Practical Strategies to Reduce Ochratoxin A in Foods


Abstract
:1. Introduction
2. Ochratoxin A (OTA)
2.1. Chemistry
2.2. Toxicology and Toxicity Mechanism
2.3. Natural Occurrence of OTA
2.4. Risk Assessment and Regulations
3. Effects of Food Processing
3.1. Non-Thermal Food Processing
3.1.1. Physical Separation and Cleaning
3.1.2. Milling
3.1.3. Fermentation/Brewing
3.2. Thermal Food Processing
3.2.1. Roasting
3.2.2. Coffee Brewing
3.2.3. Extrusion
3.2.4. Other Thermal Food Processing
3.3. Effects of Additives
3.3.1. Baking Soda
3.3.2. Sugars
3.3.3. Salts (NaCl)
3.3.4. Organic Acids
3.3.5. Adsorbents or Binders
3.3.6. Combination of Additives
3.4. Thermal Degradation Products of OTA
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- NTP, N.T.P. Toxicology and carcinogenesis studies of ochratoxin A (CAS No. 303-47-9) in F344/N Rats (Gavage studies). Natl. Toxicol. Program Tech. Rep. Ser. 1989, 358, 1–146. [Google Scholar]
- IARC. Some naturally occurring substances: Food items and constituents, heterocyclic aromatic amines and mycotoxins. IARC Monogr. Eval. Carcinog. Risk Chem. Hum. 1993, 56, 599. [Google Scholar]
- IARC. Some Traditional Herbal Medicines, Some Mycotoxins, Naphthalene and Styrene; World Health Organization: Geneva, Switzerland, 2002. [Google Scholar]
- Pfohl-Leszkowicz, A.; Manderville, R.A. Ochratoxin A: An overview on toxicity and carcinogenicity in animals and humans. Mol. Nutr. Food Res. 2007, 51, 61–99. [Google Scholar] [CrossRef] [PubMed]
- El Khoury, A.; Atoui, A. Ochratoxin A: General overview and actual molecular status. Toxins 2010, 2, 461–493. [Google Scholar] [CrossRef] [PubMed]
- Malir, F.; Ostry, V.; Pfohl-Leszkowicz, A.; Malir, J.; Toman, J. Ochratoxin A: 50 years of research. Toxins 2016, 8, 191. [Google Scholar] [CrossRef] [PubMed]
- Kuiper-Goodman, T.; Scott, P. Risk assessment of the mycotoxin ochratoxin A. Biomed. Environ. Sci. BES 1989, 2, 179–248. [Google Scholar] [PubMed]
- Speijers, G.; Van Egmond, H. Worldwide ochratoxin A levels in food and feeds. Colloq.-Inst. Natl. Sante Rech. Med Colloq. Semin. 1993, 231, 85–100. [Google Scholar]
- Bucheli, P.; Taniwaki, M. Research on the origin, and on the impact of post-harvest handling and manufacturing on the presence of ochratoxin A in coffee. Food Addit. Contam. 2002, 19, 655–665. [Google Scholar] [CrossRef]
- Iamanaka, B.; Taniwaki, M.; Menezes, H.; Vicente, E.; Fungaro, M. Incidence of toxigenic fungi and ochratoxin A in dried fruits sold in Brazil. Food Addit. Contam. 2005, 22, 1258–1263. [Google Scholar] [CrossRef]
- Chulze, S.; Magnoli, C.; Dalcero, A. Occurrence of ochratoxin A in wine and ochratoxigenic mycoflora in grapes and dried vine fruits in South America. Int. J. Food Microbiol. 2006, 111, 5–9. [Google Scholar] [CrossRef]
- Trucksess, M.; Scott, P. Mycotoxins in botanicals and dried fruits: A review. Food Addit. Contam. 2008, 25, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, K.; Campos, V.; Blaszkewicz, M.; Vega, M.; Alvarez, A.; Neira, J.; Degen, G.H. Exposure of neonates to ochratoxin A: First biomonitoring results in human milk (Colostrum) from Chile. Mycotoxin Res. 2010, 26, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Ryu, D. Significance of ochratoxin a in breakfast cereals from the united states. J. Agric. Food Chem. 2015, 63, 9404–9409. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.; Ryu, D. Concentration of ochratoxin A in breakfast cereals and snacks consumed in the United States. Food Control 2014, 40, 140–144. [Google Scholar] [CrossRef]
- Cappozzo, J.; Jackson, L.; Lee, H.J.; Zhou, W.; Al-Taher, F.; Zweigenbaum, J.; Ryu, D. Occurrence of Ochratoxin A in Infant Foods in the United States. J. Food Prot. 2017, 80, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, N.J.; Chen, C.; Palumbo, J.D.; Bianchini, A.; Cappozzo, J.; Stratton, J.; Ryu, D.; Wu, F. A risk assessment of dietary Ochratoxin a in the United States. Food Chem. Toxicol. 2017, 100, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Scottt, P.; Kanhere, S.; Lau, B.; Levvis, D.; Hayward, S.; Ryan, J.; Kuiper-Goodman, T. Survey of Canadian human blood plasma for ochratoxin A. Food Addit. Contam. 1998, 15, 555–562. [Google Scholar] [CrossRef]
- Studer-Rohr, I.; Schlatter, J.; Dietrich, D.R. Kinetic parameters and intraindividual fluctuations of ochratoxin A plasma levels in humans. Arch. Toxicol. 2000, 74, 499–510. [Google Scholar] [CrossRef]
- Tola, M.; Kebede, B. Occurrence, importance and control of mycotoxins: A review. Cogent Food Agric. 2016, 2, 1191103. [Google Scholar] [CrossRef]
- Gimeno, A. Micotoxinas, Introducción. XVII Congreso Centroamericano y del Caribe de Avicultura. 2000. Available online: http://www.engormix.com (accessed on 10 March 2008).
- Wang, L.; Hua, X.; Shi, J.; Jing, N.; Ji, T.; Lv, B.; Liu, L.; Chen, Y. Ochratoxin A: Occurrence and recent advances in detoxification. Toxicon 2022, 210, 11–18. [Google Scholar] [CrossRef]
- Chu, F.S. Studies ion ochratoxins. CRC Crit. Rev. Toxicol. 1974, 2, 499–524. [Google Scholar] [CrossRef] [PubMed]
- Galtier, P. Pharmacokinetics of Ochratoxin a in Animals; IARC Scientific Publications: Lyon, France, 1991; pp. 187–200. [Google Scholar]
- Valenta, H. Chromatographic methods for the determination of ochratoxin A in animal and human tissues and fluids. J. Chromatogr. A 1998, 815, 75–92. [Google Scholar] [CrossRef] [PubMed]
- Abrunhosa, L.; Paterson, R.R.; Venâncio, A. Biodegradation of ochratoxin A for food and feed decontamination. Toxins 2010, 2, 1078–1099. [Google Scholar] [CrossRef] [PubMed]
- Dahal, S.; Lee, H.J.; Gu, K.; Ryu, D. Heat Stability of Ochratoxin A in an Aqueous Buffered Model System. J. Food Prot. 2016, 79, 1748–1752. [Google Scholar] [CrossRef] [PubMed]
- Cramer, B.; Osteresch, B.; Munoz, K.A.; Hillmann, H.; Sibrowski, W.; Humpf, H.U. Biomonitoring using dried blood spots: Detection of ochratoxin A and its degradation product 2’R-ochratoxin A in blood from coffee drinkers. Mol. Nutr. Food Res. 2015, 59, 1837–1843. [Google Scholar] [CrossRef] [PubMed]
- Bittner, A.; Cramer, B.; Harrer, H.; Humpf, H.-U. Structure elucidation and in vitro cytotoxicity of ochratoxin α amide, a new degradation product of ochratoxin A. Mycotoxin Res. 2015, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bittner, A.; Cramer, B.; Humpf, H.-U. Matrix binding of ochratoxin a during roasting. J. Agric. Food Chem. 2013, 61, 12737–12743. [Google Scholar] [CrossRef]
- Cramer, B.; Harrer, H.; Nakamura, K.; Uemura, D.; Humpf, H.-U. Total synthesis and cytotoxicity evaluation of all ochratoxin A stereoisomers. Bioorganic Med. Chem. 2010, 18, 343–347. [Google Scholar] [CrossRef]
- Cramer, B.; Königs, M.; Humpf, H.-U. Identification and in vitro cytotoxicity of ochratoxin A degradation products formed during coffee roasting. J. Agric. Food Chem. 2008, 56, 5673–5681. [Google Scholar] [CrossRef]
- Ringot, D.; Chango, A.; Schneider, Y.; Larondelle, Y. Toxicokinetics and toxicodynamics of ochratoxin A, an update. Chem.-Biol. Interact. 2006, 159, 18–46. [Google Scholar] [CrossRef]
- Meisner, H.; Chan, S. Ochratoxin A, an inhibitor of mitochondrial transport systems. Biochemistry 1974, 13, 2795–2800. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Lu, C.; Lin, T.; Wei, R. Effect of ochratoxin A on rat liver mitochondrial respiration and oxidative phosphorylation. Toxicology 1985, 36, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Bunge, I.; Dirheimer, G.; Röschenthaler, R. In vivo in vitroinhibition of protein synthesis in Bacillus stearothermophilus by ochratoxin A. Biochem. Biophys. Res. Commun. 1978, 83, 398–405. [Google Scholar] [CrossRef] [PubMed]
- Creppy, E.; Röschenthaler, R.; Dirheimer, G. Inhibition of protein synthesis in mice by ochratoxin A and its prevention by phenylalanine. Food Chem. Toxicol. 1984, 22, 883–886. [Google Scholar] [CrossRef] [PubMed]
- Creppy, E.; Størmer, F.; Kern, D.; Röschenthaler, R.; Dirheimer, G. Effects of ochratoxin A metabolites on yeast phenylalanyl-tRNA synthetase and on the growth and in vivo protein synthesis of hepatoma cells. Chem.-Biol. Interact. 1983, 47, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Baudrimont, I.; Ahouandjivo, R.; Creppy, E. Prevention of lipid peroxidation induced by ochratoxin A in Vero cells in culture by several agents. Chem.-Biol. Interact. 1997, 104, 29–40. [Google Scholar] [CrossRef]
- Kamp, H.G.; Eisenbrand, G.; Janzowski, C.; Kiossev, J.; Latendresse, J.R.; Schlatter, J.; Turesky, R.J. Ochratoxin A induces oxidative DNA damage in liver and kidney after oral dosing to rats. Mol. Nutr. Food Res. 2005, 49, 1160–1167. [Google Scholar] [CrossRef]
- Mally, A.; Pepe, G.; Ravoori, S.; Fiore, M.; Gupta, R.C.; Dekant, W.; Mosesso, P. Ochratoxin A causes DNA damage and cytogenetic effects but no DNA adducts in rats. Chem. Res. Toxicol. 2005, 18, 1253–1261. [Google Scholar] [CrossRef]
- Rahimtula, A.; Bereziat, J.; Bussacchini-Griot, V.; Bartsch, H. Lipid peroxidation as a possible cause of ochratoxin A toxicity. Biochem. Pharmacol. 1988, 37, 4469–4477. [Google Scholar] [CrossRef]
- Omar, R.F.; Hasinoff, B.B.; Mejilla, F.; Rahimtula, A.D. Mechanism of ochratoxin A stimulated lipid peroxidation. Biochem. Pharmacol. 1990, 40, 1183–1191. [Google Scholar] [CrossRef]
- Omar, R.F.; Rahimtula, A.D.; Bartsch, H. Role of cytochrome P-450 in ochratoxin a-stimulated lipid peroxidation. J. Biochem. Toxicol. 1991, 6, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Baudrimont, I.; Betbeder, A.-M.; Gharbi, A.; Pfohl-Leszkowicz, A.; Dirheimer, G.; Creppy, E. Effect of superoxide dismutase and catalase on the nephrotoxicity induced by subchronical administration of ochratoxin A in rats. Toxicology 1994, 89, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Cavin, C.; Delatour, T.; Marin-Kuan, M.; Holzhäuser, D.; Higgins, L.; Bezencon, C.; Guignard, G.; Junod, S.; Richoz-Payot, J.; Gremaud, E. Reduction in antioxidant defenses may contribute to ochratoxin A toxicity and carcinogenicity. Toxicol. Sci. 2007, 96, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Petrik, J.; Žanić-Grubišić, T.; Barišić, K.; Pepeljnjak, S.; Radić, B.; Ferenčić, Ž.; Čepelak, I. Apoptosis and oxidative stress induced by ochratoxin A in rat kidney. Arch. Toxicol. 2003, 77, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Schaaf, G.; Nijmeijer, S.; Maas, R.; Roestenberg, P.; De Groene, E.; Fink-Gremmels, J. The role of oxidative stress in the ochratoxin A-mediated toxicity in proximal tubular cells. Biochim. Et Biophys. Acta (BBA)-Mol. Basis Dis. 2002, 1588, 149–158. [Google Scholar] [CrossRef]
- Klaunig, J.E.; Kamendulis, L.M. The role of oxidative stress in carcinogenesis. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 239–267. [Google Scholar] [CrossRef]
- Marin-Kuan, M.; Cavin, C.; Delatour, T.; Schilter, B. Ochratoxin A carcinogenicity involves a complex network of epigenetic mechanisms. Toxicon 2008, 52, 195–202. [Google Scholar] [CrossRef]
- Lee, H.J.; Pyo, M.C.; Shin, H.S.; Ryu, D.; Lee, K.-W. Renal toxicity through AhR, PXR, and Nrf2 signaling pathway activation of ochratoxin A-induced oxidative stress in kidney cells. Food Chem. Toxicol. 2018, 122, 59–68. [Google Scholar] [CrossRef]
- Shin, H.S.; Lee, H.J.; Pyo, M.C.; Ryu, D.; Lee, K.-W. Ochratoxin A-induced hepatotoxicity through phase I and phase II reactions regulated by AhR in liver cells. Toxins 2019, 11, 377. [Google Scholar] [CrossRef]
- Marin-Kuan, M.; Nestler, S.; Verguet, C.; Bezencon, C.; Piguet, D.; Mansourian, R.; Holzwarth, J.; Grigorov, M.; Delatour, T.; Mantle, P. A toxicogenomics approach to identify new plausible epigenetic mechanisms of ochratoxin a carcinogenicity in rat. Toxicol. Sci. 2005, 89, 120–134. [Google Scholar] [CrossRef]
- Boesch-Saadatmandi, C.; Loboda, A.; Jozkowicz, A.; Huebbe, P.; Blank, R.; Wolffram, S.; Dulak, J.; Rimbach, G. Effect of ochratoxin A on redox-regulated transcription factors, antioxidant enzymes and glutathione-S-transferase in cultured kidney tubulus cells. Food Chem. Toxicol. 2008, 46, 2665–2671. [Google Scholar] [CrossRef] [PubMed]
- Boesch-Saadatmandi, C.; Wagner, A.; Graeser, A.; Hundhausen, C.; Wolffram, S.; Rimbach, G. Ochratoxin A impairs Nrf2-dependent gene expression in porcine kidney tubulus cells. J. Anim. Physiol. Anim. Nutr. 2009, 93, 547–554. [Google Scholar] [CrossRef] [PubMed]
- García-Pérez, E.; Ryu, D.; Kim, H.-Y.; Kim, H.D.; Lee, H.J. Human Proximal Tubule Epithelial Cells (HK-2) as a Sensitive In Vitro System for Ochratoxin A Induced Oxidative Stress. Toxins 2021, 13, 787. [Google Scholar] [CrossRef] [PubMed]
- García-Pérez, E.; Ryu, D.; Lee, C.; Lee, H.J. Ochratoxin a induces oxidative stress in hepg2 cells by impairing the gene expression of antioxidant enzymes. Toxins 2021, 13, 271. [Google Scholar] [CrossRef] [PubMed]
- Descamps-Latscha, B.; Witko-Sarsat, V. Importance of oxidatively modified proteins in chronic renal failure. Kidney Int. 2001, 59, S108–S113. [Google Scholar] [CrossRef]
- Beisswenger, P.J.; Drummond, K.S.; Nelson, R.G.; Howell, S.K.; Szwergold, B.S.; Mauer, M. Susceptibility to diabetic nephropathy is related to dicarbonyl and oxidative stress. Diabetes 2005, 54, 3274–3281. [Google Scholar] [CrossRef]
- Coşkun, C.; Kural, A.; DÖventaş, Y.; Koldaş, M.; Oezturk, H.; Inal, B.B.; GÜmÜş, A. Hemodialysis and protein oxidation products. Ann. N. Y. Acad. Sci. 2007, 1100, 404–408. [Google Scholar] [CrossRef]
- Nguyen, K.; Ryu, D. Ultrasonic extraction with ultra-performance liquid chromatography/tandem mass spectrometry for the determination of ochratoxin A in processed cereal products. J. AOAC Int. 2014, 97, 1384–1386. [Google Scholar] [CrossRef]
- Vega, M.; Muñoz, K.; Sepúlveda, C.; Aranda, M.; Campos, V.; Villegas, R.; Villarroel, O. Solid-phase extraction and HPLC determination of ochratoxin A in cereals products on Chilean market. Food Control 2009, 20, 631–634. [Google Scholar] [CrossRef]
- Bakker, M.; Pieters, M. Risk Assessment of Ochratoxin a in The Netherlands. 2003. Available online: https://rivm.openrepository.com/handle/10029/9185 (accessed on 30 May 2021).
- Roscoe, V.; Lombaert, G.; Huzel, V.; Neumann, G.; Melietio, J.; Kitchen, D.; Kotello, S.; Krakalovich, T.; Trelka, R.; Scott, P. Mycotoxins in breakfast cereals from the Canadian retail market: A 3-year survey. Food Addit. Contam. 2008, 25, 347–355. [Google Scholar] [CrossRef]
- HC. Information Document on Health Canada’s Proposed Maximum Limits (Standards) for the Presence of the Mycotoxin Ochratoxin a in Foods; F.D. Bureau of Chemical Safety, Health Products and Food Branch, 2009. Available online: https://www.canada.ca/en/health-canada/services/food-nutrition/public-involvement-partnerships/information-document-proposed-maximum-limits-standards-presence-mycotoxin-ochratoxin-foods.html (accessed on 15 January 2018).
- Kuzdraliński, A.; Solarska, E.; Mazurkiewicz, J. Mycotoxin content of organic and conventional oats from southeastern Poland. Food Control 2013, 33, 68–72. [Google Scholar] [CrossRef]
- Kuruc, J.; Hegstad, J.; Lee, H.J.; Simons, K.; Ryu, D.; Wolf-Hall, C. Infestation and quantification of ochratoxigenic fungi in barley and wheat naturally contaminated with ochratoxin A. J. Food Prot. 2015, 78, 1350–1356. [Google Scholar] [CrossRef] [PubMed]
- Al-Taher, F.; Cappozzo, J.; Zweigenbaum, J.; Lee, H.J.; Jackson, L.; Ryu, D. Detection and quantitation of mycotoxins in infant cereals in the US market by LC-MS/MS using a stable isotope dilution assay. Food Control 2017, 72, 27–35. [Google Scholar] [CrossRef]
- De Jesus, C.L.; Bartley, A.; Welch, A.Z.; Berry, J.P. High incidence and levels of ochratoxin A in wines sourced from the United States. Toxins 2018, 10, 1. [Google Scholar] [CrossRef] [PubMed]
- Bellver Soto, J.; Fernandez-Franzon, M.; Ruiz, M.-J.; Juan-Garcia, A. Presence of ochratoxin A (OTA) mycotoxin in alcoholic drinks from southern European countries: Wine and beer. J. Agric. Food Chem. 2014, 62, 7643–7651. [Google Scholar] [CrossRef]
- Romani, S.; Sacchetti, G.; Chaves López, C.; Pinnavaia, G.G.; Dalla Rosa, M. Screening on the occurrence of ochratoxin A in green coffee beans of different origins and types. J. Agric. Food Chem. 2000, 48, 3616–3619. [Google Scholar] [CrossRef]
- Martins, M.; Martins, H.; Gimeno, A. Incidence of microflora and of ochratoxin A in green coffee beans (Coffea arabica). Food Addit. Contam. 2003, 20, 1127–1131. [Google Scholar] [CrossRef]
- Pardo, E.; Marin, S.; Ramos, A.; Sanchis, V. Occurrence of ochratoxigenic fungi and ochratoxin A in green coffee from different origins. Food Sci. Technol. Int. 2004, 10, 45–49. [Google Scholar] [CrossRef]
- EC. Commission regulation (EC) no 1881/2006 of 19 December 2006 Setting levels for certain contaminants in food stuffs. Off. J. Eur. Union 2006, 364, 5–24. [Google Scholar]
- MHRF. National Sanitary and Epidemiological Rules and Regulations Sanpin 2.3.2.1078-01 ‘Hygienic Requirements for Safety and Nutrition Value of Foodstuff’; Russian Federation: Moscow, Russia, 2001. [Google Scholar]
- ANVISA. Agência Nacional de Vigilância Sanitária. Dispõe Sobre Limites Máximos Tolerados (LMT) Para Micotoxinas em Alimentos. Diário Oficial da União; Brasília, Brazil: 2011. Resolução RDC n. 7, 18/02/2011. Available online: https://bvsms.saude.gov.br/bvs/saudelegis/anvisa/2011/res0007_18_02_2011_rep.html (accessed on 18 January 2020).
- MOHC. Gb 7718-2011 National Food Safety Standard General Standard for the Labeling of Prepackaged Foods; Ministry of Health of the People’s Republic of China: Beijing, China, 2011. [Google Scholar]
- MOHI. Food Safety and Standard (Contaminants, Toxins and Residues) Regulation; Ministry of Health and Family Welfare: New Delhi, India; Food Safety and Standards Authority of India: New Delhi, India, 2011; pp. 1–19. [Google Scholar]
- KFDA. Enforcement Rule of Food Sanitation Act; Food Fode Korea Food and Drug Administration: Seoul, Republic of Korea, 2012. [Google Scholar]
- Miraglia, M.; Brera, C. Assessment of dietary intake of ochratoxin A by the population of EU member states. Reports on tasks for scientific cooperation. Rep. Experts Particip. Task 3.2.7. 2002, 3, 20–22. [Google Scholar]
- Rousseau, J. Ochratoxin a in wines: Current knowledge-mycotoxins and wine. Vinidea.Net Wine Internet Tech. J. 2004, 5, 1–5. [Google Scholar]
- Walker, R. Risk assessment of ochratoxin: Current views of the european scientific committee on food, the jecfa and the codex committee on food additives and contaminants. In Mycotoxins and Food Safety; Springer: Berlin/Heidelberg, Germany, 2002; pp. 249–255. [Google Scholar]
- EFSA. Opinion of the Scientific Panel on Contaminants in the Food Chain on a Request from the Commission Related to Ochratoxin a in Food; Authority, EFSA: Palma, Italy, 2006; pp. 1–56. [Google Scholar]
- EFSA Panel on Contaminants in the Food Chain; Schrenk, D.; Bodin, L.; Chipman, J.K.; del Mazo, J.; Grasl-Kraupp, B.; Hogstrand, C.; Hoogenboom, L.; Leblanc, J.C.; Nebbia, C.S.; et al. Risk assessment of ochratoxin A in food. EFSA J. 2020, 18, e06113. [Google Scholar]
- Alldrick, A. The effects of processing on the occurrence of ochratoxin A in cereals. Food Addit. Contam. 1996, 13, 27–28. [Google Scholar] [PubMed]
- Scudamore, K.; Banks, J.; MacDonald, S. Fate of ochratoxin A in the processing of whole wheat grains during milling and bread production. Food Addit. Contam. 2003, 20, 1153–1163. [Google Scholar] [CrossRef]
- Valle-Algarra, F.M.; Mateo, E.M.; Medina, A.; Mateo, F.; Gimeno-Adelantado, J.V.; Jiménez, M. Changes in ochratoxin A and type B trichothecenes contained in wheat flour during dough fermentation and bread-baking. Food Addit. Contam. 2009, 26, 896–906. [Google Scholar] [CrossRef]
- Simsek, S.; Burgess, K.; Whitney, K.L.; Gu, Y.; Qian, S.Y. Analysis of deoxynivalenol and deoxynivalenol-3-glucoside in wheat. Food Control 2012, 26, 287–292. [Google Scholar] [CrossRef]
- Young, J.C.; Fulcher, R.G.; Hayhoe, J.H.; Scott, P.M.; Dexter, J.E. Effect of milling and baking on deoxynivalenol (vomitoxin) content of eastern Canadian wheats. J. Agric. Food Chem. 1984, 32, 659–664. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, B. Fate of deoxynivalenol and deoxynivalenol-3-glucoside during wheat milling and Chinese steamed bread processing. Food Control 2014, 44, 86–91. [Google Scholar] [CrossRef]
- Kupski, L.; Queiroz, M.I.; Badiale-Furlong, E. Application of carboxypeptidase A to a baking process to mitigate contamination of wheat flour by ochratoxin A. Process Biochem. 2018, 64, 248–254. [Google Scholar] [CrossRef]
- Manda, P.; Dano, D.S.; Kouadio, J.H.; Diakite, A.; Sangare-Tigori, B.; Ezoulin, M.J.M.; Soumahoro, A.; Dembele, A.; Fourny, G. Impact of industrial treatments on ochratoxin A content in artificially contaminated cocoa beans. Food Addit. Contam. 2009, 26, 1081–1088. [Google Scholar] [CrossRef]
- Amézqueta, S.; Gonzalez-Penas, E.; Murillo, M.; Lopez de Cerain, A. Occurrence of ochratoxin A in cocoa beans: Effect of shelling. Food Addit. Contam. 2005, 22, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Scudamore, K.A. Prevention of ochratoxin A in commodities and likely effects of processing fractionation and animal feeds. Food Addit. Contam. 2005, 22, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Chung, S.-H.; Lee, C.; Kim, Y.-B. Fate of ochratoxin A during cooking of naturally contaminated polished rice. J. Food Prot. 2005, 68, 2107–2111. [Google Scholar] [CrossRef] [PubMed]
- Mansouri-Nasrabadi, R.; Milani, J.M.; Nazari, S.S.J. Optimization of washing and cooking processes of rice for Ochratoxin A decrement by RSM. Food Sci. Nutr. 2018, 6, 2523–2529. [Google Scholar] [CrossRef] [PubMed]
- Blanc, M.; Pittet, A.; Muñoz-Box, R.; Viani, R. Behavior of ochratoxin A during green coffee roasting and soluble coffee manufacture. J. Agric. Food Chem. 1998, 46, 673–675. [Google Scholar] [CrossRef] [PubMed]
- Jalili, M.; Jinap, S.; Son, R. The effect of chemical treatment on reduction of aflatoxins and ochratoxin A in black and white pepper during washing. Food Addit. Contam. 2011, 28, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Iha, M.; Trucksess, M.; Tournas, V. Effect of processing on ochratoxin A content in dried beans. Food Addit. Contam. 2009, 26, 1389–1395. [Google Scholar] [CrossRef]
- Amézqueta, S.; Gonzalez-Penas, E.; Lizarraga, T.; Murillo-Arbizu, M.; De Cerain, A.L. A simple chemical method reduces ochratoxin A in contaminated cocoa shells. J. Food Prot. 2008, 71, 1422–1426. [Google Scholar] [CrossRef]
- Osborne, B.; Ibe, F.; Brown, G.; Petagine, F.; Scudamore, K.; Banks, J.; Hetmanski, M.; Leonard, C. The effects of milling and processing on wheat contaminated with ochratoxin A. Food Addit. Contam. 1996, 13, 141–153. [Google Scholar] [CrossRef]
- Peng, C.; Wang, L.; An, F.; Zhang, L.; Wang, Y.; Li, S.; Wang, C.; Liu, H. Fate of ochratoxin A during wheat milling and some Chinese breakfast processing. Food Control 2015, 57, 142–146. [Google Scholar] [CrossRef]
- Scott, P.; Kanhere, S.; Lawrence, G.; Daley, E.; Farber, J. Fermentation of wort containing added ochratoxin A and fumonisins B1 and B2. Food Addit. Contam. 1995, 12, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Krogh, P.; Hald, B.; Gjertsen, P.; Myken, F. Fate of ochratoxin A and citrinin during malting and brewing experiments. Appl. Microbiol. 1974, 28, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Chu, F.; Chang, C.; Ashoor, S.H.; Prentice, N. Stability of aflatoxin B1 and ochratoxin A in brewing. Appl. Microbiol. 1975, 29, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Nip, W.; Chang, F.C.; Chu, F.; Prentice, N. Fate of ochratoxin A in brewing. Appl. Microbiol. 1975, 30, 1048–1049. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Nagatomi, Y.; Uyama, A.; Mochizuki, N. Fate of mycotoxins during beer brewing and fermentation. Biosci. Biotechnol. Biochem. 2013, 77, 1410–1415. [Google Scholar] [CrossRef] [PubMed]
- Delage, N.; d’Harlingue, A.; Colonna Ceccaldi, B.; Bompeix, G. Occurrence of mycotoxins in fruit juices and wine. Food Control 2003, 14, 225–227. [Google Scholar] [CrossRef]
- Zimmerli, B.; Dick, R. Ochratoxin A in table wine and grape-juice: Occurrence and risk assessment. Food Addit. Contam. 1996, 13, 655–668. [Google Scholar] [CrossRef]
- Freire, L.; Braga, P.A.; Furtado, M.M.; Delafiori, J.; Dias-Audibert, F.L.; Pereira, G.E.; Reyes, F.G.; Catharino, R.R.; Sant’Ana, A.S. From grape to wine: Fate of ochratoxin A during red, rose, and white winemaking process and the presence of ochratoxin derivatives in the final products. Food Control 2020, 113, 107167. [Google Scholar] [CrossRef]
- Csutorás, C.; Rácz, L.; Rácz, K.; Fűtő, P.; Forgó, P.; Kiss, A. Monitoring of ochratoxin A during the fermentation of different wines by applying high toxin concentrations. Microchem. J. 2013, 107, 182–184. [Google Scholar] [CrossRef]
- Cecchini, F.; Morassut, M.; Moruno, E.G.; Di Stefano, R. Influence of yeast strain on ochratoxin A content during fermentation of white and red must. Food Microbiol. 2006, 23, 411–417. [Google Scholar] [CrossRef]
- Meca, G.; Blaiotta, G.; Ritieni, A. Reduction of ochratoxin A during the fermentation of Italian red wine Moscato. Food Control 2010, 21, 579–583. [Google Scholar] [CrossRef]
- Yu, J.; Smith, I.N.; Mikiashvili, N. Reducing Ochratoxin A Content in Grape Pomace by Different Methods. Toxins 2020, 12, 424. [Google Scholar] [CrossRef] [PubMed]
- Milani, J.; Heidari, S. Stability of ochratoxin A during bread making process. J. Food Saf. 2017, 37, e12283. [Google Scholar] [CrossRef]
- Kabak, B. The fate of mycotoxins during thermal food processing. J. Sci. Food Agric. 2009, 89, 549–554. [Google Scholar] [CrossRef]
- Boudra, H.; Le Bars, P.; Le Bars, J. Thermostability of Ochratoxin A in wheat under two moisture conditions. Appl. Environ. Microbiol. 1995, 61, 1156–1158. [Google Scholar] [CrossRef] [PubMed]
- Studer-Rohr, I.; Dietrich, D.R.; Schlatter, J.; Schlatter, C. The occurrence of ochratoxin A in coffee. Food Chem. Toxicol. 1995, 33, 341–355. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Quiroz, M.; Louise, B.D.; Gonzalez-Rios, O.; Barel, M.; Guyot, B.; Schorr-Galindo, S.; Guiraud, J.P. The impact of roasting on the ochratoxin A content of coffee. Int. J. Food Sci. Technol. 2005, 40, 605–611. [Google Scholar] [CrossRef]
- Nehad, E.; Farag, M.; Kawther, M.; Abdel-Samed, A.; Naguib, K. Stability of ochratoxin A (OTA) during processing and decaffeination in commercial roasted coffee beans. Food Addit. Contam. 2005, 22, 761–767. [Google Scholar] [CrossRef]
- Castellanos-Onorio, O.; Gonzalez-Rios, O.; Guyot, B.; Fontana, T.A.; Guiraud, J.-P.; Schorr-Galindo, S.; Durand, N.; Suárez-Quiroz, M. Effect of two different roasting techniques on the Ochratoxin A (OTA) reduction in coffee beans (Coffea arabica). Food Control 2011, 22, 1184–1188. [Google Scholar] [CrossRef]
- Romani, S.; Pinnavaia, G.G.; Dalla Rosa, M. Influence of roasting levels on ochratoxin A content in coffee. J. Agric. Food Chem. 2003, 51, 5168–5171. [Google Scholar] [CrossRef]
- Tsubouchi, H.; Yamamoto, K.; Hisada, K.; Sakabe, Y.; Udagawa, S.-i. Effect of roasting on ochratoxin A level in green coffee beans inoculated with Aspergillus ochraceus. Mycopathologia 1987, 97, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Pérez de Obanos, A.; González-Peñas, E.; Lopez de Cerain, A. Influence of roasting and brew preparation on the ochratoxin A content in coffee infusion. Food Addit. Contam. 2005, 22, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Viani, R. Fate of ochratoxin A (OTA) during processing of coffee. Food Addit. Contam. 1996, 13, 29–33. [Google Scholar] [PubMed]
- Lee, H.J. Stability of ochratoxin A in oats during roasting with reducing sugars. Food Control 2020, 118, 107382. [Google Scholar] [CrossRef]
- Carbon, H.; Lee, H. Varied reduction of ochratoxin A in brown and white rice during roasting. World Mycotoxin J. 2022, 15, 361–368. [Google Scholar] [CrossRef]
- Lee, H.J.; Kim, S.; Suh, H.J.; Ryu, D. Effects of explosive puffing process on the reduction of ochratoxin A in rice and oats. Food Control 2019, 95, 334–338. [Google Scholar] [CrossRef]
- Osborne, B.G. Reversed-phase high performance liquid chromatography determination of ochratoxin a in flour and bakery products. J. Sci. Food Agric. 1979, 30, 1065–1070. [Google Scholar] [CrossRef]
- Milanez, T.; Leitão, M. The effect of cooking on ochratoxin A content of beans, variety ‘carioca’. Food Addit. Contam. 1996, 13, 89–93. [Google Scholar] [CrossRef]
- Trenk, H.L.; Butz, M.E.; Chu, F.S. Production of ochratoxins in different cereal products by Aspergillus ochraceus. Appl. Microbiol. 1971, 21, 1032–1035. [Google Scholar] [CrossRef]
- Lee, H.J.; Lee, C.; Ryu, D. Effects of baking soda and fructose in reduction of ochratoxin A in rice and oat porridge during retorting process. Food Control 2020, 116, 107325. [Google Scholar] [CrossRef]
- Lee, H.J.; Gu, B.-J.; Ganjyal, G.; Ryu, D. Reduction of ochratoxin A in direct steam injected oat-based infant cereals with baking soda. Food Control 2019, 96, 441–444. [Google Scholar] [CrossRef]
- Lee, H.J.; Li, S.; Gu, K.; Ryu, D. Reduction of Ochratoxin A during the Preparation of Porridge with Sodium Bicarbonate and Fructose. Toxins 2021, 13, 224. [Google Scholar] [CrossRef] [PubMed]
- Gahlawat, P.; Sehgal, S. Phytic acid, saponins, and polyphenols in weaning foods prepared from oven-heated green gram and cereals. Cereal Chem. 1992, 69, 463–464. [Google Scholar]
- Karlovsky, P.; Suman, M.; Berthiller, F.; De Meester, J.; Eisenbrand, G.; Perrin, I.; Oswald, I.P.; Speijers, G.; Chiodini, A.; Recker, T. Impact of food processing and detoxification treatments on mycotoxin contamination. Mycotoxin Res. 2016, 32, 179–205. [Google Scholar] [CrossRef] [PubMed]
- La Pera, L.; Avellone, G.; Lo Turco, V.; Di Bella, G.; Agozzino, P.; Dugo, G. Influence of roasting and different brewing processes on the ochratoxin A content in coffee determined by high-performance liquid chromatography-fluorescence detection (HPLC-FLD). Food Addit. Contam. 2008, 25, 1257–1263. [Google Scholar] [CrossRef] [PubMed]
- Malir, F.; Ostry, V.; Pfohl-Leszkowicz, A.; Toman, J.; Bazin, I.; Roubal, T. Transfer of ochratoxin A into tea and coffee beverages. Toxins 2014, 6, 3438–3453. [Google Scholar] [CrossRef] [PubMed]
- Harper, J.M. A comparative analysis of single and twin-screw extruders. In Food Extrusion Science and Technology; Kokini, J.L., Ho, C., Karwe, M.V., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 1992; pp. 139–148. [Google Scholar]
- Humpf, H.U.; Voss, K.A. Effects of thermal food processing on the chemical structure and toxicity of fumonisin mycotoxins. Mol. Nutr. Food Res. 2004, 48, 255–269. [Google Scholar] [CrossRef]
- Cetin, Y.; Bullerman, L.B. Evaluation of reduced toxicity of zearalenone by extrusion processing as measured by the MTT cell proliferation assay. J. Agric. Food Chem. 2005, 53, 6558–6563. [Google Scholar] [CrossRef]
- Voss, K.A.; Bullerman, L.B.; Bianchini, A.; Hanna, M.A.; Ryu, D. Reduced toxicity of fumonisin B1 in corn grits by single-screw extrusion. J. Food Prot. 2008, 71, 2036–2041. [Google Scholar] [CrossRef]
- Scudamore, K.; Banks, J.; Guy, R. Fate of ochratoxin A in the processing of whole wheat grain during extrusion. Food Addit. Contam. 2004, 21, 488–497. [Google Scholar] [CrossRef]
- Castells, M.; Pardo, E.; Ramos, A.J.; Sanchis, V.; Marín, S. Reduction of ochratoxin A in extruded barley meal. J. Food Prot. 2006, 69, 1139–1143. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Dahal, S.; Perez, E.G.; Kowalski, R.J.; Ganjyal, G.; Ryu, D. Reduction of ochratoxin A in oat flakes by twin screw extrusion processing. J. Food Prot. 2017, 80, 1628–1634. [Google Scholar] [CrossRef] [PubMed]
- Ryu, D.; Kowalski, R.J.; Ganjyal, G.; Lee, H.J. Reduction of ochratoxin A in oats and rice by twin-screw extrusion processing with baking soda. Food Control 2019, 105, 21–28. [Google Scholar] [CrossRef]
- Subirade, I. Fate of ochratoxin A during breadmaking. Food Addit. Contam. 1995, 13, 25–26. [Google Scholar]
- Vidal, A.; Sanchis, V.; Ramos, A.J.; Marín, S. Thermal stability and kinetics of degradation of deoxynivalenol, deoxynivalenol conjugates and ochratoxin A during baking of wheat bakery products. Food Chem. 2015, 178, 276–286. [Google Scholar] [CrossRef]
- Bol, E.K.; Araujo, L.; Veras, F.F.; Welke, J.E. Estimated exposure to zearalenone, ochratoxin A and aflatoxin B1 through the consume of bakery products and pasta considering effects of food processing. Food Chem. Toxicol. 2016, 89, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Bryła, M.; Ksieniewicz-Woźniak, E.; Stępniewska, S.; Modrzewska, M.; Waśkiewicz, A.; Szymczyk, K.; Szafrańska, A. Transformation of ochratoxin A during bread-making processes. Food Control 2021, 125, 107950. [Google Scholar] [CrossRef]
- Smith, I.N.; Yu, J. Nutritional and sensory quality of bread containing different quantities of grape pomace from different grape cultivars. EC Nutr. 2015, 2, 291–301. [Google Scholar]
- Yu, J.; Smith, I.N.; Chen, G. Influence of Grape Pomace Inclusion on Physical and Chemical Properties of Corn-Based Extrudates. SDRP J. Food Sci. Technol. 2018, 3, 516–526. [Google Scholar] [CrossRef]
- Maman, R.; Yu, J. Chemical composition and particle size of grape seed flour and their effects on the characteristics of cookies. J. Food Res. 2019, 8, 111. [Google Scholar] [CrossRef]
- Hussain, A.; Luttfullah, G. Reduction of aflatoxin-beta/sub 1/and ochratoxin-A levels in polished basmati rice (oryza sativa linn.) by different cooking methods. J. Chem. Soc. Pak. 2009, 31, 911–915. [Google Scholar]
- Lewis, M.; Heppell, N.; Hastings, A. Continuous Thermal Processing of Foods—Pasteurization and Uht Sterilization; Aspen Publishers: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Katofsky, R.E. The Production of Fluid Fuels from Biomass; Princeton University: Princeton, NJ, USA, 1993. [Google Scholar]
- Sullivan, J.; Craig, J., Jr. The development of explosion puffing. Food Technol. 1984, 38, 52–55. [Google Scholar]
- Hoke, K.; Houška, M.; Průchová, J.; Gabrovská, D.; Vaculová, K.; Paulíčková, I. Optimisation of puffing naked barley. J. Food Eng. 2007, 80, 1016–1022. [Google Scholar] [CrossRef]
- Hui, Y.H.; Evranuz, E.Ö. Handbook of Vegetable Preservation and Processing; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Pleadin, J.; Perši, N.; Kovačević, D.; Vulić, A.; Frece, J.; Markov, K. Ochratoxin A reduction in meat sausages using processing methods practiced in households. Food Addit. Contam. Part B 2014, 7, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Josefsson, B.E.; Möller, T.E. Heat stability of ochratoxin A in pig products. J. Sci. Food Agric. 1980, 31, 1313–1315. [Google Scholar] [CrossRef] [PubMed]
- OECD. Sodium Bicarbonate. (cas no. 144-55-8), Sids Initial Assessment Report for Siam 15; OECD: Boston, MA, USA, 2002. [Google Scholar]
- FDA. CFR—Code of Federal Regulations Title 21; FDA: Rockville, MD, USA, 2017. [Google Scholar]
- Howard, P.C.; Churchwell, M.I.; Couch, L.H.; Marques, M.M.; Doerge, D.R. Formation of N-(carboxymethyl) fumonisin B1, following the reaction of fumonisin B1 with reducing sugars. J. Agric. Food Chem. 1998, 46, 3546–3557. [Google Scholar] [CrossRef]
- Lu, Y.; Clifford, L.; Hauck, C.C.; Hendrich, S.; Osweiler, G.; Murphy, P.A. Characterization of fumonisin B1− glucose reaction kinetics and products. J. Agric. Food Chem. 2002, 50, 4726–4733. [Google Scholar] [CrossRef]
- Fernández-Surumay, G.; Osweiler, G.D.; Yaeger, M.J.; Rottinghaus, G.E.; Hendrich, S.; Buckley, L.K.; Murphy, P.A. Fumonisin B− glucose reaction products are less toxic when fed to swine. J. Agric. Food Chem. 2005, 53, 4264–4271. [Google Scholar] [CrossRef]
- Murphy, P.A.; Hendrich, S.; Hopmans, E.C.; Hauck, C.C.; Lu, Z.; Buseman, G.; Munkvold, G. Effect of processing on fumonisin content of corn. Fumonisins Food 1996, 392, 323–334. [Google Scholar]
- Gu, K.; Ryu, D.; Lee, H.J. Ochratoxin A and its reaction products affected by sugars during heat processing. Food Chem. 2021, 348, 129038. [Google Scholar] [CrossRef]
- Mine Kurtbay, H.; Bekçi, Z.; Merdivan, M.; Yurdakoç, K. Reduction of ochratoxin A levels in red wine by bentonite, modified bentonites, and chitosan. J. Agric. Food Chem. 2008, 56, 2541–2545. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Heydt, M.; Cramer, B.; Graf, I.; Lerch, S.; Humpf, H.-U.; Geisen, R. Wavelength-dependent degradation of ochratoxin and citrinin by light in vitro and in vivo and its implications on Penicillium. Toxins 2012, 4, 1535–1551. [Google Scholar] [CrossRef] [PubMed]
- Sueck, F.; Hemp, V.; Specht, J.; Torres, O.; Cramer, B.; Humpf, H.-U. Occurrence of the ochratoxin A degradation product 2′ R-ochratoxin a in coffee and other food: An update. Toxins 2019, 11, 329. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Authority | Commodity | Max. Level of OTA (ng/g) |
|---|---|---|
| EU | Unprocessed cereals | 5.0 |
| All product derived from unprocessed cereals intended for direct consumption | 3.0 | |
| Dried vine fruits (raisins, currents, sultanas) | 10.0 | |
| Roasted coffee beans, ground roasted coffee | 5.0 | |
| Instant coffee | 10.0 | |
| Beer and wine | 2.0 | |
| Grape juice (not concentrated) | 2.0 | |
| Infant cereal-based foods, dietary foods for medical purposes | 0.5 | |
| Codex | Unprocessed cereals of wheat, barley, and rye | 5.0 |
| China | Grains and beans | 5.0 |
| Singapore | Beer, cereal and cereal products, coffee | 3.0 |
| Brazil | Cereals and cereal-based products excl. maize and maize derivates | 10.0 |
| Roasted coffee and soluble coffee | 10.0 | |
| Wine | 10.0 | |
| Cereal-based baby food | 2.0 | |
| Dried fruits | 10.0 | |
| India | Unprocessed cereals of wheat, barley, rye | 20.0 |
| Russia | Wheat, barley, rye, oat and rice cereals and cereal products | 5.0 |
| Specific products for children | 0.5 | |
| Korea | Grain and their processed foods (grinding, cutting, etc.), coffee beans and roasted coffee | 5.0 |
| Instant coffee | 10.0 | |
| Grape juice, concentrated grape juice as reconstituted, wine | 2.0 | |
| Raisins | 10.0 | |
| Baby foods for infants and young children | 0.5 | |
| Malaysia | Cereal-based food for infants and children | 0.5 |
| Coffee or ground coffee or coffee powder | 5.0 | |
| Instant coffee or soluble coffee, Decaffeinated coffee | 10.0 | |
| Indonesia | Cereal (rice, corn, sorghum, wheat) and their products | 5.0 |
| Coffee | 5.0 | |
| Instant coffee | 10.0 |
| Type of Thermal Processing | Matrix | Reduction (%) | Reference |
|---|---|---|---|
| Roasting | Coffee | 0–12% | [123] |
| Coffee | 17–86% | [119] | |
| Coffee | 16% | [97] | |
| Coffee | 67% | [124] | |
| Coffee | 86–94% | [125] | |
| Oat | 2–18% | [126] | |
| White/brown rice | 15–60% | [127] | |
| Cocoa beans, shells, and nibs | 24–41% | [92] | |
| Explosive puffing | Oat | 38–52% | [128] |
| Rice | 15–28% | ||
| Baking | Wheat | 20–40% | [86] |
| Wheat | 0% (bread), 67% (biscuits) | [129] | |
| Pressure cooking or autoclaving | Bean | Up to 84% | [130] |
| Oatmeal, rice | 59–89% | [131] | |
| Retorting | Oat | 17 | [132] |
| Rice | 54 | ||
| Direct steam injection | Oat | 20–28% | [133] |
| Indirect steaming | Oat | 13% | [134] |
| Rice | 59% | ||
| Brewing | Beer (barley) | 14–28% | [105] |
| Coffee | 0–97% | [122] | |
| Coffee | 0–25% | [119] | |
| Coffee | 15–50% | [124] |
| Type of Extruder | Matrix | Mode of Contamination and Level | MC * (%) | Mass Flow (g/min) | Temp. (°C) | Screw Speed (rpm) | Die Size (mm) | OTA Reduction (%) | Conditions for Maximum OTA Reduction | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| Single-screw | Barley meal | P. verrucosum; 0.82, 1.43, 0.74 ng/g | 24, 27, 30 | - | 140, 160, 180 | 40–150 | 3 | 17.5–86.5 | 30% MC, 180 °C, 70 s residence time | [144] |
| Twin-screw | Wheat flour | P. verrucosum; 8.5, 41.1 ng/g | 30 | 600 | 120, 135 | 200, 250 | 4 | 10.6–39.2 | 30% MC, 135 °C, 200 rpm | [143] |
| 17.5 | 150, 180 | 350, 400 | 4 | 8.3–34.5 | 17.5% MC, 180 °C, 400 rpm, | |||||
| P. verrucosum; 14.2 ng/g | 17.6–25 | 400, 500 | 160, 180, 200 | 350, 400 | 4 | 21.3–30.7 | 20% MC, 180 °C, 300 rpm, 400 g/min | |||
| P. verrucosum; 49.8 ng/g | 17.6–25 | 400, 500 | 180, 200 | 350, 400 | 4 | 31.6–42.2 | 17.6% MC, 180 °C, 300 rpm, 500 g/min | |||
| Twin-screw | Oat flakes | OTA standard spiking; 100 ng/g | 20, 25, 30 | 66.7 | 140, 160, 180 | 150, 200, 250 | 1.5, 2, 3 | 0–20 | 30% MC, 162 °C, 221 rpm, 3 mm | [145] |
| Twin-screw | Rice flour | OTA standard spiking; 100 ng/g | 16 | 66.7 | 120, 150 | 150, 200, 250 | 3 | 78.4–82.2 | 16% MC, 120 °C, 200 rpm | [146] |
| Oat flakes | 23 | 160 | 150, 200, 250 | 3 | 39.5–42.7 | 23% MC, 160 °C, 150 rpm |
| Type of Additives | Type of Processing | Type of Matrix | OTA Contamination Method and Level | Amount of Additives (%) * | OTA Reduction during Thermal Processing | References | |||
|---|---|---|---|---|---|---|---|---|---|
| Without Additives | With Additives | Effect of Additives | |||||||
| Baking soda | Frying | Wheat flour | A. ochraceus, 93.2 and 248 ng/g | 0.4 | 46.3 ng/g | 38.8 ng/g | ↓ | [102] | |
| 146.5 ng/g | 129.0 ng/g | ↓ | |||||||
| Direct steam injection | Oat flour | OTA standard spiking, 100 ng/g | 0.5 | 19.8–27.9% | 36.1–44.3% | ↓ | [133] | ||
| 1 | 43.4–51.4% | ↓↓ | |||||||
| Twin-screw extrusion | Rice flour | OTA standard spiking, 100 ng/g | 0.5 | 77.9–82.2% | 74.9–78.2% | ↑ | [146] | ||
| 1 | 71.5–77.4% | ↑ | |||||||
| Oat flakes | 0.5 | 39.5–42.7% | 56.2–57.7% | ↓ | |||||
| 1 | 64.6–65.4% | ↓ | |||||||
| Retorting | Rice flour | OTA standard spiking, 20 ng/g | 0.5 | 53.8% | 55.5% | = | [132] | ||
| 1 | 66.4% | ↓ | |||||||
| Oat flour | 0.5 | 17.2% | 30.3% | ↓ | |||||
| 1 | 40.8% | ↓↓ | |||||||
| Indirect steaming | Rice flour | OTA standard spiking, 20 ng/g | 0.5 | 59.4% | 78.1% | ↓ | [134] | ||
| 1 | 68.7% | ↓ | |||||||
| Oat flour | 0.5 | 13.6% | 57.7% | ↓↓↓↓ | |||||
| 1 | 72.6% | ↓↓↓↓ | |||||||
| Sugar | Glucose | Roasting | Oat grain | OTA standard spiking, 100 ng/g | 1 | 10.0% | 11.4% | ↓ | [126] |
| Fructose | 1 | 15.2% | ↓ | ||||||
| Fructose | Retorting | Rice flour | OTA standard spiking, 20 ng/g | 0.5 | 53.8% | 38.7% | ↑ | [132] | |
| 5 | 18.2% | ↑↑↑ | |||||||
| Oat flour | 0.5 | 17.2% | 40.8% | ↓↓↓ | |||||
| 5 | 35.5% | ↓↓ | |||||||
| Fructose | Indirect steaming | Rice flour | OTA standard spiking, 20 ng/g | 0.5 | 59.4% | 62.5% | ↓ | [134] | |
| 1 | 80.7% | ↓↓ | |||||||
| 5 | 66.1% | ↓ | |||||||
| 10 | 60.0% | = | |||||||
| Oat flour | 0.5 | 13.6% | 47.3% | ↓↓↓↓ | |||||
| 1 | 69.3% | ↓↓↓↓ | |||||||
| 5 | 47.5% | ↓↓↓↓ | |||||||
| 10 | 40.7% | ↓↓↓ | |||||||
| Combination | Baking soda + Fructose | Retorting | Rice flour | OTA standard spiking, 20 ng/g | 0.5 + 0.5 | 53.8% | 35.8% | ↑ | [132] |
| Oat flour | 17.2% | 39.8% | ↓↓ | ||||||
| Baking soda + Fructose | Indirect steaming | Rice flour | OTA standard spiking, 20 ng/g | 0.5 + 0.5 | 59.4% | 78.6% | ↓ | [134] | |
| Oat flour | 13.6% | 67.2% | ↓↓↓↓ | ||||||
| ↑ | Less than 20% inhibition of reduction | ||||||||
| ↑↑ | Less than 50% inhibition of reduction | ||||||||
| ↑↑↑ | More than 50% inhibition of reduction | ||||||||
| = | No reduction | ||||||||
| ↓ | <1 times reduction | ||||||||
| ↓↓ | <2 times reduction | ||||||||
| ↓↓↓ | <3 times reduction | ||||||||
| ↓↓↓↓ | more than 3-times reduction | ||||||||
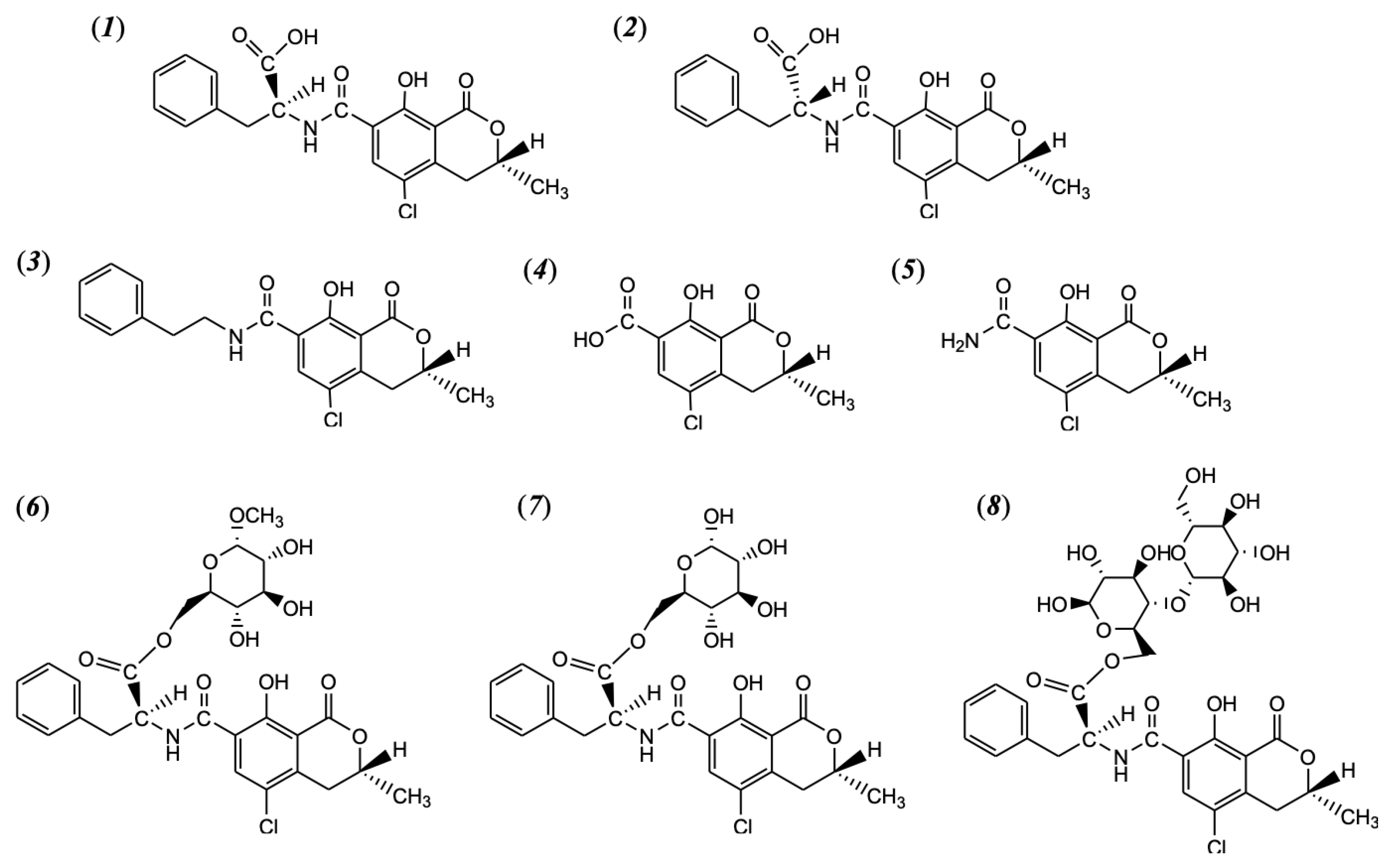
| Compound | Conditions to Form | Analysis | Ref. | |||
|---|---|---|---|---|---|---|
| Initial OTA | Temp (°C) | Time (min) | Additive or Matrix | |||
| 14R-ochratoxin A | 50 μg | 175, 200, and 225 | 5, 10, and 20 | Only OTA | HPLC-FLD | [32] |
| 13 mg | 200 | 10 | Only OTA | ESI-MS, LC-MS/MS, NMR | ||
| 14-decarboxy-ochratoxin A | 50 μg | 175, 200, and 225 | 5, 10, and 20 | Only OTA | HPLC-FLD | |
| 14R-ochratoxin A | 50 μg | 225 | 5 | α-D-glucopyranoside | HPLC-FLD | [30] |
| 14-decarboxy-ochratoxin A | 50 μg | 225 | 5 | α-D-glucopyranoside | HPLC-FLD | |
| OTA-methyl-α-D-glucopyranoside ester | 50 μg | 225 | 5 | α-D-glucopyranoside | HPLC-FLD | |
| 2.5 mg | 200 | 30 | α-D-glucopyranoside | ESI-MS, NMR | ||
| 2.0 mg | 240 | 9 | Cellulose | ESI-MS | ||
| 5 μg/g of coffee | 240 | 9 | Whole coffee beans | LC-MS/MS | ||
| OTA cellobiose ester | 2.0 mg | 240 | 9 | Cellulose | ESI-MS | |
| 5 μg/g of coffee | 240 | 9 | Whole coffee beans | LC-MS/MS | ||
| 14R-ochratoxin A | 100 μg | 200, 220 and 240 | 9 | Only OTA | HPLC-FLD | [29] |
| 14-decarboxy-ochratoxin A | 100 μg | 200, 220 and 240 | 9 | Only OTA | HPLC-FLD | |
| ochratoxin α amide | 100 μg | 200, 220 and 240 | 9 | Only OTA | HPLC-FLD/FTMS | |
| 214.4 mg | 240 | 20 | Only OTA | ESI-MS, NMR, HPLC-ELSD | ||
| ochratoxin α | 100 μg | 200, 220 and 240 | 9 | Only OTA | HPLC-FLD/FTMS | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.J.; Kim, H.D.; Ryu, D. Practical Strategies to Reduce Ochratoxin A in Foods. Toxins 2024, 16, 58. https://doi.org/10.3390/toxins16010058
Lee HJ, Kim HD, Ryu D. Practical Strategies to Reduce Ochratoxin A in Foods. Toxins. 2024; 16(1):58. https://doi.org/10.3390/toxins16010058
Chicago/Turabian StyleLee, Hyun Jung, Hae Dun Kim, and Dojin Ryu. 2024. "Practical Strategies to Reduce Ochratoxin A in Foods" Toxins 16, no. 1: 58. https://doi.org/10.3390/toxins16010058
APA StyleLee, H. J., Kim, H. D., & Ryu, D. (2024). Practical Strategies to Reduce Ochratoxin A in Foods. Toxins, 16(1), 58. https://doi.org/10.3390/toxins16010058