High-Voltage Toxin’Roll: Electrostatic Charge Repulsion as a Dynamic Venom Resistance Trait in Pythonid Snakes




Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
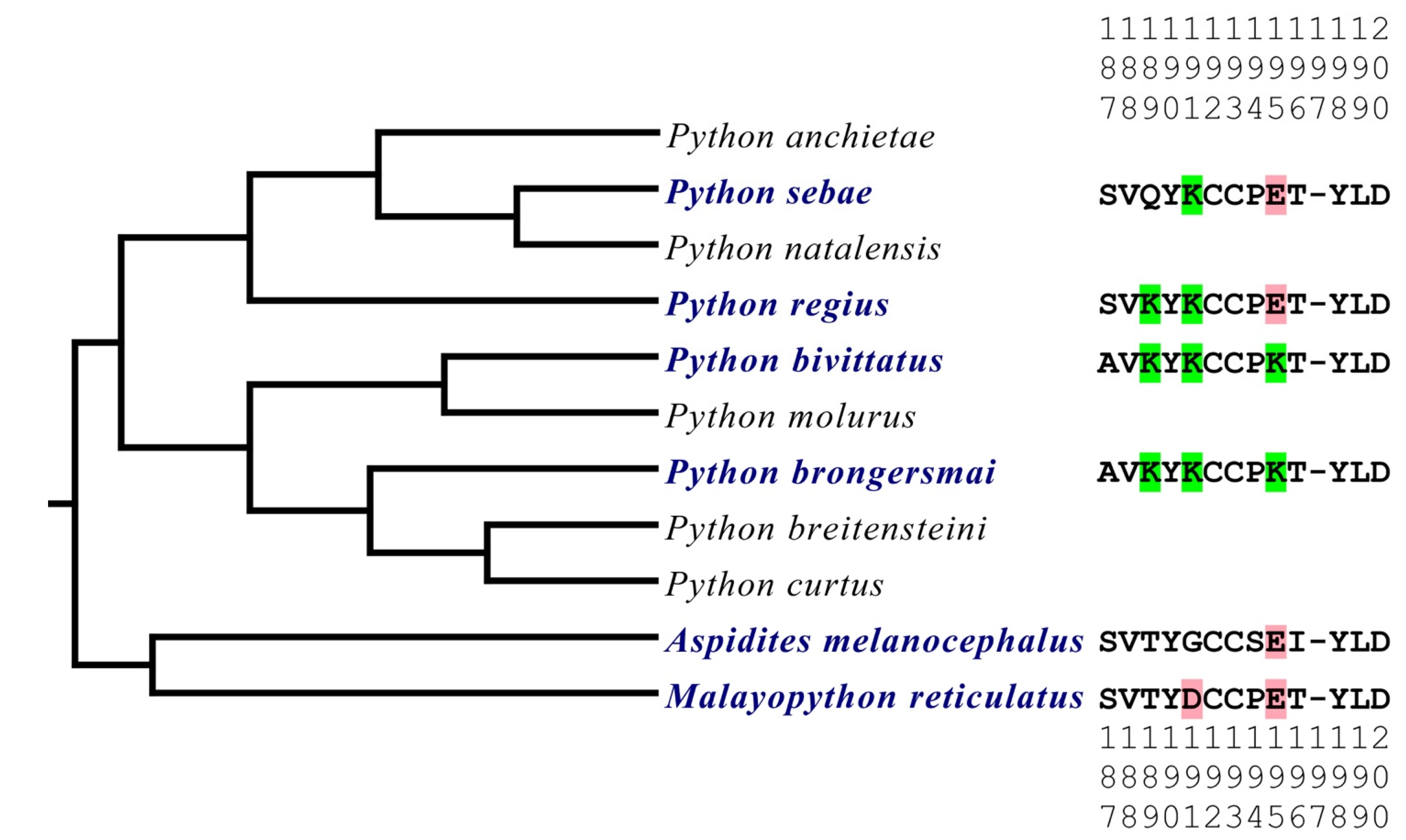
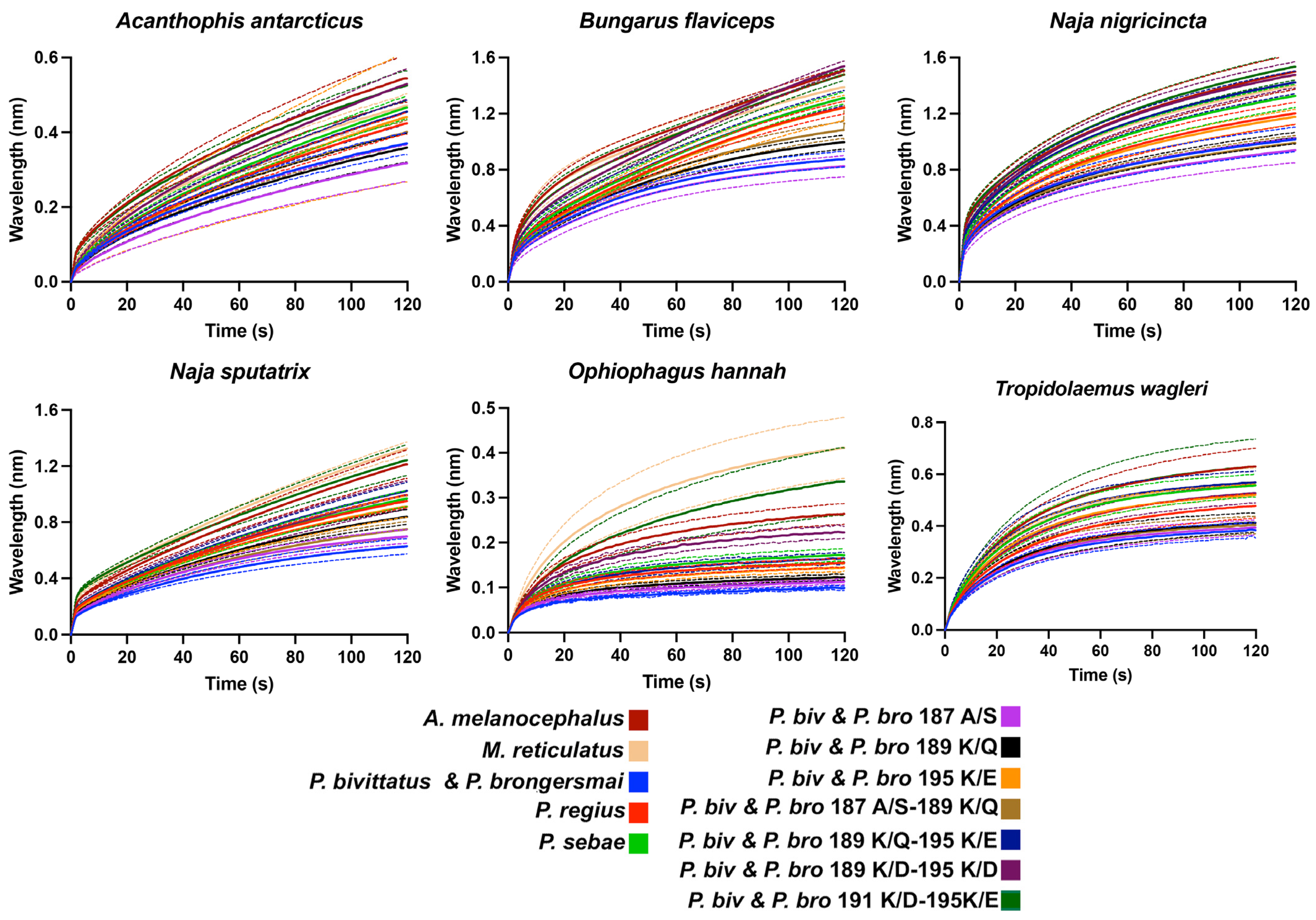
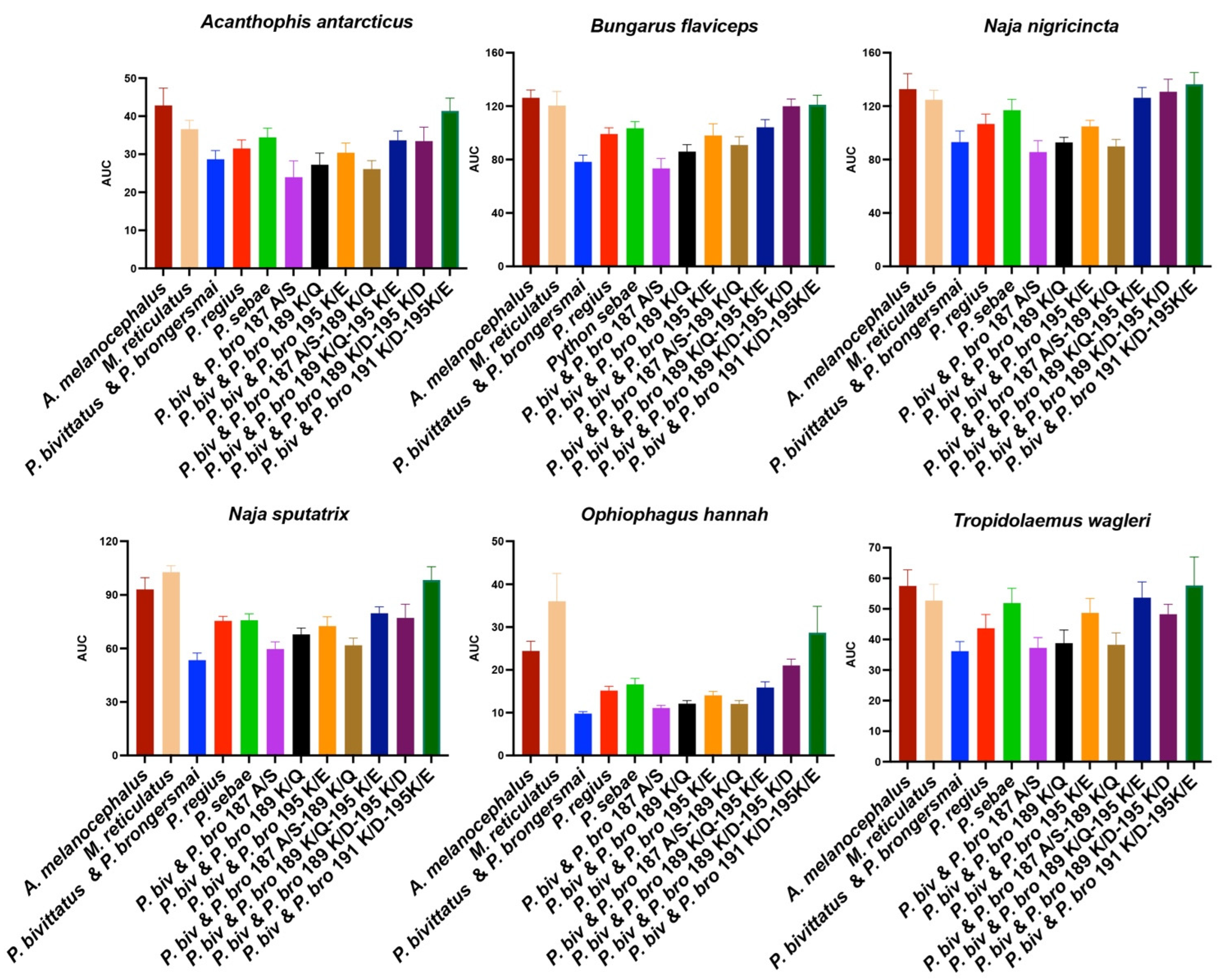
- A. melanocephalus was bound the strongest by all venoms, consistent with this species retaining both the Pythonidae family’s negatively charged amino acids (191D and 195E) and the lack of any positive charges in the orthosteric site.
- M. reticulatus was strongly bound but at a level less than A. melanocephalus, consistent with it having only one negatively charged amino acid in the orthosteric site (195E) due to the secondary loss of 191D.
- P. sebae was bound less strongly than M. reticulatus but higher than P. bivittatus, P. brongersmai, or Python regius, consistent with it retaining only one of the Python genus lysine mutations (191K), with 189K replaced by glutamine (Q).
- P. regius displayed a lower venom binding affinity than P. sebae but bound stronger than P. bivittatus and P. brongersmai, consistent with it having two lysine mutations (189K and 191K), one more than P. sebae but one fewer than P. bivittatus and P. brongersmai.
- P. bivittatus and P. brongersmai were the most resistant to binding by any of the venoms, consistent with both species having three lysine mutations (189K, 191K, and 195K).
- A stepwise replacement of the lysine residues [26] confirmed the relative role of the positively charged amino acids in the evolution of venom resistance in P. bivittatus and P. brongersmai.




3. Discussion
4. Materials and Methods
- DNeasy Blood & Tissue kit (QIAGEN, Carlsbad, CA, USA) was used to isolated the DNA, using the spin column protocol for all species, except for the P. brongersmai, whose data was extracted using the E.Z.N.A. Tissue DNA Kit (Omega Bio-tek, Norcross, GA, USA).
- In total, 25 mg of homogenized tissue samples were mixed with a lysis buffer and Proteinase K solution and 56 °C shake-incubated for 3 h. Several centrifugation steps were undertaken followed by washing with wash buffer solutions.
- Prior to the DNA extraction, the tissues were rinsed with 10% of phosphate-buffered saline (PBS) to remove the 70% ethanol preservative.
- Post-elution, the DNA concentration and purity were determined using the Nanodrop 2000 UV–VIS Spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA).
- The isolated genomic DNA was stored at −20 °C.
- A ~200 base pair range corresponding to chrna1 (muscular nAChR gene) was amplified by locus-specific primer-directed PCR.
- Primers specific for the orthosteric site of the nAChR were designed using the published Python bivittatus CHRNA1 sequence XM_007444717.2.
- ○
- Python-F = 5′ TGAATAACTACATGCCGAGTGG 3′.
- ○
- Python-R = 5′ CGTGGGTAGATAAAATACTAATCC 3′.
- The following were the PCR reaction contents:
- ○
- 25 μL of Taq PCR master mix;
- ○
- 3 μL of each primer (10 μM);
- ○
- 500 ng of DNA;
- ○
- PCR water to adjust to the 50 μL total PCR reaction volume.
- The PCR reaction conditions were as follows:
- ○
- Initial denaturation at 95 °C for 3 min (all subsequent denaturation steps were at 95 °C for 30 s);
- ○
- Annealing was at 55 °C for 30 s;
- ○
- Extension was at 72 °C for 1 min;
- ○
- The PCR steps of denaturation, annealing, and extension were repeated for 35 cycles;
- ○
- Final extension at 72 °C for 10 min.
- Sequencing of the primer-directed locus-specific amplified PCR products was undertaken at the Australian Genome Research Facility, University of Queensland, Australia, and Florida State University’s Core Facilities DNA Sequencing Laboratory, Tallahassee, Florida, using the automated dideoxy sequencing method dual-direction sequencing.
- The sequence reads were aligned and manually curated using the Aliview v.1.1 software (alignment viewer and editor) and Expasy (translate tool) to ascertain the relative absence or presence of the resistance elements in the ligand binding domain of the α1subunit of the nAChR of each of the python species tested.
- The venom samples were sourced from the long-term cryogenic collection of the Adaptive Biotoxicology Lab, University of Queensland, St Lucia, Australia.
- All the venom study protocols of this work were performed with the University of Queensland Biosafety Approval #IBC134BSBS2015 and the University of Queensland Animal Ethics Approval 2021/AE000075.
- The lyophilized crude venom samples were reconstituted with double-deionized water (ddH2O) before use. The centrifugation was performed at 14,000 RCF for 10 min with a temperature of 4 °C.
- Subsequently, the pellet (if any) was discarded, and the supernatant was used to make a working venom stock of 1 mg/mL in 50% of glycerol to preserve the enzymatic action while avoiding freezing upon storage at −20 °C.
- The concentrations of the prepared venom stocks were checked at 280 nm with a NanoDrop 2000 UV–VIS Spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA).
- Uncontrollable postsynthetic thiol oxidation was prevented by the synthetic peptides having a serine doublet in place of the cysteine doublet [44].
- The mimotopes were dissolved in 100% of dimethyl sulfoxide (DMSO) followed by a 1:10 dilution with double-deionized water in order to make 50 µg/mL of working stocks.
- All prepared mimotope stock solutions were stored at −20 °C for future use.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Casewell, N.R.; Petras, D.; Card, D.C.; Suranse, V.; Mychajliw, A.M.; Richards, D.; Koludarov, I.; Albulescu, L.O.; Slagboom, J.; Hempel, B.F.; et al. Solenodon Genome Reveals Convergent Evolution of Venom in Eulipotyphlan Mammals. Proc. Natl. Acad. Sci. USA 2019, 116, 25745–25755. [Google Scholar] [CrossRef] [PubMed]
- Kazandjian, T.D.; Petras, D.; Robinson, S.D.; van Thiel, J.; Greene, H.W.; Arbuckle, K.; Barlow, A.; Carter, D.A.; Wouters, R.M.; Whiteley, G.; et al. Convergent Evolution of Pain-Inducing Defensive Venom Components in Spitting Cobras. Science 2021, 371, 386–390. [Google Scholar] [CrossRef]
- Schendel, V.; Rash, L.D.; Jenner, R.A.; Undheim, E.A.B. The Diversity of Venom: The Importance of Behavior and Venom System Morphology in Understanding Its Ecology and Evolution. Toxins 2019, 11, 666. [Google Scholar] [CrossRef]
- Zancolli, G.; Casewell, N.R. Venom Systems as Models for Studying the Origin and Regulation of Evolutionary Novelties. Mol. Biol. Evol. 2020, 37, 2777–2790. [Google Scholar] [CrossRef] [PubMed]
- Jackson, T.N.; Fry, B.G. A Tricky Trait: Applying the Fruits of the “Function Debate” in the Philosophy of Biology to the “Venom Debate” in the Science of Toxinology. Toxins 2016, 8, 263. [Google Scholar] [CrossRef]
- Chandrasekara, U.; Harris, R.J.; Fry, B.G. The Target Selects the Toxin: Specific Amino Acids in Snake-Prey Nicotinic Acetylcholine Receptors That Are Selectively Bound by King Cobra Venoms. Toxins 2022, 14, 528. [Google Scholar] [CrossRef]
- Xie, B.; Dashevsky, D.; Rokyta, D.; Ghezellou, P.; Fathinia, B.; Shi, Q.; Richardson, M.K.; Fry, B.G. Dynamic genetic differentiation drives the widespread structural and functional convergent evolution of snake venom proteinaceous toxins. BMC Biol. 2022, 20, 4. [Google Scholar] [CrossRef]
- van Thiel, J.; Khan, M.A.; Wouters, R.M.; Harris, R.J.; Casewell, N.R.; Fry, B.G.; Kini, R.M.; Mackessy, S.P.; Vonk, F.J.; Wuster, W.; et al. Convergent Evolution Of Toxin Resistance in Animals. Biol. Rev. Camb. Philos. Soc. 2022, 97, 1823–1843. [Google Scholar] [CrossRef] [PubMed]
- Holding, M.L.; Drabeck, D.H.; Jansa, S.A.; Gibbs, H.L. Venom Resistance as a Model for Understanding the Molecular Basis of Complex Coevolutionary Adaptations. Integr. Comp. Biol. 2016, 56, 1032–1043. [Google Scholar] [CrossRef]
- Arbuckle, K.; Rodriguez de la Vega, R.C.; Casewell, N.R. Coevolution Takes The Sting Out of It: Evolutionary Biology and Mechanisms of Toxin Resistance In Animals. Toxicon 2017, 140, 118–131. [Google Scholar] [CrossRef]
- Perez, J.C.; Pichyangkul, S.; Garcia, V.E. The Resistance of Three Species of Warm-Blooded Animals to Western Diamondback Rattlesnake (Crotalus atrox) Venom. Toxicon 1979, 17, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Biardi, J.E.; Coss, R.G. Rock Squirrel (Spermophilus Variegatus) Blood Sera Affects Proteolytic and Hemolytic Activities of Rattlesnake Venoms. Toxicon 2011, 57, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, H.L.; Sanz, L.; Perez, A.; Ochoa, A.; Hassinger, A.T.B.; Holding, M.L.; Calvete, J.J. The Molecular Basis of Venom Resistance in a Rattlesnake-Squirrel Predator-Prey System. Mol. Ecol. 2020, 29, 2871–2888. [Google Scholar] [CrossRef] [PubMed]
- Barchan, D.; Kachalsky, S.; Neumann, D.; Vogel, Z.; Ovadia, M.; Kochva, E.; Fuchs, S. How the Mongoose Can Fight the Snake—The Binding-Site of the Mongoose Acetylcholine-Receptor. Proc. Natl. Acad. Sci. USA 1992, 89, 7717–7721. [Google Scholar] [CrossRef] [PubMed]
- Geffeney, S.L.; Fujimoto, E.; Brodie, E.D., 3rd; Brodie, E.D., Jr.; Ruben, P.C. Evolutionary Diversification of TTX-Resistant Sodium Channels in a Predator-Prey Interaction. Nature 2005, 434, 759–763. [Google Scholar] [CrossRef] [PubMed]
- Jansa, S.A.; Voss, R.S. Adaptive Evolution of the Venom-Targeted Vwf Protein in Opossums That Eat Pitvipers. PLoS ONE 2011, 6, e20997. [Google Scholar] [CrossRef] [PubMed]
- Tarvin, R.D.; Borghese, C.M.; Sachs, W.; Santos, J.C.; Lu, Y.; O’Connell, L.A.; Cannatella, D.C.; Harris, R.A.; Zakon, H.H. Interacting Amino Acid Replacements Allow Poison Frogs to Evolve Epibatidine Resistance. Science 2017, 357, 1261–1266. [Google Scholar] [CrossRef]
- Karageorgi, M.; Groen, S.C.; Sumbul, F.; Pelaez, J.N.; Verster, K.I.; Aguilar, J.M.; Hastings, A.P.; Bernstein, S.L.; Matsunaga, T.; Astourian, M.; et al. Genome Editing Retraces the Evolution of Toxin Resistance in the Monarch Butterfly. Nature 2019, 574, 409–412. [Google Scholar] [CrossRef]
- Barchan, D.; Ovadia, M.; Kochva, E.; Fuchs, S. The Binding-Site of the Nicotinic Acetylcholine-Receptor in Animal Species Resistant to Alpha-Bungarotoxin. Biochemistry 1995, 34, 9172–9176. [Google Scholar] [CrossRef]
- Kachalsky, S.G.; Jensen, B.S.; Barchan, D.; Fuchs, S. Two Subsites in the Binding Domain of the Acetylcholine Receptor: An Aromatic Subsite and a Proline Subsite. Proc. Natl. Acad. Sci. USA 1995, 92, 10801–10805. [Google Scholar] [CrossRef]
- Asher, O.; Jensen, B.S.; Lupu-Meiri, M.; Oron, Y.; Fuchs, S. The Mongoose Acetylcholine Receptor α-Subunit: Analysis of Glycosylation and α -Bungarotoxin Binding. FEBS Lett. 1998, 426, 212–216. [Google Scholar] [CrossRef]
- Takacs, Z.; Wilhelmsen, K.C.; Sorota, S. Snake α-neurotoxin Binding Site on the Egyptian Cobra (Naja haje) Nicotinic Acetylcholine Receptor is Conserved. Mol. Biol. Evol. 2001, 18, 1800–1809. [Google Scholar] [CrossRef]
- Takacs, Z.; Wilhelmsen, K.C.; Sorota, S. Cobra (Naja spp.) Nicotinic Acetylcholine Receptor Exhibits Resistance to Erabu Sea Snake (Laticauda semifasciata) Short-Chain α-Neurotoxin. J. Mol. Evol. 2004, 58, 516–526. [Google Scholar] [CrossRef]
- Dellisanti, C.D.; Yao, Y.; Stroud, J.C.; Wang, Z.Z.; Chen, L. Structural Determinants for Alpha-Neurotoxin Sensitivity in Muscle Nachr and Their Implications for the Gating Mechanism. Channels 2007, 1, 234–237. [Google Scholar] [CrossRef]
- Rahman, M.M.; Teng, J.; Worrell, B.T.; Noviello, C.M.; Lee, M.; Karlin, A.; Stowell, M.H.B.; Hibbs, R.E. Structure of the Native Muscle-type Nicotinic Receptor and Inhibition by Snake Venom Toxins. Neuron 2020, 106, 952–962.e955. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.J.; Fry, B.G. Electrostatic Resistance to Alpha-Neurotoxins Conferred by Charge Reversal Mutations in Nicotinic Acetylcholine Receptors. Proc. Biol. Sci./R. Soc. 2021, 288, 20202703. [Google Scholar] [CrossRef]
- Harris, R.J.; Nekaris, K.A.; Fry, B.G. Monkeying Around With Venom: An Increased Resistance to Alpha-Neurotoxins Supports an Evolutionary Arms Race Between Afro-Asian Primates And Sympatric Cobras. BMC Biol. 2021, 19, 253. [Google Scholar] [CrossRef] [PubMed]
- Jones, L.; Harris, R.J.; Fry, B.G. Not Goanna Get Me: Mutations in the Savannah Monitor Lizard (Varanus exanthematicus) Nicotinic Acetylcholine Receptor Confer Reduced Susceptibility to Sympatric Cobra Venoms. Neurotox. Res. 2021, 39, 1116–1122. [Google Scholar] [CrossRef] [PubMed]
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; et al. The Toxicogenomic Multiverse: Convergent Recruitment of Proteins Into Animal Venoms. Annu. Rev. Genom. Hum. Genet. 2009, 10, 483–511. [Google Scholar] [CrossRef]
- Dashevsky, D.; Debono, J.; Rokyta, D.; Nouwens, A.; Josh, P.; Fry, B.G. Three-Finger Toxin Diversification in the Venoms of Cat-Eye Snakes (Colubridae: Boiga). J. Mol. Evol. 2018, 86, 531–545. [Google Scholar] [CrossRef]
- Dashevsky, D.; Fry, B.G. Ancient Diversification of Three-Finger Toxins in Micrurus Coral Snakes. J. Mol. Evol. 2018, 86, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Fry, B.G.; Lumsden, N.G.; Wuster, W.; Wickramaratna, J.C.; Hodgson, W.C.; Kini, R.M. Isolation of a Neurotoxin (Alpha-Colubritoxin) From a Nonvenomous Colubrid: Evidence for Early Origin of Venom in Snakes. J. Mol. Evol. 2003, 57, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Fry, B.G.; Scheib, H.; van der Weerd, L.; Young, B.; McNaughtan, J.; Ramjan, S.F.; Vidal, N.; Poelmann, R.E.; Norman, J.A. Evolution of an Arsenal: Structural and Functional Diversification of the Venom System in the Advanced Snakes (Caenophidia). Mol. Cell. Proteomics 2008, 7, 215–246. [Google Scholar] [CrossRef] [PubMed]
- Fry, B.G.; Wuster, W.; Kini, R.M.; Brusic, V.; Khan, A.; Venkataraman, D.; Rooney, A.P. Molecular Evolution and Phylogeny of Elapid Snake Venom Three-Finger Toxins. J. Mol. Evol. 2003, 57, 110–129. [Google Scholar] [CrossRef] [PubMed]
- Fry, B.G.; Wuster, W.; Ramjan, S.F.R.; Jackson, T.; Martelli, P.; Kini, R.M. Analysis of Colubroidea Snake Venoms by Liquid Chromatography with Mass Spectrometry: Evolutionary and Toxinological Implications. Rapid Commun. Mass Spectrom. 2003, 17, 2047–2062. [Google Scholar] [CrossRef]
- Solé, R. Revisiting Leigh Van Valen’s “A New Evolutionary Law” (1973). Biol. Theory 2022, 17, 120–125. [Google Scholar] [CrossRef]
- Drabeck, D.H.; Holt, J.; McGaugh, S.E. Widespread Convergent Evolution of Alpha-Neurotoxin Resistance in African Mammals. Biol. Lett. 2022, 18, 20220361. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Dashevsky, D.; Kerkkamp, H.; Kordis, D.; de Bakker, M.A.G.; Wouters, R.; van Thiel, J.; Op den Brouw, B.; Vonk, F.; Kini, R.M.; et al. Widespread Evolution of Molecular Resistance to Snake Venom Alpha-Neurotoxins in Vertebrates. Toxins 2020, 12, 638. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, M.; Zaman, S.; Maddock, S.T.; Kamei, R.G.; Salazar-Valenzuela, D.; Wilkinson, M.; Roelants, K.; Fry, B.G. Resistance Is Not Futile: Widespread Convergent Evolution of Resistance to Alpha-Neurotoxic Snake Venoms in Caecilians (Amphibia: Gymnophiona). Int. J. Mol. Sci. 2023, 24, 11353. [Google Scholar] [CrossRef]
- Drabeck, D.H.; Dean, A.M.; Jansa, S.A. Why the Honey Badger Don’t Care: Convergent Evolution of Venom-Targeted Nicotinic Acetylcholine Receptors in Mammals That Survive Venomous Snake Bites. Toxicon 2015, 99, 68–72. [Google Scholar] [CrossRef]
- Gowri Shankar, P.; Swamy, P.; Williams, R.C.; Ganesh, S.R.; Moss, M.; Hoglund, J.; Das, I.; Sahoo, G.; Vijayakumar, S.P.; Shanker, K.; et al. King or Royal Family? Testing for Species Boundaries in the King Cobra, Ophiophagus hannah (Cantor, 1836), Using Morphology and Multilocus DNA Analyses. Mol. Phylogenet. Evol. 2021, 165, 107300. [Google Scholar] [CrossRef] [PubMed]
- Zdenek, C.N.; Harris, R.J.; Kuruppu, S.; Youngman, N.J.; Dobson, J.S.; Debono, J.; Khan, M.; Smith, I.; Yarski, M.; Harrich, D.; et al. A Taxon-Specific and High-Throughput Method for Measuring Ligand Binding to Nicotinic Acetylcholine Receptors. Toxins 2019, 11, 600. [Google Scholar] [CrossRef] [PubMed]
- Esquerre, D.; Brennan, I.G.; Donnellan, S.; Keogh, J.S. Evolutionary Models Demonstrate Rapid and Adaptive Diversification Of Australo-Papuan Pythons. Biol. Lett. 2022, 18, 20220360. [Google Scholar] [CrossRef] [PubMed]
- Bracci, L.; Lozzi, L.; Lelli, B.; Pini, A.; Neri, P. Mimotopes of the Nicotinic Receptor Binding Site Selected by a Combinatorial Peptide Library. Biochemistry 2001, 40, 6611–6619. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, M. The Book of Snakes: A Life-Size Guide to Six Hundred Species from around the World; University of Chicago Press: Chicago, IL, USA, 2018. [Google Scholar]
- Shine, R. Australian Snakes: A Natural History; Cornell University Press: Ithaca, NY, USA, 1995. [Google Scholar]
- Harris, R.J.; Youngman, N.J.; Chan, W.; Bosmans, F.; Cheney, K.L.; Fry, B.G. Getting Stoned: Characterisation of the Coagulotoxic and Neurotoxic Effects of Reef Stonefish (Synanceia verrucosa) Venom. Toxicol. Lett. 2021, 346, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.J.; Youngman, N.J.; Zdenek, C.N.; Huynh, T.M.; Nouwens, A.; Hodgson, W.C.; Harrich, D.; Dunstan, N.; Portes-Junior, J.A.; Fry, B.G. Assessing the Binding of Venoms from Aquatic Elapids to the Nicotinic Acetylcholine Receptor Orthosteric Site of Different Prey Models. Int. J. Mol. Sci. 2020, 21, 7377. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.J.; Zdenek, C.N.; Debono, J.; Harrich, D.; Fry, B.G. Evolutionary Interpretations of Nicotinic Acetylcholine Receptor Targeting Venom Effects by a Clade of Asian Viperidae Snakes. Neurotox. Res. 2020, 38, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.J.; Zdenek, C.N.; Harrich, D.; Frank, N.; Fry, B.G. An Appetite for Destruction: Detecting Prey-Selective Binding of alpha-Neurotoxins in the Venom of Afro-Asian Elapids. Toxins 2020, 12, 205. [Google Scholar] [CrossRef]
- Harris, R.J.; Zdenek, C.N.; Nouwens, A.; Sweeney, C.; Dunstan, N.; Fry, B.G. A Symmetry or Asymmetry: Functional and Compositional Comparison of Venom From the Left and Right Glands of the Indochinese Spitting Cobra (Naja siamensis). Toxicon X 2020, 7, 100050. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chandrasekara, U.; Broussard, E.M.; Rokyta, D.R.; Fry, B.G. High-Voltage Toxin’Roll: Electrostatic Charge Repulsion as a Dynamic Venom Resistance Trait in Pythonid Snakes. Toxins 2024, 16, 176. https://doi.org/10.3390/toxins16040176
Chandrasekara U, Broussard EM, Rokyta DR, Fry BG. High-Voltage Toxin’Roll: Electrostatic Charge Repulsion as a Dynamic Venom Resistance Trait in Pythonid Snakes. Toxins. 2024; 16(4):176. https://doi.org/10.3390/toxins16040176
Chicago/Turabian StyleChandrasekara, Uthpala, Emilie M. Broussard, Darin R. Rokyta, and Bryan G. Fry. 2024. "High-Voltage Toxin’Roll: Electrostatic Charge Repulsion as a Dynamic Venom Resistance Trait in Pythonid Snakes" Toxins 16, no. 4: 176. https://doi.org/10.3390/toxins16040176
APA StyleChandrasekara, U., Broussard, E. M., Rokyta, D. R., & Fry, B. G. (2024). High-Voltage Toxin’Roll: Electrostatic Charge Repulsion as a Dynamic Venom Resistance Trait in Pythonid Snakes. Toxins, 16(4), 176. https://doi.org/10.3390/toxins16040176