Comparative Analysis of Transcriptomic Changes including mRNA and microRNA Expression Induced by the Xenoestrogens Zearalenone and Bisphenol A in Human Ovarian Cells

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Results
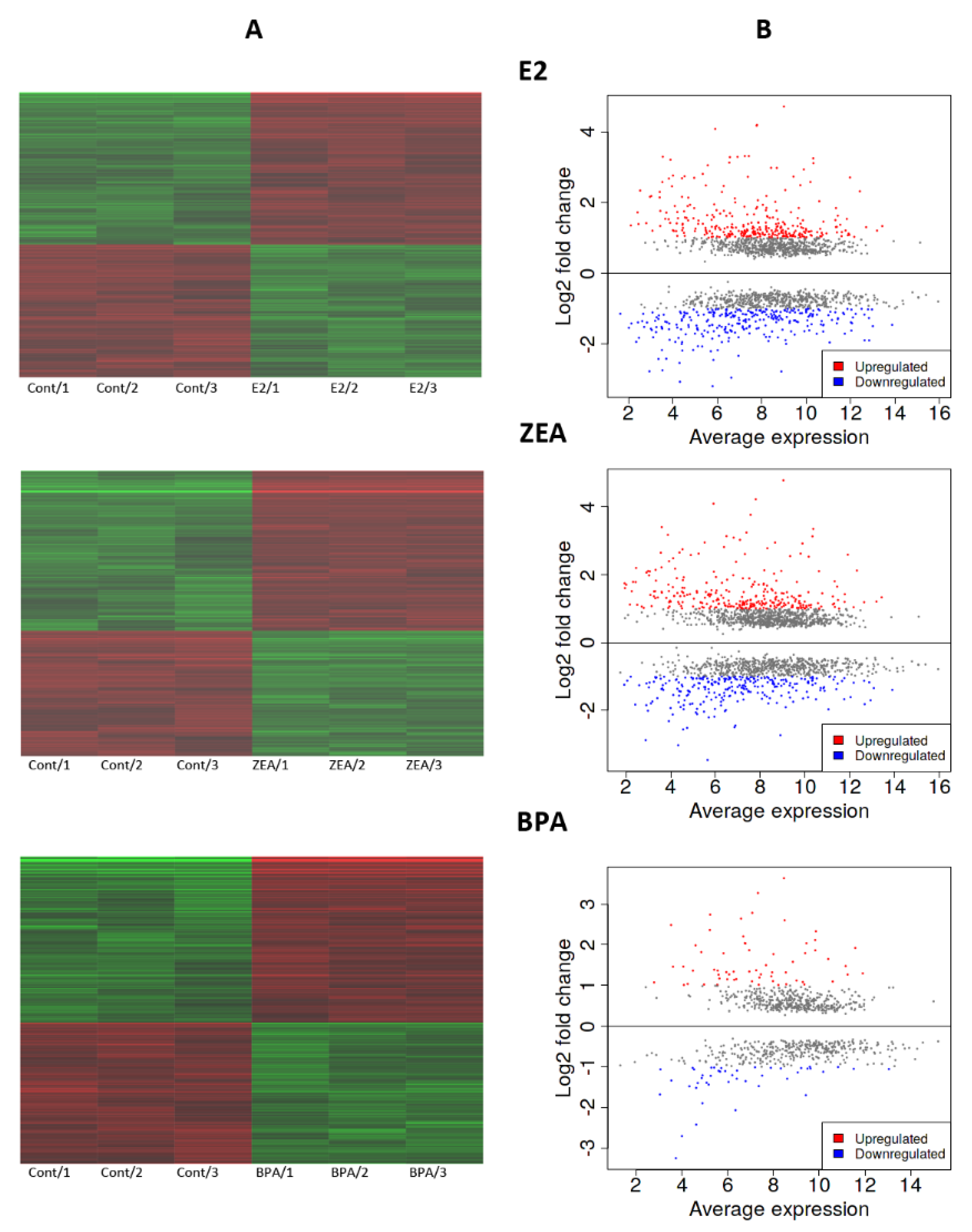
2.1. ZEA-Induced Changes in mRNA Expression Are More Comparable to That of E2 Than BPA
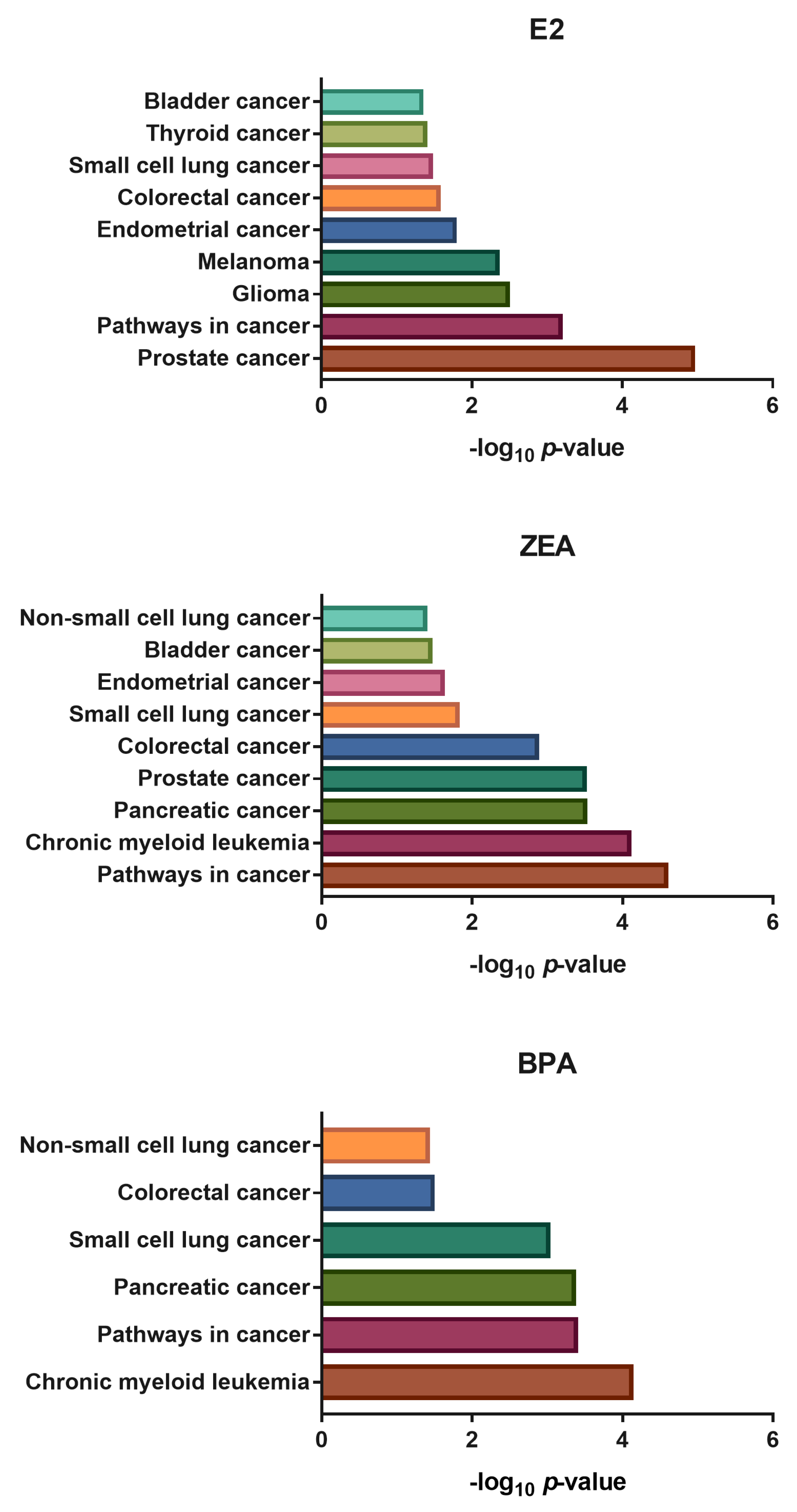
2.2. Transcriptomic Changes Induced by E2, ZEA and BPA Favored Cell Proliferation and Migration
2.3. E2, ZEA and BPA Altered the Expression of miRNAs
2.4. Alterations in miRNA Expression Induced by E2, ZEA and BPA Might Support Tumor Growth and Migration
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Cell Culturing
5.2. MRNA Isolation and Quantification by qPCR
5.3. MiRNA Isolation and Quantification by qPCR
5.4. Transcriptomic Analysis by RNA Sequencing
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fuentes, N.; Silveyra, P. Estrogen receptor signaling mechanisms. Adv. Protein. Chem. Struct. Biol. 2019, 116, 135–170. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.R.; Zhang, R.; Lian, Z.X.; Deng, S.L.; Yu, K. Estrogen-Receptor Expression and Function in Female Reproductive Disease. Cells 2019, 8, 1123. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, S.C.; Korach, K.S. Estrogen Receptors: New Directions in the New Millennium. Endocr. Rev. 2018, 39, 664–675. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed]
- Klinge, C.M. miRNAs and estrogen action. Trends Endocrinol. Metab. 2012, 23, 223–233. [Google Scholar] [CrossRef]
- Vrtačnik, P.; Ostanek, B.; Mencej-Bedrač, S.; Marc, J. The many faces of estrogen signaling. Biochem. Med. 2014, 24, 329–342. [Google Scholar] [CrossRef]
- Rahman, M.M.; Brane, A.C.; Tollefsbol, T.O. MicroRNAs and Epigenetics Strategies to Reverse Breast Cancer. Cells 2019, 8, 1214. [Google Scholar] [CrossRef]
- Howard, E.W.; Yang, X. microRNA Regulation in Estrogen Receptor-Positive Breast Cancer and Endocrine Therapy. Biol. Proced. Online 2018, 20, 17. [Google Scholar] [CrossRef]
- La Merrill, M.A.; Vandenberg, L.N.; Smith, M.T.; Goodson, W.; Browne, P.; Patisaul, H.B.; Guyton, K.Z.; Kortenkamp, A.; Cogliano, V.J.; Woodruff, T.J.; et al. Consensus on the key characteristics of endocrine-disrupting chemicals as a basis for hazard identification. Nat. Rev. Endocrinol. 2020, 16, 45–57. [Google Scholar] [CrossRef]
- Zheng, W.; Feng, N.; Wang, Y.; Noll, L.; Xu, S.; Liu, X.; Lu, N.; Zou, H.; Gu, J.; Yuan, Y.; et al. Effects of zearalenone and its derivatives on the synthesis and secretion of mammalian sex steroid hormones. Food Chem. Toxicol. 2019, 126, 262–276. [Google Scholar] [CrossRef]
- Pivonello, C.; Muscogiuri, G.; Nardone, A.; Garifalos, F.; Provvisiero, D.P.; Verde, N.; de Angelis, C.; Conforti, A.; Piscopo, M.; Auriemma, R.S.; et al. Bisphenol A: An emerging threat to female fertility. Reprod. Biol. Endocrinol. 2020, 18, 22. [Google Scholar] [CrossRef]
- Khan, N.G.; Correia, J.; Adiga, D.; Rai, P.S.; Dsouza, H.S.; Chakrabarty, S.; Kabekkodu, S.P. A comprehensive review on the carcinogenic potential of bisphenol A: Clues and evidence. Environ. Sci. Pollut. Res. Int. 2021, 28, 19643–19663. [Google Scholar] [CrossRef]
- Amir, S.; Shah, S.T.A.; Mamoulakis, C.; Docea, A.O.; Kalantzi, O.I.; Zachariou, A.; Calina, D.; Carvalho, F.; Sofikitis, N.; Makrigiannakis, A.; et al. Endocrine Disruptors Acting on Estrogen and Androgen Pathways Cause Reproductive Disorders through Multiple Mechanisms. Int. J. Environ. Res. Public Health 2021, 18, 1464. [Google Scholar] [CrossRef]
- Zheng, W.; Wang, B.; Li, X.; Wang, T.; Zou, H.; Gu, J.; Yuan, Y.; Liu, X.; Bai, J.; Bian, J.; et al. Zearalenone Promotes Cell Proliferation or Causes Cell Death? Toxins 2018, 10, 184. [Google Scholar] [CrossRef]
- Han, X.; Huangfu, B.; Xu, T.; Xu, W.; Asakiya, C.; Huang, K.; He, X. Research Progress of Safety of Zearalenone. Toxins 2022, 14, 386. [Google Scholar] [CrossRef]
- Gao, H.; Yang, B.J.; Li, N.; Feng, L.M.; Shi, X.Y.; Zhao, W.H.; Liu, S.J. Bisphenol A and hormone-associated cancers: Current progress and perspectives. Medicine 2015, 94, e211. [Google Scholar] [CrossRef]
- Mahato, D.K.; Devi, S.; Pandhi, S.; Sharma, B.; Maurya, K.K.; Mishra, S.; Dhawan, K.; Selvakumar, R.; Kamle, M.; Mishra, A.K.; et al. Occurrence, Impact on Agriculture, Human Health, and Management Strategies of Zearalenone in Food and Feed. Toxins 2021, 13, 92. [Google Scholar] [CrossRef]
- EFSA. Panel on Contaminants in the Food Chain. Scientific Opinion on the risks for public health related to the presence of zearalenone in food. EFSA J. 2011, 9, 2197. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Li, H.; Liu, Y.; Wang, Y.; Zhao, X.; Qi, X. Hormone therapy for ovarian cancer: Emphasis on mechanisms and applications. Oncol. Rep. 2021, 46, 223. [Google Scholar] [CrossRef]
- Mungenast, F.; Thalhammer, T. Estrogen biosynthesis and action in ovarian cancer. Front. Endocrinol. 2014, 5, 192. [Google Scholar] [CrossRef] [PubMed]
- Voutsadakis, I.A. Hormone Receptors in Serous Ovarian Carcinoma: Prognosis, Pathogenesis, and Treatment Considerations. Clin. Med. Insights Oncol. 2016, 10, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Márton, É.; Varga, A.; Széles, L.; Göczi, L.; Penyige, A.; Nagy, B.; Szilágyi, M. The Cell-Free Expression of MiR200 Family Members Correlates with Estrogen Sensitivity in Human Epithelial Ovarian Cells. Int. J. Mol. Sci. 2020, 21, 9725. [Google Scholar] [CrossRef] [PubMed]
- Márton, É.; Varga, A.; Soltész, B.; Penyige, A.; Lukács, J.; Póka, R.; Nagy, B.; Szilágyi, M. Comparative analysis of cell-free miR-205-5p, let-7f-5p, and miR-483-5p expression in ovarian cell cultures and plasma samples of patients with ovarian cancer. Appl. Sci. 2021, 11, 1735. [Google Scholar] [CrossRef]
- Varga, A.; Márton, É.; Markovics, A.; Penyige, A.; Balogh, I.; Nagy, B.; Szilágyi, M. Suppressing the PI3K/AKT Pathway by miR-30d-5p Mimic Sensitizes Ovarian Cancer Cells to Cell Death Induced by High-Dose Estrogen. Biomedicines 2022, 10, 2060. [Google Scholar] [CrossRef]
- Cheng, M.; Michalski, S.; Kommagani, R. Role for Growth Regulation by Estrogen in Breast Cancer 1 (GREB1) in Hormone-Dependent Cancers. Int. J. Mol. Sci. 2018, 19, 2543. [Google Scholar] [CrossRef]
- Hodgkinson, K.; Forrest, L.A.; Vuong, N.; Garson, K.; Djordjevic, B.; Vanderhyden, B.C. GREB1 is an estrogen receptor-regulated tumour promoter that is frequently expressed in ovarian cancer. Oncogene 2018, 37, 5873–5886. [Google Scholar] [CrossRef]
- Cuesta, R.; Gritsenko, M.A.; Petyuk, V.A.; Shukla, A.K.; Tsai, C.F.; Liu, T.; McDermott, J.E.; Holz, M.K. Phosphoproteome Analysis Reveals Estrogen-ER Pathway as a Modulator of mTOR Activity via DEPTOR. Mol. Cell. Proteom. 2019, 18, 1607–1618. [Google Scholar] [CrossRef]
- Li, Y.; Lei, B.; Zou, J.; Wang, W.; Chen, A.; Zhang, J.; Fu, Y.; Li, Z. High expression of carbonic anhydrase 12 (CA12) is associated with good prognosis in breast cancer. Neoplasma 2019, 66, 420–426. [Google Scholar] [CrossRef]
- Franke, C.M.; Gu, V.W.; Grimm, B.G.; Cassady, V.C.; White, J.R.; Weigel, R.J.; Kulak, M.V. TFAP2C regulates carbonic anhydrase XII in human breast cancer. Oncogene 2020, 39, 1290–1301. [Google Scholar] [CrossRef]
- Wang, Y.; Pringle, K.G.; Lumbers, E.R. The effects of cyclic AMP, sex steroids and global hypomethylation on the expression of genes controlling the activity of the renin-angiotensin system in placental cell lines. Placenta 2013, 34, 275–280. [Google Scholar] [CrossRef]
- de Aguiar Greca, S.C.; Kyrou, I.; Pink, R.; Randeva, H.; Grammatopoulos, D.; Silva, E.; Karteris, E. Involvement of the Endocrine-Disrupting Chemical Bisphenol A (BPA) in Human Placentation. J. Clin. Med. 2020, 9, 405. [Google Scholar] [CrossRef]
- Gao, J.; Song, T.; Che, D.; Li, C.; Jiang, J.; Pang, J.; Yang, Y.; Goma; Li, P. The effect of bisphenol a exposure onto endothelial and decidualized stromal cells on regulation of the invasion ability of trophoblastic spheroids in in vitro co-culture model. Biochem. Biophys. Res. Commun. 2019, 516, 506–514. [Google Scholar] [CrossRef]
- Piryaei, Z.; Salehi, Z.; Tahsili, M.R.; Ebrahimie, E.; Ebrahimi, M.; Kavousi, K. Agonist/antagonist compounds’ mechanism of action on estrogen receptor-positive breast cancer: A system-level investigation assisted by meta-analysis. Inform. Med. Unlocked 2022, 31, 100985. [Google Scholar] [CrossRef]
- Kimura, R.; Otani, T.; Shiraishi, N.; Hagiyama, M.; Yoneshige, A.; Wada, A.; Kajiyama, H.; Takeuchi, F.; Mizuguchi, N.; Morishita, K.; et al. Expression of cell adhesion molecule 1 in human and murine endometrial glandular cells and its increase during the proliferative phase by estrogen and cell density. Life Sci. 2021, 283, 119854. [Google Scholar] [CrossRef]
- Men, X.; Ma, J.; Wu, T.; Pu, J.; Wen, S.; Shen, J.; Wang, X.; Wang, Y.; Chen, C.; Dai, P. Transcriptome profiling identified differentially expressed genes and pathways associated with tamoxifen resistance in human breast cancer. Oncotarget 2017, 9, 4074–4089. [Google Scholar] [CrossRef]
- Wang, J.; Shidfar, A.; Ivancic, D.; Ranjan, M.; Liu, L.; Choi, M.R.; Parimi, V.; Gursel, D.B.; Sullivan, M.E.; Najor, M.S.; et al. Overexpression of lipid metabolism genes and PBX1 in the contralateral breasts of women with estrogen receptor-negative breast cancer. Int. J. Cancer 2017, 140, 2484–2497. [Google Scholar] [CrossRef]
- Du, F.; Zheng, F.; Han, Y.; Zhao, J.; Yuan, P. Novel Immune-Related Gene Signature for Risk Stratification and Prognosis of Survival in ER (+) and/or PR (+) and HER2 (−) Breast Cancer. Front. Pharmacol. 2022, 13, 820437. [Google Scholar] [CrossRef]
- Böckers, M.; Paul, N.W.; Efferth, T. Organophosphate ester tri-o-cresyl phosphate interacts with estrogen receptor α in MCF-7 breast cancer cells promoting cancer growth. Toxicol. Appl. Pharmacol. 2020, 395, 114977. [Google Scholar] [CrossRef]
- Böckers, M.; Paul, N.W.; Efferth, T. Bisphenolic compounds alter gene expression in MCF-7 cells through interaction with estrogen receptor α. Toxicol. Appl. Pharmacol. 2020, 399, 115030. [Google Scholar] [CrossRef]
- Chow, L.W.C.; Morita, S.; Chow, C.Y.C.; Ng, W.K.; Toi, M. Neoadjuvant palbociclib on ER+ breast cancer (N007): Clinical response and EndoPredict’s value. Endocr. Relat. Cancer 2018, 25, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Barrow, T.M.; Barault, L.; Ellsworth, R.E.; Harris, H.R.; Binder, A.M.; Valente, A.L.; Shriver, C.D.; Michels, K.B. Aberrant methylation of imprinted genes is associated with negative hormone receptor status in invasive breast cancer. Int. J. Cancer 2015, 137, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Alfarsi, L.H.; El-Ansari, R.; Craze, M.L.; Masisi, B.K.; Mohammed, O.J.; Ellis, I.O.; Rakha, E.A.; Green, A.R. Co-Expression Effect of SLC7A5/SLC3A2 to Predict Response to Endocrine Therapy in Oestrogen-Receptor-Positive Breast Cancer. Int. J. Mol. Sci. 2020, 21, 1407. [Google Scholar] [CrossRef] [PubMed]
- Hisada, T.; Kondo, N.; Wanifuchi-Endo, Y.; Osaga, S.; Fujita, T.; Asano, T.; Uemoto, Y.; Nishikawa, S.; Katagiri, Y.; Terada, M.; et al. Co-expression effect of LLGL2 and SLC7A5 to predict prognosis in ERα-positive breast cancer. Sci. Rep. 2022, 12, 16515. [Google Scholar] [CrossRef]
- Thakkar, A.; Raj, H.; Ravishankar; Muthuvelan, B.; Balakrishnan, A.; Padigaru, M. High Expression of Three-Gene Signature Improves Prediction of Relapse-Free Survival in Estrogen Receptor-Positive and Node-Positive Breast Tumors. Biomark. Insights 2015, 10, 103–112. [Google Scholar] [CrossRef]
- Shi, Y.; Zhao, Y.; Zhang, Y.; AiErken, N.; Shao, N.; Ye, R.; Lin, Y.; Wang, S. AFF3 upregulation mediates tamoxifen resistance in breast cancers. J. Exp. Clin. Cancer Res. 2018, 37, 254. [Google Scholar] [CrossRef]
- Liang, X.; Zhao, Y.; Fang, Z.; Shao, N.; Zhai, D.; Zhang, M.; Yu, L.; Shi, Y. DLGAP1-AS2 promotes estrogen receptor signalling and confers tamoxifen resistance in breast cancer. Mol. Biol. Rep. 2022, 49, 3939–3947. [Google Scholar] [CrossRef]
- Kim, S.; You, D.; Jeong, Y.; Yoon, S.Y.; Kim, S.A.; Lee, J.E. Inhibition of platelet-derived growth factor receptor synergistically increases the pharmacological effect of tamoxifen in estrogen receptor α positive breast cancer. Oncol. Lett. 2021, 21, 294. [Google Scholar] [CrossRef]
- Galmés-Pascual, B.M.; Nadal-Casellas, A.; Bauza-Thorbrügge, M.; Sbert-Roig, M.; García-Palmer, F.J.; Proenza, A.M.; Gianotti, M.; Lladó, I. 17β-estradiol improves hepatic mitochondrial biogenesis and function through PGC1B. J. Endocrinol. 2017, 232, 297–308. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, S.; Wang, Z.; Feng, X.; Liu, P.; Lv, X.B.; Li, F.; Yu, F.X.; Sun, Y.; Yuan, H.; et al. Estrogen regulates Hippo signaling via GPER in breast cancer. J. Clin. Investig. 2015, 125, 2123–2135. [Google Scholar] [CrossRef]
- Ye, L.; Li, F.; Song, Y.; Yu, D.; Xiong, Z.; Li, Y.; Shi, T.; Yuan, Z.; Lin, C.; Wu, X.; et al. Overexpression of CDCA7 predicts poor prognosis and induces EZH2-mediated progression of triple-negative breast cancer. Int. J. Cancer 2018, 143, 2602–2613. [Google Scholar] [CrossRef]
- Zhang, W.; Qiu, X.; Sun, D.; Zhang, D.; Qi, Y.; Li, X.; Liu, B.; Liu, J.; Lin, B. Systematic Analysis of the Clinical Relevance of Cell Division Cycle Associated Family in Endometrial Carcinoma. J. Cancer 2020, 11, 5588–5600. [Google Scholar] [CrossRef]
- Kim, H.R.; Kim, Y.S.; Yoon, J.A.; Lyu, S.W.; Shin, H.; Lim, H.J.; Hong, S.H.; Lee, D.R.; Song, H. Egr1 is rapidly and transiently induced by estrogen and bisphenol A via activation of nuclear estrogen receptor-dependent ERK1/2 pathway in the uterus. Reprod. Toxicol. 2014, 50, 60–67. [Google Scholar] [CrossRef]
- Shajahan-Haq, A.N.; Boca, S.M.; Jin, L.; Bhuvaneshwar, K.; Gusev, Y.; Cheema, A.K.; Demas, D.D.; Raghavan, K.S.; Michalek, R.; Madhavan, S.; et al. EGR1 regulates cellular metabolism and survival in endocrine resistant breast cancer. Oncotarget 2017, 8, 96865–96884. [Google Scholar] [CrossRef]
- Zahra, A.; Dong, Q.; Hall, M.; Jeyaneethi, J.; Silva, E.; Karteris, E.; Sisu, C. Identification of Potential Bisphenol A (BPA) Exposure Biomarkers in Ovarian Cancer. J. Clin. Med. 2021, 10, 1979. [Google Scholar] [CrossRef]
- Zhou, W.J.; Zhang, J.; Yang, H.L.; Wu, K.; Xie, F.; Wu, J.N.; Wang, Y.; Yao, L.; Zhuang, Y.; Xiang, J.D.; et al. Estrogen inhibits autophagy and promotes growth of endometrial cancer by promoting glutamine metabolism. Cell Commun. Signal. 2019, 17, 99. [Google Scholar] [CrossRef]
- Zhang, X.; Guo, N.; Jin, H.; Liu, R.; Zhang, Z.; Cheng, C.; Fan, Z.; Zhang, G.; Xiao, M.; Wu, S.; et al. Bisphenol A drives di (2-ethylhexyl) phthalate promoting thyroid tumorigenesis via regulating HDAC6/PTEN and c-MYC signaling. J. Hazard. Mater. 2022, 425, 127911. [Google Scholar] [CrossRef]
- Cheng, R.; Qi, L.; Kong, X.; Wang, Z.; Fang, Y.; Wang, J. Identification of the Significant Genes Regulated by Estrogen Receptor in Estrogen Receptor-Positive Breast Cancer and Their Expression Pattern Changes When Tamoxifen or Fulvestrant Resistance Occurs. Front. Genet. 2020, 11, 538734. [Google Scholar] [CrossRef]
- Kallak, T.K.; Baumgart, J.; Nilsson, K.; Åkerud, H.; Poromaa, I.S.; Stavreus-Evers, A. Vaginal Gene Expression During Treatment with Aromatase Inhibitors. Clin. Breast Cancer 2015, 15, 527–535.e2. [Google Scholar] [CrossRef]
- Yin, L.; Duan, J.J.; Bian, X.W.; Yu, S.C. Triple-negative breast cancer molecular subtyping and treatment progress. Breast Cancer Res. 2020, 22, 61. [Google Scholar] [CrossRef]
- Jorgensen, E.M.; Alderman, M.H., III; Taylor, H.S. Preferential epigenetic programming of estrogen response after in utero xenoestrogen (bisphenol-A) exposure. FASEB J. 2016, 30, 3194–3201. [Google Scholar] [CrossRef] [PubMed]
- Nallasamy, S.; Yoshida, K.; Akins, M.; Myers, K.; Iozzo, R.; Mahendroo, M. Steroid Hormones Are Key Modulators of Tissue Mechanical Function via Regulation of Collagen and Elastic Fibers. Endocrinology 2017, 158, 950–962. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, D.J.; Kuo, W.W.; Lai, Y.P.; Shibu, M.A.; Shen, C.Y.; Pai, P.; Yeh, Y.L.; Lin, J.Y.; Viswanadha, V.P.; Huang, C.Y. 17β-Estradiol and/or Estrogen Receptor β Attenuate the Autophagic and Apoptotic Effects Induced by Prolonged Hypoxia Through HIF-1α-Mediated BNIP3 and IGFBP-3 Signaling Blockage. Cell. Physiol. Biochem. 2015, 36, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, J.C.; Vanoni, S.; Zeng, C.; Waggoner, L.; Yang, Y.; Wu, D.; Uddin, J.; Karns, R.; Kottyan, L.; Mukkada, V.; et al. 17β-Estradiol protects the esophageal epithelium from IL-13-induced barrier dysfunction and remodeling. J. Allergy Clin. Immunol. 2019, 143, 2131–2146. [Google Scholar] [CrossRef]
- Casa, A.J.; Potter, A.S.; Malik, S.; Lazard, Z.; Kuiatse, I.; Kim, H.T.; Tsimelzon, A.; Creighton, C.J.; Hilsenbeck, S.G.; Brown, P.H.; et al. Estrogen and insulin-like growth factor-I (IGF-I) independently down-regulate critical repressors of breast cancer growth. Breast Cancer Res. Treat. 2012, 132, 61–73. [Google Scholar] [CrossRef]
- Northrop-Albrecht, E.J.; Rich, J.J.J.; Cushman, R.A.; Yao, R.; Ge, X.; Perry, G.A. Influence of conceptus presence and preovulatory estradiol exposure on uterine gene transcripts and proteins around maternal recognition of pregnancy in beef cattle. Mol. Cell. Endocrinol. 2022, 540, 111508. [Google Scholar] [CrossRef]
- Mi, Y.; Liu, F.; Liang, X.; Liu, S.; Huang, X.; Sang, M.; Geng, C. Tumor suppressor let-7a inhibits breast cancer cell proliferation, migration and invasion by targeting MAGE-A1. Neoplasma 2019, 66, 54–62. [Google Scholar] [CrossRef]
- Fan, D.; Ren, B.; Yang, X.; Liu, J.; Zhang, Z. Upregulation of miR-501-5p activates the wnt/β-catenin signaling pathway and enhances stem cell-like phenotype in gastric cancer. J. Exp. Clin. Cancer Res. 2016, 35, 177. [Google Scholar] [CrossRef]
- Gao, S.; Bian, T.; Su, M.; Liu, Y.; Zhang, Y. miR-26a inhibits ovarian cancer cell proliferation, migration and invasion by targeting TCF12. Oncol. Rep. 2020, 43, 368–374. [Google Scholar] [CrossRef]
- Jain, N.; Roy, J.; Das, B.; Mallick, B. miR-197-5p inhibits sarcomagenesis and induces cellular senescence via repression of KIAA0101. Mol. Carcinog. 2019, 58, 1376–1388. [Google Scholar] [CrossRef]
- Dai, T.; Liang, J.; Liu, W.; Zou, Y.; Niu, F.; Li, M.; Zhang, H.; Li, C.; Fan, M.; Cui, G. The miRNA mir-582-3p suppresses ovarian cancer progression by targeting AKT/MTOR signaling via lncRNA TUG1. Bioengineered 2021, 12, 10771–10781. [Google Scholar] [CrossRef]
- Mauro, T.; Hao, L.; Pop, L.C.; Buckley, B.; Schneider, S.H.; Bandera, E.V.; Shapses, S.A. Circulating zearalenone and its metabolites differ in women due to body mass index and food intake. Food Chem. Toxicol. 2018, 116 Pt B, 227–232. [Google Scholar] [CrossRef]
- Kong, L.; Zhao, A.H.; Wang, Q.W.; Feng, Y.Q.; Yan, Z.H.; Li, M.H.; Zhang, F.L.; Wang, H.; Shen, K.Y.; Liu, Y.; et al. Maternal Zearalenone exposure impacted ovarian follicle formation and development of suckled offspring. Sci. Total Environ. 2021, 788, 147792. [Google Scholar] [CrossRef]
- Kowalska, K.; Habrowska-Górczyńska, D.E.; Urbanek, K.A.; Domińska, K.; Piastowska-Ciesielska, A.W. Estrogen Receptor α Is Crucial in Zearalenone-Induced Invasion and Migration of Prostate Cancer Cells. Toxins 2018, 10, 98. [Google Scholar] [CrossRef]
- Lecomte, S.; Demay, F.; Pham, T.H.; Moulis, S.; Efstathiou, T.; Chalmel, F.; Pakdel, F. Deciphering the Molecular Mechanisms Sustaining the Estrogenic Activity of the Two Major Dietary Compounds Zearalenone and Apigenin in ER-Positive Breast Cancer Cell Lines. Nutrients 2019, 11, 237. [Google Scholar] [CrossRef]
- Parveen, M.; Zhu, Y.; Kiyama, R. Expression profiling of the genes responding to zearalenone and its analogues using estrogen-responsive genes. FEBS Lett. 2009, 583, 2377–2384. [Google Scholar] [CrossRef]
- Zhang, G.L.; Zhang, R.Q.; Sun, X.F.; Cheng, S.F.; Wang, Y.F.; Ji, C.L.; Feng, Y.Z.; Yu, J.; Ge, W.; Zhao, Y.; et al. RNA-seq based gene expression analysis of ovarian granulosa cells exposed to zearalenone in vitro: Significance to steroidogenesis. Oncotarget 2017, 8, 64001–64014. [Google Scholar] [CrossRef]
- So, M.Y.; Tian, Z.; Phoon, Y.S.; Sha, S.; Antoniou, M.N.; Zhang, J.; Wu, R.S.; Tan-Un, K.C. Gene expression profile and toxic effects in human bronchial epithelial cells exposed to zearalenone. PLoS ONE 2014, 9, e96404. [Google Scholar] [CrossRef]
- Li, Z.; Yin, H.; Shen, Y.; Ren, M.; Xu, X. The influence of phenolic environmental estrogen on the transcriptome of uterine leiomyoma cells: A whole transcriptome profiling-based analysis. Ecotoxicol. Environ. Saf. 2021, 211, 111945. [Google Scholar] [CrossRef]
- Kim, C.H.; Kim, M.J.; Park, J.; Kim, J.; Kim, J.Y.; An, M.J.; Shin, G.S.; Lee, H.M.; Kim, J.W. Bisphenol a Exposure Changes the Transcriptomic and Proteomic Dynamics of Human Retinoblastoma Y79 Cells. Genes 2021, 12, 264. [Google Scholar] [CrossRef]
- Li, Z.; Lyu, C.; Ren, Y.; Wang, H. Role of TET Dioxygenases and DNA Hydroxymethylation in Bisphenols-Stimulated Proliferation of Breast Cancer Cells. Environ. Health Perspect. 2020, 128, 27008. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yin, H.; Chen, K.; Ding, B.; Xu, J.; Ren, M.; Zhang, C.; Shen, Y. Effects of bisphenol A on uterine leiomyoma: In vitro and in vivo evaluation with mechanistic insights related to XBP1. Ecotoxicol. Environ. Saf. 2022, 247, 114201. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Burns, K.A.; Arao, Y.; Luh, C.J.; Korach, K.S. Differential estrogenic actions of endocrine-disrupting chemicals bisphenol A, bisphenol AF, and zearalenone through estrogen receptor α and β in vitro. Environ. Health Perspect. 2012, 120, 1029–1035. [Google Scholar] [CrossRef] [PubMed]
- Gertz, J.; Reddy, T.E.; Varley, K.E.; Garabedian, M.J.; Myers, R.M. Genistein and bisphenol A exposure cause estrogen receptor 1 to bind thousands of sites in a cell type-specific manner. Genome Res. 2012, 22, 2153–2162. [Google Scholar] [CrossRef]
- Acconcia, F.; Pallottini, V.; Marino, M. Molecular Mechanisms of Action of BPA. Dose Response 2015, 13, 1559325815610582. [Google Scholar] [CrossRef]
- Lo, E.K.K.; Lee, J.C.; Turner, P.C.; El-Nezami, H. Low dose of zearalenone elevated colon cancer cell growth through G protein-coupled estrogenic receptor. Sci. Rep. 2021, 11, 7403. [Google Scholar] [CrossRef]
- Sang, C.; Song, Y.; Jin, T.W.; Zhang, S.; Fu, L.; Zhao, Y.; Zou, X.; Wang, Z.; Gao, H.; Liu, S. Bisphenol A induces ovarian cancer cell proliferation and metastasis through estrogen receptor-α pathways. Environ. Sci. Pollut. Res. Int. 2021, 28, 36060–36068. [Google Scholar] [CrossRef]
- Deng, P.; Tan, M.; Zhou, W.; Chen, C.; Xi, Y.; Gao, P.; Ma, Q.; Liang, Y.; Chen, M.; Tian, L.; et al. Bisphenol A promotes breast cancer cell proliferation by driving miR-381-3p-PTTG1-dependent cell cycle progression. Chemosphere 2021, 268, 129221. [Google Scholar] [CrossRef]
- Jun, J.H.; Oh, J.E.; Shim, J.K.; Kwak, Y.L.; Cho, J.S. Effects of bisphenol A on the proliferation, migration, and tumor growth of colon cancer cells: In vitro and in vivo evaluation with mechanistic insights related to ERK and 5-HT3. Food Chem. Toxicol. 2021, 158, 112662. [Google Scholar] [CrossRef]
- Wang, K.; Huang, D.; Zhou, P.; Su, X.; Yang, R.; Shao, C.; Ma, A.; Wu, J. Individual and Combined Effect of Bisphenol A and Bisphenol AF on Prostate Cell Proliferation through NF-κB Signaling Pathway. Int. J. Mol. Sci. 2022, 23, 12283. [Google Scholar] [CrossRef]
- Pelletier, J.; Thomas, G.; Volarević, S. Ribosome biogenesis in cancer: New players and therapeutic avenues. Nat. Rev. Cancer 2018, 18, 51–63. [Google Scholar] [CrossRef]
- Ray, S.; Johnston, R.; Campbell, D.C.; Nugent, S.; McDade, S.S.; Waugh, D.; Panov, K.I. Androgens and estrogens stimulate ribosome biogenesis in prostate and breast cancer cells in receptor dependent manner. Gene 2013, 526, 46–53. [Google Scholar] [CrossRef]
- Saito, Y.; Soga, T. Amino acid transporters as emerging therapeutic targets in cancer. Cancer Sci. 2021, 112, 2958–2965. [Google Scholar] [CrossRef]
- Karantza, V. Keratins in health and cancer: More than mere epithelial cell markers. Oncogene 2011, 30, 127–138. [Google Scholar] [CrossRef]
- Kim, H.J.; Choi, W.J.; Lee, C.H. Phosphorylation and Reorganization of Keratin Networks: Implications for Carcinogenesis and Epithelial Mesenchymal Transition. Biomol. Ther. 2015, 23, 301–312. [Google Scholar] [CrossRef]
- Tzanakakis, G.; Kavasi, R.M.; Voudouri, K.; Berdiaki, A.; Spyridaki, I.; Tsatsakis, A.; Nikitovic, D. Role of the extracellular matrix in cancer-associated epithelial to mesenchymal transition phenomenon. Dev. Dyn. 2018, 247, 368–381. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, Z.; Cenciarini, M.E.; Proietti, C.J.; Amasino, M.; Hong, T.; Yang, M.; Liao, Y.; Chiang, H.C.; Kaklamani, V.G.; et al. Tamoxifen Resistance in Breast Cancer Is Regulated by the EZH2-ERα-GREB1 Transcriptional Axis. Cancer Res. 2018, 78, 671–684. [Google Scholar] [CrossRef]
- Thakkar, A.D.; Raj, H.; Chakrabarti, D.; Ravishankar; Saravanan, N.; Muthuvelan, B.; Balakrishnan, A.; Padigaru, M. Identification of gene expression signature in estrogen receptor positive breast carcinoma. Biomark. Cancer 2010, 2, BIC.S3793. [Google Scholar] [CrossRef]
- Yip, K.Y.; Wan, M.L.Y.; Wong, A.S.T.; Korach, K.S.; El-Nezami, H. Combined low-dose zearalenone and aflatoxin B1 on cell growth and cell-cycle progression in breast cancer MCF-7 cells. Toxicol. Lett. 2017, 281, 139–151. [Google Scholar] [CrossRef]
- Kowalska, K.; Habrowska-Górczyńska, D.E.; Domińska, K.; Urbanek, K.A.; Piastowska-Ciesielska, A.W. ERβ and NFκB—Modulators of zearalenone-induced oxidative stress in human prostate cancer cells. Toxins 2020, 12, 199. [Google Scholar] [CrossRef] [Green Version]
- Taranu, I.; Braicu, C.; Marin, D.E.; Pistol, G.C.; Motiu, M.; Balacescu, L.; Burlacu, R. Exposure to zearalenone mycotoxin alters in vitro porcine intestinal epithelial cells by differential gene expression. Toxicol. Lett. 2015, 232, 310–325. [Google Scholar] [CrossRef] [PubMed]
- Heneweer, M.; Houtman, R.; Poortman, J.; Groot, M.; Maliepaard, C.; Peijnenburg, A. Estrogenic effects in the immature rat uterus after dietary exposure to ethinylestradiol and zearalenone using a systems biology approach. Toxicol. Sci. 2007, 99, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Alderman, M.H.; Asgari, C.; Taylor, H.S. Fetal Bisphenol-a induced changes in murine behavior and brain gene expression persisted in adult-aged offspring. Endocrinology 2020, 161, bqaa164. [Google Scholar] [CrossRef] [PubMed]
- Szilágyi, M.; Pös, O.; Márton, É.; Buglyó, G.; Soltész, B.; Keserű, J.; Penyige, A.; Szemes, T.; Nagy, B. Circulating Cell-Free Nucleic Acids: Main Characteristics and Clinical Application. Int. J. Mol. Sci. 2020, 21, 6827. [Google Scholar] [CrossRef] [PubMed]
- Rupaimoole, R.; Calin, G.A.; Lopez-Berestein, G.; Sood, A.K. miRNA Deregulation in Cancer Cells and the Tumor Microenvironment. Cancer Discov. 2016, 6, 235–246. [Google Scholar] [CrossRef]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef]
- Klinge, C.M. miRNAs regulated by estrogens, tamoxifen, and endocrine disruptors and their downstream gene targets. Mol. Cell. Endocrinol. 2015, 418 Pt 3, 273–297. [Google Scholar] [CrossRef]
- Wang, H.; Jin, J.; Wu, J.; Qu, H.; Wu, S.; Bao, W. Transcriptome and chromatin accessibility in porcine intestinal epithelial cells upon Zearalenone exposure. Sci. Data 2019, 6, 298. [Google Scholar] [CrossRef]
- Wang, M.; Wu, W.; Li, L.; He, J.; Huang, S.; Chen, S.; Chen, J.; Long, M.; Yang, S.; Li, P. Analysis of the miRNA Expression Profiles in the Zearalenone-Exposed TM3 Leydig Cell Line. Int. J. Mol. Sci. 2019, 20, 635. [Google Scholar] [CrossRef]
- Grenier, B.; Hackl, M.; Skalicky, S.; Thamhesl, M.; Moll, W.D.; Berrios, R.; Schatzmayr, G.; Nagl, V. MicroRNAs in porcine uterus and serum are affected by zearalenone and represent a new target for mycotoxin biomarker discovery. Sci. Rep. 2019, 9, 9408. [Google Scholar] [CrossRef] [Green Version]
- Mao, J.; Kinkade, J.A.; Bivens, N.J.; Rosenfeld, C.S. miRNA changes in the mouse placenta due to bisphenol A exposure. Epigenomics 2021, 13, 1909–1919. [Google Scholar] [CrossRef]
- Rosenfeld, C.S. Transcriptomics and Other Omics Approaches to Investigate Effects of Xenobiotics on the Placenta. Front. Cell Dev. Biol. 2021, 9, 723656. [Google Scholar] [CrossRef]
- Rasdi, Z.; Kamaludin, R.; Ab Rahim, S.; Syed Ahmad Fuad, S.B.; Othman, M.H.D.; Siran, R.; Mohd Nor, N.S.; Abdul Hamid Hasani, N.; Sheikh Abdul Kadir, S.H. The impacts of intrauterine Bisphenol A exposure on pregnancy and expression of miRNAs related to heart development and diseases in animal model. Sci. Rep. 2020, 10, 5882. [Google Scholar] [CrossRef]
- Ostry, V.; Malir, F.; Toman, J.; Grosse, Y. Mycotoxins as human carcinogens-the IARC Monographs classification. Mycotoxin Res. 2017, 33, 65–73. [Google Scholar] [CrossRef]
- Márton, É.; Lukács, J.; Penyige, A.; Janka, E.; Hegedüs, L.; Soltész, B.; Méhes, G.; Póka, R.; Nagy, B.; Szilágyi, M. Circulating epithelial-mesenchymal transition-associated miRNAs are promising biomarkers in ovarian cancer. J. Biotechnol. 2019, 297, 58–65. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | E2 | ZEA | BPA | Reference | |||
|---|---|---|---|---|---|---|---|
| Log2FC | FDR | Log2FC | FDR | Log2FC | FDR | ||
| GREB1 | 4.71 | 7.39 × 10−11 | 4.79 | 3.95 × 10−8 | 3.64 | 3.61 × 10−12 | [26,27] |
| KCNF1 | 4.19 | 2.15 × 10−10 | 4.23 | 7.10 × 10−8 | 3.27 | 8.96 × 10−12 | |
| DEPTOR | 4.17 | 7.39 × 10−11 | 3.77 | 5.25 × 10−8 | 2.79 | 8.96 × 10−12 | [28] |
| CA12 | 4.08 | 7.39 × 10−11 | 4.10 | 2.98 × 10−8 | 2.74 | 4.80 × 10−12 | [29,30] |
| AGT | 3.31 | 2.69 × 10−10 | 3.03 | 5.25 × 10−8 | 2.21 | 1.38 × 10−11 | [31] |
| MGAT3 | 3.31 | 2.15 × 10−10 | 3.25 | 3.15 × 10−8 | 2.03 | 4.05 × 10−12 | |
| CBLN1 | 3.29 | 2.71 × 10−10 | 3.10 | 1.20 × 10−7 | 2.64 | 3.74 × 10−12 | [32] |
| OLFM1 | 3.28 | 1.43 × 10−9 | 3.14 | 6.37 × 10−7 | 1.79 | 1.29 × 10−8 | [33,34] |
| ARHGAP26 | 3.25 | 2.22 × 10−10 | 3.35 | 3.95 × 10−8 | 2.33 | 8.95 × 10−12 | |
| SEZ6 | 3.21 | 1.04 × 10−8 | 3.18 | 1.61 × 10−7 | 2.49 | 3.15 × 10−12 | |
| PPP1R1A | 3.13 | 2.08 × 10−10 | 2.82 | 2.98 × 10−8 | 1.26 | 4.58 × 10−10 | |
| CADM1 | 3.11 | 2.69 × 10−10 | 3.13 | 3.37 × 10−8 | 2.11 | 6.99 × 10−12 | [35] |
| PIPOX | 3.05 | 9.83 × 10−8 | 2.95 | 5.97 × 10−6 | 2.36 | 4.42 × 10−8 | |
| DHRS2 | 2.98 | 5.99 × 10−10 | 2.78 | 1.17 × 10−7 | 1.50 | 2.70 × 10−9 | [36,37] |
| PKDCC | 2.97 | 6.56 × 10−10 | 2.95 | 2.98 × 10−8 | 2.60 | 4.05 × 10−12 | |
| SLC7A2 | 2.93 | 1.43 × 10−9 | 2.92 | 2.89 × 10−7 | 2.04 | 2.70 × 10−10 | [38,39,40] |
| HPDL | 2.81 | 2.51 × 10−8 | 2.63 | 3.42 × 10−6 | 1.82 | 7.26 × 10−8 | |
| CABLES1 | 2.73 | 2.94 × 10−10 | 2.55 | 3.14 × 10−8 | 1.58 | 2.63 × 10−11 | |
| RHOBTB1 | 2.72 | 9.85 × 10−10 | 2.61 | 8.73 × 10−8 | 1.86 | 9.33 × 10−11 | |
| RBBP8 | 2.71 | 2.69 × 10−10 | 2.60 | 3.95 × 10−8 | 1.92 | 1.49 × 10−11 | [41] |
| RUBCNL | 2.69 | 1.05 × 10−8 | 2.58 | 1.00 × 10−6 | 1.98 | 1.35 × 10−8 | |
| DIRAS3 | 2.58 | 6.56 × 10−10 | 2.54 | 6.41 × 10−8 | 1.86 | 1.94 × 10−11 | [42] |
| LONRF2 | 2.49 | 1.43 × 10−9 | 2.63 | 1.61 × 10−7 | 1.79 | 1.78 × 10−10 | |
| RIMS4 | 2.35 | 3.09 × 10−9 | 2.11 | 9.53 × 10−8 | 1.32 | 3.73 × 10−10 | |
| SLC7A5 | 2.32 | 9.02 × 10−10 | 2.13 | 8.63 × 10−8 | 1.29 | 1.20 × 10−9 | [43,44] |
| PLAT | 2.30 | 6.56 × 10−10 | 1.94 | 2.36 × 10−6 | 1.17 | 9.17 × 10−9 | [45] |
| CISH | 2.18 | 1.06 × 10−8 | 2.13 | 6.54 × 10−7 | 1.77 | 2.70 × 10−10 | |
| AFF3 | 2.11 | 1.77 × 10−8 | 2.38 | 6.58 × 10−7 | 2.04 | 2.11 × 10−10 | [46,47] |
| PDGFRL | 2.06 | 1.41 × 10−8 | 1.65 | 7.60 × 10−7 | 1.19 | 4.13 × 10−9 | [48] |
| RASGRP1 | 2.03 | 2.41 × 10−8 | 2.11 | 8.43 × 10−7 | 1.65 | 9.16 × 10−10 | |
| BEND3 | 2.00 | 1.47 × 10−8 | 1.99 | 9.05 × 10−7 | 1.34 | 7.40 × 10−8 | |
| PPARGC1B | 1.95 | 1.49 × 10−7 | 2.09 | 6.75 × 10−6 | 1.36 | 9.28 × 10−8 | [49] |
| MTHFD2P7 | 1.93 | 3.40 × 10−8 | 1.61 | 8.43 × 10−7 | 1.36 | 1.40 × 10−8 | |
| ERICH3 | 1.88 | 1.37 ×10−7 | 1.90 | 3.96 × 10−6 | 1.34 | 5.68 × 10−8 | |
| PLCB1 | 1.87 | 9.19 × 10−8 | 1.79 | 2.36 × 10−6 | 1.01 | 5.31 × 10−7 | [50] |
| CDCA7 | 1.85 | 1.04 × 10−8 | 1.74 | 4.73 × 10−7 | 1.32 | 2.00 × 10−9 | [51,52] |
| OLFML3 | 1.84 | 1.41 × 10−8 | 1.79 | 6.54 × 10−7 | 1.08 | 1.20 × 10−8 | |
| SLC22A3 | 1.84 | 9.64 × 10−9 | 1.81 | 2.89 × 10−7 | 1.47 | 1.31 × 10−10 | |
| EGR1 | 1.78 | 2.67 × 10−8 | 1.71 | 9.58 × 10−7 | 1.22 | 9.03 × 10−9 | [53,54] |
| SLITRK4 | 1.76 | 5.89 × 10−8 | 1.76 | 1.40 × 10−6 | 1.15 | 3.05 × 10−7 | |
| PNPT1P1 | 1.73 | 8.04 × 10−8 | 1.19 | 3.83 × 10−5 | 1.46 | 4.42 × 10−8 | |
| RRS1 | 1.72 | 1.49 × 10−8 | 1.49 | 9.13 × 10−7 | 1.12 | 2.30 × 10−8 | [55] |
| ZNF239 | 1.70 | 4.87 × 10−8 | 1.55 | 2.42 × 10−6 | 1.14 | 3.21 × 10−8 | |
| NPY1R | 1.69 | 1.99 × 10−8 | 1.54 | 3.45 × 10−6 | 1.32 | 2.19 × 10−9 | [36] |
| SLC6A15 | 1.66 | 1.75 × 10−7 | 1.72 | 2.65 × 10−6 | 1.27 | 3.28 × 10−8 | |
| TSEN2 | 1.66 | 2.68 × 10−7 | 1.49 | 1.14 × 10−5 | 1.10 | 3.82 × 10−7 | |
| ZNF485 | 1.64 | 5.17 × 10−7 | 1.77 | 9.08 × 10−7 | 1.38 | 3.22 × 10−9 | |
| MYC | 1.63 | 1.49 ×10−8 | 1.65 | 4.36 × 10−7 | 1.26 | 4.51 × 10−10 | [56,57] |
| NCR3LG1 | 1.62 | 1.94 × 10−7− | 1.86 | 3.42 × 10−6 | 1.34 | 8.40 × 10−8 | [40] |
| TFAP4 | 1.61 | 1.36 × 10−7 | 1.58 | 2.23 × 10−6 | 1.10 | 5.87 × 10−8 | |
| KBTBD8 | 1.59 | 2.61 × 10−8 | 1.55 | 9.05 × 10−7 | 1.01 | 1.29 × 10−8 | |
| SLC19A2 | 1.57 | 8.04 × 10−8 | 1.58 | 9.05 × 10−7 | 1.01 | 1.51 × 10−8 | |
| LYAR | 1.56 | 5.51 × 10−8 | 1.33 | 1.40 × 10−6 | 1.02 | 5.39 × 10−8 | |
| KAZN | 1.46 | 4.96 × 10−8 | 1.45 | 9.08 × 10−7 | 1.09 | 9.52 × 10−9 | |
| PUS7 | 1.41 | 5.51 × 10−8 | 1.35 | 9.28 × 10−7 | 1.03 | 9.66 × 10−9 | [58] |
| ERG | 1.40 | 1.84 × 10−8 | 1.20 | 4.52 × 10−5 | 1.03 | 7.90 × 10−8 | |
| NXNL2 | 1.28 | 1.69 × 10−6 | 1.25 | 4.68 × 10−5 | 1.27 | 8.75 × 10−8 | |
| CLDN8 | −1.32 | 3.21 × 10−7 | −2.11 | 5.65 × 10−7 | −1.03 | 3.19 × 10−7 | [59,60] |
| LINC01133 | −1.42 | 2.42 × 10−7 | −1.26 | 5.64 × 10−6 | −1.01 | 2.91 × 10−7 | |
| MFAP2 | −1.55 | 1.31 × 10−7 | −1.80 | 5.65 × 10−7 | −1.22 | 7.07 × 10−8 | [61,62] |
| IGSF9 | −1.60 | 5.90 × 10−8 | −1.57 | 7.60 × 10−7 | −1.03 | 4.15 × 10−8 | |
| KRTAP2-3 | −1.60 | 5.51 × 10−8 | −1.35 | 9.58 × 10−7 | −1.02 | 2.80 × 10−8 | |
| UNC5B-AS1 | −1.62 | 6.74 × 10−8 | −1.38 | 5.76 × 10−8 | −1.12 | 5.76 ×10−8 | |
| BMF | −1.64 | 1.99 × 10−8 | −1.65 | 4.90 × 10−9 | −1.12 | 4.90 × 10−9 | |
| EPHB3 | −1.64 | 2.84 × 10−8 | −1.62 | 4.08 × 10−8 | −1.04 | 4.08 × 10−8 | |
| KRT4 | −1.71 | 2.13 × 10−8 | −1.67 | 1.10 × 10−8 | −1.01 | 1.10 × 10−8 | [55] |
| CYSRT1 | −1.72 | 1.69 × 10−8 | −2.06 | 1.33 × 10−8 | −1.10 | 1.33 × 10−8 | |
| RNF224 | −1.77 | 2.59 × 10−8 | −1.96 | 1.68 × 10−7 | −1.09 | 1.68 × 10−7 | |
| RASSF2 | −1.80 | 1.06 × 10−6 | −1.64 | 4.32 × 10−9 | −1.29 | 4.32 × 10−9 | |
| IKZF2 | −1.89 | 6.80 × 10−8 | −1.47 | 3.52 × 10−7 | −1.04 | 3.52 × 10−7 | |
| IGFBP3 | −1.89 | 1.04 × 10−8 | −1.82 | 5.02 × 10−9 | −1.06 | 5.02 × 10−9 | [63] |
| POU2F3 | −1.92 | 3.87 × 10−7 | −1.99 | 1.46 × 10−8 | −1.35 | 1.46 × 10−8 | |
| NDRG1 | −1.96 | 4.01 × 10−9 | −1.97 | 2.70 × 10−9 | −1.05 | 2.70 × 10−9 | |
| TGM1 | −2.00 | 2.22 × 10−8 | −1.91 | 3.82 × 10−8 | −1.00 | 3.82 × 10−8 | [64] |
| LINC01559 | −2.00 | 3.15 × 10−7 | −1.90 | 1.34 × 10−7 | −1.40 | 1.34 × 10−7 | |
| LRRC4 | −2.20 | 4.11 × 10−8 | −2.21 | 2.76 × 10−8 | −1.21 | 2.76 × 10−8 | |
| FILIP1L | −2.33 | 1.49 × 10−8 | −2.45 | 6.82 × 10−10 | −1.22 | 6.82 × 10−10 | |
| C4ORF26 | −2.45 | 1.61 × 10−7 | −2.18 | 2.30 × 10−8 | −1.29 | 7.14 × 10−8 | |
| NECTIN4 | −2.78 | 1.43 × 10−9 | −2.73 | 1.07 × 10−10 | −1.70 | 1.07 × 10−10 | |
| AIM1L | −2.95 | 1.04 × 10−8 | −2.50 | 3.21 × 10−10 | −1.48 | 3.21 × 10−10 | |
| BLNK | −3.07 | 2.53 × 10−7 | −3.03 | 2.70 × 10−9 | −1.90 | 2.70 × 10−9 | [65] |
| RBBP8NL | −3.19 | 1.28 × 10−7 | −3.46 | 1.42 × 10−8 | −2.07 | 1.42 × 10−8 | [66] |
| E2 | ZEA | BPA | ||||||
|---|---|---|---|---|---|---|---|---|
| miRNA | Log2FC | FDR | miRNA | Log2FC | FDR | miRNA | Log2FC | FDR |
| let-7a-2-3p | −1.99 | 0.001 | miR-6795-3p | −1.83 | 0.0002 | miR-6795-3p | −1.66 | 0.029 |
| miR-501-5p | −1.98 | 0.001 | miR-3661 | −1.68 | 0.052 | miR-597-5p | −1.33 | 0.05 |
| let-7g-3p | −1.98 | 0.017 | miR-501-5p | −1.55 | 0.033 | miR-197-5p | −1.21 | 0.046 |
| miR-3679-5p | −1.95 | 0.001 | miR-197-5p | −1.10 | 0.066 | miR-5008-5p | −1.09 | 0.037 |
| miR-26a-2-3p | −1.92 | 0.023 | miR-5008-5p | −1.07 | 0.013 | miR-320c | −1.03 | 0.065 |
| miR-326 | −1.65 | 0.035 | miR-451a | 1.09 | 0.039 | miR-6879-3p | 1.01 | 0.046 |
| miR-6795-3p | −1.41 | 0.035 | miR-3065-3p | 1.33 | 0.095 | miR-3934-5p | 1.11 | 0.065 |
| miR-1305 | −1.27 | 0.092 | miR-3620-3p | 1.46 | 0.049 | miR-590-5p | 1.11 | 0.046 |
| miR-197-5p | −1.27 | 0.026 | miR-6806-3p | 1.49 | 0.033 | miR-636 | 1.22 | 0.05 |
| miR-6765-3p | −1.16 | 0.035 | miR-4747-3p | 1.93 | 0.033 | miR-6806-3p | 1.45 | 0.046 |
| miR-5008-5p | −1.05 | 0.026 | miR-548u | 2.34 | 0.013 | |||
| miR-582-3p | 1.48 | 0.023 | ||||||
| miR-6775-3p | 2.48 | 0.026 | ||||||
| E2 | ZEA | BPA | ||||||
|---|---|---|---|---|---|---|---|---|
| miRNA | Deg. | Betw. | miRNA | Deg. | Betw. | miRNA | Deg. | Betw. |
| miR-1305 | 195 | 141,840 | miR-501-5p | 158 | 48,544 | miR-320c | 108 | 19,875 |
| miR-501-5p | 158 | 107,695 | miR-3620-3p | 95 | 41,791 | miR-197-5p | 68 | 13,869 |
| miR-326 | 138 | 96,436 | miR-197-5p | 68 | 27,959 | miR-590-5p | 66 | 20,688 |
| let-7g-3p | 106 | 43,469 | miR-3661 | 49 | 16,350 | |||
| let-7a-2-3p | 102 | 40,279 | miR-451a | 31 | 11,295 | |||
| miR-197-5p | 68 | 46,136 | ||||||
| miR-3679-5p | 63 | 45,319 | ||||||
| miR-26a-2-3p | 59 | 42,825 | ||||||
| miR-582-3p | 55 | 39,732 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Márton, É.; Varga, A.; Penyige, A.; Birkó, Z.; Balogh, I.; Nagy, B.; Szilágyi, M. Comparative Analysis of Transcriptomic Changes including mRNA and microRNA Expression Induced by the Xenoestrogens Zearalenone and Bisphenol A in Human Ovarian Cells. Toxins 2023, 15, 140. https://doi.org/10.3390/toxins15020140
Márton É, Varga A, Penyige A, Birkó Z, Balogh I, Nagy B, Szilágyi M. Comparative Analysis of Transcriptomic Changes including mRNA and microRNA Expression Induced by the Xenoestrogens Zearalenone and Bisphenol A in Human Ovarian Cells. Toxins. 2023; 15(2):140. https://doi.org/10.3390/toxins15020140
Chicago/Turabian StyleMárton, Éva, Alexandra Varga, András Penyige, Zsuzsanna Birkó, István Balogh, Bálint Nagy, and Melinda Szilágyi. 2023. "Comparative Analysis of Transcriptomic Changes including mRNA and microRNA Expression Induced by the Xenoestrogens Zearalenone and Bisphenol A in Human Ovarian Cells" Toxins 15, no. 2: 140. https://doi.org/10.3390/toxins15020140
APA StyleMárton, É., Varga, A., Penyige, A., Birkó, Z., Balogh, I., Nagy, B., & Szilágyi, M. (2023). Comparative Analysis of Transcriptomic Changes including mRNA and microRNA Expression Induced by the Xenoestrogens Zearalenone and Bisphenol A in Human Ovarian Cells. Toxins, 15(2), 140. https://doi.org/10.3390/toxins15020140