Patulin Biodegradation Mechanism Study in Pichia guilliermondii S15-8 Based on PgSDR-A5D9S1


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
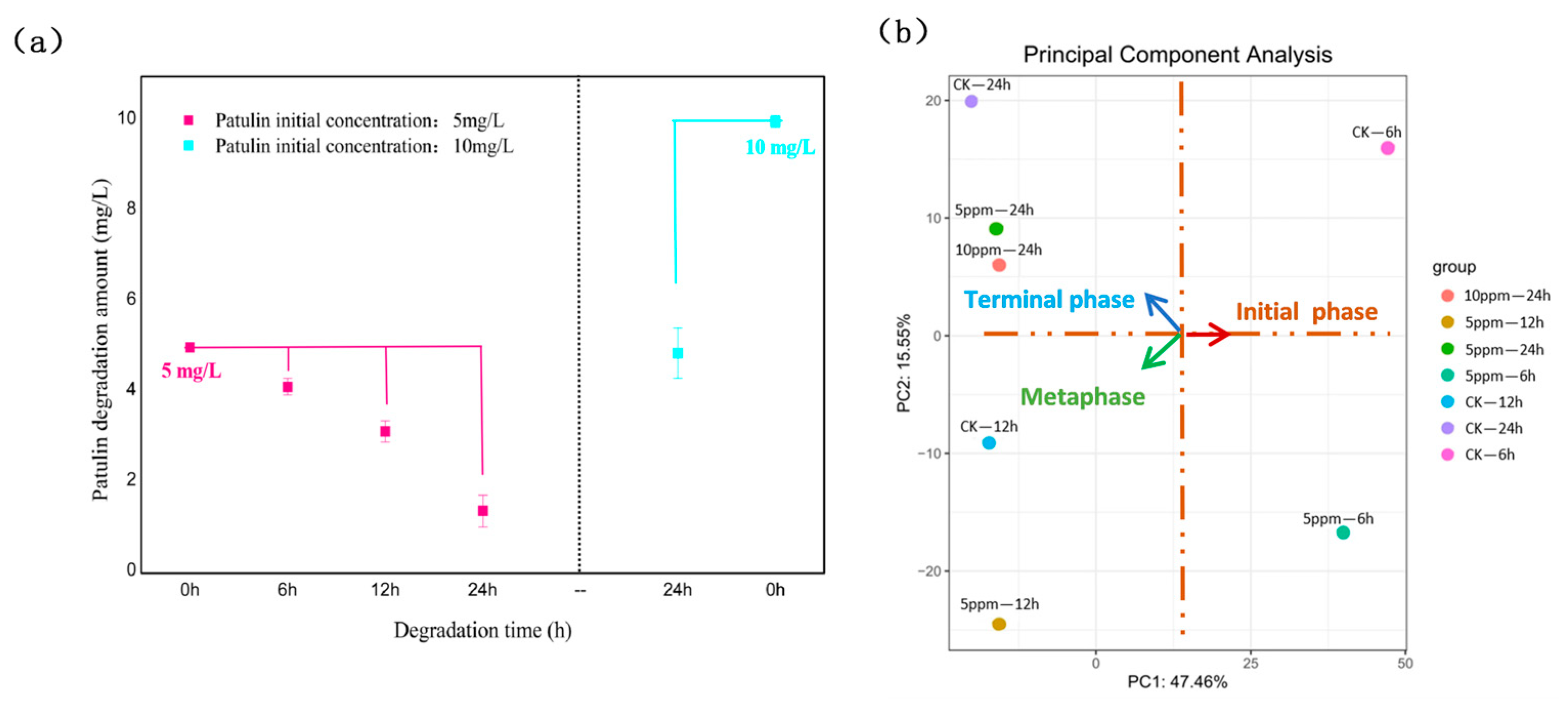
2.1. Patulin Degradation at Different Times and Concentrations
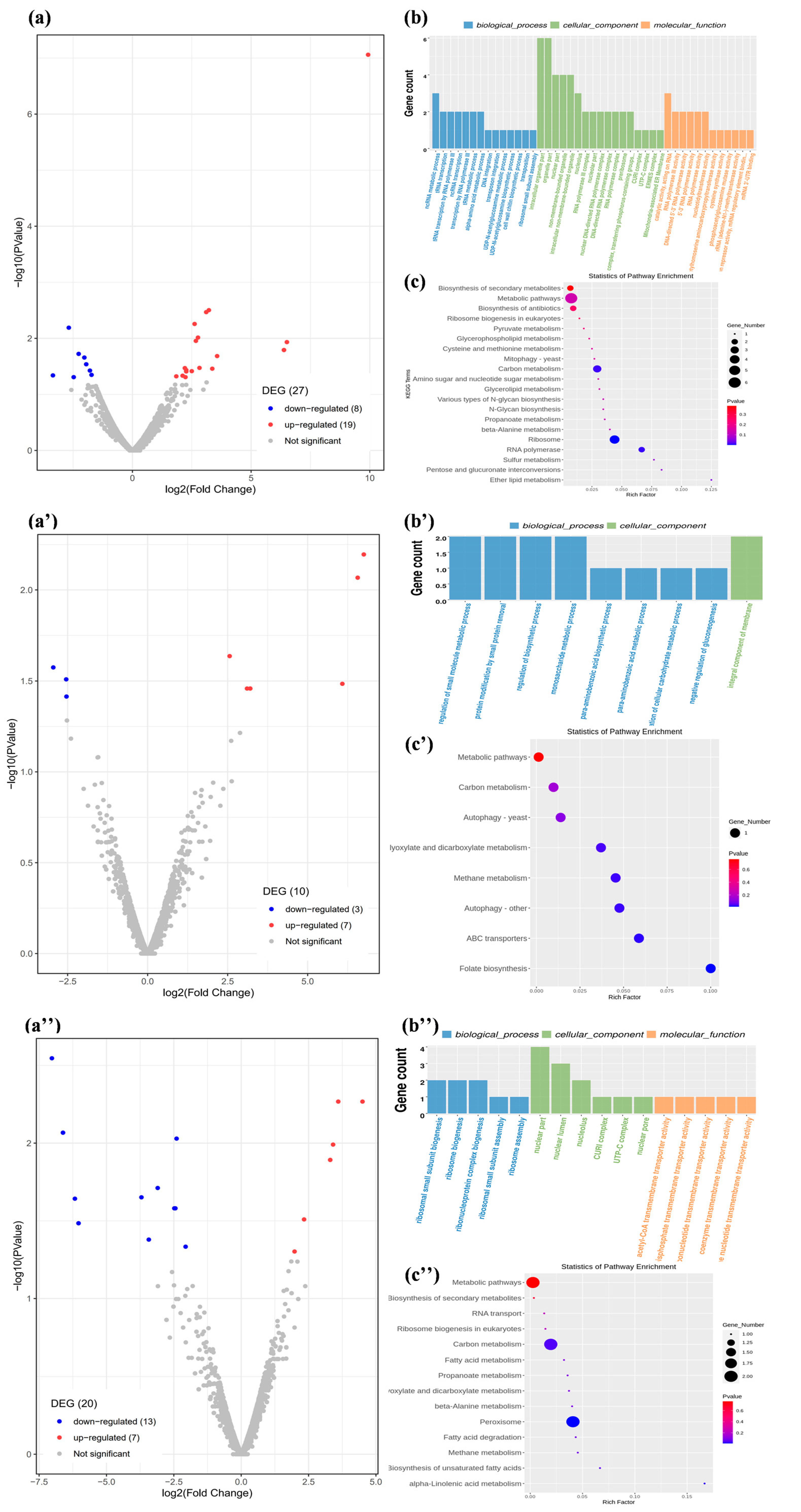
2.2. Function and Metabolic Pathway Enrichment Analysis of DEGs
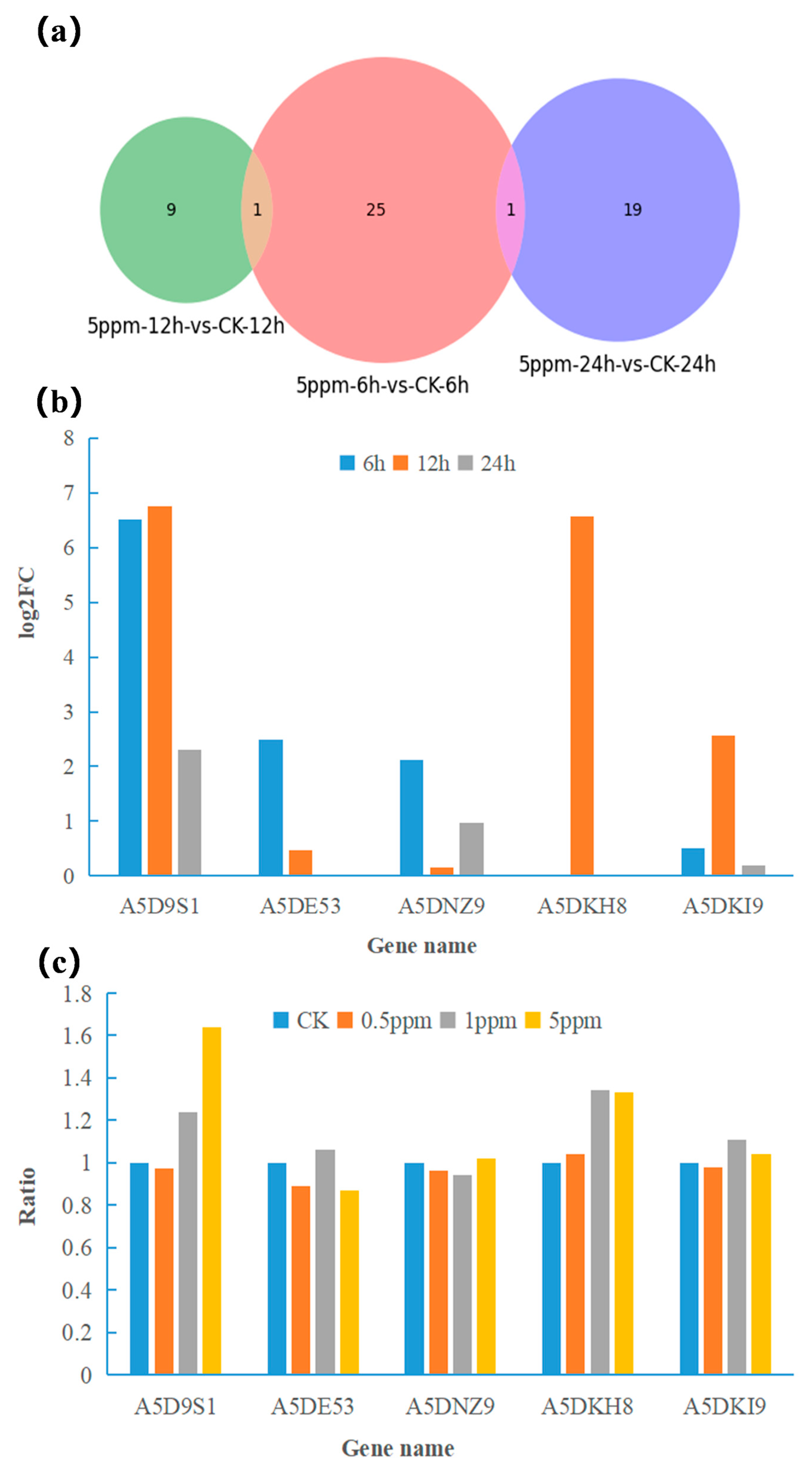
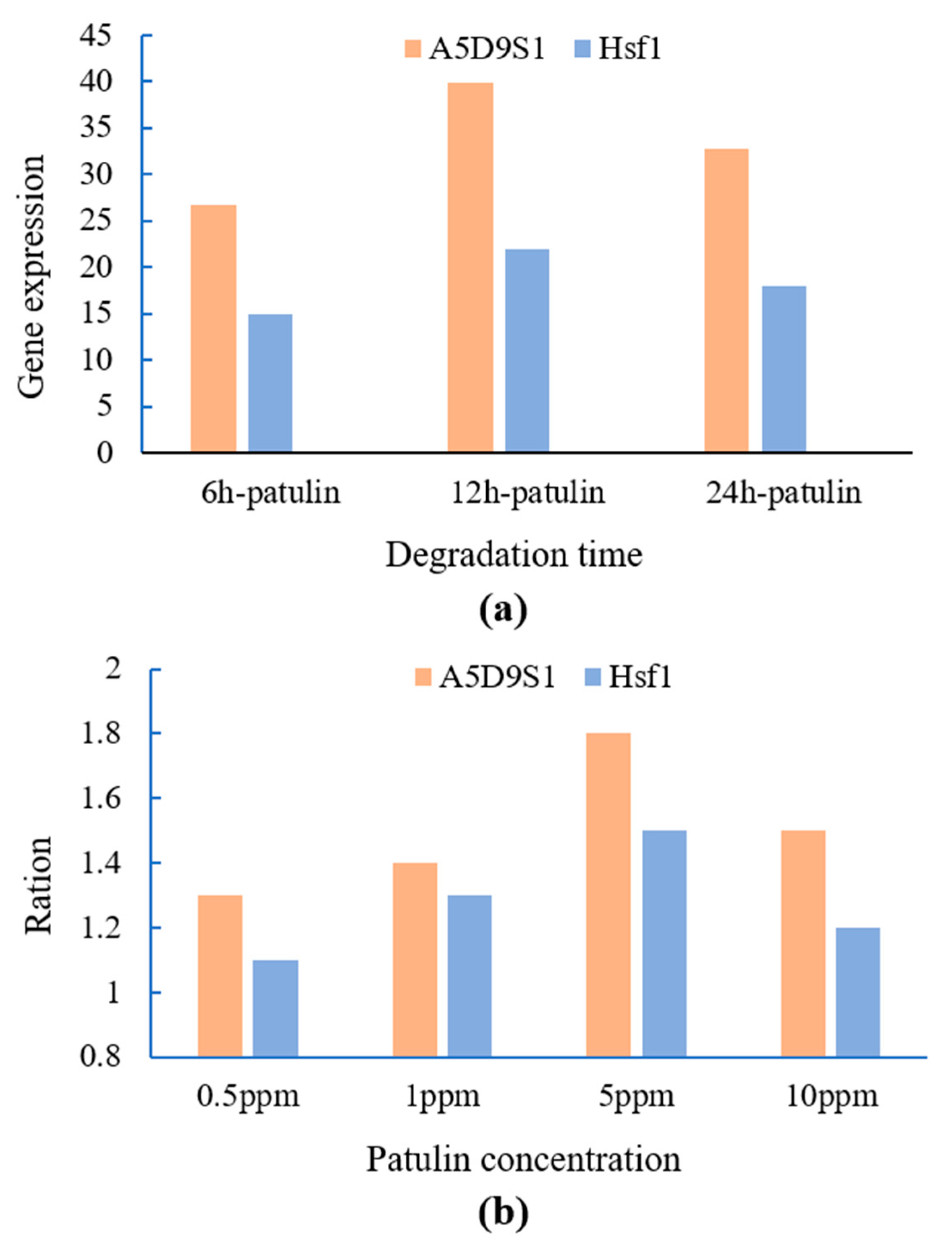
2.3. qRT-PCR Verification
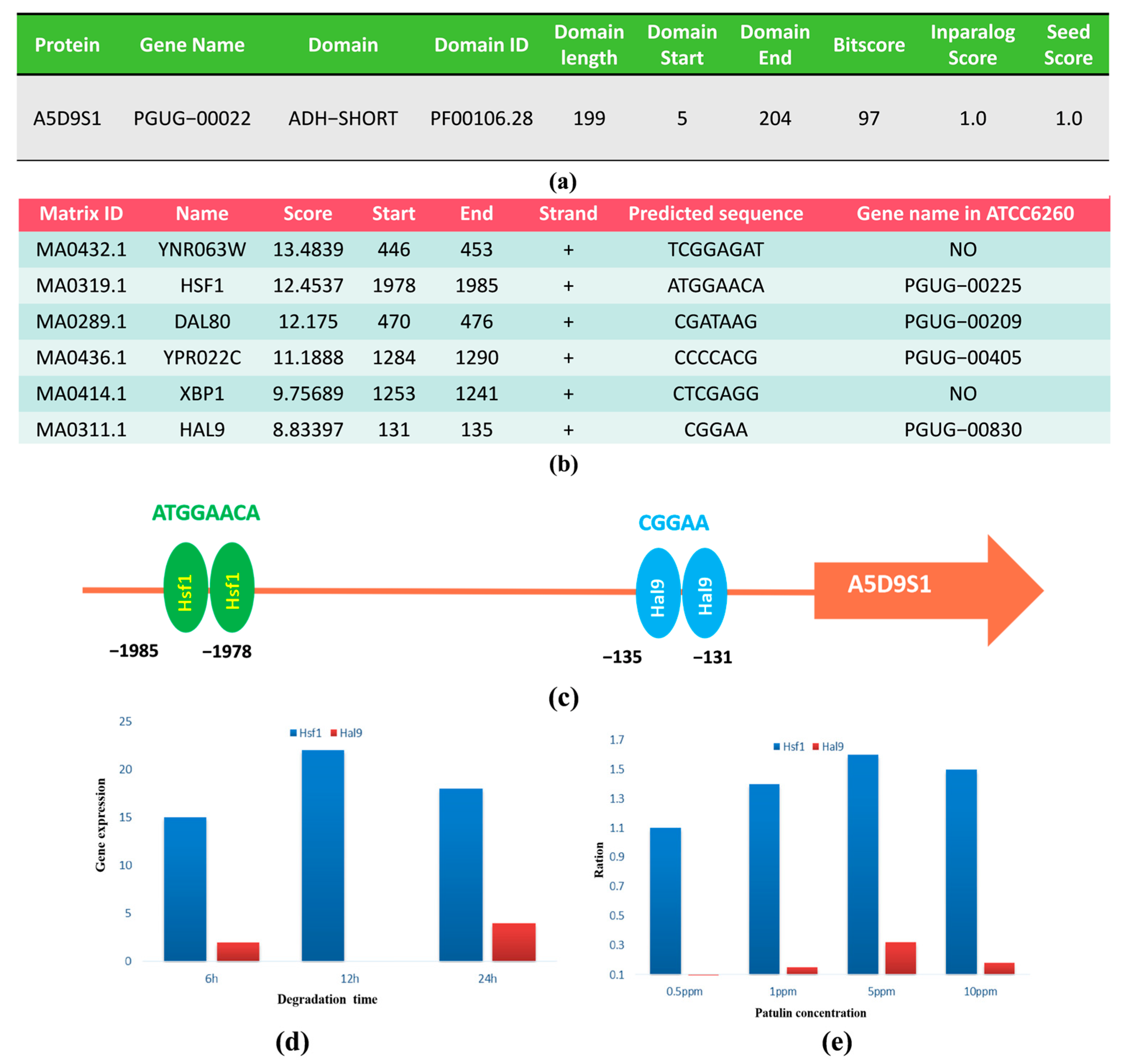
2.4. Gene A5D9S1 Characteristics and Regulatory Factor Prediction
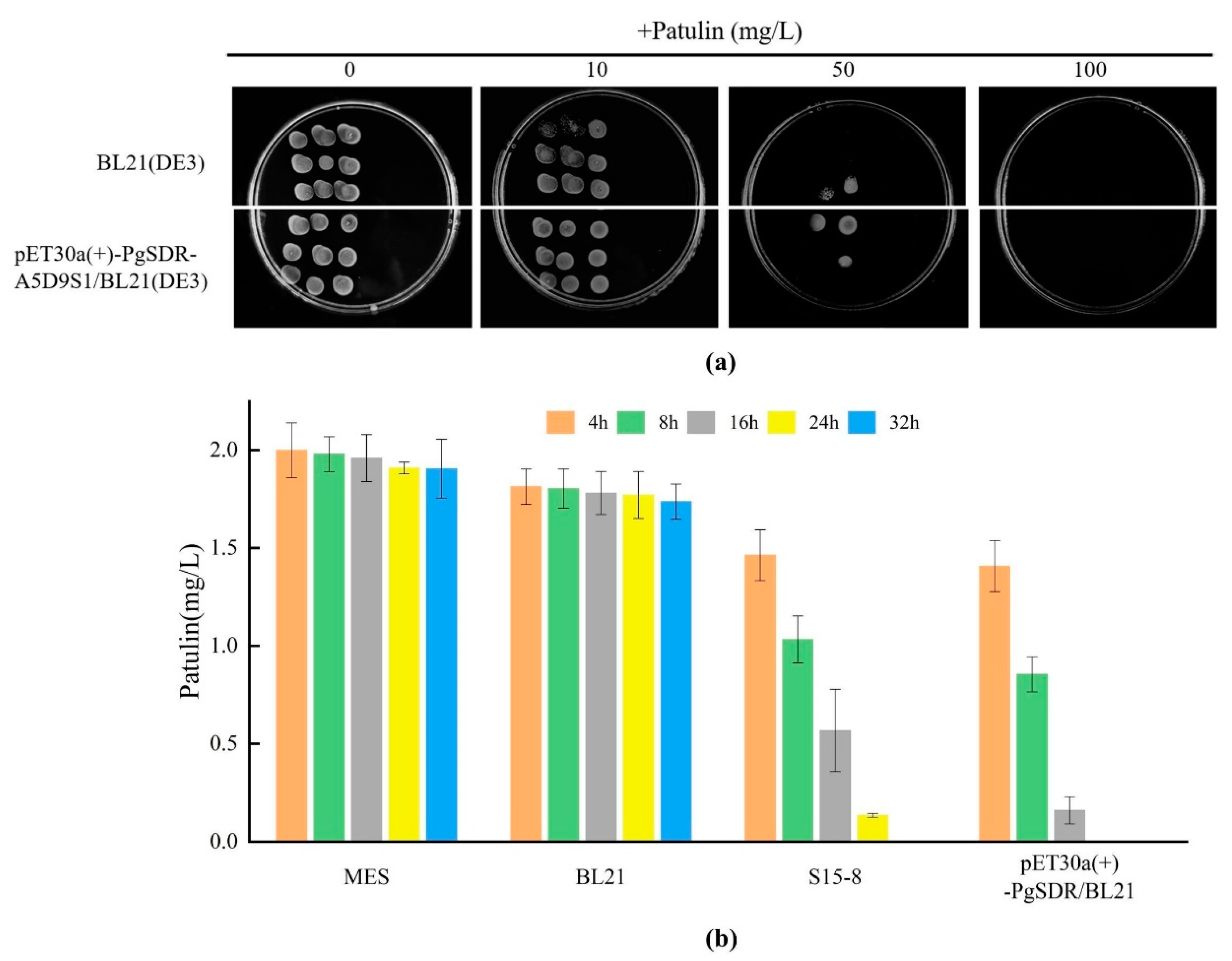
2.5. Patulin Degradation Ability for PgSDR-A5D9S1-Expressed E. coli
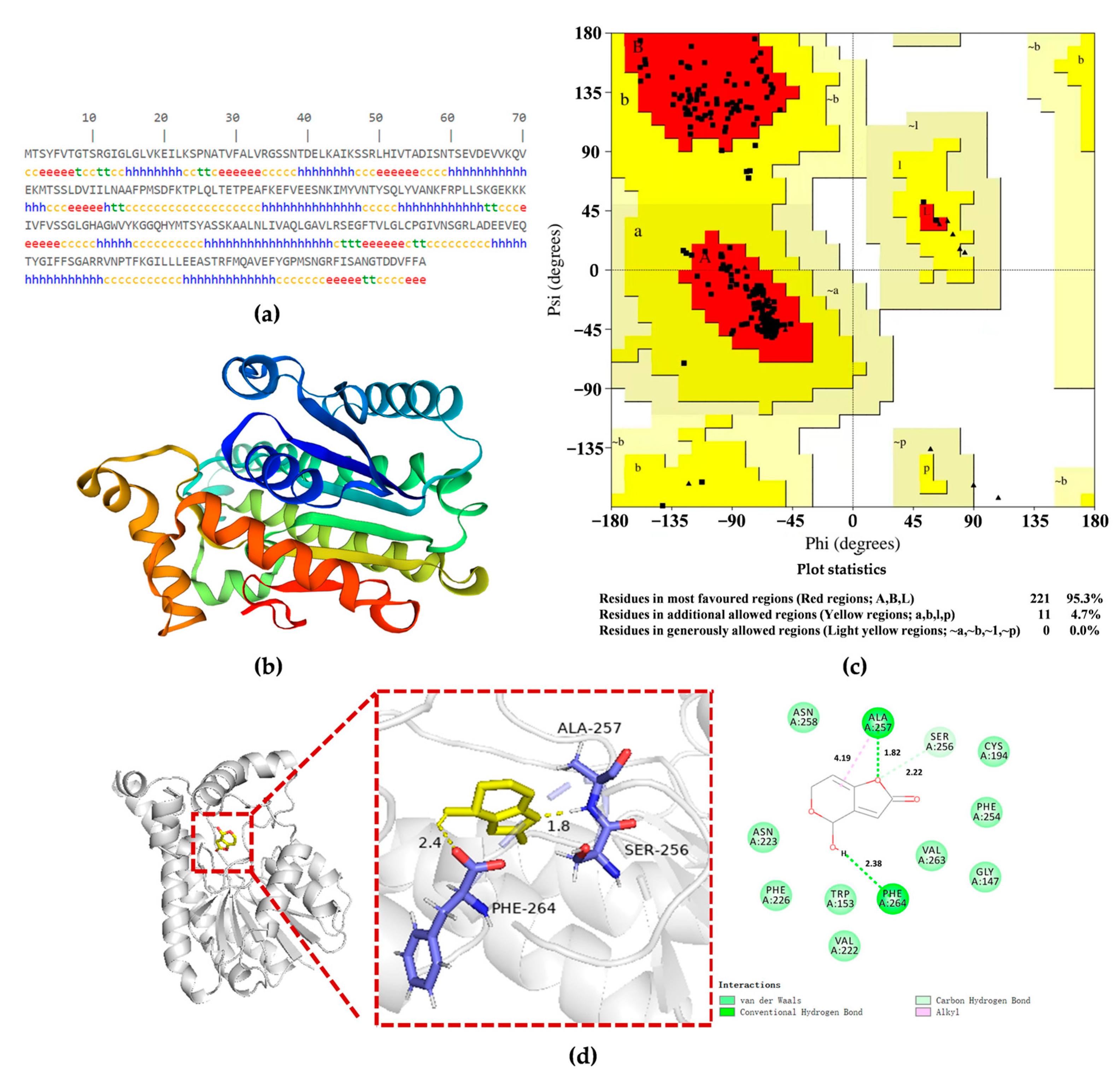
2.6. PgSDR-A5D9S1 Structure Prediction and PgSDR-Patulin Docking
3. Conclusions
4. Materials and Methods
4.1. Patulin and Yeast Strain Preparation
4.2. Patulin Degradation and Quantification
4.3. RNA Extraction and Sequencing
4.4. Enrichment Analysis and qRT-PCR Verification
4.5. Target Gene Characteristics and Regulatory Factor Prediction
4.6. Target Gene Clone, Plasmid Construction, and Transformation
4.7. Patulin Sensitivity and Degradation Assay in Target Gene Expressed E. coli Strain
4.8. Three-Dimensional (3D) Structure Prediction and Molecular Docking of Patulin Degradation Protein
4.9. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Barad, S.; Sionov, E.; Prusky, D. Role of patulin in post-harvest diseases. Fungal Biol. Rev. 2016, 30, 24–32. [Google Scholar] [CrossRef]
- Sajid, M.; Mehmood, S.; Yuan, Y.; Yue, T. Mycotoxin patulin in food matrices: Occurrence and its biological degradation strategies. Drug Metab. Rev. 2019, 51, 1–48. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, A.; Babaali, E.; Berizi, E. Effect of radiation, heating, high pressure, and the commercial processing method on reduction and/or elimination of patulin in fruit and vegetable products: A systematic review. Toxin Rev. 2021, 40, 338–346. [Google Scholar] [CrossRef]
- Ma, D.; Ji, D.; Liu, J.; Xu, Y.; Chen, T.; Tian, S. Efficacy of methyl thujate in inhibiting Penicillium expansum growth and possible mechanism involved. Postharvest Biol. Technol. 2020, 161, 111070. [Google Scholar] [CrossRef]
- Zheng, X.; Wei, W.; Zhou, W.; Li, H.; Rao, S.; Gao, L.; Yang, Z. Prevention and detoxification of patulin in apple and its products: A review. Food Res. Int. 2021, 140, 110034. [Google Scholar] [CrossRef]
- Zhu, R.; Wang, W.; Luo, Z.; Lin, H.; Li, Y.; Lu, W.; Xu, Z.; Cai, C.; Hu, S. Blue LED light treatment inhibits virulence and patulin biosynthesis in Penicillium expansum. Postharvest Biol. Technol. 2023, 200, 112340. [Google Scholar] [CrossRef]
- Zhang, X.; Li, B.; Zhang, Z.; Chen, Y.; Tian, S. Antagonistic Yeasts: A Promising Alternative to Chemical Fungicides for Controlling Postharvest Decay of Fruit. J. Fungi 2020, 6, 158. [Google Scholar] [CrossRef]
- Cao, J.; Zhang, H.; Yang, Q.; Ren, R. Efficacy of Pichia caribbica in controlling blue mold rot and patulin degradation in apples. Int. J. Food Microbiol. 2013, 162, 167–173. [Google Scholar] [CrossRef]
- Li, B.; Chen, Y.; Zhang, Z.; Qin, G.; Chen, T.; Tian, S. Molecular basis and regulation of pathogenicity and patulin biosynthesis in Penicillium expansum. Compr. Rev. Food Sci. Food Saf. 2020, 19, 3416–3438. [Google Scholar] [CrossRef]
- Yang, C.; Peng, B. Biodegradation characteristics of patulin by Saccharomyces cerevisiae during fermentation. Food Control 2023, 145, 109463. [Google Scholar] [CrossRef]
- Yang, Y.; Ji, J.; Wu, S.; Ye, Y.; Sheng, L.; Zhang, Y.; Sun, X. Efficient Biodegradation of Patulin by Aspergillus niger FS10 and Metabolic Response of Degrading Strain. Foods 2023, 12, 382. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Zheng, L.; Xia, F.; Wei, W.; Wang, S.; Rao, S.; Gao, L.; Yang, Z. In vivo evaluation of the toxicity of patulin degradation products produced by Lactobacillus casei YZU01. Biol. Control 2022, 169, 104878. [Google Scholar] [CrossRef]
- Xing, M.; Chen, Y.; Li, B.; Tian, S. Characterization of a short-chain dehydrogenase/reductase and its function in patulin biodegradation in apple juice. Food Chem. 2021, 348, 129046. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Liu, L.; Yuan, L.; Li, J.; Wang, X. The characteristics of patulin degradation by probiotic yeast—Pichia guilliermondii S15-8. Food Control 2022, 133, 108627. [Google Scholar] [CrossRef]
- Hahn, J.S.; Hu, Z.; Thiele, D.J.; Iyer, V.R. Genome-wide analysis of the biology of stress responses through heat shock transcription factor. Mol. Cell. Biol. 2004, 24. [Google Scholar] [CrossRef] [PubMed]
- Brandman, O.; Stewart-Ornstein, J.; Wong, D.; Larson, A.; Williams, C.C.; Li, G.-W.; Zhou, S.; King, D.; Shen, P.S.; Weibezahn, J.; et al. A Ribosome-Bound Quality Control Complex Triggers Degradation of Nascent Peptides and Signals Translation Stress. Cell 2012, 151, 1042–1054. [Google Scholar] [CrossRef] [PubMed]
- Cui, D.; Liu, L.; Sun, L.; Lin, X.; Lin, L.; Zhang, C. Genome-wide analysis reveals Hsf1 maintains high transcript abundance of target genes controlled by strong constitutive promoter in Saccharomyces cerevisiae. Biotechnol. Biofuels Bioprod. 2023, 16, 72. [Google Scholar] [CrossRef]
- Wang, H.; Li, Q.; Kuang, X.; Xiao, D.; Han, X.; Hu, X.; Li, X.; Ma, M. Functions of aldehyde reductases from Saccharomyces cerevisiae in detoxification of aldehyde inhibitors and their biotechnological applications. Appl. Microbiol. Biotechnol. 2018, 102, 10439–10456. [Google Scholar] [CrossRef]
- Liu, Z.L. Understanding the tolerance of the industrial yeast Saccharomyces cerevisiae against a major class of toxic aldehyde compounds. Appl. Microbiol. Biotechnol. 2018, 102, 5369–5390. [Google Scholar] [CrossRef]
- Du, G.; Zhang, X.; Gao, Y.; Sun, C.; Wang, L.; Zhao, W.; Meng, D.; Guan, W.; Zhao, H. Heat shock transcriptional factor HSF1 is activated by phosphorylation in response to ginger oleoresin stress in S. cerevisiae. LWT 2023, 184, 115116. [Google Scholar] [CrossRef]
- Studer, G.; Rempfer, C.; Waterhouse, A.M.; Gumienny, R.; Haas, J.; Schwede, T. QMEANDisCo-distance constraints applied on model quality estimation. Bioinformatics 2020, 36, 1765–1771. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.; Chudasama, R.; Solanki, R.; Patel, P.; Parmar, K.; Munshi, N.S. Structure prediction and molecular docking studies of aromatic hydrocarbon sensing proteins TbuT, HbpR and PhnR to detect priority pollutants. J. Environ. Sci. Health Part A Tox. Hazard. Subst. Environ. Eng. 2020, 55, 126–141. [Google Scholar] [CrossRef]
- Luo, Y.; Wang, Z.L.; Yuan, Y.H.; Zhou, Z.K.; Yue, T.L. Patulin adsorption of a superior microorganism strain with low flavour-affection of kiwi fruit juice. World Mycotoxin J. 2016, 9, 195–203. [Google Scholar] [CrossRef]
- Gene Ontology Consortium; Aleksander, S.A.; Balhoff, J.; Carbon, S.; Cherry, J.M.; Drabkin, H.J.; Ebert, D.; Feuermann, M.; Gaudet, P.; Harris, N.L.; et al. The Gene Ontology knowledgebase in 2023. Genetics 2023, 224, iyad031. [Google Scholar] [CrossRef] [PubMed]
- Huckvale, E.; Moseley, H.N.B. kegg_pull: A software package for the RESTful access and pulling from the Kyoto Encyclopedia of Gene and Genomes. BMC Bioinform. 2023, 24, 78. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Deng, X.; Luo, Y.; Lu, M.; Guan, T.; Li, Y.; Guo, X. Unraveling the Mechanism of Zhibaidihuang Decoction against IgA Nephropathy Using Network Pharmacology and Molecular Docking Analyses. Tohoku J. Exp. Med. 2023, 259, 37–47. [Google Scholar] [CrossRef]
- Kp, A.D.; Shimoga Janakirama, A.R.; Martin, A. SIRT1 activation by Taurine: In vitro evaluation, molecular docking and molecular dynamics simulation studies. J. Nutr. Biochem. 2022, 102, 108948. [Google Scholar] [CrossRef]
- Tang, Y.; Zhou, Y.; Zhao, Q.; Zhao, Q.; Wang, Z. Enzymatic synthesis of an orlistat intermediate using a mutant short-chain dehydrogenase from Novosphingobium aromaticivorans. Process Biochem. 2020, 95, 235–241. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xi, H.; Wang, Y.; Ni, X.; Zhang, M.; Luo, Y. Patulin Biodegradation Mechanism Study in Pichia guilliermondii S15-8 Based on PgSDR-A5D9S1. Toxins 2024, 16, 177. https://doi.org/10.3390/toxins16040177
Xi H, Wang Y, Ni X, Zhang M, Luo Y. Patulin Biodegradation Mechanism Study in Pichia guilliermondii S15-8 Based on PgSDR-A5D9S1. Toxins. 2024; 16(4):177. https://doi.org/10.3390/toxins16040177
Chicago/Turabian StyleXi, Huijuan, Yebo Wang, Xulei Ni, Minjie Zhang, and Ying Luo. 2024. "Patulin Biodegradation Mechanism Study in Pichia guilliermondii S15-8 Based on PgSDR-A5D9S1" Toxins 16, no. 4: 177. https://doi.org/10.3390/toxins16040177
APA StyleXi, H., Wang, Y., Ni, X., Zhang, M., & Luo, Y. (2024). Patulin Biodegradation Mechanism Study in Pichia guilliermondii S15-8 Based on PgSDR-A5D9S1. Toxins, 16(4), 177. https://doi.org/10.3390/toxins16040177