Multiplexed Host-Induced Gene Silencing of Aspergillus flavus Genes Confers Aflatoxin Resistance in Groundnut


 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
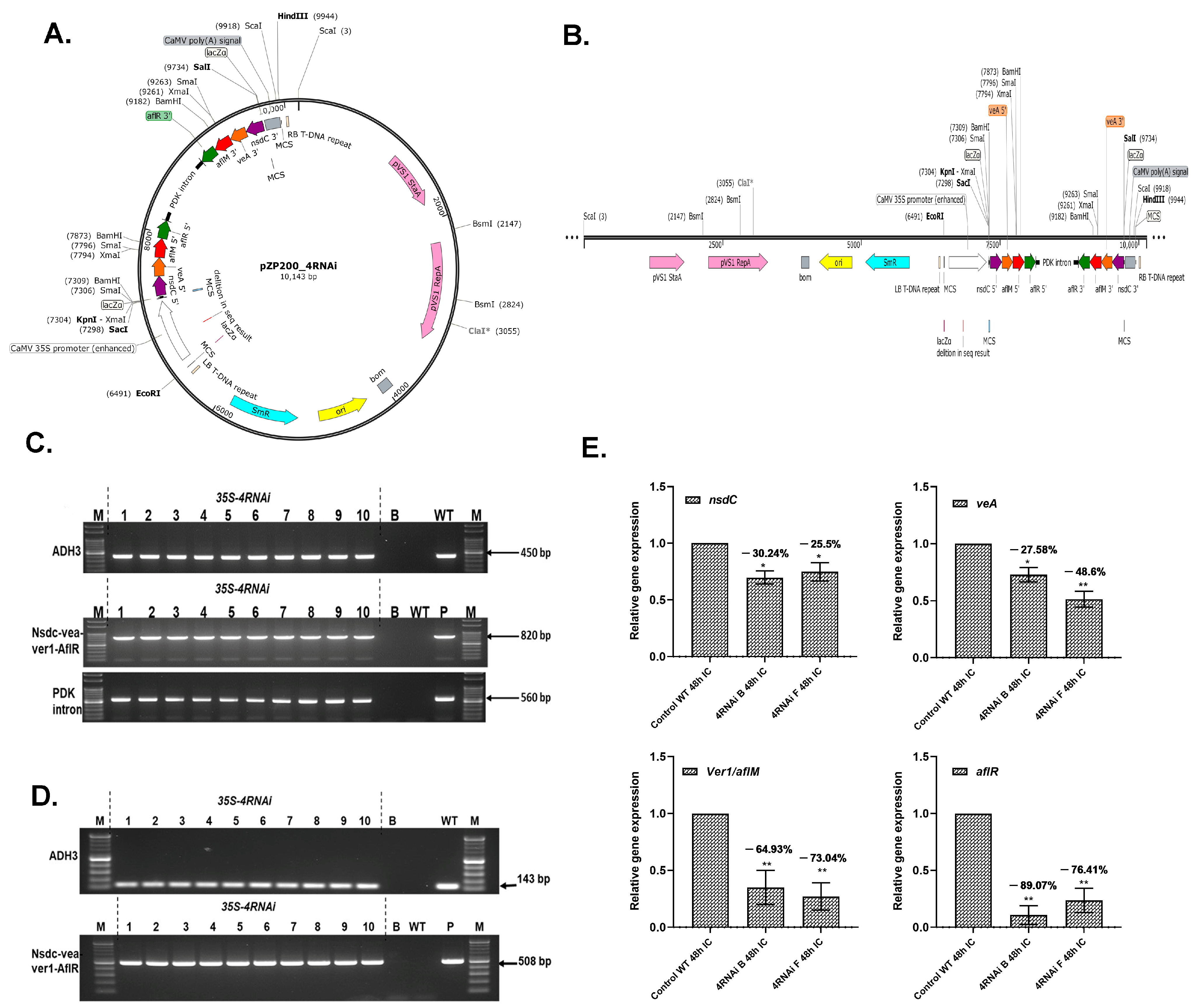
2.1. Generation of HIGS Lines Overexpressing 4RNAi Cassette
2.2. 4RNAi-Expressing HIGS Lines Demonstrate no Substantial Alterations in Gene Expression
2.3. HIGS Cotyledons Showed Significant Resistance to A. flavus Infection and Aflatoxin Contamination
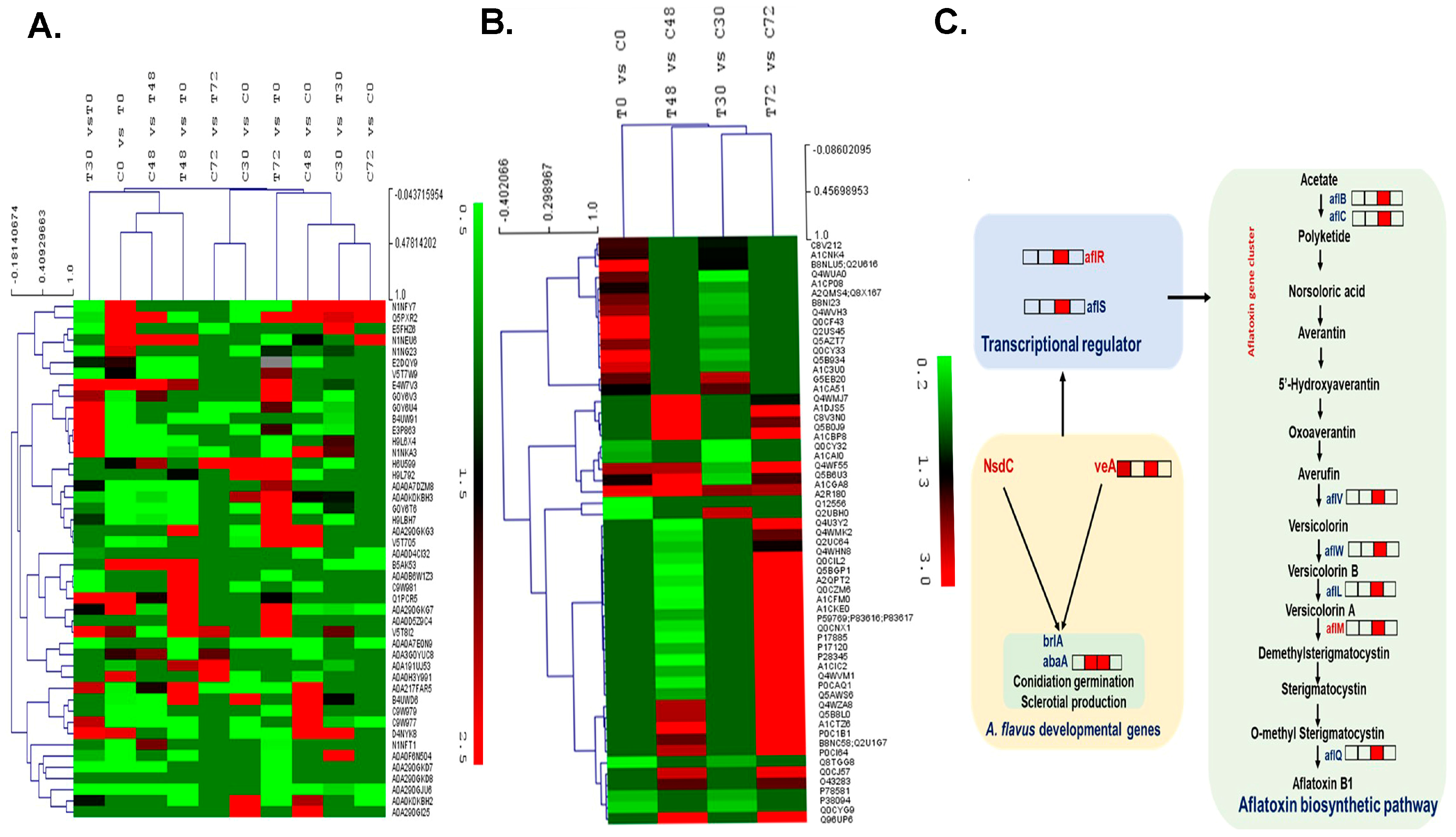
2.4. Impact of Aspergillus Infection on Groundnut Proteomes
2.5. Effect of Host-Induced Gene Silencing on the A. flavus Proteome
2.6. Differentially Expressed Proteins in the Groundnut Host System and Identification of Host Resistance-Associated Proteins
2.7. Identified Host Susceptibility-Associated Proteins
2.8. Validation of DEPs by qRT-PCR
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Generation of HIGS Groundnut Expressing RNAi Genes
5.2. Genotyping of HIGS Plants
5.3. Gene Expression Analyses
5.4. Copy Number Detection and Inheritance Studies
5.5. Aspergillus flavus Growth Conditions
5.6. Colonization of A. flavus on Kernels and Aflatoxin Analysis
5.7. Detection of Fungal Load
5.8. Extraction and Digestion of Total Proteins
5.9. UPLC-MS Analysis of Peptides
5.10. Identification and Quantification of Proteins
5.11. Functional Annotation and Pathway Mapping
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Amaike, S.; Keller, N.P. Aspergillus flavus. Ann. Rev. Phytopathol. 2011, 49, 107–133. [Google Scholar] [CrossRef] [PubMed]
- Antiga, L.; La Starza, S.R.; Miccoli, C.; D’angeli, S.; Scala, V.; Zaccaria, M.; Shu, X.; Obrian, G.; Beccaccioli, M.; Payne, G.A.; et al. Aspergillus flavus exploits maize kernels using an “Orphan” secondary metabolite cluster. Int. J. Mol. Sci. 2020, 21, 8213. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Groopman, J.D.; Pestka, J.J. Public health impacts of foodborne mycotoxins. Annu. Rev. Food Sci. Technol. 2014, 5, 351–372. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Pathak, H.; Bhadauria, S.; Sudan, J. Aflatoxin contamination in food crops: Causes, detection, and management: A review. Food Prod. Proc. Nutr. 2021, 3, 17. [Google Scholar] [CrossRef]
- Bhatnagar-Mathur, P.; Sunkara, S.; Bhatnagar-Panwar, M.; Waliyar, F.; Sharma, K.K. Biotechnological advances for combating Aspergillus flavus and aflatoxin contamination in crops. Plant Sci. 2015, 234, 119–132. [Google Scholar] [CrossRef]
- Pandey, M.K.; Kumar, R.; Pandey, A.K.; Soni, P.; Gangurde, S.S.; Sudini, H.K.; Fountain, J.C.; Liao, B.; Desmae, H.; Okori, P.; et al. Mitigating aflatoxin contamination in groundnut through a combination of genetic resistance and post-harvest management practices. Toxins 2019, 11, 315. [Google Scholar] [CrossRef]
- Nigam, S.; Waliyar, F.; Aruna, R.; Reddy, S.; Kumar, P.L.; Craufurd, P.Q.; Diallo, A.T.; Ntare, B.R.; Upadhyaya, H.D. Breeding peanut for resistance to aflatoxin contamination at ICRISAT. Peanut Sci. 2009, 36, 42–49. [Google Scholar] [CrossRef]
- Arias, R.S.; Dang, P.M.; Sobolev, V.S. RNAi-mediated control of aflatoxins in peanut: Method to analyze mycotoxin production and transgene expression in the peanut/Aspergillus pathosystem. J. Vis. Exp. 2015, 106, e53398. [Google Scholar] [CrossRef]
- Sharma, K.K.; Pothana, A.; Prasad, K.; Shah, D.; Kaur, J.; Bhatnagar, D.; Chen, Z.Y.; Raruang, Y.; Cary, J.W.; Rajasekaran, K.; et al. Peanuts that keep aflatoxin at bay: A threshold that matters. Plant Biotechnol. J. 2018, 16, 1024–1033. [Google Scholar] [CrossRef]
- Thakare, D.; Zhang, J.; Wing, R.A.; Cotty, P.J.; Schmidt, M.A. Aflatoxin-free transgenic maize using host-induced gene silencing. Sci. Adv. 2017, 3, e1602382. [Google Scholar] [CrossRef]
- Caceres, I.; Al Khoury, A.; El Khoury, R.; Lorber, S.P.; Oswald, I.; El Khoury, A.; Atoui, A.; Puel, O.; Bailly, J.D. Aflatoxin biosynthesis and genetic regulation: A review. Toxins 2020, 12, 150. [Google Scholar] [CrossRef]
- Khan, R.; Ghazali, F.M.; Mahyudin, N.A.; Samsudin, N.I.P. Aflatoxin biosynthesis, genetic regulation, toxicity, and control strategies: A review. J. Fungi 2021, 7, 606. [Google Scholar] [CrossRef] [PubMed]
- Masanga, J.O.; Matheka, J.M.; Omer, R.A.; Ommeh, S.C.; Monda, E.O.; Alakonya, A.E. Downregulation of transcription factor aflR in Aspergillus flavus confers reduction to aflatoxin accumulation in transgenic maize with alteration of host plant architecture. Plant Cell Rep. 2015, 34, 1379–1387. [Google Scholar] [CrossRef] [PubMed]
- Raruang, Y.; Omolehin, O.; Hu, D.; Wei, Q.; Han, Z.-Q.; Rajasekaran, K.; Cary, J.W.; Wang, K.; Chen, Z.Y. Host induced gene silencing targeting Aspergillus flavus aflM reduced aflatoxin contamination in transgenic maize under field conditions. Front. Microbiol. 2020, 11, 754. [Google Scholar] [CrossRef]
- Power, I.L.; Faustinelli, P.C.; Orner, V.A.; Sobolev, V.S.; Arias, R.S. Analysis of small RNA populations generated in peanut leaves after exogenous application of dsRNA and dsDNA targeting aflatoxin synthesis genes. Sci. Rep. 2020, 10, 13820. [Google Scholar] [CrossRef] [PubMed]
- Omolehin, O.; Raruang, Y.; Hu, D.; Han, Z.-Q.; Wei, Q.; Wang, K.; Rajasekaran, K.; Cary, J.W.; Chen, Z.Y. Resistance to aflatoxin accumulation in maize mediated by host-induced silencing of the Aspergillus flavus alkaline protease (alk) gene. J. Fungi 2021, 7, 904. [Google Scholar] [CrossRef]
- Gilbert, M.K.; Majumdar, R.; Rajasekaran, K.; Chen, Z.-Y.; Wei, Q.; Sickler, C.M.; Lebar, M.D.; Cary, J.W.; Frame, B.R.; Wang, K. RNA interference-based silencing of the alpha-amylase (amy1) gene in Aspergillus flavus decreases fungal growth and aflatoxin production in maize kernels. Planta 2018, 247, 1465–1473. [Google Scholar] [CrossRef]
- Cary, J.W.; Harris-Coward, P.Y.; Ehrlich, K.C.; Mack, B.M.; Kale, S.P.; Larey, C.; Calvo, A.M. NsdC and NsdD affect Aspergillus flavus morphogenesis and aflatoxin production. Eukaryot. Cell 2012, 11, 1104–1111. [Google Scholar] [CrossRef]
- Calvo, A.M.; Bok, J.; Brooks, W.; Keller, N.P. veA is required for toxin and sclerotial production in Aspergillus parasiticus. Appl. Environ. Microbiol. 2004, 70, 4733–4739. [Google Scholar] [CrossRef] [PubMed]
- Price, M.S.; Yu, J.; Nierman, W.C.; Kim, H.S.; Pritchard, B.; Jacobus, C.A.; Bhatnagar, D.; Cleveland, T.E.; Payne, G.A. The aflatoxin pathway regulator AflR induces gene transcription inside and outside of the aflatoxin biosynthetic cluster. FEMS Microbiol. Lett. 2006, 255, 275–279. [Google Scholar] [CrossRef]
- Yu, J. Current understanding on aflatoxin biosynthesis and future perspective in reducing aflatoxin contamination. Toxins 2012, 4, 1024–1057. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, F.; Senthil-Kumar, M.; Dai, X.; Ramu, V.S.; Lee, S.; Mysore, K.S.; Zhao, P.X. pssRNAit: A web server for designing effective and specific plant siRNAs with genome-wide off-target assessment. Plant Physiol. 2020, 184, 65–81. [Google Scholar] [CrossRef] [PubMed]
- Koch, A.; Wassenegger, M. Host-induced gene silencing–mechanisms and applications. New Phytol. 2021, 231, 54–59. [Google Scholar] [CrossRef]
- Nunes, C.C.; Dean, R.A. Host-induced gene silencing: A tool for understanding fungal host interaction and for developing novel disease control strategies. Mol. Plant Pathol. 2012, 13, 519–529. [Google Scholar] [CrossRef]
- Qi, T.; Guo, J.; Peng, H.; Liu, P.; Kang, Z.; Guo, J. Host-induced gene silencing: A powerful strategy to control diseases of wheat and barley. Int. J. Mol. Sci. 2019, 20, 206. [Google Scholar] [CrossRef] [PubMed]
- Baidya, S.; Duran, R.M.; Lohmar, J.M.; Harris-Coward, P.Y.; Cary, J.W.; Hong, S.Y.; Roze, L.V.; Linz, J.E.; Calvo, A.M. VeA is associated with the response to oxidative stress in the aflatoxin producer Aspergillus flavus. Eukaryot. Cell 2014, 13, 1095–1103. [Google Scholar] [CrossRef]
- Cary, J.; Ehrlich, K.; Wright, M.; Chang, P.-K.; Bhatnagar, D. Generation of aflR disruption mutants of Aspergillus parasiticus. Appl. Microbiol. Biotechnol. 2000, 53, 680–684. [Google Scholar] [CrossRef]
- Payne, G.; Nystrom, G.; Bhatnagar, D.; Cleveland, T.; Woloshuk, C. Cloning of the afl-2 gene involved in aflatoxin biosynthesis from Aspergillus flavus. Appl. Environ. Microbiol. 1993, 59, 156–162. [Google Scholar] [CrossRef]
- Chang, P.-K.; Scharfenstein, L.L.; Li, P.; Ehrlich, K.C. Aspergillus flavus VelB acts distinctly from VeA in conidiation and may coordinate with FluG to modulate sclerotial production. Fungal Genet. Biol. 2013, 58, 71–79. [Google Scholar] [CrossRef]
- Duran, R.M.; Cary, J.W.; Calvo, A.M. Production of cyclopiazonic acid, aflatrem, and aflatoxin by Aspergillus flavus is regulated by veA, a gene necessary for sclerotial formation. Appl. Microbiol. Biotechnol. 2007, 73, 1158–1168. [Google Scholar] [CrossRef]
- Power, I.L.; Dang, P.M.; Sobolev, V.S.; Orner, V.A.; Powell, J.L.; Lamb, M.C.; Arias, R.S. Characterization of small RNA populations in non-transgenic and aflatoxin-reducing-transformed peanut. Plant Sci. 2017, 257, 106–125. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Hadi, A.M.; Caley, D.P.; Carter, D.R.; Magan, N. Control of aflatoxin production of Aspergillus flavus and Aspergillus parasiticus using RNA silencing technology by targeting aflD (nor-1) gene. Toxins 2011, 3, 647–659. [Google Scholar] [CrossRef] [PubMed]
- McDonald, T.; Brown, D.; Keller, N.P.; Hammond, T.M. RNA silencing of mycotoxin production in Aspergillus and Fusarium species. Mol. Plant Microbe Int. 2005, 18, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Freeman, B.; Beattie, G. An overview of plant defenses against pathogens and herbivores. Plant Health Instr. 2008. [Google Scholar] [CrossRef]
- Görlach, A.; Bertram, K.; Hudecova, S.; Krizanova, O. Calcium and ROS: A mutual interplay. Redox Biol. 2015, 6, 260–271. [Google Scholar] [CrossRef]
- Millana Fañanás, E.; Todesca, S.; Sicorello, A.; Masino, L.; Pompach, P.; Magnani, F.; Pastore, A.; Mattevi, A. On the mechanism of calcium-dependent activation of NADPH oxidase 5 (NOX5). FEBS J. 2020, 287, 2486–2503. [Google Scholar] [CrossRef]
- Haslbeck, M.; Vierling, E. A first line of stress defense: Small heat shock proteins and their function in protein homeostasis. J. Mol. Biol. 2015, 427, 1537–1548. [Google Scholar] [CrossRef]
- Van Ooijen, G.; Lukasik, E.; Van Den Burg, H.A.; Vossen, J.H.; Cornelissen, B.J.; Takken, F.L. The small heat shock protein 20 RSI2 interacts with and is required for stability and function of tomato resistance protein I-2. Plant J. 2010, 63, 563–572. [Google Scholar] [CrossRef]
- Banzet, N.; Richaud, C.; Deveaux, Y.; Kazmaier, M.; Gagnon, J.; Triantaphylidès, C. Accumulation of small heat shock proteins, including mitochondrial HSP22, induced by oxidative stress and adaptive response in tomato cells. Plant J. 1998, 13, 519–527. [Google Scholar] [CrossRef]
- Liu, X.; Khajuria, C.; Li, J.; Trick, H.N.; Huang, L.; Gill, B.S.; Reeck, G.R.; Antony, G.; White, F.F.; Chen, M.-S. Wheat Mds-1 encodes a heat-shock protein and governs susceptibility towards the Hessian fly gall midge. Nat. Commun. 2013, 4, 2070. [Google Scholar] [CrossRef]
- Bhatnagar-Mathur, P.; Yogendra, K.; Parankusam, S.; Sanivarapu, H.; Prasad, K.; Lingampali, S.B.; Sharma, K.K. Comparative proteomics provide insights on the basis of resistance to Aspergillus flavus infection and aflatoxin production in peanut (Arachis hypogea L.). J. Plant Interact. 2021, 16, 494–509. [Google Scholar] [CrossRef]
- Dodds, P.N.; Rathjen, J.P. Plant immunity: Towards an integrated view of plant–pathogen interactions. Nat. Rev. Genet. 2010, 11, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Kushalappa, A.C.; Yogendra, K.N.; Karre, S. Plant innate immune response: Qualitative and quantitative resistance. Crit. Rev. Plant Sci. 2016, 35, 38–55. [Google Scholar] [CrossRef]
- Pieterse, C.M.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C. Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef]
- Clevenger, J.; Marasigan, K.; Liakos, V.; Sobolev, V.; Vellidis, G.; Holbrook, C.; Ozias-Akins, P. RNA sequencing of contaminated seeds reveals the state of the seed permissive for pre-harvest aflatoxin contamination and points to a potential susceptibility factor. Toxins 2016, 8, 317. [Google Scholar] [CrossRef]
- Fountain, J.C.; Khera, P.; Yang, L.; Nayak, S.N.; Scully, B.T.; Lee, R.D.; Chen, Z.Y.; Kemerait, R.C.; Varshney, R.K.; Guo, B. Resistance to Aspergillus flavus in maize and peanut: Molecular biology, breeding, environmental stress, and future perspectives. Crop J. 2015, 3, 229–237. [Google Scholar] [CrossRef]
- Raffaele, S.; Vailleau, F.; Léger, A.; Joubès, J.; Miersch, O.; Huard, C.; Blée, E.; Mongrand, S.; Domergue, F.; Roby, D. A MYB transcription factor regulates very-long-chain fatty acid biosynthesis for activation of the hypersensitive cell death response in Arabidopsis. Plant Cell 2008, 20, 752–767. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Zhang, C.L.; Wang, G.L.; Wang, Y.X.; Qi, C.H.; Zhao, Q.; You, C.X.; Li, Y.Y.; Hao, Y.J. The R2R3 MYB transcription factor MdMYB30 modulates plant resistance against pathogens by regulating cuticular wax biosynthesis. BMC Plant Biol. 2019, 19, 362. [Google Scholar] [CrossRef]
- Song, G.; Li, X.; Munir, R.; Khan, A.R.; Azhar, W.; Yasin, M.U.; Jiang, Q.; Bancroft, I.; Gan, Y. The WRKY6 transcription factor affects seed oil accumulation and alters fatty acid compositions in Arabidopsis thaliana. Physiol. Plant. 2020, 169, 612–624. [Google Scholar] [CrossRef]
- Choi, C.; Hwang, S.H.; Fang, I.R.; Kwon, S.I.; Park, S.R.; Ahn, I.; Kim, J.B.; Hwang, D.J. Molecular characterization of Oryza sativa WRKY 6, which binds to W-box-like element 1 of the Oryza sativa pathogenesis-related (PR) 10a promoter and confers reduced susceptibility to pathogens. New Phytol. 1015, 208, 846–859. [Google Scholar] [CrossRef]
- Jayashree, T.; Subramanyam, C. Oxidative stress as a prerequisite for aflatoxin production by Aspergillus parasiticus. Free Radic. Biol. Med. 2000, 29, 981–985. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Kolomiets, M.V. Host-derived lipids and oxylipins are crucial signals in modulating mycotoxin production by fungi. Toxin Rev. 2009, 28, 79–88. [Google Scholar] [CrossRef]
- Laxalt, A.M.; Munnik, T. Phospholipid signalling in plant defence. Curr. Opin. Plant Biol. 2002, 5, 332–338. [Google Scholar] [CrossRef]
- Munnik, T.; Testerink, C. Plant phospholipid signaling: “In a nutshell”. J. Lipid Res. 2009, 50, S260–S265. [Google Scholar] [CrossRef]
- de Torres Zabela, M.; Fernandez-Delmond, I.; Niittyla, T.; Sanchez, P.; Grant, M. Differential expression of genes encoding Arabidopsis phospholipases after challenge with virulent or avirulent Pseudomonas isolates. Mol. Plant Microbe Interact. 2002, 15, 808–816. [Google Scholar] [CrossRef]
- Canavar, Ö.; Kaynak, M.A. Prevention of pre-harvest aflatoxin production and the effect of different harvest times on peanut (Arachis hypogaea L.) fatty acids. Food Addit. Contam. Part A 2013, 30, 1807–1818. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ruiz, H.; Szurek, B.; Van den Ackerveken, G. Stop helping pathogens: Engineering plant susceptibility genes for durable resistance. Curr. Opin. Biotechnol. 2021, 70, 187–195. [Google Scholar] [CrossRef]
- Yang, C.F.; Tsai, W.C. Calmodulin: The switch button of calcium signaling. Tzu Chi Med. J. 2002, 34, 15–22. [Google Scholar] [CrossRef]
- Zhang, J.; Zou, A.; Wen, Y.; Wei, X.; Liu, C.; Lv, X.; Ma, X.; Fan, G.; Sun, X. SlCML55, a novel Solanum lycopersicum calmodulin-like gene, negatively regulates plant immunity to Phytophthora pathogens. Sci. Hortic. 2022, 299, 111049. [Google Scholar] [CrossRef]
- Long, Q.; Xie, Y.; He, Y.; Li, Q.; Zou, X.; Chen, S. Abscisic acid promotes jasmonic acid accumulation and plays a key role in citrus canker development. Front. Plant Sci. 2019, 10, 1634. [Google Scholar] [CrossRef]
- Li, X.; Liu, Y.; He, Q.; Li, S.; Liu, W.; Lin, C.; Miao, W. A candidate secreted effector protein of rubber tree powdery mildew fungus contributes to infection by regulating plant ABA biosynthesis. Front. Microbiol. 2020, 11, 591387. [Google Scholar] [CrossRef] [PubMed]
- Ulferts, S.; Delventhal, R.; Splivallo, R.; Karlovsky, P.; Schaffrath, U. Abscisic acid negatively interferes with basal defence of barley against Magnaporthe oryzae. BMC Plant Biol. 2015, 15, 7. [Google Scholar] [CrossRef] [PubMed]
- Kusch, S.; Panstruga, R. mlo-based resistance: An apparently universal “weapon” to defeat powdery mildew disease. Mol. Plant Microbe Interact. 2017, 30, 179–189. [Google Scholar] [CrossRef]
- Rescher, U.; Gerke, V. Annexins-unique membrane binding proteins with diverse functions. J. Cell Sci. 2004, 117, 2631–2639. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Schipper, D.; Jacobsen, E.; Visser, R.G.; Govers, F.; Bouwmeester, K.; Bai, Y. Silencing susceptibility genes in potato hinders primary infection with Phytophthora infestans at different stages. Hortic. Res. 2022, 9, uhab058. [Google Scholar] [CrossRef]
- He, X.; Huo, Y.; Liu, X.; Zhou, Q.; Feng, S.; Shen, X.; Li, B.; Wu, S.; Chen, X. Activation of disease resistance against Botryosphaeria dothidea by downregulating the expression of MdSYP121 in apple. Hortic. Res. 2018, 5, 24. [Google Scholar] [CrossRef] [PubMed]
- Nierman, W.C.; Yu, J.; Fedorova-Abrams, N.D.; Losada, L.; Cleveland, T.E.; Bhatnagar, D.; Bennett, J.W.; Dean, R.; Payne, G.A. Genome sequence of Aspergillus flavus NRRL 3357, a strain that causes aflatoxin contamination of food and feed. Genome Announc. 2015, 3, e00168-15. [Google Scholar] [CrossRef]
- Sharma, K.K.; Anjaiah, V. An efficient method for the production of transgenic plants of peanut (Arachis hypogaea L.) through Agrobacterium tumefaciens-mediated genetic transformation. Plant Sci. 2000, 159, 7–19. [Google Scholar] [CrossRef]
- Sharma, K.K.; Bhatnagar-Mathur, P. Peanut (Arachis hypogaea L.). In Methods in Molecular Biology Vol. 343: Agrobacterium Protocols, 2/e; Wang, K., Ed.; Humana Press Inc.: Totowa, NJ, USA, 2006; Volume 1, pp. 347–358. [Google Scholar]
- Reddy, D.S.; Bhatnagar-Mathur, P.; Cindhuri, K.S.; Sharma, K.K. Evaluation and validation of reference genes for normalization of quantitative real-time PCR based gene expression studies in peanut. PLoS ONE 2013, 8, e78555. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Weng, H.; Pan, A.; Yang, L.; Zhang, C.; Liu, Z.; Zhang, D. Estimating number of transgene copies in transgenic rapeseed by real-time PCR assay with HMG I/Y as an endogenous reference gene. Plant Mol. Biol. Rep. 2004, 22, 289–300. [Google Scholar] [CrossRef]
- Mehan, V.; Ramakrishna, N.; Rao, R.; McDonald, D. Preharvest aflatoxin contamination of groundnuts subjected to terminal drought stress in postrainy season. Mycotoxin Res. 1995, 11, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.; Reddy, C.; Muralidharan, K. Detection of Aspergillus spp. and aflatoxin B1 in rice in India. Food Microbiol. 2009, 26, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Karre, S.; Dhokane, D.; Kage, U.; Hukkeri, S.; Kushalappa, A.C. Real-time quantitative PCR based method for the quantification of fungal biomass to discriminate quantitative resistance in barley and wheat genotypes to fusarium head blight. J. Cereal Sci. 2015, 64, 16–22. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. B Stat. Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Chou, K.-C.; Shen, H.-B. Plant-mPLoc: A top-down strategy to augment the power for predicting plant protein subcellular localization. PLoS ONE 2010, 5, e11335. [Google Scholar] [CrossRef]
- Okuda, S.; Watanabe, Y.; Moriya, Y.; Kawano, S.; Yamamoto, T.; Matsumoto, M.; Takami, T.; Kobayashi, D.; Araki, N.; Yoshizawa, A.C.; et al. jPOSTrepo: An international standard data repository for proteomes. Nucleic Acids Res. 2017, 45, D1107–D1111. [Google Scholar] [CrossRef]
- Lohse, M.; Nagel, A.; Herter, T.; May, P.; Schroda, M.; Zrenner, R.; Tohge, T.; Fernie, A.R.; Stitt, M.; Usadel, B. Mercator: A fast and simple web server for genome scale functional annotation of plant sequence data. Plant Cell Environ. 2014, 37, 1250–1258. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Mass (kDa) | Protein Name | 0 h | 30 h | 48 h | 72 h |
|---|---|---|---|---|---|---|
| A. Fungal differentiation, development, and pathogenicity | ||||||
| P60204; P60205 | 17.01 | Calmodulin | −46.44 | |||
| A0A1R3RGK4 | 27.39 | Ochratoxin biosynthesis cluster transcription factor | 19.16 | |||
| A2QK82 | 34.97 | Probable pectinesterase A | 77.76 | |||
| Q2UNJ0 | 37.39 | Chitin synthase export chaperone | −14.57 | |||
| Q5B8Y3; Q2UQ34 | 37.54 | Eukaryotic translation initiation factor 3 subunit I | −35.84 | |||
| Q1XGE2; Q8TFU8 | 37.79 | Transcriptional activator hacA | −23.18 | |||
| Q5ATQ3 | 39.87 | Endopolygalacturonase AN8327 | −26.66 | |||
| Q5AQJ1 | 39.94 | Probable pectin lyase D | 21.05 | |||
| C8VDI2 | 40.83 | Autophagy-related protein 3 | 1.95 | 13.92 | ||
| A1CLZ1 | 41.63 | Diels-Alderase ccsF | 13.77 | |||
| C8VQG9; Q6TLK5 | 43.31 | Secondary metabolism regulator laeA | −10.42 | |||
| Q2U6D5 | 45.27 | Autophagy-related protein 18 | 11.70 | |||
| A2QMH1 | 55.14 | Kynurenine 3-monooxygenase 2 | −11.03 | |||
| Q12730 | 56.63 | Protein disulfide-isomerase | −10.25 | |||
| Q5B9G5 | 56.87 | Mannitol 2-dehydrogenase | 72.84 | |||
| B0Y7U1; Q4WMR0 | 58.54 | Probable feruloyl esterase B-2 | −15.05 | |||
| P28298 | 60.49 | Isocitrate lyase | 2.21 | 37.51 | ||
| A1DG37 | 66.19 | Autophagy-related protein 22-1 | −22.57 | 22.77 | ||
| A1CEH4 | 69.49 | Vacuolar fusion protein mon1 | 83.61 | |||
| Q12062 | 70.39 | Versicolorin B synthase | −30.71 | |||
| Q9HFB3; Q96UW0 | 71.31 | pH-response transcription factor pacC/RIM101 | 10.99 | −16.96 | ||
| Q2UB56 | 80.15 | Sorting nexin mvp1 | −12.61 | |||
| B8NBX4 | 84.06 | Cell pattern formation-associated protein stuA | −12.90 | |||
| P20945 | 89.34 | Conidiophore development regulator abaA | −2.64 | −11.04 | ||
| Q9R1S8 | 93.36 | Calpain-7 | 12.10 | |||
| Q92197 | 101.78 | Chitin synthase C | 20.18 | |||
| Q4WPF2 | 106.42 | Serine/threonine-protein kinase atg1 | −1.57 | −12.29 | ||
| Q00078 | 123.26 | Protein kinase C-like | −4.39 | −17.63 | ||
| B. Aflatoxin biosynthetic pathway | ||||||
| Q8TGA1 | 21.22 | Fatty acid synthase beta subunit (aflB) | 2.15 | |||
| P50161 | 28.15 | Versicolorin reductase 1 (aflM) | −4.26 | |||
| B9WYE6 | 38.87 | Versiconal hemiacetal acetate reductase (vrdA) | −1.90 | |||
| B8NUL8; Q2U4H2 | 46.30 | Lipoyl synthase_ mitochondrial (aflA) | −1.67 | −3.75 | ||
| O42716 | 47.63 | Aflatoxin cluster transcriptional coactivator (aflS) | −5.71 | |||
| P52957 | 47.25 | Sterigmatocystin biosynthesis regulatory protein (aflR) | −2.01 | |||
| P0C1B3; P30292 | 55.32 | Alpha-amylase A type-1/2 (amy1) | 2.82 | |||
| Q6UEF3 | 55.48 | FAD-binding monooxygenase (aflW) | −1.75 | −9.20 | ||
| Q6UEG2 | 55.65 | Cytochrome P450 monooxygenase (aflN) | 2.74 | |||
| Q6UEF1 | 56.06 | Oxidoreductase (AflY) | −2.50 | |||
| Q6UEH4 | 56.23 | Cytochrome P450 monooxygenase (aflU) | −8.42 | |||
| Q9UW95 | 56.73 | Versicolorin B desaturase (aflL) | −3.79 | |||
| Q6UEF4 | 56.73 | Cytochrome P450 monooxygenase (aflV) | −3.92 | |||
| Q5BBM1 | 57.79 | Sexual development regulator (velC) | −3.47 | |||
| O13345 | 60.47 | O-methyl sterigmatocystin oxidoreductase (aflQ) | −3.46 | |||
| E9RCK4 | 63.20 | Developmental and secondary metabolism regulator (veA) | −2.21 | −2.44 | ||
| Q12062 | 70.38 | Versicolorin B synthase (AflK) | −30.71 | |||
| Q8TGA1 | 212.26 | Fatty acid synthase beta subunit (aflB) | −1.71 | |||
| Q12053 | 232.94 | Norsolorinic acid synthase (aflC) | −7.06 | |||
| Accession | Mass | Description | Fold Change against WT Control | |||
|---|---|---|---|---|---|---|
| 0 h | 30 h | 48 h | 72 h | |||
| Heat shock proteins and calcium signaling-related proteins | ||||||
| B4UW51 | 14.52 | Class II small heat shock protein Le-HSP17.6 | 2.50 | |||
| B4UW89 | 18.10 | Heat shock protein 2 | 1.97 | 2.50 | ||
| E3NYT2 | 19.04 | Heat shock protein 70 | 3.62 | |||
| A0A068VVA2 | 24.85 | Ca2+ hinding-protein SOS3 | 2.14 | 2.27 | ||
| E7CQA1 | 40.70 | Heat shock transcription factor A-2 | 2.10 | |||
| V5M2Y8 | 61.53 | Calcium-dependent protein kinase | 2.28 | 5.94 | ||
| Phytohormones | ||||||
| Q5QET3 | 8.39 | Auxin-induced putative CP12 domain-containing protein | 2.40 | |||
| Q5QET8 | 9.69 | Auxin-induced putative aldo/keto reductase family protein | 4.44 | |||
| B4UW77 | 12.57 | Gibberellin-regulated protein | 1.61 | 1.94 | ||
| E3NYH5 | 14.39 | S-adenosyl methionine synthase | 2.00 | 3.04 | ||
| M4TG02 | 30.48 | Auxin signaling F-box 3 | 3.38 | |||
| A0A023IUN1 | 33.82 | Abscisic acid 8-hydroxylase 3 | 8.95 | |||
| D7RJM3 | 39.89 | S-adenosylmethionine decarboxylase proenzyme | 2.02 | |||
| K0FB33 | 45.18 | Ethylene-responsive element binding factor 6 | 2.03 | |||
| G4X5C7 | 48.26 | ABA response element binding protein 1 | 2.86 | 12.57 | ||
| U6NJF1; K4PM24 | 55.28 | ABA 8′-hydroxylase | 4.35 | 3.19 | ||
| Transcription factors | ||||||
| V5T7X7 | 8.47 | Putative MYB-related protein 25 | 5.33 | 1.86 | ||
| M4SZY9 | 22.35 | Ethylene-responsive transcription factor | 1.64 | 2.03 | 66.16 | |
| V5T7W6 | 22.51 | Putative R2R3 MYB protein 8 | 2.10 | |||
| A0A0H3Y991 | 24.02 | Wuschel-related homeobox 13B1 | 3.00 | |||
| A0A0H3Y7V8 | 24.25 | Wuschel-related homeobox 13A | 1.78 | |||
| E4W7V3 | 24.80 | Putative DREB transcription factor | 1.91 | 9.65 | ||
| M4SZZ4 | 24.80 | Nuclear transcription factor Y subunit A-3 | 2.03 | |||
| M4T2P8 | 25.06 | F-box family protein 6 | 2.55 | |||
| V5T684 | 25.58 | Putative R2R3 MYB protein 9 | 1.55 | 2.40 | ||
| V5T6N4 | 27.20 | Putative R2R3 MYB protein 1 | 4.73 | |||
| A0A1L1VTR5 | 32.84 | MYB-like transcript factor 6 | 3.73 | |||
| V5T7W9 | 33.01 | Putative MYB-related protein 14 | 7.39 | |||
| V5T688 | 33.56 | Putative MYB-related protein 16 | 1.58 | 4.19 | ||
| J9Q9Z8 | 33.74 | Ethylene-responsive element binding factor 3 | 24.17 | 70.87 | ||
| V5T6Q5 | 34.36 | Putative MYB-related protein 28 | 1.91 | |||
| K0FBW3 | 34.41 | Ethylene-responsive element binding factor 4 | 6.26 | 3.14 | ||
| V5T8I2 | 35.73 | Putative R2R3 MYB protein 7 | 1.85 | 2.21 | ||
| V5T714 | 36.32 | Putative MYB-related protein 22 | 2.66 | |||
| C6EU67 | 37.94 | NAC-like transcription factor 3 | 2.20 | 4.28 | ||
| V5T7Y1 | 38.12 | Putative MYB-related protein 30 | 1.65 | 4.50 | ||
| C6EU68 | 39.37 | NAC-like transcription factor | 1.67 | 2.62 | 4.04 | |
| B5AK53 | 39.67 | WRKY transcription factor 15 | 12.15 | |||
| V5T8I4 | 48.21 | Putative MYB-related protein 13 | 1.60 | 1.99 | ||
| Fatty acids | ||||||
| D3YM77 | 14.28 | Acyl carrier protein | 3.92 | 29.37 | ||
| A0A0A6ZDY1 | 15.56 | Peptidyl-prolyl cis-trans isomerase | 2.56 | 2.96 | ||
| N1NKF7 | 15.99 | Glycerol-3-phosphate dehydrogenase | 22.47 | |||
| A0A0A6ZDP1 | 20.10 | Glyceraldehyde-3-phosphate dehydrogenase C2 | 2.04 | 3.26 | ||
| B4UW57 | 24.67 | Putative dihydroflavonol reductase | 1.67 | |||
| B4UW49 | 26.61 | Putative lipase | 1.58 | |||
| B4UWB9 | 26.79 | Lipoxygenase 1 | 7.97 | |||
| A0A0U3E0B1 | 27.41 | Phosphatidyl inositol phosphate kinase | 1.98 | 4.68 | ||
| D8KXY5 | 41.11 | Malonyl-CoA:ACP transacylase 1-1 | 3.86 | |||
| A0A384QWC2 | 43.91 | Acyl-[acyl-carrier-protein] desaturase | 2.88 | 1.86 | 6.09 | |
| A0A3G0YUC8 | 44.36 | Lysophosphatidyl acyltransferase 5 | 5.40 | 3.90 | 1.82 | |
| A0A384QZQ3 | 45.64 | Palmitoyl-monogalactosyldiacylglycerol delta-7 desaturase | 1.97 | 6.01 | ||
| N1NG06 | 48.15 | 3-ketoacyl-CoA synthase | 1.67 | 3.48 | ||
| N1NFY7 | 48.58 | Putative ceramide kinase-related protein | 3.18 | |||
| A0A0R4UXQ1 | 48.86 | 3-ketoacyl-CoA thiolase | 4.58 | |||
| A0A0R4VXV1 | 50.46 | Phosphatidylserine decarboxylase proenzyme 1 | 3.59 | |||
| E6Y9A7 | 50.47 | Beta-ketoacyl-ACP synthetase I | 1.67 | 5.86 | ||
| A0A384QZP9 | 52.49 | Sphingolipid delta8 desaturase | 2.29 | 2.22 | 2.39 | 9.71 |
| A0A0K0K9Q6 | 53.75 | Glyceraldehyde-3-phosphate dehydrogenase | 1.82 | 3.87 | 3.85 | |
| A0A0R4UXP7 | 56.05 | Glycerol-3-phosphate acyltransferase 6 | 3.25 | 2.53 | ||
| A0A385I5T0 | 57.54 | 3-ketoacyl-CoA synthase | 2.36 | |||
| A0A109Z9U2 | 75.55 | Long chain acyl-CoA synthetase 1 | 1.51 | 2.02 | ||
| F1AM70 | 79.43 | Triacylglycerol lipase 1 | 1.65 | |||
| A0A0R4VUF1 | 91.12 | Digalactosyldiacylglycerol synthase 1 | 2.61 | |||
| Q2HWT7 | 91.27 | Phospholipase D | 2.63 | 4.15 | ||
| Q4JME6 | 97.76 | Lipoxygenase | 1.63 | 9.88 | 3.43 | 3.36 |
| Accession | Mass | Description | Change against WT at Different Times after Infection (In Folds) | |||
|---|---|---|---|---|---|---|
| 0 h | 30 h | 48 h | 72 h | |||
| E2DQY9 | 8.92 | Heat shock protein DnaJ | −2.57 | −3.69 | ||
| B4UW90 | 12.43 | Heat shock protein 3 | −1.58 | |||
| B1PMD1 | 13.50 | Zinc finger protein ZFP133 | −1.87 | |||
| Q6R2U6 | 16.64 | Calmodulin | −3.33 | |||
| Q06H39 | 17.10 | Syntaxin | −2.05 | −1.93 | −4.02 | |
| B4UWB2 | 22.09 | Kunitz trypsin inhibitor 4 | −1.64 | |||
| B4UW91 | 25.92 | Putative heat shock protein 4 | −1.53 | −11.67 | −3.21 | |
| V5T7W3 | 27.81 | Putative R2R3 MYB protein 3 | −1.63 | −2.60 | ||
| E3NYG8 | 28.02 | Zinc finger protein, ZAT10-like | −3.51 | |||
| V5T6P8 | 28.83 | Putative MYB-related protein 18 | −1.85 | −2.49 | −1.71 | |
| B5AK52 | 30.03 | NAC-like transcription factor 2 | −5.56 | −4.30 | ||
| V5T692 | 32.43 | Putative MYB-related protein 21 | −1.51 | −5.59 | ||
| V5T8J4 | 32.74 | Putative MYB-related protein 29 | −1.83 | −2.05 | ||
| B2ZHY3 | 34.34 | NAC-like transcription factor | −1.83 | −1.68 | ||
| A0A0F6VX63 | 36.25 | Annexin | −6.26 | |||
| V5T7X2 | 37.51 | Putative MYB-related protein 20 | −3.00 | −2.97 | ||
| K0FAV2 | 41.50 | Ethylene-responsive element binding factor 1 | −2.31 | −2.70 | −2.52 | |
| D8KXZ7 | 41.53 | Enoyl-ACP reductase 1–2 | −2.00 | −6.41 | ||
| E6Y9A8 | 52.10 | Chloroplast omega-6 fatty acid desaturase | −2.96 | −1.63 | −1.95 | |
| A0A290G010 | 56.63 | MLO-like protein | −1.60 | −3.82 | ||
| A0A385I5T0 | 57.54 | 3-ketoacyl-CoA synthase | −2.16 | |||
| B4YA12 | 58.21 | Calcium calmodulin-dependent protein kinase | −2.28 | |||
| D8KXZ0 | 59.54 | Beta-ketoacyl-ACP synthase II-1 | −41.34 | |||
| Q70KY0 | 67.36 | 9-cis-epoxy carotenoid dioxygenase | −1.89 | −7.53 | ||
| A0A109QJM5 | 75.06 | Long chain acyl-CoA synthetase 4 | −12.32 | |||
| N1NFY2 | 105.41 | Putative Zinc finger_ C3HC4 type (RING finger) | −7.70 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prasad, K.; Yogendra, K.; Sanivarapu, H.; Rajasekaran, K.; Cary, J.W.; Sharma, K.K.; Bhatnagar-Mathur, P. Multiplexed Host-Induced Gene Silencing of Aspergillus flavus Genes Confers Aflatoxin Resistance in Groundnut. Toxins 2023, 15, 319. https://doi.org/10.3390/toxins15050319
Prasad K, Yogendra K, Sanivarapu H, Rajasekaran K, Cary JW, Sharma KK, Bhatnagar-Mathur P. Multiplexed Host-Induced Gene Silencing of Aspergillus flavus Genes Confers Aflatoxin Resistance in Groundnut. Toxins. 2023; 15(5):319. https://doi.org/10.3390/toxins15050319
Chicago/Turabian StylePrasad, Kalyani, Kalenahalli Yogendra, Hemalatha Sanivarapu, Kanniah Rajasekaran, Jeffrey W. Cary, Kiran K. Sharma, and Pooja Bhatnagar-Mathur. 2023. "Multiplexed Host-Induced Gene Silencing of Aspergillus flavus Genes Confers Aflatoxin Resistance in Groundnut" Toxins 15, no. 5: 319. https://doi.org/10.3390/toxins15050319
APA StylePrasad, K., Yogendra, K., Sanivarapu, H., Rajasekaran, K., Cary, J. W., Sharma, K. K., & Bhatnagar-Mathur, P. (2023). Multiplexed Host-Induced Gene Silencing of Aspergillus flavus Genes Confers Aflatoxin Resistance in Groundnut. Toxins, 15(5), 319. https://doi.org/10.3390/toxins15050319