Dynamics of the Toxin Cylindrospermopsin and the Cyanobacterium Chrysosporum (Aphanizomenon) ovalisporum in a Mediterranean Eutrophic Reservoir


Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Chrysosporum ovalisporum |
|---|---|
| Laboratory optimal growth temperature (°C) | 28 ± 2 a |
| 33 ± 2 b | |
| 32.8 ± 0.9 c | |
| 26 ± 1 d | |
| Maximum growth rate at optimal temperature (day−1) | 0.3 a |
| 0.36 c | |
| Filament flotation rate (m h−1) | <0.04 e |
| Optimal solar irradiation (W m−2) | 80 a |
2. Results
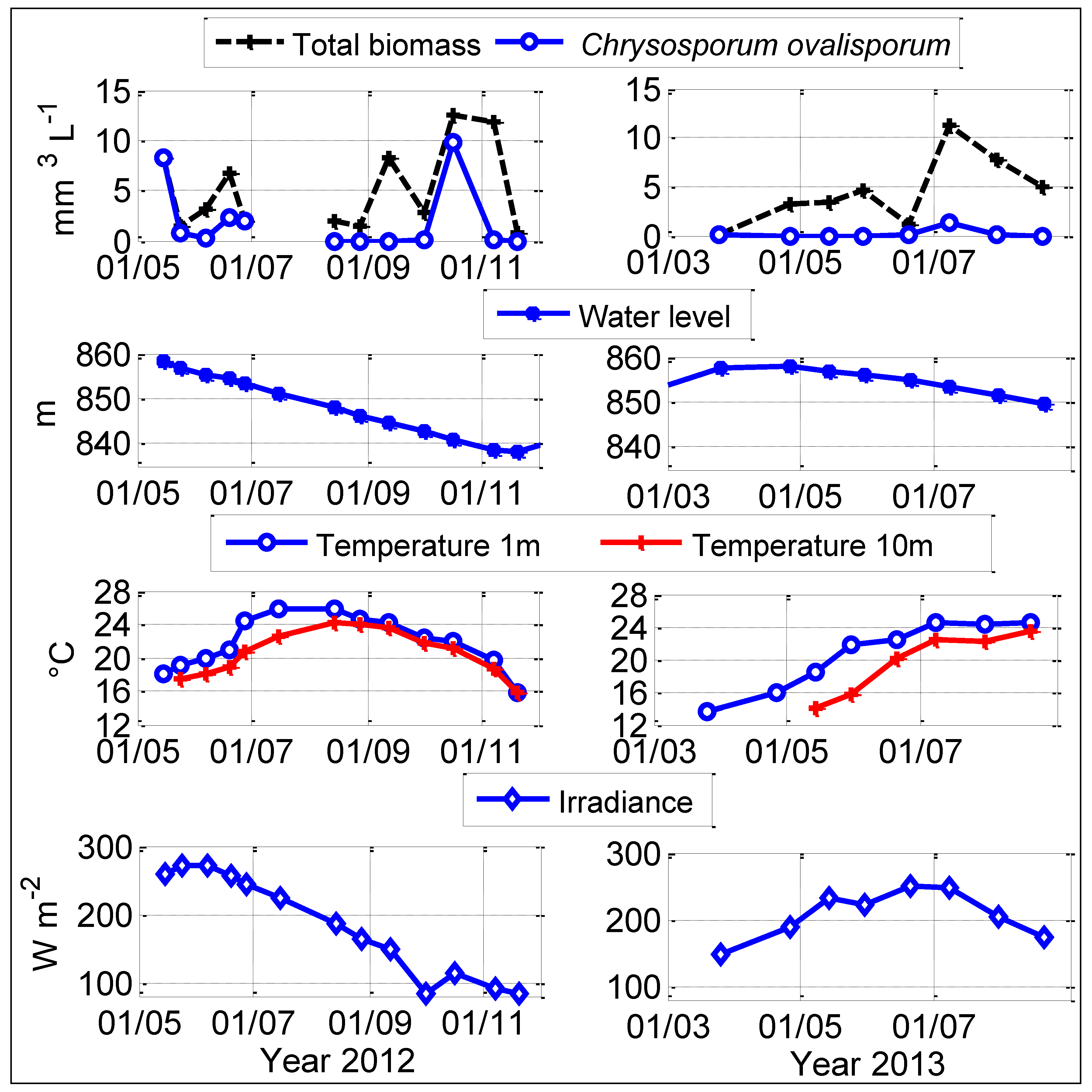
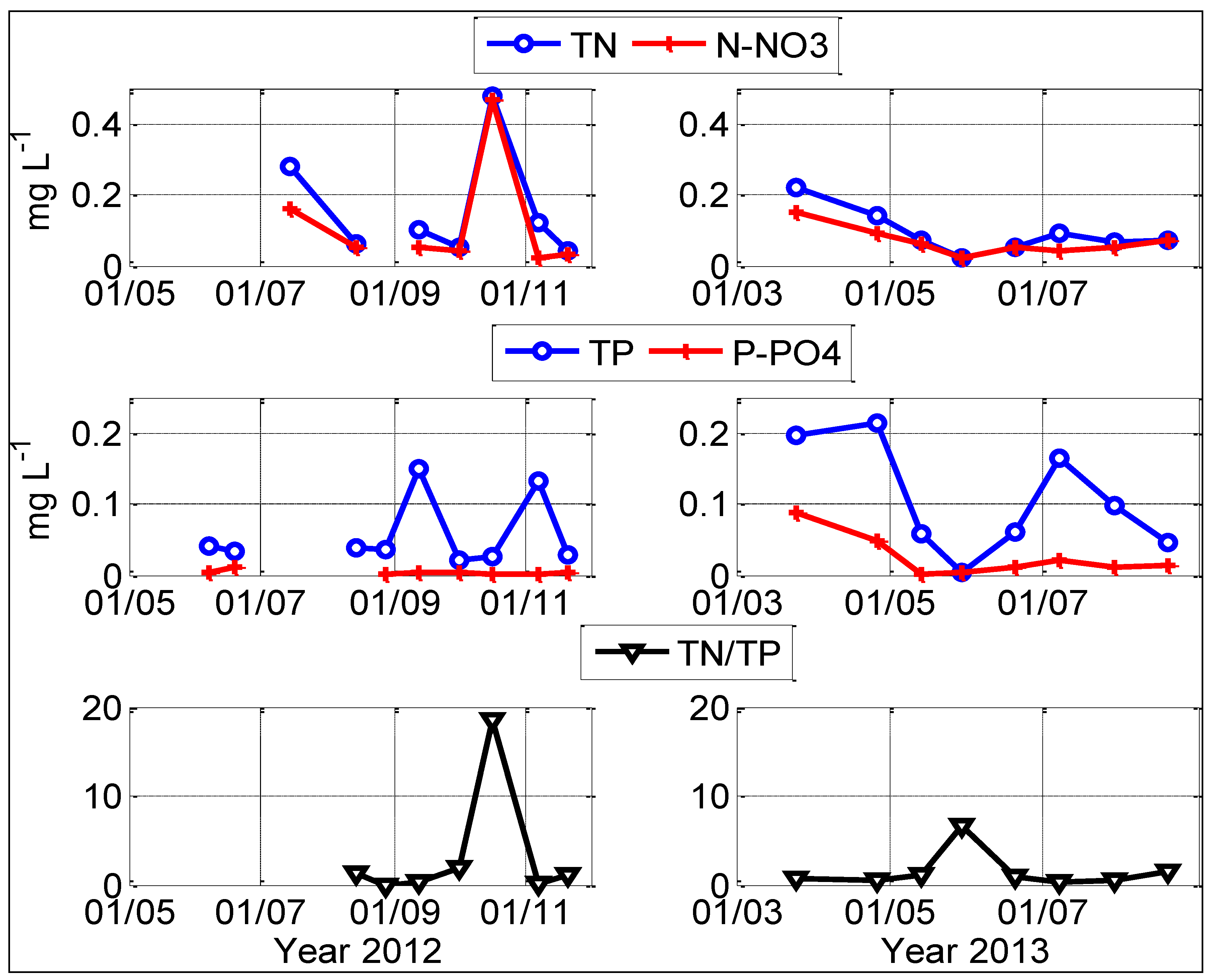
2.1. Physical-Chemical Conditions
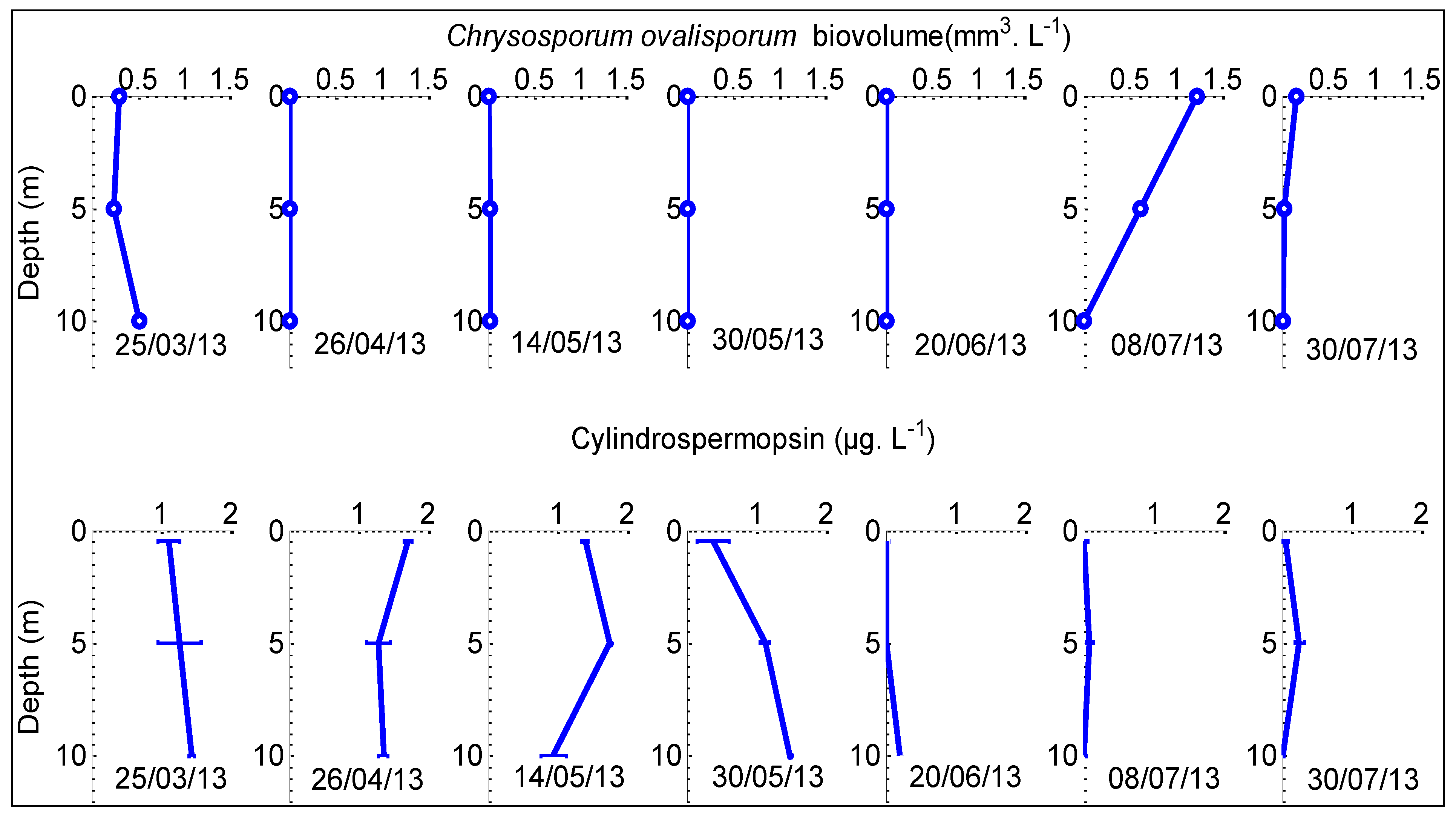
2.2. Dynamics of Chrysosporum ovalisporum in Karaoun Reservoir




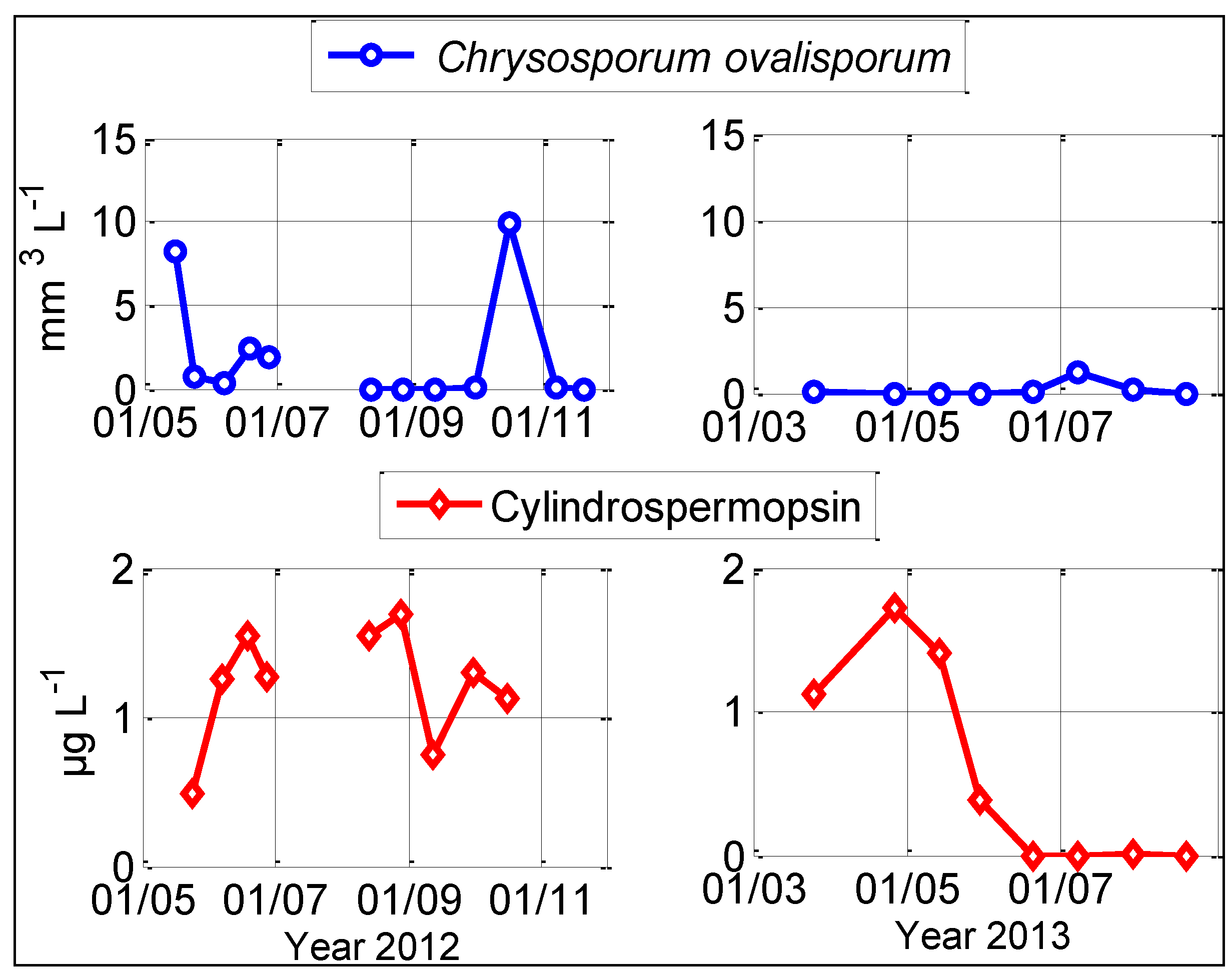
2.3. Cylindrospermopsin Quantification

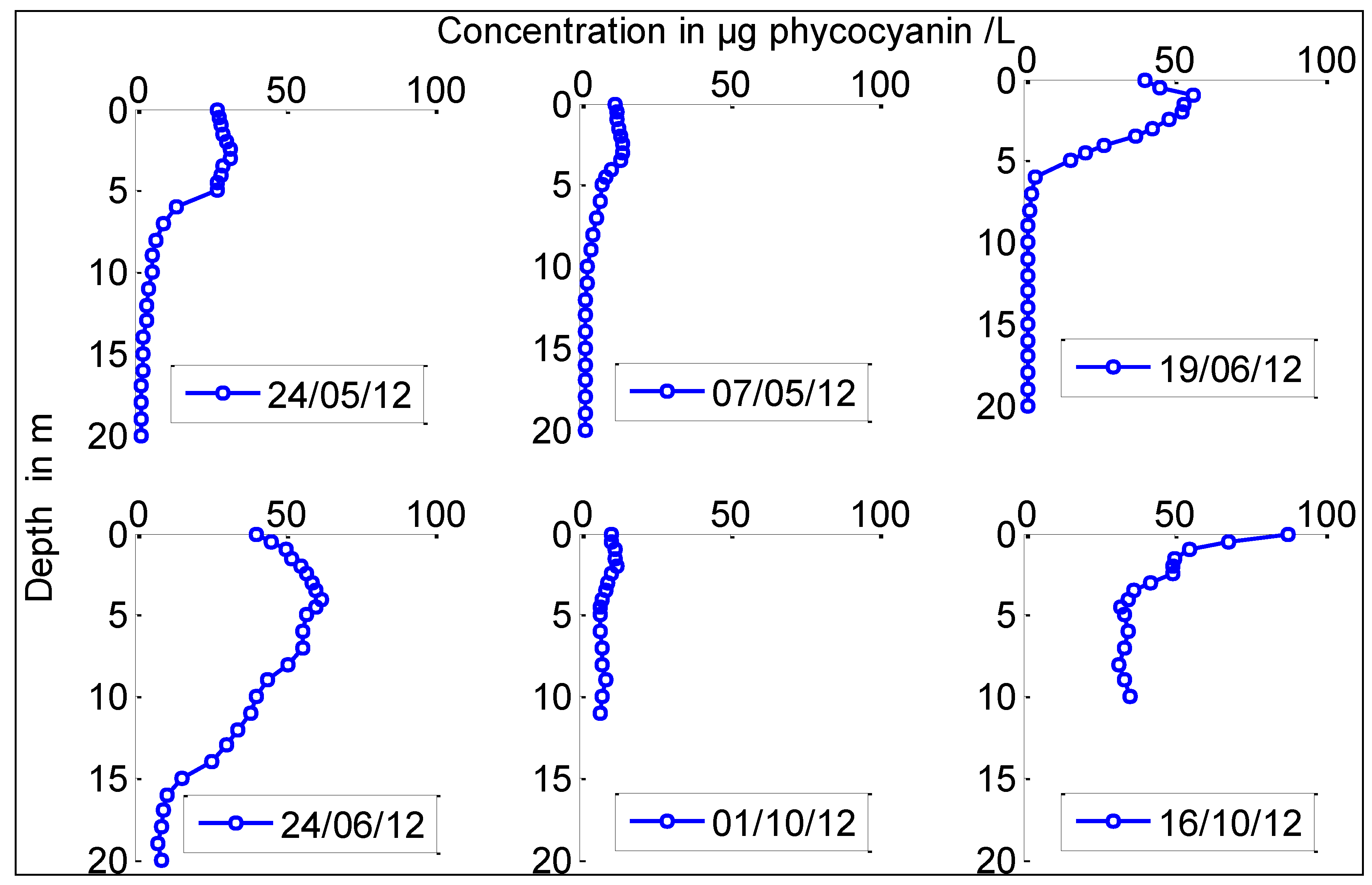
2.4. Comparison Between Chrysosporum ovalisporum and CYN Distribution in the Water Column

2.5. Absence of Correlation between Cylindrospermopsin Concentration and Chrysosporum ovalisporum Biovolumes
3. Discussion
3.1. Chrysosporum ovalisporum Blooms in Karaoun Reservoir
3.2. Relation between Cylindrospermopsin Concentrations and Chrysosporum ovalisporum
3.3. Disappearance of CYN from Water Column by Degradation or Sedimentation
4. Experimental Section
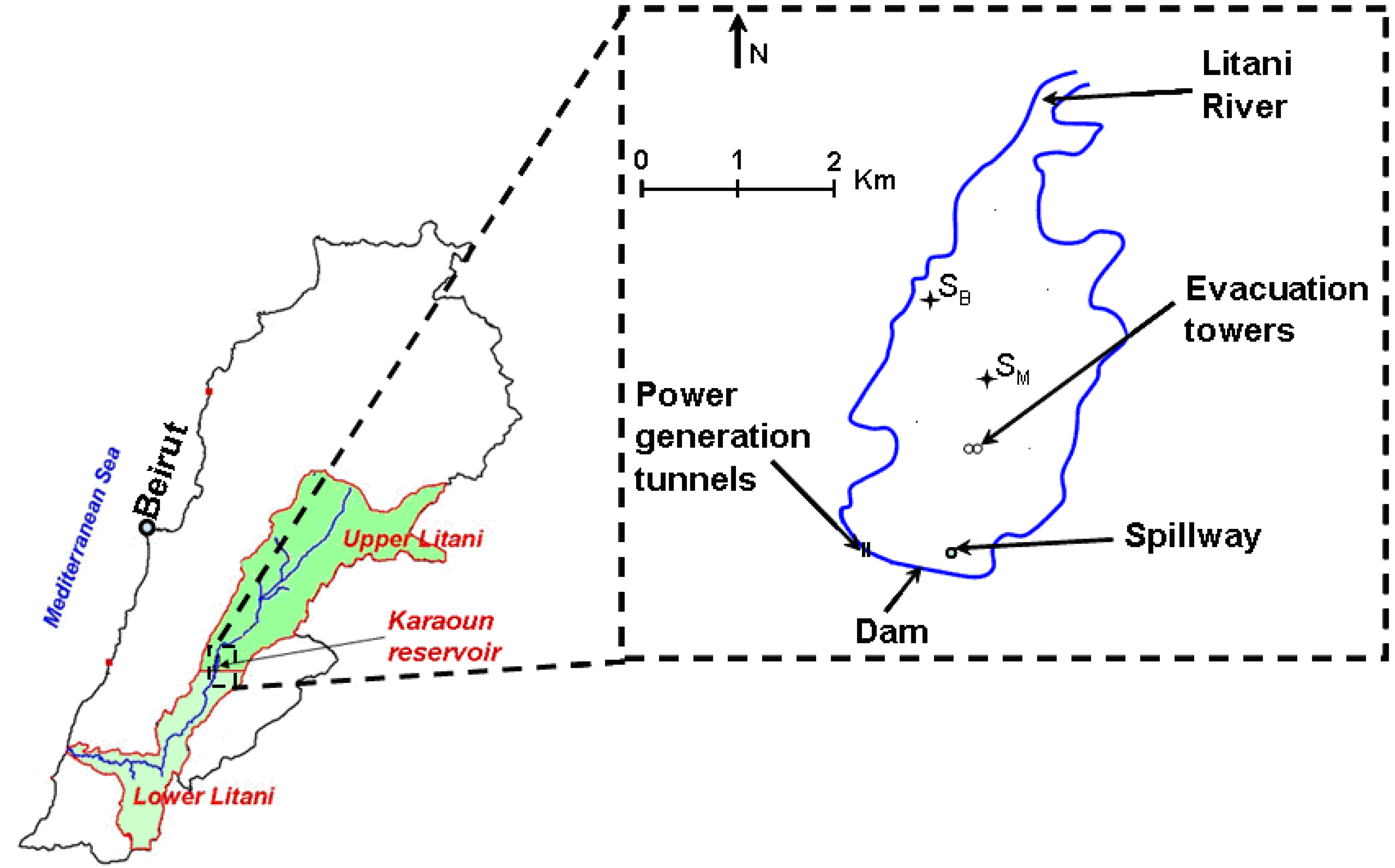
4.1. Study Site

| Morphometry and hydrology | Values |
|---|---|
| Surface area at full capacity | 12 km2 |
| Maximum storage capacity | 224 × 106 m3 |
| Maximum depth | 60 m |
| Mean depth at full capacity | 19 m |
| Altitude at maximum level (m) | 860 m |
| Catchment area | 1600 km2 |
| Mean residence time of water | 9 months |
4.2. Sampling Procedure
4.3. Water Temperature and Phycocyanin Measurements
4.4. Microscopic Identification and Counting
4.5. Nutrient Analysis
4.6. Cylindrospermopsin Analysis
4.7. Meteorological and Hydrological Data
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Li, D.; Kong, F.; Shi, X.; Ye, L.; Yu, Y.; Yang, Z. Quantification of microcystin-producing and non-microcystin producing Microcystis populations during the 2009 and 2010 blooms in Lake Taihu using quantitative real-time PCR. J. Environ. Sci. 2012, 24, 284–290. [Google Scholar] [CrossRef]
- Liu, Y.; Tan, W.; Wu, X.; Wu, Z.; Yu, G.; Li, R. First report of microcystin production in Microcystis smithii Komárek and Anagnostidis (Cyanobacteria) from a water bloom in Eastern China. J. Environ. Sci. 2012, 23, 102–107. [Google Scholar] [CrossRef]
- Messineo, V.; Bogialli, S.; Melchiorre, S.; Sechi, N.; Luglia, A.; Casiddu, P.; Mariani, M.A.; Padedda, B.M.; Corcia, A.D.; Mazza, R.; et al. Cyanobacterial toxins in Italian freshwaters. Limnologica 2009, 39, 95–106. [Google Scholar] [CrossRef]
- Ballot, A.; Pflugmacher, S.; Wiegand, C.; Kotut, K.; Krienitz, L. Cyanobacterial toxins in Lake Baringo, Kenya. Limnologica 2003, 33, 2–9. [Google Scholar] [CrossRef]
- Rejmánková, E.; Komárek, J.; Dix, M.; Rejmánková, J.; Girón, N. Cyanobacterial blooms in Lake Atitlan, Guatemala. Limnologica 2011, 41, 296–302. [Google Scholar] [CrossRef]
- Zapomělová, E.; Skácelová, O.; Pumann, P.; Kopp, R.; Janeček, E. Biogeographically interesting planktonic Nostocales (Cyanobacteria) in the Czech Republic and their polyphasic evaluation resulting in taxonomic revisions of Anabaena bergii Ostenfeld 1908 (Chrysosporum gen. nov.) and A. tenericaulis Nygaard 1949 (Dolichospermum tenericaule comb. nova). Hydrobiologia 2012, 698, 353–365. [Google Scholar] [CrossRef]
- Gkelis, S.; Moustaka-Gouni, M.; Sivonen, K.; Lanaras, T. First report of the cyanobacterium Aphanizomenon ovalisporum Forti in two Greek lakes and cyanotoxin occurrence. J. Plankton Res. 2005, 27, 1295–1300. [Google Scholar] [CrossRef]
- Messineo, V.; Melchiorre, S.; Di Corcia, A.; Gallo, P.; Bruno, M. Seasonal succession of Cylindrospermopsis raciborskii and Aphanizomenon ovalisporum blooms with cylindrospermopsin occurrence in the volcanic Lake Albano, Central Italy. Environ. Toxicol. 2010, 25, 18–27. [Google Scholar] [PubMed]
- Quesada, A.; Moreno, E.; Carrasco, D.; Paniagua, T.; Wormer, L.; Hoyos, C.D.; Sukenik, A. Toxicity of Aphanizomenon ovalisporum (Cyanobacteria) in a Spanish water reservoir. Eur. J. Phycol. 2006, 41, 39–45. [Google Scholar] [CrossRef]
- Pollingher, U.; Hadas, O.; Yacobi, Y.Z.; Zohary, T.; Berman, T. Aphanizomenon ovalisporum (Forti) in Lake Kinneret, Israel. J. Plankton Res. 1998, 20, 1321–1339. [Google Scholar] [CrossRef]
- Reynolds, C.S.; Oliver, R.L.; Walsby, A.E. Cyanobacterial dominance: The role of buoyancy regulation in dynamic lake environments. N. Z. J. Mar. Freshwater Res. 1987, 21, 379–390. [Google Scholar] [CrossRef]
- Reynolds, C.S. Nutrient uptake and assimilation in phytoplankton. In The Ecology of Phytoplankton; Cambridge University Press: Cambridge, UK, 2006; pp. 145–175. [Google Scholar]
- Kardinaal, W.; Visser, P. Dynamics of cyanobacterial toxins. In Harmful cyanobacteria; Huisman, J., Matthijs, H.C.P., Visser, P.M., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2005; pp. 41–63. [Google Scholar]
- Hadas, O.; Pinkas, R.; Malinsky-Rushansky, N.; Shalev-Alon, G.; Delphine, E.; Berner, T.; Sukenik, A.; Kaplan, A. Physiological variables determined under laboratory conditions may explain the bloom of Aphanizomenon ovalisporum in Lake Kinneret. Eur. J. Phycol. 2002, 37, 259–267. [Google Scholar] [CrossRef]
- Cirés, S.; Wörmer, L.; Timón, J.; Wiedner, C.; Quesada, A. Cylindrospermopsin production and release by the potentially invasive cyanobacterium Aphanizomenon ovalisporum under temperature and light gradients. Harmful Algae 2011, 10, 668–675. [Google Scholar] [CrossRef]
- Mehnert, G.; Leunert, F.; Cirés, S.; Jöhnk, K.D.; Rücker, J.; Nixdorf, B.; Wiedner, C. Competitiveness of invasive and native cyanobacteria from temperate freshwaters under various light and temperature conditions. J. Plankton Res. 2010, 32, 1009–1021. [Google Scholar] [CrossRef]
- Porat, R.; Teltsch, B.; Perelman, A.; Dubinsky, Z. Diel Buoyancy Changes by the Cyanobacterium Aphanizomenon ovalisporum from a Shallow Reservoir. J. Plankton Res. 2001, 23, 753–763. [Google Scholar] [CrossRef]
- Boopathi, T.; Ki, J.S. Impact of Environmental Factors on the Regulation of Cyanotoxin Production. Toxins 2014, 6, 1951–1978. [Google Scholar] [CrossRef] [PubMed]
- Stucken, K.; John, U.; Cembella, A.; Soto-Liebe, K.; Vásquez, M. Impact of nitrogen sources on gene expression and toxin production in the diazotroph Cylindrospermopsis raciborskii CS-505 and non-diazotroph Raphidiopsis brookii D9. Toxins 2014, 6, 1896–1915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths, D.J.; Saker, M.L. The Palm Island mystery disease 20 years on: A review of research on the cyanotoxin cylindrospermopsin. Environ. Toxicol. 2003, 18, 78–93. [Google Scholar] [CrossRef] [PubMed]
- Humpage, A.R.; Fenech, M.; Thomas, P.; Falconer, I.R. Micronucleus induction and chromosome loss in transformed human white cells indicate clastogenic and aneugenic action of the cyanobacterial toxin, cylindrospermopsin. Mutat. Res. Genet. Toxicol. Environ. Mutagenesis 2000, 472, 155–161. [Google Scholar] [CrossRef]
- Falconer, I.R.; Humpage, A.R. Cyanobacterial (blue-green algal) toxins in water supplies: Cylindrospermopsins. Environ. Toxicol. 2006, 21, 299–304. [Google Scholar] [PubMed]
- Preußel, K.; Wessel, G.; Fastner, J.; Chorus, I. Response of cylindrospermopsin production and release in Aphanizomenon flos-aquae (Cyanobacteria) to varying light and temperature conditions. Harmful Algae 2009, 8, 645–650. [Google Scholar] [CrossRef]
- Chiswell, R.K.; Shaw, G.R.; Eaglesham, G.; Smith, M.J.; Norris, R.L.; Seawright, A.A.; Moore, M.R. Stability of cylindrospermopsin, the toxin from the cyanobacterium, Cylindrospermopsis raciborskii: Effect of pH, temperature, and sunlight on decomposition. Environ. Toxicol. 1999, 14, 155–161. [Google Scholar] [CrossRef]
- Bogialli, S.; Bruno, M.; Curini, R.; Di Corcia, A.; Fanali, C.; Laganà, A. Monitoring algal toxins in lake water by liquid chromatography tandem mass spectrometry. Environ. Sci. Technol. 2006, 40, 2917–2923. [Google Scholar] [CrossRef] [PubMed]
- Rücker, J.; Stüken, A.; Nixdorf, B.; Fastner, J.; Chorus, I.; Wiedner, C. Concentrations of particulate and dissolved cylindrospermopsin in 21 Aphanizomenon-dominated temperate lakes. Toxicon 2007, 50, 800–809. [Google Scholar] [CrossRef] [PubMed]
- Slim, K. Contribution to the study of the flora of the basin of the Litani basin. Leb. Sci. Res. Rep. 1996, 1, 65–73. [Google Scholar]
- Saad, Z.; Slim, K.; Elzein, G.; Elsamad, O. Evaluation of the water quality of Karaoun reservoir (Lebanon). Bull. Soc. Neuchatel. Sci. Nat. 2005, 128, 71–80. [Google Scholar]
- Slim, K.; Fadel, A.; Atoui, A.; Lemaire, B.J.; Vinçon-Leite, B.; Tassin, B. Global warming as a driving factor for cyanobacterial blooms in Lake Karaoun, Lebanon. Desalination Water Treat. 2014, 52, 2094–2101. [Google Scholar] [CrossRef] [Green Version]
- Atoui, A.; Hafez, H.; Slim, K. Occurrence of toxic cyanobacterial blooms for the first time in Lake Karaoun, Lebanon. Water Environ. J. 2013, 27, 42–49. [Google Scholar] [CrossRef]
- Pollingher, U. Phytoplankton periodicity in a subtropical lake (Lake Kinneret, Israel). Hydrobiologia 1986, 138, 127–138. [Google Scholar] [CrossRef]
- Chen, Y.; Qin, B.; Teubner, K.; Dokulil, M.T. Long-term dynamics of phytoplankton assemblages: Microcystis-domination in Lake Taihu, a large shallow lake in China. J. Plankton Res. 2003, 25, 445–453. [Google Scholar] [CrossRef]
- Moreno-Ostos, E.; Cruz-Pizarro, L.; Basanta-Alvés, A.; Escot, C.; George, D.G. Algae in the motion: Spatial distribution of phytoplankton in thermally stratified reservoirs. Limnetica. 2006, 25, 205–216. [Google Scholar]
- Whitton, B.; Potts, M.; Oliver, R.; Ganf, G. Freshwater Blooms. In The Ecology of Cyanobacteria; Springer Netherlands: Dordrecht, The Netherlands, 2002; pp. 149–194. [Google Scholar]
- Smith, V.H.; Bierman, V.J.; Jones, B.L.; Havens, K.E. Historical trends in the Lake Okeechobee ecosystem IV. Nitrogen:phosphorus ratios, cyanobacterial dominance, and nitrogen fixation potential. Arch. Hydrobiol. 1995, 107, 71–88. [Google Scholar]
- Gophen, M.; Smith, V.H.; Nishri, A.; Threlkeld, S.T. Nitrogen deficiency, phosphorus sufficiency, and the invasion of Lake Kinneret, Israel, by the N2-fixing cyanobacterium Aphanizomenon ovalisporum. Aquat. Sci. 1999, 61, 293–306. [Google Scholar]
- Hadas, O.; Pinkas, R.; Malinsky-Rushansky, N.; Nishri, A.; Kaplan, A.; Rimmer, A.; Sukenik, A. Appearance and establishment of diazotrophic cyanobacteria in Lake Kinneret, Israel. Freshw. Biol. 2012, 57, 1214–1227. [Google Scholar] [CrossRef]
- Vinçon-Leite, B.; Lemaire, B.J.; Khac, V.; Tassin, B. Long-term temperature evolution in a deep sub-alpine lake, Lake Bourget, France: How a one-dimensional model improves its trend assessment. Hydrobiologia 2014, 1–16. [Google Scholar]
- Wu, Q.L.; Hahn, M.W. High predictability of the seasonal dynamics of a species-like Polynucleobacter population in a freshwater lake. Environ. Microbiol. 2006, 8, 1660–1666. [Google Scholar] [CrossRef] [PubMed]
- Peeters, F.; Livingstone, D.M.; Goudsmit, G.H.; Kipfer, R.; Forster, R. Modeling 50 years of historical temperature profiles in a large central European lake. Limnol. Oceanogr. 2002, 47, 186–197. [Google Scholar] [CrossRef]
- Everson, S.; Fabbro, L.; Kinnear, S.; Eaglesham, G.; Wright, P. Distribution of the cyanobacterial toxins cylindrospermopsin and deoxycylindrospermopsin in a stratified lake in north-eastern New South Wales, Australia. Mar. Freshw. Res. 2009, 60, 25–33. [Google Scholar] [CrossRef]
- Shaw, G.R.; Sukenik, A.; Livne, A.; Chiswell, R.K.; Smith, M.J.; Seawright, A.A.; Norris, R.L.; Eaglesham, G.K.; Moore, M.R. Blooms of the cylindrospermopsin containing cyanobacterium, Aphanizomenon ovalisporum (Forti), in newly constructed lakes, Queensland, Australia. Environ. Toxicol. 1999, 14, 167–177. [Google Scholar] [CrossRef]
- Ho, L.; Sawade, E.; Newcombe, G. Biological treatment options for cyanobacteria metabolite removal: A review. Water Res. 2012, 46, 1536–1548. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.J.; Shaw, G.R.; Eaglesham, G.K.; Ho, L.; Brookes, J.D. Elucidating the factors influencing the biodegradation of cylindrospermopsin in drinking water sources. Environ. Toxicol. 2008, 23, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Wörmer, L.; Cirés, S.; Carrasco, D.; Quesada, A. Cylindrospermopsin is not degraded by co-occurring natural bacterial communities during a 40-day study. Harmful Algae 2008, 7, 206–213. [Google Scholar] [CrossRef]
- Cyanobacteria and Cyanotoxins: Information for Drinking Water Systems. Available online: http://water.epa.gov/scitech/swguidance/standards/criteria/nutrients/upload/cyanobacteria_factsheet.pdf (accessed on 23 October 2014).
- Fadel, A.; Lemaire, B.J.; Atoui, A.; Vinçon-Leite, B.; Amacha, N.; Slim, K.; Tassin, B. First assessment of the ecological status of Karaoun Reservoir, Lebanon. Lakes Res. 2014, 19, 142–157. [Google Scholar] [CrossRef]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota, part 1: Chroococcales. In Süßwasserflora von Mitteleuropa; Elsevier: Heidelberg, Germany, 1998. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota, part 2: Oscillatoriales. In Süsswasserflora von Mitteleuropa; Elsevier: Heidelberg, Germany, 2005. [Google Scholar]
- Humpage, A.R.; Froscio, S.M.; Lau, H.M.; Murphy, D.; Blackbeard, J. Evaluation of the Abraxis Strip Test for Microcystins™ for use with wastewater effluent and reservoir water. Water Res. 2012, 46, 1556–1565. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fadel, A.; Atoui, A.; Lemaire, B.J.; Vinçon-Leite, B.; Slim, K. Dynamics of the Toxin Cylindrospermopsin and the Cyanobacterium Chrysosporum (Aphanizomenon) ovalisporum in a Mediterranean Eutrophic Reservoir. Toxins 2014, 6, 3041-3057. https://doi.org/10.3390/toxins6113041
Fadel A, Atoui A, Lemaire BJ, Vinçon-Leite B, Slim K. Dynamics of the Toxin Cylindrospermopsin and the Cyanobacterium Chrysosporum (Aphanizomenon) ovalisporum in a Mediterranean Eutrophic Reservoir. Toxins. 2014; 6(11):3041-3057. https://doi.org/10.3390/toxins6113041
Chicago/Turabian StyleFadel, Ali, Ali Atoui, Bruno J. Lemaire, Brigitte Vinçon-Leite, and Kamal Slim. 2014. "Dynamics of the Toxin Cylindrospermopsin and the Cyanobacterium Chrysosporum (Aphanizomenon) ovalisporum in a Mediterranean Eutrophic Reservoir" Toxins 6, no. 11: 3041-3057. https://doi.org/10.3390/toxins6113041
APA StyleFadel, A., Atoui, A., Lemaire, B. J., Vinçon-Leite, B., & Slim, K. (2014). Dynamics of the Toxin Cylindrospermopsin and the Cyanobacterium Chrysosporum (Aphanizomenon) ovalisporum in a Mediterranean Eutrophic Reservoir. Toxins, 6(11), 3041-3057. https://doi.org/10.3390/toxins6113041