Hidden Hazards Revealed: Mycotoxins and Their Masked Forms in Poultry



Abstract
:1. Introduction
2. Masked Mycotoxins
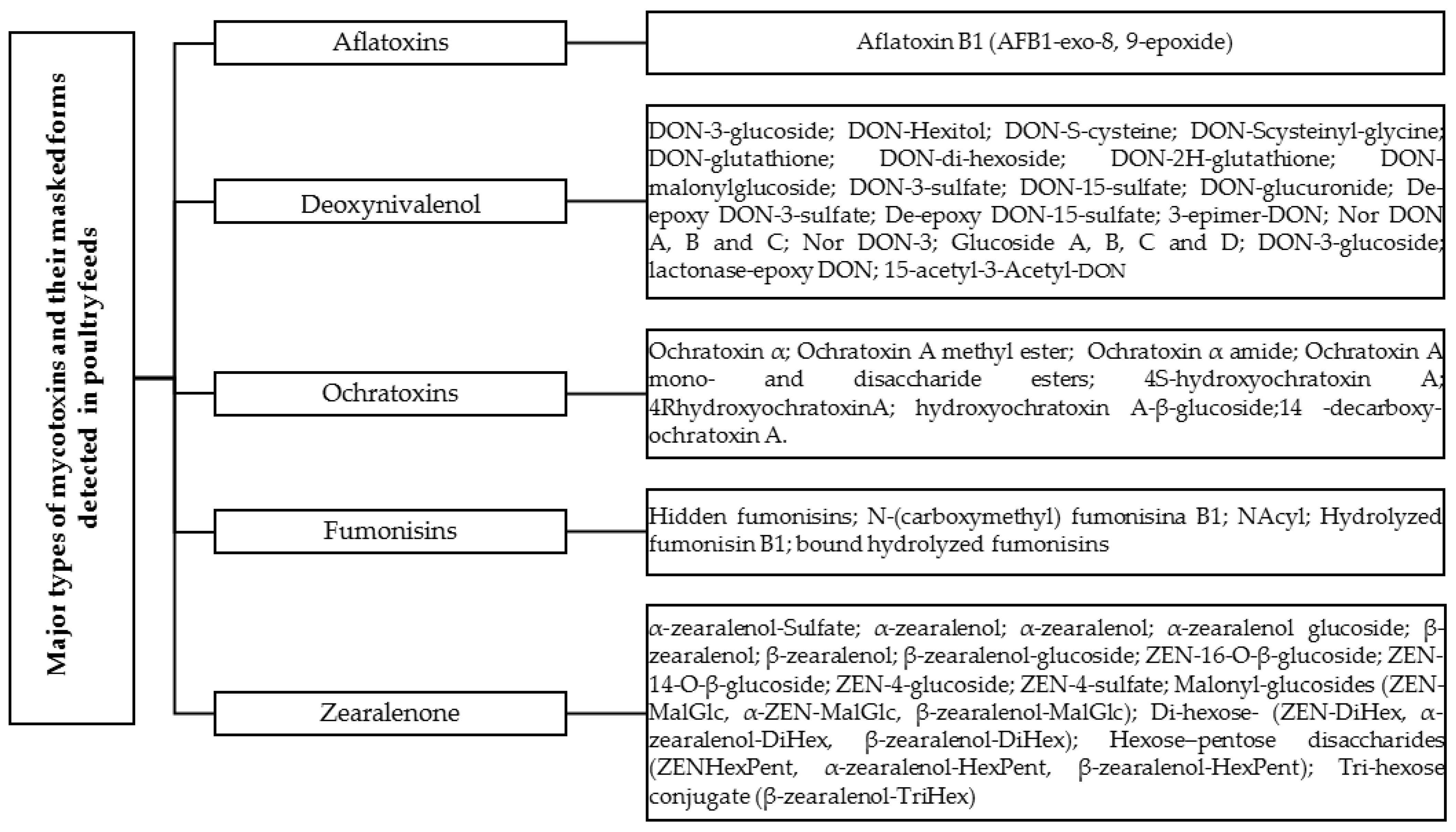
3. Major Types of Mycotoxins and Their Masked Forms in Chickens
3.1. Aflatoxin
3.2. Deoxynivalenol (DON)
3.3. Ochratoxins
3.4. Fumonisins
3.5. Zearalenone
4. Approaches to Mitigate Mycotoxins in Poultry Diets
Mitigating Masked Mycotoxins in Poultry Feed Can Be Achieved through the Use of Specific Feed Additives: Here Are Some Strategies to Consider
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Patil, R.D.; Sharma, R.; Asrani, R.K. Mycotoxicosis and its control in poultry: A review. J. Poult. Sci. Technol. 2014, 2, 1–10. [Google Scholar]
- Karlovsky, P.; Suman, M.; Berthiller, F.; De Meester, J.; Eisenbrand, G.; Perrin, I.; Oswald, I.P.; Speijers, G.; Chiodini, A.; Recker, T.; et al. Impact of food processing and detoxification treatments on mycotoxin contamination. Mycotoxin Res. 2016, 32, 179–205. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Zhang, L.; Liu, M.; Su, Y.-T.; Xie, W.-M.; Zhang, N.-Y.; Dai, J.-F.; Wang, Y.; Rajput, S.A.; Qi, D.-S. Individual and combined occurrence of mycotoxins in feed ingredients and complete feeds in China. Toxins 2018, 10, 113. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, L.; Xu, Z.; Liu, X.; Chen, L.; Dai, J.; Karrow, N.A.; Sun, L. Occurrence of Aflatoxin B1, deoxynivalenol and zearalenone in feeds in China during 2018–2020. J. Anim. Sci. Biotechnol. 2021, 12, 74. [Google Scholar] [CrossRef]
- Ostry, V.; Malir, F.; Toman, J.; Grosse, Y. Mycotoxins as human carcinogens—The IARC Monographs classification. Mycotoxin Res. 2017, 33, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Rushing, B.R.; Selim, M.I. Aflatoxin B1: A review on metabolism, toxicity, occurrence in food, occupational exposure, and detoxification methods. Food Chem. Toxicol. 2019, 124, 81–100. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Qin, X.; Tang, Y.; Ma, Q.; Zhang, J.; Zhao, L. CotA laccase, a novel aflatoxin oxidase from Bacillus licheniformis, transforms aflatoxin B1 to aflatoxin Q1 and epi-aflatoxin Q1. Food Chem. 2020, 325, 126877. [Google Scholar] [CrossRef]
- Fan, Y.; Zhao, L.; Ji, C.; Li, X.; Jia, R.; Xi, L.; Zhang, J.; Ma, Q. Protective effects of Bacillus subtilis ANSB060 on serum biochemistry, histopathological changes and antioxidant enzyme activities of broilers fed moldy peanut meal naturally contaminated with aflatoxins. Toxins 2015, 7, 3330–3343. [Google Scholar] [CrossRef]
- Kumar, P.; Mahato, D.K.; Kamle, M.; Mohanta, T.K.; Kang, S.G. Aflatoxins: A global concern for food safety, human health and their management. Front. Microbiol. 2017, 7, 2170. [Google Scholar] [CrossRef]
- Chang, J.; Wang, T.; Wang, P.; Yin, Q.; Liu, C.; Zhu, Q.; Lu, F.; Gao, T. Compound probiotics alleviating aflatoxin B(1) and zearalenone toxic effects on broiler production performance and gut microbiota. Ecotoxicol. Environ. Saf. 2020, 194, 110420. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed]
- Tsiouris, V.; Tassis, P.; Raj, J.; Mantzios, T.; Kiskinis, K.; Vasiljević, M.; Delić, N.; Petridou, E.; Brellou, G.D.; Polizopoulou, Z.; et al. Investigation of a Novel Multicomponent Mycotoxin Detoxifying Agent in Amelioration of Mycotoxicosis Induced by Aflatoxin-B1 and Ochratoxin A in Broiler Chicks. Toxins 2021, 13, 367. [Google Scholar] [CrossRef]
- Berthiller, F.; Crews, C.; Dall’Asta, C.; Saeger, S.D.; Haesaert, G.; Karlovsky, P.; Oswald, I.P.; Seefelder, W.; Speijers, G.; Stroka, J. Masked mycotoxins: A review. Mol. Nutr. Food Res. 2013, 57, 165–186. [Google Scholar] [CrossRef] [PubMed]
- Kovalsky Paris, M.P.; Schweiger, W.; Hametner, C.; Stückler, R.; Muehlbauer, G.J.; Varga, E.; Krska, R.; Berthiller, F.; Adam, G. Zearalenone-16-O-glucoside: A new masked mycotoxin. J. Agric. Food Chem. 2014, 62, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Dall’Erta, A.; Cirlini, M.; Dall’Asta, M.; Del Rio, D.; Galaverna, G.; Dall’Asta, C. Masked mycotoxins are efficiently hydrolyzed by human colonic microbiota releasing their aglycones. Chem. Res. Toxicol. 2013, 26, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Riahi, I.; Ramos, A.J.; Raj, J.; Jakovčević, Z.; Farkaš, H.; Vasiljević, M.; Pérez-Vendrell, A.M. Effect of a mycotoxin binder (MMDA) on the growth performance, blood and carcass characteristics of broilers fed ochratoxin a and t-2 mycotoxin contaminated diets. Animals 2021, 11, 3205. [Google Scholar] [CrossRef]
- Wu, K.; Ren, C.; Gong, Y.; Gao, X.; Rajput, S.A.; Qi, D.; Wang, S. The insensitive mechanism of poultry to zearalenone: A review. Anim. Nutr. 2021, 7, 587–594. [Google Scholar] [CrossRef]
- Rychlik, M.; Humpf, H.U.; Marko, D.; Dänicke, S.; Mally, A.; Berthiller, F.; Klaffke, H.; Lorenz, N. Proposal of a comprehensive definition of modified and other forms of mycotoxins including “masked” mycotoxins. Mycotoxin Res. 2014, 30, 197–205. [Google Scholar] [CrossRef]
- Gareis, M.; Bauer, J.; Thiem, J.; Plank, G.; Grabley, S.; Gedek, B. Cleavage of zearalenone-glycoside, a “masked” mycotoxin, during digestion in swine. Zentralblatt Veterinarmed. Reihe B 1990, 37, 236–240. [Google Scholar] [CrossRef]
- Zhang, Z.; Nie, D.; Fan, K.; Yang, J.; Guo, W.; Meng, J.; Zhao, Z.; Han, Z. A systematic review of plant-conjugated masked mycotoxins: Occurrence, toxicology, and metabolism. Crit. Rev. Food Sci. Nutr. 2020, 60, 1523–1537. [Google Scholar] [CrossRef]
- Kim, J.H.; Park, G.H.; Han, G.P.; Kil, D.Y. Effect of feeding corn distillers dried grains with solubles naturally contaminated with deoxynivalenol on growth performance, meat quality, intestinal permeability, and utilization of energy and nutrients in broiler chickens. Poult. Sci. 2021, 100, 101215. [Google Scholar] [CrossRef] [PubMed]
- Nabizadeh, S.; Shariatifar, N.; Shokoohi, E.; Shoeibi, S.; Gavahian, M.; Fakhri, Y.; Azari, A.; Mousavi Khaneghah, A. Prevalence and probabilistic health risk assessment of aflatoxins B1, B2, G1, and G2 in Iranian edible oils. Environ. Sci. Pollut. Res. 2018, 25, 35562–35570. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, J.; Alipour, S.H.D.; Miri, A.; Fakhri, Y.; Riahi, S.M.; Keramati, H.; Moradi, M.; Amanidaz, N.; Pouya, R.H.; Bahmani, Z.; et al. The prevalence of aflatoxin M1 in milk of Middle East region: A systematic review, meta-analysis and probabilistic health risk assessment. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2018, 118, 653–666. [Google Scholar] [CrossRef] [PubMed]
- Freire, L.; Sant’Ana, A.S. Modified mycotoxins: An updated review on their formation, detection, occurrence, and toxic effects. Food Chem. Toxicol. 2018, 111, 189–205. [Google Scholar] [CrossRef] [PubMed]
- Freire, L.; Guerreiro, T.M.; Caramês, E.T.; Lopes, L.S.; Orlando, E.A.; Pereira, G.E.; Lima Pallone, J.A.; Catharino, R.R.; Sant’Ana, A.S. Influence of maturation stages in different varieties of wine grapes (Vitis vinifera) on the production of ochratoxin A and its modified forms by Aspergillus carbonarius and Aspergillus niger. J. Agric. Food Chem. 2018, 66, 8824–8831. [Google Scholar] [CrossRef] [PubMed]
- Kępińska-Pacelik, J.; Biel, W. Alimentary risk of mycotoxins for humans and animals. Toxins 2021, 13, 822. [Google Scholar] [CrossRef] [PubMed]
- Scarpino, V.; Vanara, F.; Sulyok, M.; Krska, R.; Blandino, M. Fate of regulated, masked, emerging mycotoxins and secondary fungal metabolites during different large-scale maize dry-milling processes. Food Res. Int. 2021, 140, 109861. [Google Scholar] [CrossRef]
- Liu, J.D.; Doupovec, B.; Schatzmayr, D.; Murugesan, G.R.; Bortoluzzi, C.; Villegas, A.M.; Applegate, T.J. The impact of deoxynivalenol, fumonisins, and their combination on performance, nutrient, and energy digestibility in broiler chickens. Poult. Sci. 2020, 99, 272–279. [Google Scholar] [CrossRef]
- Palumbo, R.; Crisci, A.; Venâncio, A.; Cortiñas Abrahantes, J.; Dorne, J.L.; Battilani, P.; Toscano, P. Occurrence and Co-Occurrence of Mycotoxins in Cereal-Based Feed and Food. Microorganisms 2020, 8, 74. [Google Scholar] [CrossRef]
- Ochieng, P.E.; Scippo, M.L.; Kemboi, D.C.; Croubels, S.; Okoth, S.; Kang’ethe, E.K.; Doupovec, B.; Gathumbi, J.K.; Lindahl, J.F.; Antonissen, G. Mycotoxins in Poultry Feed and Feed Ingredients from Sub-Saharan Africa and Their Impact on the Production of Broiler and Layer Chickens: A Review. Toxins 2021, 13, 633. [Google Scholar] [CrossRef]
- Hernández-Ramírez, J.O.; Nava-Ramírez, M.J.; Merino-Guzmán, R.; Téllez-Isaías, G.; Vázquez-Durán, A.; Méndez-Albores, A. The effect of moderate-dose aflatoxin B(1) and Salmonella Enteritidis infection on intestinal permeability in broiler chickens. Mycotoxin Res. 2020, 36, 31–39. [Google Scholar] [CrossRef]
- Saleemi, M.K.; Ashraf, K.; Gul, S.T.; Naseem, M.N.; Sajid, M.S.; Mohsin, M.; He, C.; Zubair, M.; Khan, A. Toxicopathological effects of feeding aflatoxins B1 in broilers and its ameliosration with indigenous mycotoxin binder. Ecotoxicol. Environ. Saf. 2020, 187, 109712. [Google Scholar] [CrossRef]
- Antonissen, G.; De Baere, S.; Novak, B.; Schatzmayr, D.; den Hollander, D.; Devreese, M.; Croubels, S. Toxicokinetics of Hydrolyzed Fumonisin B1 after Single Oral or Intravenous Bolus to Broiler Chickens Fed a Control or a Fumonisins-Contaminated Diet. Toxins 2020, 12, 413. [Google Scholar] [CrossRef]
- Peillod, C.; Laborde, M.; Travel, A.; Mika, A.; Bailly, J.; Cleva, D.; Boissieu, C.; Le Guennec, J.; Albaric, O.; Labrut, S.; et al. Toxic Effects of Fumonisins, Deoxynivalenol and Zearalenone Alone and in Combination in Ducks Fed the Maximum EUTolerated Level. Toxins 2021, 13, 152. [Google Scholar] [CrossRef]
- Grenier, B.; Dohnal, I.; Shanmugasundaram, R.; Eicher, S.; Selvaraj, R.K.; Schatzmayr, G.; Applegate, T.J. Susceptibility of Broiler Chickens to Coccidiosis When Fed Subclinical Doses of Deoxynivalenol and Fumonisins—Special Emphasis on the Immunological Response and the Mycotoxin Interaction. Toxins 2016, 8, 231. [Google Scholar] [CrossRef]
- Haque, M.A.; Wang, Y.; Shen, Z.; Li, X.; Saleemi, M.K.; He, C. Mycotoxin contamination and control strategy in human, domestic animal and poultry: A review. Microb. Pathog. 2020, 142, 104095. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, X.; Li, Q. Aflatoxin B1 in poultry liver: Toxic mechanism. Toxicon 2023, 107262. [Google Scholar] [CrossRef]
- Motbaynor, A.; Kassaye, D.; Keffale, M.; Wasihun, P. Magnitude of Aflatoxigenic Aspergillus Species, Level of Aflatoxin B1, and Associated Factors in Stored Feed at Poultry Farms in Dire Dawa, Ethiopia. Vet. Med. Int. 2021, 2021, 6638083. [Google Scholar] [CrossRef]
- Gbashi, S.; Madala, N.E.; Saeger, S.D.; Boevre, M.D.; Adekoya, I.; Adebo, O.A.; Njobeh, P.B. The Socio-Economic Impact of Mycotoxin Contamination in Africa. In Mycotoxins—Impact and Management Strategies; IntechOpen: London, UK, 2018. [Google Scholar]
- Rashidi, N.; Khatibjoo, A.; Taherpour, K.; Akbari-Gharaei, M.; Shirzadi, H. Effects of licorice extract, probiotic, toxin binder and poultry litter biochar on performance, immune function, blood indices and liver histopathology of broilers exposed to aflatoxin-B1. Poult. Sci. 2020, 99, 5896–5906. [Google Scholar] [CrossRef]
- Zhang, J.; Fang, Y.; Fu, Y.; Jalukar, S.; Ma, J.; Liu, Y.; Guo, Y.; Ma, Q.; Ji, C.; Zhao, L. Yeast polysaccharide mitigated oxidative injury in broilers induced by mixed mycotoxins via regulating intestinal mucosal oxidative stress and hepatic metabolic enzymes. Poult. Sci. 2023, 102, 102862. [Google Scholar] [CrossRef]
- Magnoli, A.P.; Rodriguez, M.; Gonzalez Pereyra, M.L.; Poloni, V.L.; Peralta, M.; Nilson, A.; Miazzo, R.D.; Bagnis, G.; Chiacchiera, S.M.; Cavaglieri, L. Use of yeast (Pichia kudriavzevii) as a novel feed additive to ameliorate the effects of aflatoxin B 1 on broiler chicken performance. Mycotoxin Res. 2017, 33, 273–283. [Google Scholar] [CrossRef]
- Mgbeahuruike, A.C.; Ejiofor, T.E.; Ashang, M.U.; Ojiako, C.; Obasi, C.C.; Ezema, C.; Okoroafor, O.; Mwanza, M.; Karlsson, M.; Chah, K.F. Reduction of the adverse impacts of fungal mycotoxin on proximate composition of feed and growth performance in broilers by combined adsorbents. Toxins 2021, 13, 430. [Google Scholar] [CrossRef]
- Aikore, M.O.S.; Ortega-Beltran, A.; Eruvbetine, D.; Atehnkeng, J.; Falade, T.D.O.; Cotty, P.J.; Bandyopadhyay, R. Performance of Broilers Fed with Maize Colonized by Either Toxigenic or Atoxigenic Strains of Aspergillus flavus with and without an Aflatoxin-Sequestering Agent. Toxins 2019, 11, 565. [Google Scholar] [CrossRef]
- Rosa, C.A.; Miazzo, R.; Magnoli, C.; Salvano, M.; Chiacchiera, S.M.; Ferrero, S.; Saenz, M.; Carvalho, E.C.; Dalcero, A. Evaluation of the efficacy of bentonite from the south of Argentina to ameliorate the toxic effects of aflatoxin in broilers. Poult. Sci. 2001, 80, 139–144. [Google Scholar] [CrossRef]
- Miazzo, R.D.; Peralta, M.F.; Magnoli, C.E.; Salvano, M.A.; Ferrero, S.; Chiacchiera, S.M.; Carvalho, E.; Rosa, C.A.R.; Dalcero, A.M. Efficacy of sodium bentonite as a detoxifier of broiler feed contaminated with aflatoxin and fumonisin. Poult. Sci. 2005, 84, 1–8. [Google Scholar] [CrossRef]
- Zaghini, A.; Martelli, G.; Roncada, P.; Simioli, M.; Rizzi, L. Mannanoligosaccharides and aflatoxin B1 in feed for laying hens: Effects on egg quality, aflatoxins B1 and M1 residues in eggs, and aflatoxin B1 levels in liver. Poult. Sci. 2005, 84, 825–832. [Google Scholar] [CrossRef]
- Bintvihok, A.; Kositcharoenkul, S. Effect of dietary calcium propionate on performance, hepatic enzyme activities and aflatoxin residues in broilers fed a diet containing low levels of aflatoxin B1. Toxicon 2006, 47, 41–46. [Google Scholar] [CrossRef]
- Tessari, E.N.; Oliveira, C.A.; Cardoso, A.L.; Ledoux, D.R.; Rottinghaus, G.E. Effects of aflatoxin B1 and fumonisin B1 on body weight, antibody titres and histology of broiler chicks. Br. Poult. Sci. 2006, 47, 357–364. [Google Scholar] [CrossRef]
- Balogh, K.; Kövesi, B.; Zándoki, E.; Kulcsár, S.; Ancsin, Z.; Erdélyi, M.; Dobolyi, C.; Bata-Vidács, I.; Inotai, K.; Szekeres, A.; et al. Effect of Sterigmatocystin or Aflatoxin Contaminated Feed on Lipid Peroxidation and Glutathione Redox System and Expression of Glutathione Redox System Regulatory Genes in Broiler Chicken. Antioxidants 2019, 8, 201. [Google Scholar] [CrossRef]
- Rauber, R.H.; Dilkin, P.; Mallmann, A.O.; Marchioro, A.; Mallmann, C.A.; Borsoi, A.; Nascimento, V.P. Individual and combined effects of Salmonella typhimurium lipopolysaccharide and fumonisin B1 in broiler chickens. Poult. Sci. 2012, 91, 2785–2791. [Google Scholar] [CrossRef]
- Muhammad, D.; Chand, N.; Khan, S.; Sultan, A.; Mushtaq, M. Hepatoprotective Role of Milk Thistle (Silybum marianum) in Meat Type Chicken Fed Aflatoxin B1 Contaminated Feed. Pak. Vet. J. 2012, 32, 443. [Google Scholar]
- Fani Makki, O.; Afzali, N.; Omidi, A. Effect of milk thistle on the immune system, intestinal related variables, appearance and mortality of broilers contaminated with Aflatoxin B1. J. Med. Herbs 2013, 4, 33–38. [Google Scholar]
- Dos Anjos, F.R.; Ledoux, D.R.; Rottinghaus, G.E.; Chimonyo, M. Efficacy of adsorbents (bentonite and diatomaceous earth) and turmeric (Curcuma longa) in alleviating the toxic effects of aflatoxin in chicks. Br. Poult. Sci. 2015, 56, 459–469. [Google Scholar] [CrossRef]
- Jahanian, E.; Mahdavi, A.; Asgary, S.; Jahanian, R. Effects of dietary inclusion of silymarin on performance, intestinal morphology and ileal bacterial count in aflatoxin—Challenged broiler chicks. J. Anim. Physiol. Anim. Nutr. 2017, 101, e43–e54. [Google Scholar] [CrossRef] [PubMed]
- Afshin, M.; Afzali, N.; Mojtahedi, M.; Mohammadi, A. Effects of milk thistle seeds, whole plant and extract on performance, carcass characteristics and some blood parameters of broiler chickens fed aflatoxin B1 contaminated diet. Anim. Prod. 2017, 19, 403–414. [Google Scholar] [CrossRef]
- Feshanghchi, M.; Baghban-Kanani, P.; Kashefi-Motlagh, B.; Adib, F.; Azimi-Youvalari, S.; Hosseintabar-Ghasemabad, B.; Slozhenkina, M.; Gorlov, I.; Zangeronimo, M.G.; Swelum, A.A. Milk Thistle (Silybum marianum), Marine Algae (Spirulina platensis) and toxin binder powders in the diets of broiler chickens exposed to aflatoxin-B1: Growth performance, humoral immune response and cecal microbiota. Agriculture 2022, 12, 805. [Google Scholar] [CrossRef]
- Eraslan, G.; Akdoğan, M.; Yarsan, E.; Şahindokuyucu, F.; Eşsiz, D.; Altintaş, L. The effects of aflatoxins on oxidative stress in broiler chickens. Turk. J. Vet. Anim. Sci. 2005, 29, 701–707. [Google Scholar]
- Borutova, R.; Faix, S.; Placha, I.; Gresakova, L.; Cobanova, K.; Leng, L. Effects of deoxynivalenol and zearalenone on oxidative stress and blood phagocytic activity in broilers. Arch. Anim. Nutr. 2008, 62, 303–312. [Google Scholar] [CrossRef]
- Osselaere, A.; Santos, R.; Hautekiet, V.; De Backer, P.; Chiers, K.; Ducatelle, R.; Croubels, S. Deoxynivalenol impairs hepatic and intestinal gene expression of selected oxidative stress, tight junction and inflammation proteins in broiler chickens, but addition of an adsorbing agent shifts the effects to the distal parts of the small intestine. PLoS ONE 2013, 8, e69014. [Google Scholar] [CrossRef]
- Sridhar, M.; Suganthi, R.U.; Thammiaha, V. Effect of dietary resveratrol in ameliorating aflatoxin B1-induced changes in broiler birds. J. Anim. Physiol. Anim. Nutr. 2015, 99, 1094–1104. [Google Scholar] [CrossRef]
- Chen, J.; Chen, K.; Yuan, S.; Peng, X.; Fang, J.; Wang, F.; Cui, H.; Chen, Z.; Yuan, J.; Geng, Y. Effects of aflatoxin B1 on oxidative stress markers and apoptosis of spleens in broilers. Toxicol. Ind. Health 2016, 32, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Rajput, S.A.; Sun, L.; Zhang, N.; Khalil, M.M.; Gao, X.; Ling, Z.; Zhu, L.; Khan, F.A.; Zhang, J.; Qi, D. Ameliorative Effects of Grape Seed Proanthocyanidin Extract on Growth Performance, Immune Function, Antioxidant Capacity, Biochemical Constituents, Liver Histopathology and Aflatoxin Residues in Broilers Exposed to Aflatoxin B₁. Toxins 2017, 9, 371, Correction in Toxins 2018, 10, 366. [Google Scholar] [CrossRef] [PubMed]
- Guerrini, A.; Tedesco, D.E.A. Restoring Activity of Milk Thistle (Silybum marianum L.) on Serum Biochemical Parameters, Oxidative Status, Immunity, and Performance in Poultry and Other Animal Species, Poisoned by Mycotoxins: A Review. Animals 2023, 13, 330. [Google Scholar] [CrossRef] [PubMed]
- Tedesco, D.; Steidler, S.; Galletti, S.; Tameni, M.; Sonzogni, O.; Ravarotto, L. Efficacy of silymarin-phospholipid complex in reducing the toxicity of aflatoxin B1 in broiler chicks. Poult. Sci. 2004, 83, 1839–1843. [Google Scholar] [CrossRef] [PubMed]
- Fani Makki, O.; Omidi, A.; Afzali, N.; Sarir, H.; Frouzanmehr, M.; Shibak, A. Efficacy of Silybum marianum seeds in ameliorating the toxic effects of aflatoxin B1 in broilers. Iran. J. Toxicol. 2014, 8, 977–982. [Google Scholar]
- Malekinejad, P.; Afzali, N.; Mohammadi, A.; Sarir, H. Protective effects of milk thistle (Silybum marianum) seeds and sodium bentonite in ameliorating the toxic effects of aflatoxin b1 in broiler chicks. Arch. Med. Lab. Sci. 2015, 1. [Google Scholar]
- Amiridumari, H.; Sarir, H.; Afzali, N.; FaniMakki, O. Effects of milk thistle seed against aflatoxin B1 in broiler model. J. Res. Med. Sci. Off. J. Isfahan Univ. Med. Sci. 2013, 18, 786. [Google Scholar]
- Chand, N.; Muhammad, D.; Durrani, F.; Qureshi, M.S.; Ullah, S.S. Protective effects of milk thistle (Silybum marianum) against aflatoxin B1 in broiler chicks. Asian-Australas. J. Anim. Sci. 2011, 24, 1011–1018. [Google Scholar] [CrossRef]
- Akinmusire, O.O.; El-Yuguda, A.D.; Musa, J.A.; Oyedele, O.A.; Sulyok, M.; Somorin, Y.M.; Ezekiel, C.N.; Krska, R. Mycotoxins in poultry feed and feed ingredients in Nigeria. Mycotoxin Res. 2019, 35, 149–155. [Google Scholar] [CrossRef]
- Naseem, M.N.; Saleemi, M.K.; Khan, A.; Khatoon, A.; Gul, S.T.; Rizvi, F.; Ahmad, I.; Fayyaz, A. Pathological effects of concurrent administration of aflatoxin B(1) and fowl adenovirus-4 in broiler chicks. Microb. Pathog. 2018, 121, 147–154. [Google Scholar] [CrossRef]
- Cravens, R.L.; Goss, G.R.; Chi, F.; De Boer, E.D.; Davis, S.W.; Hendrix, S.M.; Richardson, J.A.; Johnston, S.L. The effects of necrotic enteritis, aflatoxin B1, and virginiamycin on growth performance, necrotic enteritis lesion scores, and mortality in young broilers. Poult. Sci. 2013, 92, 1997–2004. [Google Scholar] [CrossRef]
- Mavrommatis, A.; Giamouri, E.; Tavrizelou, S.; Zacharioudaki, M.; Danezis, G.; Simitzis, P.E.; Zoidis, E.; Tsiplakou, E.; Pappas, A.C.; Georgiou, C.A. Impact of mycotoxins on animals’ oxidative status. Antioxidants 2021, 10, 214. [Google Scholar] [CrossRef] [PubMed]
- Oskoueian, E.; Abdullah, N.; Zulkifli, I.; Ebrahimi, M.; Karimi, E.; Goh, Y.M.; Oskoueian, A.; Shakeri, M. Cytoprotective effect of palm kernel cake phenolics against aflatoxin B1-induced cell damage and its underlying mechanism of action. BMC Complement. Altern. Med. 2015, 15, 392. [Google Scholar] [CrossRef] [PubMed]
- Saminathan, M.; Selamat, J.; Abbasi Pirouz, A.; Abdullah, N.; Zulkifli, I. Effects of Nano-Composite Adsorbents on the Growth Performance, Serum Biochemistry, and Organ Weights of Broilers Fed with Aflatoxin-Contaminated Feed. Toxins 2018, 10, 345. [Google Scholar] [CrossRef] [PubMed]
- Shannon, T.A.; Ledoux, D.R.; Rottinghaus, G.E.; Shaw, D.P.; Dakovic, A.; Markovic, M. The efficacy of raw and concentrated bentonite clay in reducing the toxic effects of aflatoxin in broiler chicks. Poult. Sci. 2017, 96, 1651–1658. [Google Scholar] [CrossRef] [PubMed]
- Kalpana, S.; Aggarwal, M.; Rao, G.S.; Malik, J.K. Effects of aflatoxin B1 on tissue residues of enrofloxacin and its metabolite ciprofloxacin in broiler chickens. Environ. Toxicol. Pharmacol. 2012, 33, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Guan, K.; Zuo, Z.; Wang, F.; Peng, X.; Fang, J.; Cui, H.; Zhou, Y.; Ouyang, P.; Su, G. Effects of aflatoxin B1 on the cell cycle distribution of splenocytes in chickens. J. Toxicol. Pathol. 2019, 32, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Bahari, A.; Mehrzad, J.; Mahmoudi, M.; Bassami, M.R.; Dehghani, H. Cytochrome P450 isoforms are differently up-regulated in aflatoxin B1-exposed human lymphocytes and monocytes. Immunopharmacol. Immunotoxicol. 2014, 36, 1–10. [Google Scholar] [CrossRef]
- Yin, H.; Jiang, M.; Peng, X.; Cui, H.; Zhou, Y.; He, M.; Zuo, Z.; Ouyang, P.; Fan, J.; Fang, J. The molecular mechanism of G2/M cell cycle arrest induced by AFB1 in the jejunum. Oncotarget 2016, 7, 35592. [Google Scholar] [CrossRef]
- Yu, Z.; Wang, F.; Liang, N.; Wang, C.; Peng, X.; Fang, J.; Cui, H.; Jameel Mughal, M.; Lai, W. Effect of selenium supplementation on apoptosis and cell cycle blockage of renal cells in broilers fed a diet containing aflatoxin B1. Biol. Trace Elem. Res. 2015, 168, 242–251. [Google Scholar] [CrossRef]
- Bbosa, G.S.; Kitya, D.; Odda, J.; Ogwal-Okeng, J. Aflatoxins metabolism, effects on epigenetic mechanisms and their role in carcinogenesis. Health 2013, 5, 14–34. [Google Scholar] [CrossRef]
- Murugesan, G.R.; Ledoux, D.R.; Naehrer, K.; Berthiller, F.; Applegate, T.J.; Grenier, B.; Phillips, T.D.; Schatzmayr, G. Prevalence and effects of mycotoxins on poultry health and performance, and recent development in mycotoxin counteracting strategies. Poult. Sci. 2015, 94, 1298–1315. [Google Scholar] [CrossRef]
- Zhang, Y.; Jia, Z.; Yin, S.; Shan, A.; Gao, R.; Qu, Z.; Liu, M.; Nie, S. Toxic effects of maternal zearalenone exposure on uterine capacity and fetal development in gestation rats. Reprod. Sci. 2014, 21, 743–753. [Google Scholar] [CrossRef]
- Yilmaz, S.; Bag, H. Aflatoxin B1: Mechanism, Oxidative Stress and Effects on Animal Health. J. Anim. Biol. Vet. 2022, 2, 1–16. [Google Scholar]
- Fouad, A.M.; Ruan, D.; El-Senousey, H.K.; Chen, W.; Jiang, S.; Zheng, C. Harmful effects and control strategies of aflatoxin b1 produced by Aspergillus flavus and Aspergillus parasiticus strains on poultry. Toxins 2019, 11, 176. [Google Scholar] [CrossRef]
- Yilmaz, S.; Kaya, E.; Kisacam, M.A. The effect on oxidative stress of aflatoxin and protective effect of lycopene on aflatoxin damage. Aflatoxin-Control Anal. Detect. Health Risks 2017, 30, 67–90. [Google Scholar]
- Richard, E.; Heutte, N.; Sage, L.; Pottier, D.; Bouchart, V.; Lebailly, P.; Garon, D. Toxigenic fungi and mycotoxins in mature corn silage. Food Chem. Toxicol. 2007, 45, 2420–2425. [Google Scholar] [CrossRef] [PubMed]
- Pestka, J.J. Deoxynivalenol: Mechanisms of action, human exposure, and toxicological relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Kolawole, O.; Graham, A.; Donaldson, C.; Owens, B.; Abia, W.A.; Meneely, J.P.; Alcorn, M.J.; Connolly, L.; Elliott, C.T. Low Doses of Mycotoxin Mixtures below EU Regulatory Limits Can Negatively Affect the Performance of Broiler Chickens: A Longitudinal Study. Toxins 2020, 12, 433. [Google Scholar] [CrossRef]
- Vidal, A.; Sanchis, V.; Ramos, A.J.; Marín, S. The fate of deoxynivalenol through wheat processing to food products. Curr. Opin. Food Sci. 2016, 11, 34–39. [Google Scholar] [CrossRef]
- Metayer, J.-P.; Travel, A.; Mika, A.; Bailly, J.-D.; Cleva, D.; Boissieu, C.; Guennec, J.L.; Froment, P.; Albaric, O.; Labrut, S.; et al. Lack of Toxic Interaction between Fusariotoxins in Broiler Chickens Fed throughout Their Life at the Highest Level Tolerated in the European Union. Toxins 2019, 11, 455. [Google Scholar] [CrossRef]
- Wellington, M.O.; Bosompem, M.A.; Petracek, R.; Nagl, V.; Columbus, D.A. Effect of long-term feeding of graded levels of deoxynivalenol (DON) on growth performance, nutrient utilization, and organ health in finishing pigs and DON content in biological samples. J. Anim. Sci. 2020, 98, skaa378. [Google Scholar] [CrossRef]
- Lucke, A.; Doupovec, B.; Paulsen, P.; Zebeli, Q.; Böhm, J. Effects of low to moderate levels of deoxynivalenol on feed and water intake, weight gain, and slaughtering traits of broiler chickens. Mycotoxin Res. 2017, 33, 261–271. [Google Scholar] [CrossRef]
- Keçi, M.; Lucke, A.; Paulsen, P.; Zebeli, Q.; Böhm, J.; Metzler-Zebeli, B.U. Deoxynivalenol in the diet impairs bone mineralization in broiler chickens. Toxins 2019, 11, 352. [Google Scholar] [CrossRef]
- Ghareeb, K.; Awad, W.A.; Böhm, J.; Zebeli, Q. Impacts of the feed contaminant deoxynivalenol on the intestine of monogastric animals: Poultry and swine. J. Appl. Toxicol. 2015, 35, 327–337. [Google Scholar] [CrossRef]
- Yunus, A.W.; Ghareeb, K.; Twaruzek, M.; Grajewski, J.; Böhm, J. Deoxynivalenol as a contaminant of broiler feed: Effects on bird performance and response to common vaccines. Poult. Sci. 2012, 91, 844–851. [Google Scholar] [CrossRef] [PubMed]
- Riahi, I.; Marquis, V.; Ramos, A.J.; Brufau, J.; Esteve-Garcia, E.; Pérez-Vendrell, A.M. Effects of Deoxynivalenol-Contaminated Diets on Productive, Morphological, and Physiological Indicators in Broiler Chickens. Animals 2020, 10, 1795. [Google Scholar] [CrossRef]
- Awad, W.A.; Ghareeb, K.; Dadak, A.; Hess, M.; Böhm, J. Single and combined effects of deoxynivalenol mycotoxin and a microbial feed additive on lymphocyte DNA damage and oxidative stress in broiler chickens. PLoS ONE 2014, 9, e88028. [Google Scholar] [CrossRef]
- Ghareeb, K.; Awad, W.; Zebeli, Q.; Böhm, J. Deoxynivalenol in chicken feed alters the vaccinal immune response and clinical biochemical serum parameters but not the intestinal and carcass characteristics. J. Anim. Physiol. Anim. Nutr. 2016, 100, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Antonissen, G.; Martel, A.; Pasmans, F.; Ducatelle, R.; Verbrugghe, E.; Vandenbroucke, V.; Li, S.; Haesebrouck, F.; Van Immerseel, F.; Croubels, S. The impact of Fusarium mycotoxins on human and animal host susceptibility to infectious diseases. Toxins 2014, 6, 430–452. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.S.; Li, Y.-H.; Lin, M.-F. Chronic exposure to the Fusarium mycotoxin deoxynivalenol: Impact on performance, immune organ, and intestinal integrity of slow-growing chickens. Toxins 2017, 9, 334. [Google Scholar] [CrossRef]
- Girish, C.K.; Smith, T.K.; Boermans, H.J.; Karrow, N.A. Effects of feeding blends of grains naturally contaminated with Fusarium mycotoxins on performance, hematology, metabolism, and immunocompetence of turkeys. Poult. Sci. 2008, 87, 421–432. [Google Scholar] [CrossRef]
- Dänicke, S.; Matthes, S.; Halle, I.; Ueberschär, K.H.; Döll, S.; Valenta, H. Effects of graded levels of Fusarium toxin-contaminated wheat and of a detoxifying agent in broiler diets on performance, nutrient digestibility and blood chemical parameters. Br. Poult. Sci. 2003, 44, 113–126. [Google Scholar] [CrossRef]
- Swamy, H.V.; Smith, T.K.; Karrow, N.A.; Boermans, H.J. Effects of feeding blends of grains naturally contaminated with Fusarium mycotoxins on growth and immunological parameters of broiler chickens. Poult. Sci. 2004, 83, 533–543. [Google Scholar] [CrossRef]
- Awad, W.; Böhm, J.; Razzazi-Fazeli, E.; Zentek, J. Effects of feeding deoxynivalenol contaminated wheat on growth performance, organ weights and histological parameters of the intestine of broiler chickens. J. Anim. Physiol. Anim. Nutr. 2006, 90, 32–37. [Google Scholar] [CrossRef]
- Dänicke, S.; Valenta, H.; Ueberschär, K.-H.; Matthes, S. On the interactions between Fusarium toxin-contaminated wheat and non-starch-polysaccharide hydrolysing enzymes in turkey diets on performance, health and carry-over of deoxynivalenol and zearalenone. Br. Poult. Sci. 2007, 48, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Eicher, S.D.; Applegate, T.J. Effects of increasing dietary concentrations of corn naturally contaminated with deoxynivalenol on broiler and turkey poult performance and response to lipopolysaccharide. Poult. Sci. 2011, 90, 2766–2774. [Google Scholar] [CrossRef] [PubMed]
- Awad, W.A.; Hess, M.; Twarużek, M.; Grajewski, J.; Kosicki, R.; Böhm, J.; Zentek, J. The impact of the Fusarium mycotoxin deoxynivalenol on the health and performance of broiler chickens. Int. J. Mol. Sci. 2011, 12, 7996–8012. [Google Scholar] [CrossRef] [PubMed]
- Ghareeb, K.; Awad, W.A.; Böhm, J. Ameliorative effect of a microbial feed additive on infectious bronchitis virus antibody titer and stress index in broiler chicks fed deoxynivalenol. Poult. Sci. 2012, 91, 800–807. [Google Scholar] [CrossRef]
- Wang, A.; Hogan, N.S. Performance effects of feed-borne Fusarium mycotoxins on broiler chickens: Influences of timing and duration of exposure. Anim. Nutr. 2019, 5, 32–40. [Google Scholar] [CrossRef]
- Li, D.; Ye, Y.; Lin, S.; Deng, L.; Fan, X.; Zhang, Y.; Deng, X.; Li, Y.; Yan, H.; Ma, Y. Evaluation of deoxynivalenol-induced toxic effects on DF-1 cells in vitro: Cell-cycle arrest, oxidative stress, and apoptosis. Environ. Toxicol. Pharmacol. 2014, 37, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Li, L.; Duan, Y.; Yang, X. Antioxidant activity of JM113 in vitro and its protective effect on broiler chickens challenged with deoxynivalenol. J. Anim. Sci. 2017, 95, 837–846. [Google Scholar] [CrossRef] [PubMed]
- de Souza, M.; Baptista, A.A.S.; Valdiviezo, M.J.; Justino, L.; Menck-Costa, M.F.; Ferraz, C.R.; da Gloria, E.M.; Verri, W.A., Jr.; Bracarense, A.P.F. Lactobacillus spp. reduces morphological changes and oxidative stress induced by deoxynivalenol on the intestine and liver of broilers. Toxicon 2020, 185, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Faixová, Z.; Faix, Š.; Leng, L.; Váczi, P.; Renáta, S.; Maková, Z. Effects of feeding diet contaminated with deoxynivalenol on plasma chemistry in growing broiler chickens and the efficacy of glucomannan mycotoxin adsorbent. Acta Vet. 2006, 56, 479–487. [Google Scholar]
- Faixová, Z.; Faix, Š.; Bořutová, R.; Leng, L. Efficacy of dietary selenium to counteract toxicity of deoxynivalenol in growing broiler chickens. Acta Vet. Brno 2007, 76, 349–356. [Google Scholar] [CrossRef]
- Tola, S.; Bureau, D.P.; Hooft, J.M.; Beamish, F.W.; Sulyok, M.; Krska, R.; Encarnação, P.; Petkam, R. Effects of Wheat Naturally Contaminated with Fusarium Mycotoxins on Growth Performance and Selected Health Indices of Red Tilapia (Oreochromis niloticus × O. mossambicus). Toxins 2015, 7, 1929–1944. [Google Scholar] [CrossRef]
- Awad, W.; Böhm, J.; Razzazi-Fazeli, E.; Ghareeb, K.; Zentek, J. Effect of addition of a probiotic microorganism to broiler diets contaminated with deoxynivalenol on performance and histological alterations of intestinal villi of broiler chickens. Poult. Sci. 2006, 85, 974–979. [Google Scholar] [CrossRef]
- Mousavi Khaneghah, A.; Kamani, M.H.; Fakhri, Y.; Coppa, C.; de Oliveira, C.A.F.; Sant’Ana, A.S. Changes in masked forms of deoxynivalenol and their co-occurrence with culmorin in cereal-based products: A systematic review and meta-analysis. Food Chem. 2019, 294, 587–596. [Google Scholar] [CrossRef]
- Khaneghah, A.M.; Martins, L.M.; von Hertwig, A.M.; Bertoldo, R.; Sant’Ana, A.S. Deoxynivalenol and its masked forms: Characteristics, incidence, control and fate during wheat and wheat based products processing—A review. Trends Food Sci. Technol. 2018, 71, 13–24. [Google Scholar] [CrossRef]
- Mousavi Khaneghah, A.; Fakhri, Y.; Raeisi, S.; Armoon, B.; Sant’Ana, A.S. Prevalence and concentration of ochratoxin A, zearalenone, deoxynivalenol and total aflatoxin in cereal-based products: A systematic review and meta-analysis. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2018, 118, 830–848. [Google Scholar] [CrossRef]
- Broekaert, N.; Devreese, M.; van Bergen, T.; Schauvliege, S.; De Boevre, M.; De Saeger, S.; Vanhaecke, L.; Berthiller, F.; Michlmayr, H.; Malachová, A. In vivo contribution of deoxynivalenol-3-β-d-glucoside to deoxynivalenol exposure in broiler chickens and pigs: Oral bioavailability, hydrolysis and toxicokinetics. Arch. Toxicol. 2017, 91, 699–712. [Google Scholar] [CrossRef]
- Santos, R.R.; Molist, F. Effect of different dietary levels of corn naturally contaminated with DON and its derivatives 3+ 15 Ac-DON and DON-3-glucoside on the performance of broilers. Heliyon 2020, 6, e05257. [Google Scholar] [CrossRef]
- Payros, D.; Alassane-Kpembi, I.; Pierron, A.; Loiseau, N.; Pinton, P.; Oswald, I.P. Toxicology of deoxynivalenol and its acetylated and modified forms. Arch. Toxicol. 2016, 90, 2931–2957. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Contaminants in the Food Chain. Scientific Opinion on risks for animal and public health related to the presence of nivalenol in food and feed. EFSA J. 2013, 11, 3262. [Google Scholar] [CrossRef]
- Oliveira, G.; Evangelista, S.R.; Passamani, F.R.F.; Santiago, W.D.; Cardoso, M.d.G.; Batista, L.R. Influence of temperature and water activity on Ochratoxin A production by Aspergillus strain in coffee south of Minas Gerais/Brazil. LWT 2019, 102, 1–7. [Google Scholar] [CrossRef]
- Tahir, M.A.; Abbas, A.; Muneeb, M.; Bilal, R.M.; Hussain, K.; Abdel-Moneim, A.-M.E.; Farag, M.R.; Dhama, K.; Elnesr, S.S.; Alagawany, M. Ochratoxicosis in poultry: Occurrence, environmental factors, pathological alterations and amelioration strategies. World’s Poult. Sci. J. 2022, 78, 727–749. [Google Scholar] [CrossRef]
- Elaroussi, M.A.; Mohamed, F.R.; El Barkouky, E.M.; Atta, A.M.; Abdou, A.M.; Hatab, M.H. Experimental ochratoxicosis in broiler chickens. Avian Pathol. 2006, 35, 263–269. [Google Scholar] [CrossRef]
- Stoev, S.D. Studies on some feed additives and materials giving partial protection against the suppressive effect of ochratoxin A on egg production of laying hens. Res. Vet. Sci. 2010, 88, 486–491. [Google Scholar] [CrossRef]
- Solcan, C.; Pavel, G.; Floristean, V.C.; Chiriac, I.S.; Şlencu, B.G.; Solcan, G. Effect of ochratoxin A on the intestinal mucosa and mucosa-associated lymphoid tissues in broiler chickens. Acta Vet. Hung. 2015, 63, 30–48. [Google Scholar] [CrossRef] [PubMed]
- Pappas, A.C.; Tsiplakou, E.; Tsitsigiannis, D.I.; Georgiadou, M.; Iliadi, M.K.; Sotirakoglou, K.; Zervas, G. The role of bentonite binders in single or concomitant mycotoxin contamination of chicken diets. Br. Poult. Sci. 2016, 57, 551–558. [Google Scholar] [CrossRef]
- Yu, Z.; Wu, F.; Tian, J.; Guo, X.; An, R. Protective effects of compound ammonium glycyrrhizin, L-arginine, silymarin and glucurolactone against liver damage induced by ochratoxin A in primary chicken hepatocytes. Mol. Med. Rep. 2018, 18, 2551–2560. [Google Scholar] [CrossRef] [PubMed]
- Kövesi, B.; Cserháti, M.; Erdélyi, M.; Zándoki, E.; Mézes, M.; Balogh, K. Long-term effects of ochratoxin A on the glutathione redox system and its regulation in chicken. Antioxidants 2019, 8, 178. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Cao, Z.; Guo, Y.; Tong, C.; Yang, S.; Long, M.; Li, P.; He, J. Selenium Yeast Alleviates Ochratoxin A-Induced Apoptosis and Oxidative Stress via Modulation of the PI3K/AKT and Nrf2/Keap1 Signaling Pathways in the Kidneys of Chickens. Oxidative Med. Cell. Longev. 2020, 2020, 4048706. [Google Scholar] [CrossRef] [PubMed]
- Stoev, S.; Mircheva, T.; Denev, S.; Chobanova, S.; Ivanov, V. The Protective Effect of Silymarin against Ochratoxin A Induced Histopathological and Biochemical Changes in Chicks. J. Adv. Vet. Res. 2021, 11, 1–8. [Google Scholar]
- Gentles, A.; Smith, E.E.; Kubena, L.F.; Duffus, E.; Johnson, P.; Thompson, J.; Harvey, R.B.; Edrington, T.S. Toxicological evaluations of cyclopiazonic acid and ochratoxin A in broilers. Poult. Sci. 1999, 78, 1380–1384. [Google Scholar] [CrossRef] [PubMed]
- El Cafsi, I.; Bjeoui, S.; Rabeh, I.; Nechi, S.; Chelbi, E.; El Cafsi, M.; Ghram, A. Effects of ochratoxin A on membrane phospholipids of the intestine of broiler chickens, practical consequences. Animal 2020, 14, 933–941. [Google Scholar] [CrossRef]
- Surai, P.F. Antioxidants in poultry nutrition and reproduction: An update. Antioxidants 2020, 9, 105. [Google Scholar] [CrossRef]
- Ding, L.; Han, M.; Wang, X.; Guo, Y. Ochratoxin A: Overview of Prevention, Removal, and Detoxification Methods. Toxins 2023, 15, 565. [Google Scholar] [CrossRef]
- Liuzzi, V.C.; Fanelli, F.; Tristezza, M.; Haidukowski, M.; Picardi, E.; Manzari, C.; Lionetti, C.; Grieco, F.; Logrieco, A.F.; Thon, M.R.; et al. Transcriptional Analysis of Acinetobacter sp. neg1 Capable of Degrading Ochratoxin A. Front. Microbiol. 2017, 7, 2162. [Google Scholar] [CrossRef]
- Tao, Y.; Xie, S.; Xu, F.; Liu, A.; Wang, Y.; Chen, D.; Pan, Y.; Huang, L.; Peng, D.; Wang, X.; et al. Ochratoxin A: Toxicity, oxidative stress and metabolism. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2018, 112, 320–331. [Google Scholar] [CrossRef]
- EC–European Commission. Commission Recommendation 2006/576/EC of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding. Off. J. Eur. Union 2006, L229, 7–9. [Google Scholar]
- Ruan, D.; Wang, W.; Lin, C.; Fouad, A.; Chen, W.; Xia, W.; Wang, S.; Luo, X.; Zhang, W.; Yan, S. Effects of curcumin on performance, antioxidation, intestinal barrier and mitochondrial function in ducks fed corn contaminated with ochratoxin A. Animal 2019, 13, 42–52. [Google Scholar] [CrossRef]
- Wang, W.; Zhai, S.; Xia, Y.; Wang, H.; Ruan, D.; Zhou, T.; Zhu, Y.; Zhang, H.; Zhang, M.; Ye, H.; et al. Ochratoxin A induces liver inflammation: Involvement of intestinal microbiota. Microbiome 2019, 7, 151. [Google Scholar] [CrossRef]
- Yang, S.; Li, L.; Yu, L.; Sun, L.; Li, K.; Tong, C.; Xu, W.; Cui, G.; Long, M.; Li, P. Selenium-enriched yeast reduces caecal pathological injuries and intervenes changes of the diversity of caecal microbiota caused by Ochratoxin-A in broilers. Food Chem. Toxicol. 2020, 137, 111139. [Google Scholar] [CrossRef]
- Zhai, S.; Zhu, Y.; Feng, P.; Li, M.; Wang, W.; Yang, L.; Yang, Y. Ochratoxin A: Its impact on poultry gut health and microbiota, an overview. Poult. Sci. 2021, 100, 101037. [Google Scholar] [CrossRef]
- Tong, C.; Li, P.; Yu, L.-H.; Li, L.; Li, K.; Chen, Y.; Yang, S.-H.; Long, M. Selenium-rich yeast attenuates ochratoxin A-induced small intestinal injury in broiler chickens by activating the Nrf2 pathway and inhibiting NF-KB activation. J. Funct. Foods 2020, 66, 103784. [Google Scholar] [CrossRef]
- Munkvold, G.P.; Arias, S.; Taschl, I.; Gruber-Dorninger, C. Mycotoxins in corn: Occurrence, impacts, and management. In Corn; Elsevier: Amsterdam, The Netherlands, 2019; pp. 235–287. [Google Scholar]
- García-Díaz, M.; Gil-Serna, J.; Vázquez, C.; Botia, M.N.; Patiño, B. A comprehensive study on the occurrence of mycotoxins and their producing fungi during the maize production cycle in Spain. Microorganisms 2020, 8, 141. [Google Scholar] [CrossRef] [PubMed]
- Lumsangkul, C.; Chiang, H.-I.; Lo, N.-W.; Fan, Y.-K.; Ju, J.-C. Developmental toxicity of mycotoxin fumonisin b1 in animal embryogenesis: An overview. Toxins 2019, 11, 114. [Google Scholar] [CrossRef]
- Awuchi, C.G.; Ondari, E.N.; Nwozo, S.; Odongo, G.A.; Eseoghene, I.J.; Twinomuhwezi, H.; Ogbonna, C.U.; Upadhyay, A.K.; Adeleye, A.O.; Okpala, C.O.R. Mycotoxins’ Toxicological Mechanisms Involving Humans, Livestock and Their Associated Health Concerns: A Review. Toxins 2022, 14, 167. [Google Scholar] [CrossRef] [PubMed]
- Laurain, J.; Tardieu, D.; Matard-Mann, M.; Rodriguez, M.A.; Guerre, P. Fumonisin B1 accumulates in chicken tissues over time and this accumulation was reduced by feeding algo-clay. Toxins 2021, 13, 701. [Google Scholar] [CrossRef] [PubMed]
- Poersch, A.B.; Trombetta, F.; Braga, A.C.M.; Boeira, S.P.; Oliveira, M.S.; Dilkin, P.; Mallmann, C.A.; Fighera, M.R.; Royes, L.F.F.; Oliveira, M.S.; et al. Involvement of oxidative stress in subacute toxicity induced by fumonisin B1 in broiler chicks. Vet. Microbiol. 2014, 174, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Sousa, M.C.S.; Galli, G.M.; Alba, D.F.; Griss, L.G.; Gebert, R.R.; Souza, C.F.; Baldissera, M.D.; Gloria, E.M.; Mendes, R.E.; Zanelato, G.O.; et al. Pathogenetic effects of feed intake containing of fumonisin (Fusarium verticillioides) in early broiler chicks and consequences on weight gain. Microb. Pathog. 2020, 147, 104247. [Google Scholar] [CrossRef] [PubMed]
- Galli, G.M.; Griss, L.G.; Fortuoso, B.F.; Silva, A.D.; Fracasso, M.; Lopes, T.F.; Schetinger, M.R.S.; Gundel, S.; Ourique, A.F.; Carneiro, C.; et al. Feed contaminated by fumonisin (Fusarium spp.) in chicks has a negative influence on oxidative stress and performance, and the inclusion of curcumin-loaded nanocapsules minimizes these effects. Microb. Pathog. 2020, 148, 104496. [Google Scholar] [CrossRef] [PubMed]
- Nwangburuka, C.; Denton, L.; Chioma, G.; Oyekale, K.; Tayo, G.; Ajayi, B.; Olarinmoye, A.; Ezekiel, C.; Taiwo, E.; Babalola, O. Effect of levels of equisetin and fumonisin mycotoxins on blood parameters of broiler chicks. Mycopath 2019, 15. [Google Scholar]
- Armanini, E.H.; Boiago, M.M.; Cécere, B.G.d.O.; Oliveira, P.V.; Teixeira, C.J.; Strapazzon, J.V.; Bottari, N.B.; Silva, A.D.; Fracasso, M.; Vendruscolo, R.G. Protective effects of silymarin in broiler feed contaminated by mycotoxins: Growth performance, meat antioxidant status, and fatty acid profiles. Trop. Anim. Health Prod. 2021, 53, 442. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, D.; Keum, M.; Han, E.; An, B.; Chang, H.; Choi, Y.; Moon, B.; Lee, K. Effects of fumonisin B1 and mycotoxin binders on growth performance, tibia characteristics, gut physiology, and stress indicators in broiler chickens raised in different stocking densities. Poult. Sci. 2018, 97, 845–854. [Google Scholar] [CrossRef]
- Li, Z.; Yang, Z.B.; Yang, W.R.; Wang, S.J.; Jiang, S.Z.; Wu, Y.B. Effects of feed-borne Fusarium mycotoxins with or without yeast cell wall adsorbent on organ weight, serum biochemistry, and immunological parameters of broiler chickens. Poult. Sci. 2012, 91, 2487–2495. [Google Scholar] [CrossRef]
- Chen, Y.; Cheng, Y.; Wen, C.; Wang, W.; Kang, Y.; Wang, A.; Zhou, Y. The protective effects of modified palygorskite on the broilers fed a purified zearalenone-contaminated diet. Poult. Sci. 2019, 98, 3802–3810. [Google Scholar] [CrossRef]
- Antonissen, G.; Croubels, S.; Pasmans, F.; Ducatelle, R.; Eeckhaut, V.; Devreese, M.; Verlinden, M.; Haesebrouck, F.; Eeckhout, M.; De Saeger, S.; et al. Fumonisins affect the intestinal microbial homeostasis in broiler chickens, predisposing to necrotic enteritis. Vet. Res. 2015, 46, 98. [Google Scholar] [CrossRef]
- Grenier, B.; Applegate, T.J. Modulation of intestinal functions following mycotoxin ingestion: Meta-analysis of published experiments in animals. Toxins 2013, 5, 396–430. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain; Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L. Risk for animal and human health related to the presence of dioxins and dioxin-like PCBs in feed and food. EFSA J. 2018, 16, e05333. [Google Scholar]
- Lai, Y.; Sun, M.; He, Y.; Lei, J.; Han, Y.; Wu, Y.; Bai, D.; Guo, Y.; Zhang, B. Mycotoxins binder supplementation alleviates aflatoxin B(1) toxic effects on the immune response and intestinal barrier function in broilers. Poult. Sci. 2022, 101, 101683. [Google Scholar] [CrossRef] [PubMed]
- Hueza, I.M.; Raspantini, P.C.F.; Raspantini, L.E.R.; Latorre, A.O.; Górniak, S.L. Zearalenone, an estrogenic mycotoxin, is an immunotoxic compound. Toxins 2014, 6, 1080–1095. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Yang, W.; Yang, Z.; Jiang, S.; Zhang, G.; Jiang, X. Effects of low dose zearalenone and adsorbent on growth performance, serum biochemical and antioxidant indices of growing-laying hens. Chin. J. Anim. Nutr. 2016, 28, 1137–1144. [Google Scholar]
- Bibo, Z.; Lei, Z.; Weili, Y.; Yefei, C.; Chao, W.; Yanmin, Z. Effect of zearalenone on serum parameters, hepatic oxidative damage and residue of zearalenone in broilers. Anim. Husb. Vet. Med. 2016, 48, 10–14. [Google Scholar]
- Ling, A.; Guo, J.; Guo, W.; Yang, J.; Zhao, Z. Effects of zearalenone on laying performance, hematological indices and reproductive hormone levels in laying hens. Acta Agric. Shanghai 2019, 35, 100–106. [Google Scholar]
- Yuan, T.; Li, J.; Wang, Y.; Li, M.; Yang, A.; Ren, C.; Qi, D.; Zhang, N. Effects of Zearalenone on Production Performance, Egg Quality, Ovarian Function and Gut Microbiota of Laying Hens. Toxins 2022, 14, 653. [Google Scholar] [CrossRef] [PubMed]
- Rai, A.; Das, M.; Tripathi, A. Occurrence and toxicity of a fusarium mycotoxin, zearalenone. Crit. Rev. Food Sci. Nutr. 2020, 60, 2710–2729. [Google Scholar] [CrossRef]
- Grešáková, Ľ.; Bořutová, R.; Faix, Š.; Plachá, I.; Čobanová, K.; Košíková, B.; Leng, Ľ. Effect of lignin on oxidative stress in chickens fed a diet contaminated with zearalenone. Acta Vet. Hung. 2012, 60, 103–114. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Y.; Jiang, L.; Huang, H. Improvement of the enzymatic detoxification activity towards mycotoxins through structure-based engineering. Biotechnol. Adv. 2022, 56, 107927. [Google Scholar] [CrossRef]
- Bulgaru, C.V.; Marin, D.E.; Pistol, G.C.; Taranu, I. Zearalenone and the immune response. Toxins 2021, 13, 248. [Google Scholar] [CrossRef]
- Rogowska, A.; Pomastowski, P.; Sagandykova, G.; Buszewski, B. Zearalenone and its metabolites: Effect on human health, metabolism and neutralisation methods. Toxicon 2019, 162, 46–56. [Google Scholar] [CrossRef]
- Lorenz, N.; Dänicke, S.; Edler, L.; Gottschalk, C.; Lassek, E.; Marko, D.; Rychlik, M.; Mally, A. A critical evaluation of health risk assessment of modified mycotoxins with a special focus on zearalenone. Mycotoxin Res. 2019, 35, 27–46. [Google Scholar] [CrossRef]
- Tatay, E.; Espin, S.; Garcia-Fernandez, A.-J.; Ruiz, M.-J. Oxidative damage and disturbance of antioxidant capacity by zearalenone and its metabolites in human cells. Toxicol. Vitr. 2017, 45, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Hort, V.; Nicolas, M.; Travel, A.; Jondreville, C.; Maleix, C.; Baéza, E.; Engel, E.; Guérin, T. Carry-over assessment of fumonisins and zearalenone to poultry tissues after exposure of chickens to a contaminated diet–A study implementing stable-isotope dilution assay and UHPLC-MS/MS. Food Control 2020, 107, 106789. [Google Scholar] [CrossRef]
- Buranatragool, K.; Poapolathep, S.; Isariyodom, S.; Imsilp, K.; Klangkaew, N.; Poapolathep, A. Dispositions and tissue residue of zearalenone and its metabolites α-zearalenol and β-zearalenol in broilers. Toxicol. Rep. 2015, 2, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Blandino, M.; Scarpino, V.; Giordano, D.; Sulyok, M.; Krska, R.; Vanara, F.; Reyneri, A. Impact of sowing time, hybrid and environmental conditions on the contamination of maize by emerging mycotoxins and fungal metabolites. Ital. J. Agron. 2017, 12. [Google Scholar] [CrossRef]
- Dufera, L.T.; Jimma, E. Management of mycotoxin in post-harvest food chain of durable crops. Management 2020, 100, 12–20. [Google Scholar]
- Neme, K.; Mohammed, A. Mycotoxin occurrence in grains and the role of postharvest management as a mitigation strategies. A review. Food Control 2017, 78, 412–425. [Google Scholar] [CrossRef]
- Zhu, Y.; Hassan, Y.I.; Watts, C.; Zhou, T. Innovative technologies for the mitigation of mycotoxins in animal feed and ingredients—A review of recent patents. Anim. Feed. Sci. Technol. 2016, 216, 19–29. [Google Scholar] [CrossRef]
- Sherif, K.E.; Hasan, R.; El-Gogary, M.; Ismail, R.F. Effectivenesss of some Feed Additives for Detoxification of Aflatoxin B1 in Laying Hens. J. Anim. Poult. Prod. 2018, 9, 345–356. [Google Scholar] [CrossRef]
- Chen, X.; Horn, N.L.; Applegate, T.J. Efficiency of hydrated sodium calcium aluminosilicate to ameliorate the adverse effects of graded levels of aflatoxin B1 in broiler chicks. Poult. Sci. 2014, 93, 2037–2047. [Google Scholar] [CrossRef]
- Tso, K.-H.; Ju, J.-C.; Fan, Y.-K.; Chiang, H.-I. Enzyme degradation reagents effectively remove mycotoxins deoxynivalenol and zearalenone from pig and poultry artificial digestive juices. Toxins 2019, 11, 599. [Google Scholar] [CrossRef]
- Li, L.; Zhang, T.; Ren, X.; Li, B.; Wang, S. Male reproductive toxicity of zearalenone—Meta-analysis with mechanism review. Ecotoxicol. Environ. Saf. 2021, 221, 112457. [Google Scholar] [CrossRef]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP); Rychen, G.; Aquilina, G.; Azimonti, G.; Bampidis, V.; de Lourdes Bastos, M.; Bories, G.; Chesson, A.; Cocconcelli, P.S.; Flachowsky, G.; et al. Safety and efficacy of fumonisin esterase (FUMzyme®) as a technological feed additive for all avian species. EFSA J. 2016, 14, e04617. [Google Scholar]
- Murcia, H.W.; Diaz, G.J. Protective effect of glutathione S-transferase enzyme activity against aflatoxin B1 in poultry species: Relationship between glutathione S-transferase enzyme kinetic parameters, and resistance to aflatoxin B1. Poult. Sci. 2021, 100, 101235. [Google Scholar] [CrossRef] [PubMed]
- Elhady, M.A.; Khalaf, A.A.A.; Ibrahim, M.A.; Hassanen, E.I.; Abdelrahman, R.E.; Noshy, P.A. Protective effects of fermentation extract against ochratoxin A-induced nephrotoxicity and immunotoxicity in broiler chickens. J. Vet. Res. 2022, 66, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Eisa Ahmed, M.F.; Sangare, L.; Zhao, Y.; Selvaraj, J.N.; Xing, F.; Wang, Y.; Yang, H.; Liu, Y. Novel aflatoxin-degrading enzyme from Bacillusckletonii L7. Toxins 2017, 9, 36. [Google Scholar] [CrossRef]
- Jia, R.; Sadiq, F.A.; Liu, W.; Cao, L.; Shen, Z. Protective effects of Bacillus subtilis ASAG 216 on growth performance, antioxidant capacity, gut microbiota and tissues residues of weaned piglets fed deoxynivalenol contaminated diets. Food Chem. Toxicol. 2021, 148, 111962. [Google Scholar] [CrossRef] [PubMed]
- Ruan, M.-L.; Wang, J.; Xia, Z.-Y.; Li, X.-W.; Zhang, B.; Wang, G.-L.; Wu, Y.-Y.; Han, Y.; Deng, J.; Sun, L.-H. An integrated mycotoxin-mitigating agent can effectively mitigate the combined toxicity of AFB1, DON and OTA on the production performance, liver and oviduct health in broiler breeder hens. Food Chem. Toxicol. 2023, 182, 114159. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Wang, X.; Nepovimova, E.; Wang, Y.; Yang, H.; Li, L.; Zhang, X.; Kuca, K. Antioxidant agents against trichothecenes: New hints for oxidative stress treatment. Oncotarget 2017, 8, 110708. [Google Scholar] [CrossRef]
- Wang, X.; Zuo, Z.; Zhao, C.; Zhang, Z.; Peng, G.; Cao, S.; Hu, Y.; Yu, S.; Zhong, Z.; Deng, J. Protective role of selenium in the activities of antioxidant enzymes in piglet splenic lymphocytes exposed to deoxynivalenol. Environ. Toxicol. Pharmacol. 2016, 47, 53–61. [Google Scholar] [CrossRef]
- Xiao, Y.; Xu, S.; Zhao, S.; Liu, K.; Lu, Z.; Hou, Z. Protective effects of selenium against zearalenone-induced apoptosis in chicken spleen lymphocyte via an endoplasmic reticulum stress signaling pathway. Cell Stress. Chaperones 2019, 24, 77–89. [Google Scholar] [CrossRef]
- Zheng, J.; Zhang, Y.; Xu, W.; Luo, Y.; Hao, J.; Shen, X.L.; Yang, X.; Li, X.; Huang, K. Zinc protects HepG2 cells against the oxidative damage and DNA damage induced by ochratoxin A. Toxicol. Appl. Pharmacol. 2013, 268, 123–131. [Google Scholar] [CrossRef]
- Gao, X.; Xiao, Z.-H.; Liu, M.; Zhang, N.-Y.; Khalil, M.M.; Gu, C.-Q.; Qi, D.-S.; Sun, L.-H. Dietary silymarin supplementation alleviates zearalenone-induced hepatotoxicity and reproductive toxicity in rats. J. Nutr. 2018, 148, 1209–1216. [Google Scholar] [CrossRef] [PubMed]
- Vartiainen, S.; Yiannikouris, A.; Apajalahti, J.; Moran, C.A. Comprehensive evaluation of the efficiency of yeast cell wall extract to adsorb ochratoxin A and mitigate accumulation of the toxin in broiler chickens. Toxins 2020, 12, 37. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Feng, Y.; Wei, J.-T.; Zhu, M.-X.; Zhang, L.; Zhang, J.-C.; Karrow, N.A.; Han, Y.-M.; Wu, Y.-Y.; Guo, Y.-M. Mitigation effects of bentonite and yeast cell wall binders on AFB1, DON, and OTA induced changes in laying hen performance, egg quality, and health. Toxins 2021, 13, 156. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Li, Z.; Wang, G.; Yang, Z.; Yang, W.; Zhang, G.; Wu, Y. Effects of Fusarium mycotoxins with yeast cell wall absorbent on hematology, serum biochemistry, and oxidative stress in broiler chickens. J. Appl. Poult. Res. 2014, 23, 165–173. [Google Scholar] [CrossRef]
- Elliott, C.T.; Connolly, L.; Kolawole, O. Potential adverse effects on animal health and performance caused by the addition of mineral adsorbents to feeds to reduce mycotoxin exposure. Mycotoxin Res. 2020, 36, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Prapapanpong, J.; Udomkusonsri, P.; Mahavorasirikul, W.; Choochuay, S.; Tansakul, N. In vitro studies on gastrointestinal monogastric and avian models to evaluate the binding efficacy of mycotoxin adsorbents by liquid chromatography-tandem mass spectrometry. J. Adv. Vet. Anim. Res. 2019, 6, 125. [Google Scholar] [CrossRef] [PubMed]
- Kolawole, O.; Meneely, J.; Greer, B.; Chevallier, O.; Jones, D.S.; Connolly, L.; Elliott, C. Comparative in vitro assessment of a range of commercial feed additives with multiple mycotoxin binding claims. Toxins 2019, 11, 659. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Aflatoxin | Treatment | Experimental Period/Days | Impacts/Chicken Type | Publication Date | References |
|---|---|---|---|---|---|
| Feed Efficiency and Growth Performance | 5 mg/kg | 22-d | Decrease in BWG/Broiler | 2001 | [45] |
| 2.5 mg/kg | 27-d | Decrease in BWG/Broiler | 2005 | [46] | |
| 2.5 mg/kg | 28-d | Decrease in EQT/Layer | 2005 | [47] | |
| 0.05, 0.1 mg/kg | 42-d | Decrease in FI and BW/Broiler | 2006 | [48] | |
| 0.2 mg/kg | 33-d | Decrease in FI and BWG/Broiler | 2006 | [49] | |
| 149 µg/kg | 14-d | Decrease in FI and BW/Broiler | 2019 | [50] | |
| 100 mg/kg | 28-d | Increase in FCR and Decrease in FI and BWG/Broiler | 2012 | [51] | |
| 80 µ/kg | 7-d | Increase in FCR and decrease in FI and BWG/Broiler | 2012 | [52] | |
| 250–500 ppb/kg | 35-d | Decrease in BW/Broiler | 2013 | [53] | |
| 2 mg/kg | 21-d | Decrease in BW and BWG/Broiler | 2015 | [54] | |
| 0.5–2 ppm | 42-d | Increase in FCR and decrease in DFI and DWG/Broiler | 2017 | [55] | |
| 500 ppb/kg | 35-d | Increase in FCR and decrease in BW/Broiler | 2017 | [56] | |
| 0.5 mg/kg | 56-d | Increase in FCR and MR and decrease in BW and BWG/Broiler | 2019 | [44] | |
| 0.6 mg/kg | 42-d | Increase in FCR and decrease in FI and BW/Broiler | 2022 | [57] | |
| Antioxidants status | 0.05, 0.1, 0.5, 1 mg/kg | 45-d | Increase in MDA and decrease in SOD, CAT, G6PD and GSH-Px/Broiler | 2005 | [58] |
| 3.4 or 8.2 mg/kg | 41-d | Increase in MDA in liver, kidney, serum and decrease in GPx activity in liver tissue/Broiler | 2008 | [59] | |
| 7.54 mg/kg | 21-d | Increase in HIF-1α and HMOX in jejunum and xanthine oxidoreductase in liver/Broiler | 2013 | [60] | |
| 1 ppm/kg | 42-d | Increase in MDA and SOD and decrease in and TAC and CAT/Broiler | 2014 | [61] | |
| 0.15, 0.3 and 0.6 mg/kg | 21-d | Increase in MDA and GSH and decrease in spleen levels of GSH-Px, GR and CAT/broiler | 2016 | [62] | |
| 1 mg/kg | 28-d | Increase in MDA, decrease in liver and serum CAT, GPx, T-SOD, GR and GSTs/Broiler | 2017 | [63] | |
| 20 mg/kg | 42-d | Increase in MDA/Broiler | 2022 | [64] | |
| Blood parameters | 0.8 mg/kg | 35-d | Decrease in plasma ALT/Broiler | 2004 | [65] |
| 2.5 mg/kg | 27-d | Decrease serum TP, ALB, and GLB/Broiler | 2005 | [46] | |
| 0.05, 0.1 mg/kg | 42-d | Decrease in g-GGT, AST, and ALT/Broiler | 2006 | [48] | |
| 80 µ/kg | 7-d | Increase in plasma T.P and decrease in ALP, AST and ALT/Broiler | 2012 | [52] | |
| Blood parameters | 250–500 ppb | 35-d | Increase in ALB, direct bilirubin, Ca, and P; decrease in UA, Glu, total bilirubin, ALT, AST, and γ-GT/Broiler | 2014 | [66] |
| 2 mg/kg | 21-d | Decrease in serum T.P, ALB, Ca, and Glu/Broiler | 2015 | [54] | |
| 500 ppm/kg | 28-d | Decrease in ALB/Broiler | 2015 | [67] | |
| 500 ppb/kg | 35-d | Increase in ALT/Broiler | 2017 | [56] | |
| 250–500 ppm | 21-d | Decrease in Glu, Ca, HDL, Cr and increase in AST, ALT/Broiler | 2018 | [68] | |
| 20 mg/kg | 21-d | Increase in Ca and HDL; decrease in ALT, AST/Broiler | 2022 | [64] | |
| 0.6 mg/kg | 42-d | Increase in ALT and AST/Broiler | 2022 | [57] | |
| Internal organs | 2, 2.5, 5 mg/kg | 21-d | Increase weight of liver and changed hepatic histopathology/Broiler | 2001, 2005, 2015 | [45,46,54] |
| 0.8 mg/kg | 35-d | liver had necrosis along with multifocal portal infiltration/Broiler | 2004 | [65] | |
| 2.5 mg/kg | 28-d | Liver AFB1 residues/Layer | 2005 | [47] | |
| 0.05, 0.1 mg/kg | 42-d | Residues of AFB1 and AFM1 in livers and muscles/Broiler | 2006 | [48] | |
| 0.2 mg/kg | 33-d | Changed hepatic histopathology/Broiler | 2006 | [49] | |
| 80–285 µg/kg | 35-d | Decrease thymus and bursa weights/Broiler | 2011 | [69] | |
| 80 µ/kg | 7-d | Paleness, bleeding, and fragility in the liver Kidney swelling and bursal atrophy/Broiler | 2012 | [52] | |
| 250–500 ppb/kg | 35-d | reduced ileum, duodenum, and jejunum intestinal length/Broiler | 2013 | [53] | |
| 250–500 ppb/kg | 35-d | reduced ileum, duodenum, and jejunum intestinal length/Broiler | 2013 | [53] | |
| 0.5–2 ppm | 42-d | Increase heart weight/Broiler | 2017 | [55] | |
| 0.5 mg/kg | 56-d | Increase weights of liver, spleen and kidney/Broiler | 2019 | [44] |
| Deoxynivalenol | Treatment | Experimental Period | Impacts/Chicken Type | Publication Date | References |
|---|---|---|---|---|---|
| Feed Efficiency and Growth Performance | 4.7 or 8.2 mg/kg | 56-d | Increase FI and BW/Broiler | 2002 | [103] |
| Up 14 mg/kg | 35-d | Increase in FCR and decreased in FI, BW/Broiler | 2003 | [104] | |
| 5.9 or 9.5 mg/kg | 56-d | Decrease in FI and BWG/Broiler | 2004 | [105] | |
| 5 mg/kg | 21-d | Decrease in FI/Broiler | 2006 | [106] | |
| 1.5 mg/kg | 53-d | Improved in FCR and increase in BW/Broiler | 2007 | [107] | |
| Up to 18 mg/kg | 21-d | Decrease in FI and BWG/Broilers | 2011 | [108] | |
| 1 and 5 mg/kg | 35-d | Decrease in FI, BW and BWG/Broiler | 2011 | [109] | |
| 10 mg/kg | 35-d | Increase in FCR and decrease in FI, BW, BWG/Broiler | 2012 | [110] | |
| 1.68 or 12.20 mg/kg | 35-d | Decrease in FI and BWG/Broiler | 2012 | [97] | |
| 2.5, 5 or 10 mg/kg | 35-d | Decrease in FI, BW and BWG/Broiler | 2017 | [94] | |
| 7.9 mg/kg | 34-d | Increase in FCR and Decrease in FI and BW and BWG/Broiler | 2019 | [111] | |
| 5 or 15 mg/kg | 42-d | Increase in FCR and decrease in BW/Broiler | 2020 | [98] | |
| Antioxidant stat-us | 100–200 ng/mL | 6–48-h | Increase in ROS and MDA, decrease in GSH and SOD in embryo fibroblast DF-1 cells/Broiler | 2014 | [112] |
| 10 mg/kg | 35 | Increase in TBARS in jejunum/Broiler | 2014 | [99] | |
| 10 mg/kg | 20-h | Decrease in SOD in serum and MDA or TBARS in the jejunal mucosa/Broiler | 2017 | [113] | |
| 19.3 mg/kg | 7-d | Increase in TBARS GSH and decrease in ABTS in jejunum/Broiler | 2020 | [114] | |
| Blood Parameters | 3 mg/kg | 42-d | Decrease in T.P, mg, T.G, F.G and increase in ALT/Broiler | 2006 | [115] |
| 3 mg/kg | 42-d | Decrease in T.P, T.G, F.G and increase in ALT/Broiler | 2007 | [116] | |
| Blood Parameters | 2.95 mg/kg | 28-d | Decrease in T.P, ALB and increase in ALT, AST and ALP/Broiler | 2011 | [117] |
| 10 mg/kg | 35-d | Decrease in plasma T.P, T.G and U.A/Broiler | 2012 | [110] | |
| 10 mg/kg | 35-d | Decrease in ALT, Cho, and T.G/Broiler | 2016 | [100] | |
| 5 or 15 | 42-d | Decrease in CK and Cho/Broiler | 2020 | [98] | |
| 15 mg/kg | 42-d | Decrease in serum Cho/Broiler | 2020 | [98] | |
| 5 or 15 mg/kg | 42-d | Decrease in HGB and erythrocytes/Broiler | 2021 | [16] | |
| Internal organs | Up to 14 mg/kg | 35-d | Increase relative weight of heart and decrease relative weight of spleen/Broiler | 2003 | [104] |
| 5 mg/kg | 21-d | Unaffected relative weights heart, gizzard, pancreas, caecum, colon, spleen and decrease in small intestine/Broiler | 2006 | [118] | |
| 1.68 or 12.20 mg/kg | 35-d | Increase relative weights of liver and spleen and decrease relative weights of duodenum and jejunum/Broiler | 2012 | [97] | |
| 10 mg/kg | 35-d | Increase relative weight of gizzard and decrease relative weight of kidney/Broiler | 2014 | [99] | |
| 2, 5 or 10 mg/kg | 112-d | Increase relative weight of spleen/Layer | 2017 | [102] | |
| 2.5, 5 or 10 mg/kg | 35-d | Decrease relative weight of liver/Broiler | 2017 | [94] | |
| 19.3 mg/kg | 8-d | Decrease villi height and intestinal health and increase crypt depth/Broiler | 2020 | [114] | |
| 5 or 15 mg/kg | 42-d | Increase relative weight of gizzard and thymus and decrease relative weight of colon and small intestine/Broiler | 2020 | [98] |
| Ochratoxin | Treatment | Experimental Period | Impacts/Chicken Type | Publication Date | References |
|---|---|---|---|---|---|
| Feed Efficiency and Growth Performance | 0.4 or 0.8 mg/kg | 35-d | Decrease in FI and BW/Broiler | 2006 | [128] |
| 5 mg/kg | 365-d | Decrease in EPR and EW/Layer | 2010 | [129] | |
| 0.05 mg/kg | 28-d | Decrease in BWG/Broiler | 2015 | [130] | |
| 0.1 mg/kg | 42-d | Decrease in BW/Broiler | 2016 | [131] | |
| 0.5 and 1 mg/kg | 35-d | Reduced in FI and GR/Broiler | 2021 | [16] | |
| Antioxidants status | 1 mg/kg | 24-h | Increase in MDA, mRNA expression of apoptosis-associated genes, and apoptosis rate; decrease in SOD, and GSH levels/Broiler | 2018 | [132] |
| 106,654 and 1126 µg/kg | 21-d | Increase in liver and kidney MDA, GSH in blood plasma and liver/Broiler | 2019 | [133] | |
| 50 µg/kg | 21-d | Increase in kidney MDA and decrease in GSH, SOD, CAT (m-RNA expression), SOD (m-RNA expression) and GSH-Px (m-RNA expression)./Broiler | 2020 | [134] | |
| Blood Parameters | 1 mg/kg | 1-d | Increase in AST and ALT/Broiler | 2018 | [132] |
| 3 mg/kg | 10-d | Increase in blood Glu, U.A, ALT and AST/Broiler | 2021 | [135] | |
| Internal organs | 2.5 mg/kg | 21-d | An increase in kidney weight/Broiler | 1999 | [136] |
| 0.4 or 0.8 mg/kg | 35-d | An increase in gizzard mass/Broiler | 2006 | [128] | |
| 0.05 mg/kg | 28-d | Changes in the anatomy of the intestinal mucosa/Broiler | 2015 | [130] | |
| 0.1 mg/kg | 42-d | OTA residues in the liver and increase weight/Broiler | 2016 | [131] | |
| 3 mg/kg | 10-d | Hemorrhages on the epicardium and duodenal mucosa, catarrhal enteritis/Broiler | 2021 | [135] |
| Fumonisins | Treatment | Experimental Period | Impacts/Chicken Type | Publication Date | References |
|---|---|---|---|---|---|
| Feed Efficiency and Growth Performance | 5 mg/kg | 22-d | Decrease in BWG/Broiler | 2001 | [45] |
| 50,200 mg/kg | 33-d | Decrease in BW and BWG/Broiler | 2006 | [49] | |
| 200 mg/kg | 33-d | Decrease in BW and BWG/Broiler | 2006 | [49] | |
| 100 mg/kg | 21-d | Increase in FCR/Broiler | 2014 | [153] | |
| 100 mg/kg | 28-d | Increase in FCR and decrease in FI and BW/Broiler | 2016 | [130] | |
| Antioxidants status | 100 mg/kg | 21-d | Decrease in SOD, GST and NPSH | 2014 | [153] |
| 2.5, 5, or 10 ppm | 11-d | Increase in liver LPO and ROS; decrease in SOD, GPx, and GST activity; decrease in serum LPO levels and ROS levels/Broiler | 2020 | [154] | |
| 600 mg/kg | 15-d | Increase in TPA, ROS, and SOD; decrease in GPx/Broiler | 2020 | [155] | |
| Blood Parameters | 25, 50 or 100 mg/kg | 8 -15-d | Increase in plasma ALT and AST, Ur and Cr/Broiler | 2017 | [156] |
| 2.5, 5, or 10 ppm | 11-d | Decrease in T.P, ALB, Glu, ALT, T.G and U.A/Broiler | 2020 | [154] | |
| 600 mg/kg | 15-d | Increase in Cho and decrease in U.A/Broiler | 2020 | [155] | |
| 0.05–20 ppm | 42-d | Increase in ALT, AST and U.A/Broiler | 2021 | [157] | |
| Internal organs | 50 mg/kg | 22-d | Variations in hepatic histopathology/Broiler | 2006 | [49] |
| 200 mg/kg | 33-d | Increase liver weight and changes in hepatic histopathology/Broiler | 2006 | [49] | |
| 100 mg/kg | 28-d | Reduced villus height and the ratio of villus to crypt Variations in hepatic histopathology/Broiler | 2012 | [51] | |
| 100 mg/kg | 21-d | Increase liver absolut and relative weight/Broiler | 2014 | [153] |
| Zearalenone | Treatment | Experimental Period | Impacts/Chicken Type | Publication Date | References |
|---|---|---|---|---|---|
| Feed Efficiency and Growth Performance | 2 mg/kg | 42-d | Increase in FCR and decrease in BWG/Broiler | 2019 | [160] |
| 0.05–20 ppm | 42-d | Decrease in FI, FCR and WG/Broiler | 2021 | [157] | |
| Antioxidants status | 7.9 mg/kg | 14-d | Increase in GSH-Px, and levels of MDA/Broiler | 2012 | [171] |
| 0.4 mg/kg | 47-d | Increase in GSH-Px and T-SOD/Layers | 2016 | [166] | |
| Blood Parameters | 7.9 mg/kg | 14-d | Increase in GGT/Broiler | 2012 | [171] |
| 5 to 200 mg/kg | 28-d | Disrupted sex hormones b-endorphin, LH, and progesterone, occurrence of renal edema and nephremia and increase levels of urea and creatinine/Layers | 2019 | [168] | |
| 0.8 mg/kg | 35-d | Decrease in T.P and GLB, and increase in ALT/Broiler | 2022 | [172] | |
| Internal organs | 0.4 mg/kg | 112-d | Increase relative weight of oviduct and ovary, degeneration and atrophy of the ovarian tissues/Layers | 2017 | [102] |
| 2 mg/kg | 42-d | ZEN residues in the kidney and liver A rise in liver weight/Broiler | 2019 | [160] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okasha, H.; Song, B.; Song, Z. Hidden Hazards Revealed: Mycotoxins and Their Masked Forms in Poultry. Toxins 2024, 16, 137. https://doi.org/10.3390/toxins16030137
Okasha H, Song B, Song Z. Hidden Hazards Revealed: Mycotoxins and Their Masked Forms in Poultry. Toxins. 2024; 16(3):137. https://doi.org/10.3390/toxins16030137
Chicago/Turabian StyleOkasha, Hamada, Bochen Song, and Zhigang Song. 2024. "Hidden Hazards Revealed: Mycotoxins and Their Masked Forms in Poultry" Toxins 16, no. 3: 137. https://doi.org/10.3390/toxins16030137
APA StyleOkasha, H., Song, B., & Song, Z. (2024). Hidden Hazards Revealed: Mycotoxins and Their Masked Forms in Poultry. Toxins, 16(3), 137. https://doi.org/10.3390/toxins16030137