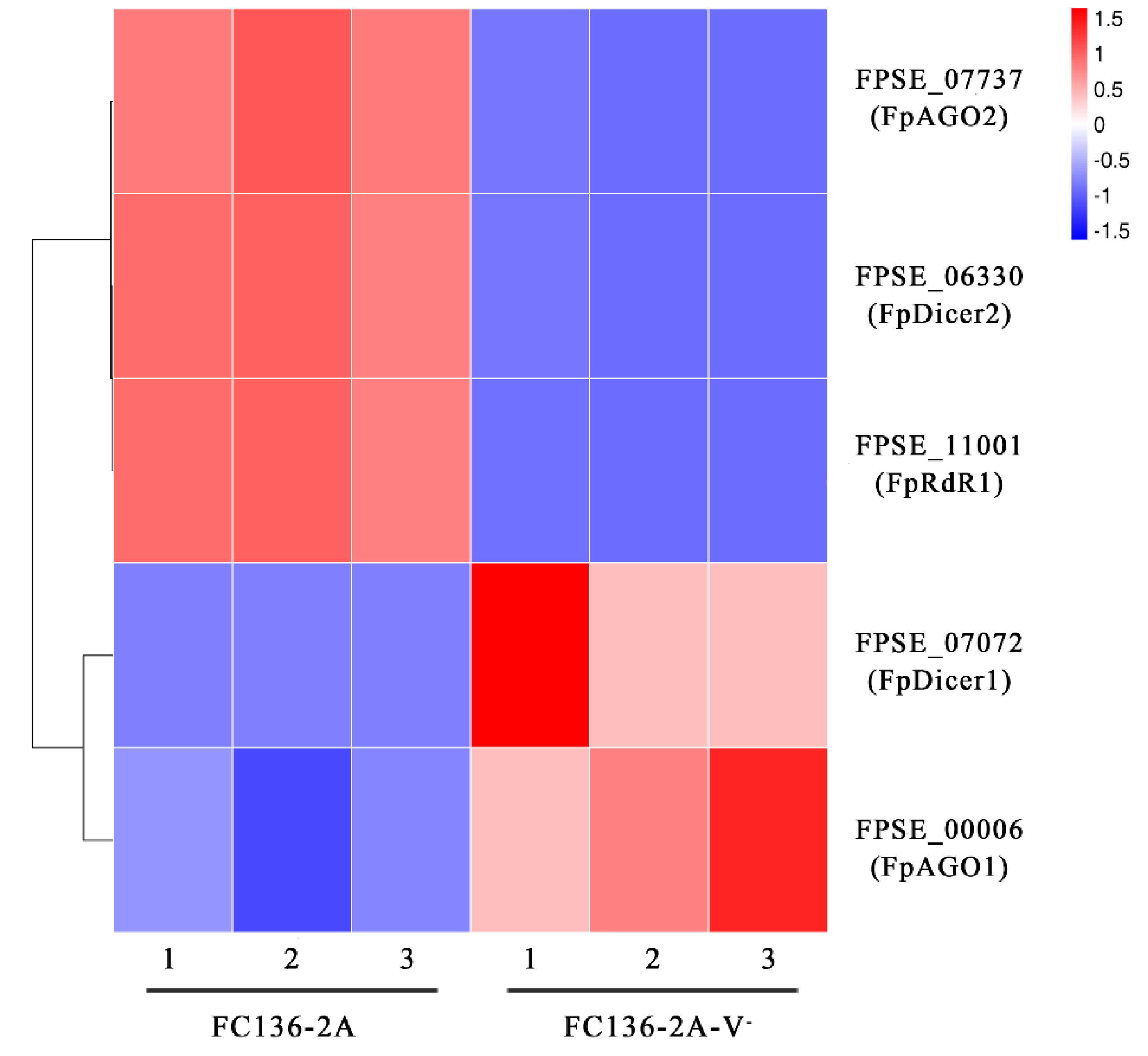
2.6. FpDicer1 and FpAGO1 Were Down-Regulated by FpgMBV1
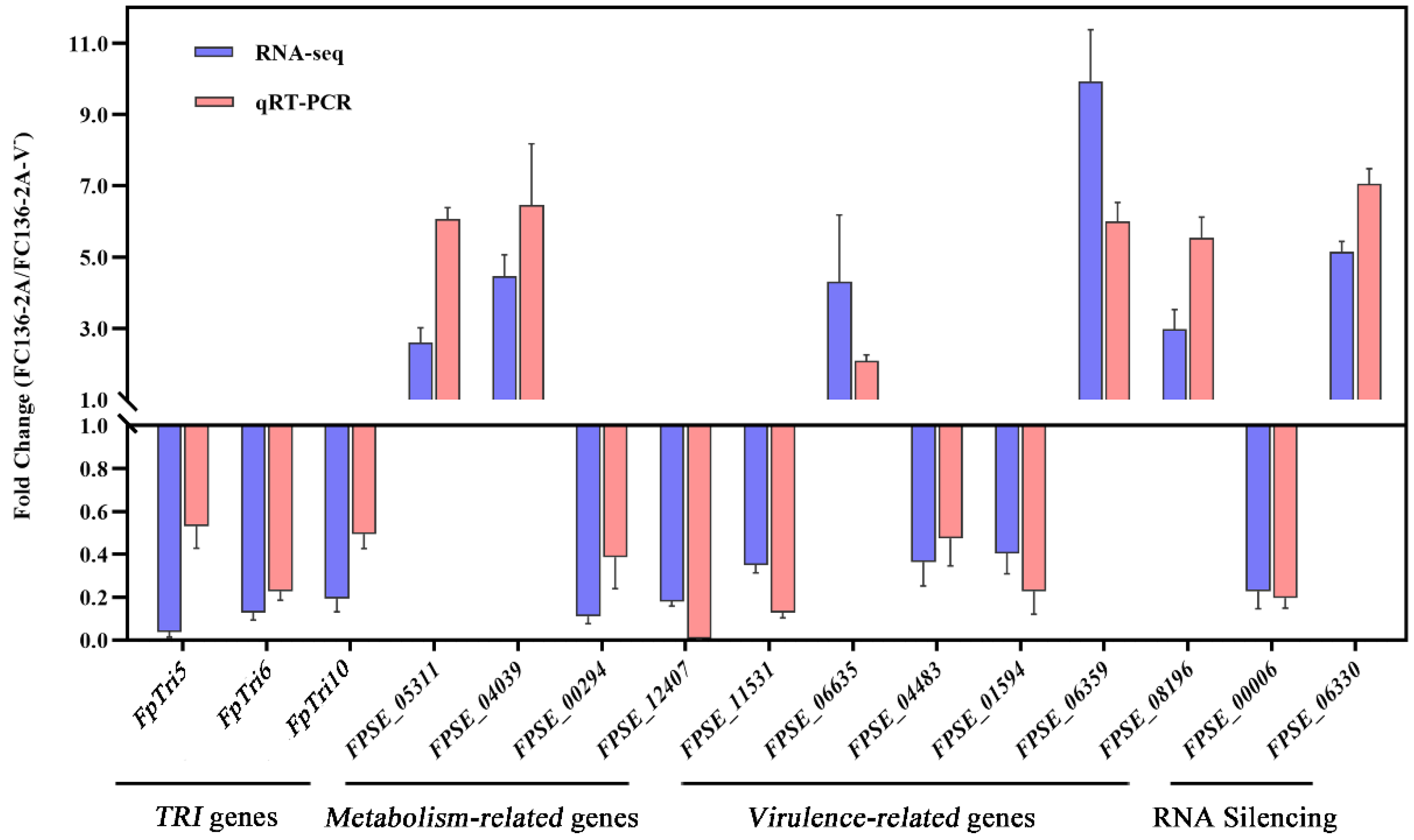
2.7. Gene Expression Level by Quantitive Real-Time RT-PCR
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Fungal Material and Growth Conditions
5.2. HPLC-MS/MS Analysis of Type B Trichothecene
5.3. Total RNA Extraction
5.4. cDNA Library Preparation and Sequencing
5.5. RNA-Seq Data Analysis
5.6. Quantitative Real-Time RT-PCR (qRT-PCR) Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burgess, L.; Bryden, W. Fusarium: A ubiquitous fungus of global significance. Microbiology 2012, 33, 22. [Google Scholar] [CrossRef]
- Ma, L.J.; Geiser, D.M.; Proctor, R.H.; Rooney, A.P.; O’Donnell, K.; Trail, F.; Gardiner, D.M.; Manners, J.M.; Kazan, K. Fusarium pathogenomics. Annu. Rev. Microbiol. 2013, 67, 399–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 314–430. [Google Scholar] [CrossRef] [Green Version]
- Rampersad, S. Pathogenomics and management of Fusarium diseases in plants. Pathogens 2020, 9, 340. [Google Scholar] [CrossRef] [PubMed]
- Petronaitis, T.; Simpfendorfer, S.; Hüberli, D. Importance of Fusarium spp. in wheat to food security: A global perspective. In Plant Pathology in the 21st Century; Springer: Berlin/Hamburg, Germany, 2021; Volume 10. [Google Scholar] [CrossRef]
- Li, H.L.; Yuan, H.X.; Fu, B.; Xing, X.P.; Sun, B.J.; Tang, W.H. First Report of Fusarium pseudograminearum Causing Crown Rot of Wheat in Henan, China. Plant Dis. 2012, 103, 18–19. [Google Scholar] [CrossRef]
- Alahmad, S.; Simpfendorfer, S.; Bentley, A.R.; Hickey, L.T. Crown rot of wheat in Australia: Fusarium pseudograminearum taxonomy, population biology and disease management. Australas. Plant Pathol. 2018, 47, 285–299. [Google Scholar] [CrossRef]
- Kazan, K.; Gardiner, D.M. Fusarium crown rot caused by Fusarium pseudograminearum in cereal crops: Recent progress and future prospects. Mol. Plant Pathol. 2018, 19, 1547–1562. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; He, X.; Wang, S.; Ma, Q.; Sun, B.; Ding, S.; Chen, L.; Zhang, M.; Li, H. Diversity of the Fusarium pathogens associated with crown rot in the Huanghuai wheat-growing region of China. Environ. Microbiol. 2019, 21, 2740–2754. [Google Scholar] [CrossRef]
- Bragard, C.; Baptista, P.; Chatzivassiliou, E.; DiSerio, F.; Gonthier, P.; JaquesMiret, J.A.; Justesen, A.F.; MacLeod, A.; Magnusson, C.S.; Milonas, P.; et al. Pest categorisation of Fusarium pseudograminearum. EFSA J. 2022, 29, e07399. [Google Scholar] [CrossRef]
- Perincherry, L.; Justyna, L.; Lukasz, S. Fusarium-Produced Mycotoxins in Plant-Pathogen Interactions. Toxins 2019, 11, 664. [Google Scholar] [CrossRef] [Green Version]
- Borbély, M.; Sipos, P.; Pelles, F.; Győri, Z. Mycotoxin contamination in cereals. J. Agroaliment. Process. Technol. 2010, 16, 96–98. Available online: https://www.researchgate.net/publication/265178683 (accessed on 10 June 2010).
- Lozowicka, B.; Iwaniuk, P.; Konecki, R.; Kaczynski, P.; Kuldybayev, N.; Dutbayev, Y. Impact of Diversified Chemical and Biostimulator Protection on Yield, Health Status, Mycotoxin Level, and Economic Profitability in Spring Wheat (Triticum aestivum L.) Cultivation. Agronomy 2022, 12, 258. [Google Scholar] [CrossRef]
- Skrzydlewski, P.; Twarużek, M.; Grajewski, J. Cytotoxicity of Mycotoxins and Their Combinations on Different Cell Lines: A Review. Toxins 2022, 14, 244. [Google Scholar] [CrossRef]
- Pierzgalski, A.; Bryła, M.; Kanabus, J.; Modrzewska, M.; Podolska, G. Updated Review of the Toxicity of Selected Fusarium Toxins and Their Modified Forms. Toxins 2021, 13, 768. [Google Scholar] [CrossRef]
- Flynn, C.M.; Broz, K.; Jonkers, W.; Schmidt-Dannert, C.; Kistler, H.C. Expression of the Fusarium graminearum terpenome and involvement of the endoplasmic reticulum-derived toxisome. Fungal Genet. Biol. 2019, 124, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Seong, K.Y.; Pasquali, M.; Zhou, X.; Song, J.; Hilburn, K.; McCormick, S.; Dong, Y.; Xu, J.R.; Kistler, H.C. Global gene regulation by Fusarium transcription factors Tri6 and Tri10 reveals adaptations for toxin biosynthesis. Mol. Microbiol. 2009, 72, 354–367. [Google Scholar] [CrossRef]
- Menke, J.; Dong, Y.N.; Kistler, H.C. Fusarium graminearum Tri12p influences virulence to wheat and trichothecene accumulation. Mol. Plant-Microbe Interact. 2012, 25, 1408–1418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, N.J.; McCormick, S.P.; Hohn, T.M. TRI12, a trichothecene efflux pump from Fusarium sporotrichioides: Gene isolation and expression in yeast. Mol. Gen. Genet. 1999, 261, 977–984. [Google Scholar] [CrossRef]
- Kimura, M.; Tokai, T.; O’Donnell, K.; Ward, T.J.; Fujimura, M.; Hamamoto, H.; Shibata, T.; Yamaguchi, I. The trichothecene biosynthesis gene cluster of Fusarium graminearum F15 contains a limited number of essential pathway genes and expressed non-essential genes. FEBS Lett. 2003, 539, 105–110. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.; Lee, S.H.; Shin, J.Y.; Kim, H.K.; Yun, S.H.; Kim, H.Y.; Lee, S.; Ryu, J.G. Comparison of trichothecene biosynthetic gene expression between Fusarium graminearum and Fusarium asiaticum. Plant Pathol. J. 2014, 30, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Lan, X.; Liao, H.; Parsley, T.B.; Nuss, D.L.; Chen, B. Genome Sequence, Full-Length Infectious cDNA Clone, and Mapping of Viral Double-Stranded RNA Accumulation Determinant of Hypovirus CHV1-EP721. J. Virol. 2007, 81, 1813–1820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Li, B.; Fu, Y.; Jiang, D.; Ghabrial, S.A.; Li, G.; Peng, Y.; Xie, J.; Cheng, J.; Huang, J.; et al. A geminivirus-related DNA mycovirus that confers hypovirulence to a plant pathogenic fungus. Proc. Natl. Acad. Sci. USA 2010, 107, 8387–8392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiba, S.; Salaipeth, L.; Lin, Y.-H.; Sasaki, A.; Kanematsu, S.; Suzuki, N. A Novel Bipartite Double-Stranded RNA Mycovirus from the White Root Rot Fungus Rosellinia necatrix: Molecular and Biological Characterization, Taxonomic Considerations, and Potential for Biological Control. J. Virol. 2009, 83, 12801–12812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Bhattacharjee, P.; Wang, S.; Zhang, L.; Ahmed, I.; Guo, L. Mycoviruses in Fusarium species: An update. Front. Cell. Infect. Microbiol. 2019, 9, 257. [Google Scholar] [CrossRef] [Green Version]
- Chu, Y.M.; Jeon, J.J.; Yea, S.J.; Kim, Y.H.; Yun, S.H.; Lee, Y.W.; Kim, K.H. Double-stranded RNA mycovirus from Fusarium graminearum. Appl. Environ. Microbiol. 2002, 68, 2529–2534. [Google Scholar] [CrossRef] [Green Version]
- Darissa, O.; Adam, G.; Schäfer, W. A dsRNA mycovirus causes hypovirulence of Fusarium graminearum to wheat and maize. Eur. J. Plant Pathol. 2012, 134, 181–189. [Google Scholar] [CrossRef]
- Li, P.; Zhang, H.; Chen, X.; Qiu, D.; Guo, L. Molecular characterization of a novel hypovirus from the plant pathogenic fungus Fusarium graminearum. Virology 2015, 481, 151–160. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Gao, F.; Zhang, F.; Xie, Y.; Zhou, L.; Yuan, H.; Zhang, S.; Li, H. The complete genomic sequence of a novel megabirnavirus from Fusarium pseudograminearum, the causal agent of wheat crown rot. Arch. Virol. 2018, 163, 3173–3175. [Google Scholar] [CrossRef]
- Xie, Y.; Wang, Z.; Li, K.; Liu, D.; Jia, Y.; Gao, F.; Dai, J.; Zhang, S.; Zhang, X.; Li, H. A megabirnavirus alleviates the pathogenicity of Fusarium pseudograminearum to wheat. Phytopathology 2021, 112, 1175–1184. [Google Scholar] [CrossRef]
- Li, P.; Lin, Y.; Zhang, H.; Wang, S.; Qiu, D.; Guo, L. Molecular characterization of a novel mycovirus of the family Tymoviridae isolated from the plant pathogenic fungus Fusarium graminearum. Virology 2016, 489, 86–94. [Google Scholar] [CrossRef] [Green Version]
- Cho, W.K.; Yu, J.; Lee, K.M.; Son, M.; Min, K.; Lee, Y.W.; Kim, K.H. Genome-wide expression profiling shows transcriptional reprogramming in Fusarium graminearum by Fusarium graminearum virus 1-DK21 infection. BMC Genom. 2012, 13, 173. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.M.; Cho, W.K.; Yu, J.; Son, M.; Choi, H.; Min, K.; Lee, Y.W.; Kim, K.H. A comparison of transcriptional patterns and mycological phenotypes following infection of Fusarium graminearum by four mycoviruses. PLoS ONE 2014, 9, e0100989. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, J.; Li, P.; Qiu, D.; Guo, L. Transcriptome-based discovery of Fusarium graminearum stress responses to FgHV1 infection. Int. J. Mol. Sci. 2016, 17, 1922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, Z.; Fu, Y.; Lin, Y.; Zhao, Z.; Zhang, X.; Cheng, J.; Xie, J.; Chen, T.; Li, B.; Jiang, D. Transcriptional Responses of Sclerotinia sclerotiorum to the Infection by SsHADV-1. J. Fungi 2021, 7, 493. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Kistler, H.C.; Ma, Z. Fusarium graminearum Trichothecene Mycotoxins: Biosynthesis, Regulation, and Management. Annu. Rev. Phytopathol. 2019, 57, 15–39. [Google Scholar] [CrossRef] [Green Version]
- Ding, S.W.; Li, H.; Lu, R.; Li, F.; Li, W.X. RNA silencing: A conserved antiviral immunity of plants and animals. Virus Res. 2004, 102, 109–115. [Google Scholar] [CrossRef]
- Nasmith, C.G.; Walkowiak, S.; Wang, L.; Leung, W.W.Y.; Gong, Y.; Johnston, A.; Harris, L.J.; Guttman, D.S.; Subramaniam, R. Tri6 is a global transcription regulator in the phytopathogen Fusarium graminearum. PLoS Pathog. 2011, 7, e1002266. [Google Scholar] [CrossRef] [PubMed]
- Proctor, R.H.; Hohn, T.M.; McCormick, S.P.; Desjardins, A.E. Tri6 encodes an unusual zinc finger protein involved in regulation of trichothecene biosynthesis in Fusarium sporotrichioides. Appl. Environ. Microbiol. 1995, 61, 1923–1930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peplow, A.W.; Tag, A.G.; Garifullina, G.F.; Beremand, M.N. Identification of new genes positively regulated by Tri10 and a regulatory network for trichothecene mycotoxin production. Appl. Environ. Microbiol. 2003, 69, 2731–2736. [Google Scholar] [CrossRef] [Green Version]
- Yun, Y.; Yin, D.; Dawood, D.H.; Liu, X.; Chen, Y.; Ma, Z. Functional characterization of FgERG3 and FgERG5 associated with ergosterol biosynthesis, vegetative differentiation and virulence of Fusarium graminearum. Fungal Genet. Biol. 2014, 68, 60–70. [Google Scholar] [CrossRef]
- Liu, X.; Jiang, J.; Yin, Y.; Ma, Z. Involvement of FgERG4 in ergosterol biosynthesis, vegetative differentiation and virulence in Fusarium graminearum. Mol. Plant Pathol. 2013, 14, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J.; Abramson, D.; Mccallum, B.; Clear, R. Comparison of Canadian Fusarium graminearum isolates for aggressiveness, vegetative compatibility, and production of ergosterol and mycotoxins. Mycopathologia 2002, 153, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Lamper, C.; Teren, J.; Bartok, T.; Komoroczy, R.; Mesterhazy, A.; Sagi, F. Predicting DON contamination in Fusarium-infected wheat grains via determination of the ergosterol content. Cereal Res. Commun. 2000, 28, 337–344. [Google Scholar] [CrossRef]
- Rodrigues, M.L. The multifunctional fungal ergosterol. mBio 2018, 9, e01755-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacob-Wilk, D.; Turina, M.; Van Alfen, N.K. Mycovirus Cryphonectria hypovirus 1 elements cofractionate with trans-Golgi network membranes of the fungal host Cryphonectria parasitica. J. Virol. 2006, 80, 6588–6596. [Google Scholar] [CrossRef] [Green Version]
- Van De Sande, W.W.J.; Lo-Ten-Foe, J.R.; Van Belkum, A.; Netea, M.G.; Kullberg, B.J.; Vonk, A.G. Mycoviruses: Future therapeutic agents of invasive fungal infections in humans? Eur. J. Clin. Microbiol. Infect. Dis. 2010, 29, 755–763. [Google Scholar] [CrossRef] [Green Version]
- Seong, K.; Li, L.; Hou, Z.; Tracy, M.; Kistler, H.C.; Xu, J.R. Cryptic promoter activity in the coding region of the HMG-CoA reductase gene in Fusarium graminearum. Fungal Genet. Biol. 2006, 43, 34–41. [Google Scholar] [CrossRef]
- Stermer, B.A.; Bianchini, G.M.; Korth, K.L. Regulation of HMG-CoA reductase activity in plants. J. Lipid Res. 1994, 35, 1133–1140. [Google Scholar] [CrossRef]
- Lee, J.; Myong, K.; Kim, J.E.; Kim, H.K.; Yun, S.H.; Lee, Y.W. FgVelB globally regulates sexual reproduction, mycotoxin production and pathogenicity in the cereal pathogen Fusarium graminearum. Microbiology 2012, 158, 1723–1733. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Yun, Y.; Liu, Y.; Ma, Z. FgVeLB is associated with vegetative differentiation, secondary metabolism and virulence in Fusarium graminearum. Fungal Genet. Biol. 2012, 49, 653–662. [Google Scholar] [CrossRef]
- Chun, J.; Ko, Y.H.; Kim, D.H. Transcriptome analysis of Cryphonectria parasitica infected with Cryphonectria hypovirus 1 (CHV1) reveals distinct genes related to fungal metabolites, virulence, antiviral RNA-silencing, and their regulation. Front. Microbiol. 2020, 11, 1711. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, R.; Narayanan, S.; Walkowiak, S.; Wang, L.; Joshi, M.; Rocheleau, H.; Ouellet, T.; Harris, L.J. Leucine metabolism regulates TRI6 expression and affects deoxynivalenol production and virulence in Fusarium graminearum. Mol. Microbiol. 2015, 98, 760–769. [Google Scholar] [CrossRef] [PubMed]
- Leonetti, P.; Stuttmann, J.; Pantaleo, V. Regulation of plant antiviral defense genes via host RNA-silencing mechanisms. Virol. J. 2021, 18, 194. [Google Scholar] [CrossRef] [PubMed]
- Nuss, D.L. Mycoviruses, RNA Silencing, and Viral RNA Recombination. Adv. Virus Res. 2011, 80, 25–48. [Google Scholar] [CrossRef] [Green Version]
- Mochama, P.; Jadhav, P.; Neupane, A.; Marzano, S.Y.L. Mycoviruses as triggers and targets of RNA silencing in white mold fungus Sclerotinia sclerotiorum. Viruses 2018, 10, 214. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Park, J.Y.; Heo, J.I.; Kim, K.H. The ORF2 protein of Fusarium graminearum virus 1 suppresses the transcription of FgDICER2 and FgAGO1 to limit host antiviral defences. Mol. Plant Pathol. 2020, 21, 230–243. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | SwissProt_Description | log2(FC136_2A/FC136_2A_V-) | Identity (%) | E-Value | PHI Accession |
|---|---|---|---|---|---|---|
| ERG3B | FPSE_12291 | Probable Delta(7)-sterol 5(6)-desaturase | −1.656 | 92.3 | 7.3 × 10−179 | PHI:3036 |
| ERG4 | FPSE_03901 | Delta(24(24(1)))-sterol reductase | −1.336 | 96.6 | 0 | PHI:2728 |
| ERG5A | FPSE_09181 | Cytochrome P450 61 | −1.584 | 99.3 | 0 | PHI:3037 |
| ERG5B | FPSE_01847 | Sterol 22-desaturase | −7.320 | 94.3 | 1.6 × 10−292 | PHI:3038 |
| TRI5 | FPSE_12160 | Trichodiene synthase | −3.030 | 100 | 1.1 × 10−228 | PHI:6846 |
| TRI6 | FPSE_12161 | Trichothecene biosynthesis transcription regulator 6 | −2.686 | 95.9 | 1.2 × 10−126 | PHI:1362 |
| TRI12 | FPSE_12157 | Trichothecene efflux pump TRI12 | −2.305 | 91.7 | 4.5 × 10−307 | PHI:2704 |
| TRI10 | FPSE_12159 | Trichothecene biosynthesis transcription regulator 10 | −2.702 | 93.3 | 1.5 × 10−232 | PHI:2328 |
| TRI15 | FPSE_02457 | Cys(2)-His(2) zinc finger protein | −1.803 | 95.7 | 8.3 × 10−183 | PHI:1363 |
| HMR1 | FPSE_03466 | Hydroxymethylglutaryl CoA reductase gene | −1.781 | 94.7 | 0 | PHI:1006 |
| VELB | FPSE_11531 | Velvet complex subunit B | −1.760 | 83.3 | 1.1 × 10−159 | PHI:2427 |
| GLX | FPSE_04483 | WSC domain-containing protein ARB_07867 | −1.238 | 87 | 0 | PHI:5393 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, K.; Liu, D.; Pan, X.; Yan, S.; Song, J.; Liu, D.; Wang, Z.; Xie, Y.; Dai, J.; Liu, J.; et al. Deoxynivalenol Biosynthesis in Fusarium pseudograminearum Significantly Repressed by a Megabirnavirus. Toxins 2022, 14, 503. https://doi.org/10.3390/toxins14070503
Li K, Liu D, Pan X, Yan S, Song J, Liu D, Wang Z, Xie Y, Dai J, Liu J, et al. Deoxynivalenol Biosynthesis in Fusarium pseudograminearum Significantly Repressed by a Megabirnavirus. Toxins. 2022; 14(7):503. https://doi.org/10.3390/toxins14070503
Chicago/Turabian StyleLi, Ke, Dongmei Liu, Xin Pan, Shuwei Yan, Jiaqing Song, Dongwei Liu, Zhifang Wang, Yuan Xie, Junli Dai, Jihong Liu, and et al. 2022. "Deoxynivalenol Biosynthesis in Fusarium pseudograminearum Significantly Repressed by a Megabirnavirus" Toxins 14, no. 7: 503. https://doi.org/10.3390/toxins14070503
APA StyleLi, K., Liu, D., Pan, X., Yan, S., Song, J., Liu, D., Wang, Z., Xie, Y., Dai, J., Liu, J., Li, H., Zhang, X., & Gao, F. (2022). Deoxynivalenol Biosynthesis in Fusarium pseudograminearum Significantly Repressed by a Megabirnavirus. Toxins, 14(7), 503. https://doi.org/10.3390/toxins14070503