Characterization of a Novel BmαTX47 Toxin Modulating Sodium Channels: The Crucial Role of Expression Vectors in Toxin Pharmacological Activity

Abstract
:1. Introduction
2. Results and Discussion
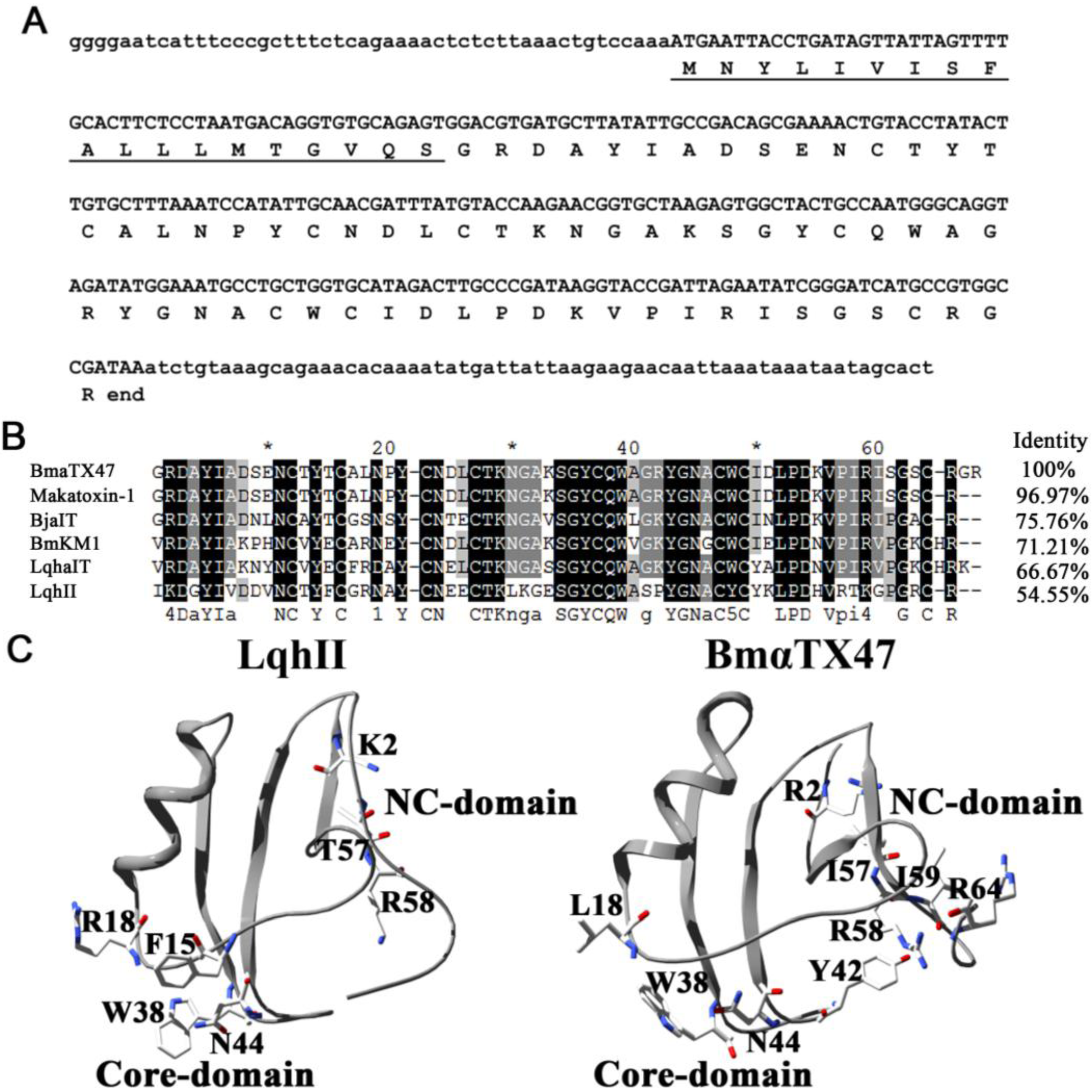
2.1. Cloning and Sequence Analysis of BmαTX47

2.2. Expression and Purification of BmαTX47 Using Expression Vector pET-14b

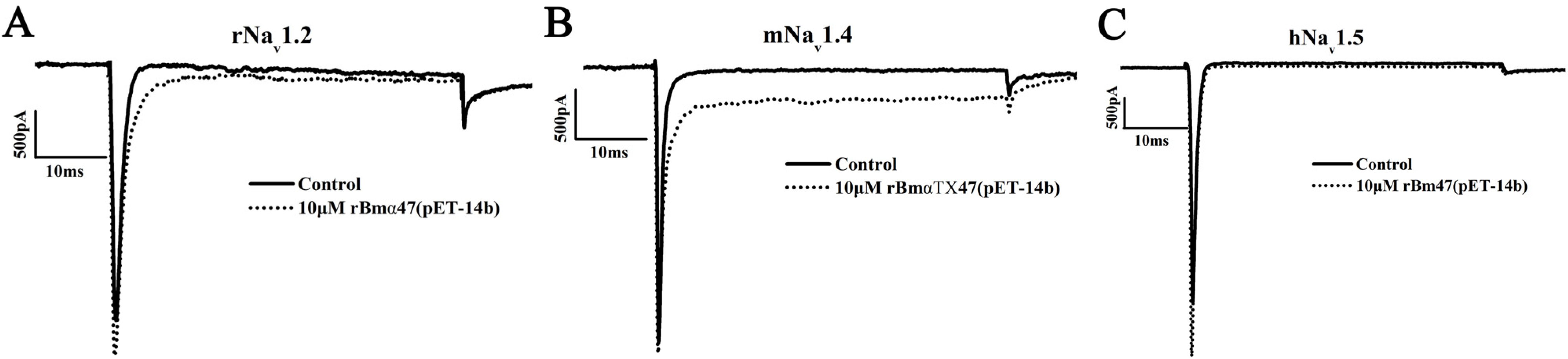
2.3. Pharmacological Characterization of rBmαTX47 from Expression Vector pET-14b


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxins (10 μM) | I5ms/Ipeak on rNav1.2 (%) | I5ms/Ipeak on mNav1.4 (%) | I5ms/Ipeak on hNav1.5 (%) |
|---|---|---|---|
| rBmαTX47 (pET-14b) | 11.33 ± 1.46 | 15.96 ± 1.87 | 5.04 ± 2.38 |
| rBmαTX47 (pET-28a) | 44.12 ± 3.17 | 25.40 ± 4.89 | 65.34 ± 3.86 |
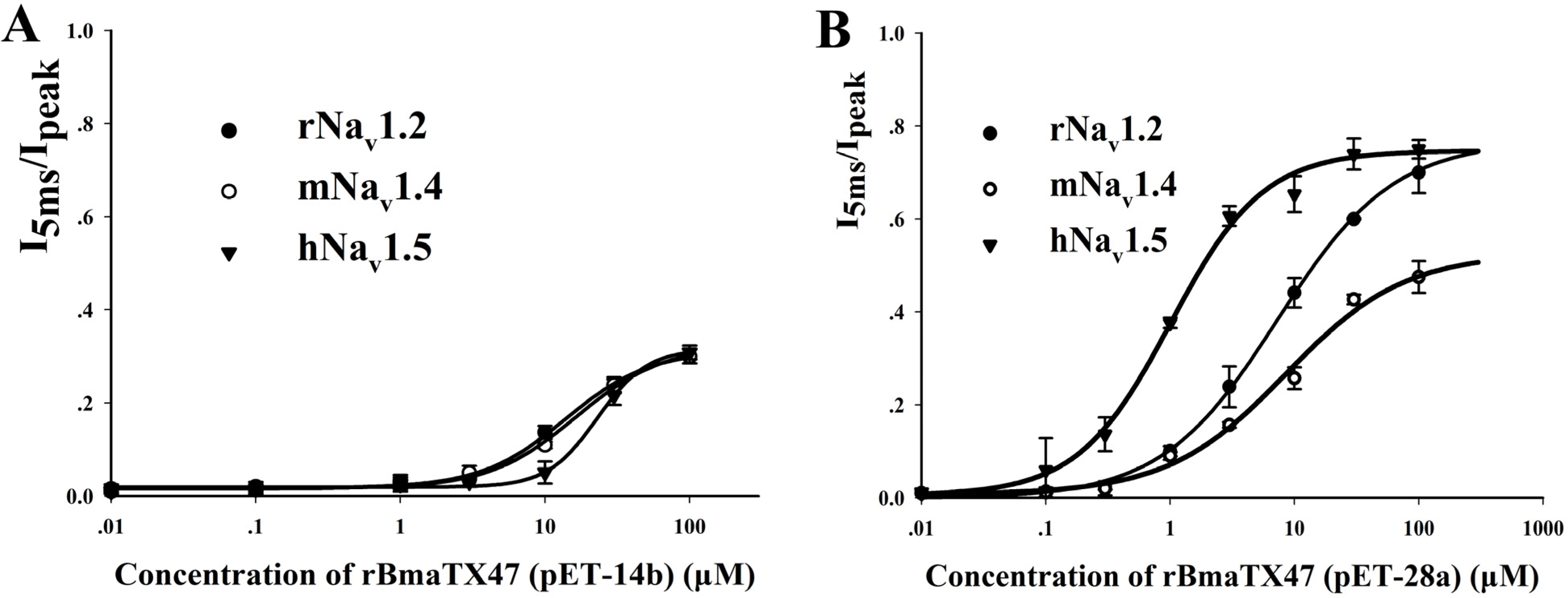
2.4. Preparation and Pharmacological Features of rBmαTX47 Using Expression Vector pET-28a

2.5. The Pivotal Role of Expression Vectors in Scorpion Toxin Pharmacological Properties

3. Experimental Section
3.1. cDNA Library Construction and Screening
3.2. Materials
3.3. Construction of Expression Vectors
3.4. Expression and Purification of BmαTX47
3.5. Electrophysiology

4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Catterall, W.A. Voltage-gated sodium channels at 60: Structure, function and pathophysiology. J. Physiol. 2012, 590, 2577–2589. [Google Scholar] [CrossRef]
- Catterall, W.A.; Goldin, A.L.; Waxman, S.G. International Union of Pharmacology. XLVII. Nomenclature and structure-function relationships of voltage-gated sodium channels. Pharmacol. Rev. 2005, 57, 397–409. [Google Scholar] [CrossRef]
- Beneski, D.A.; Catterall, W.A. Covalent labeling of protein components of the sodium channel with a photoactivable derivative of scorpion toxin. Proc. Natl. Acad. Sci. USA 1980, 77, 639–643. [Google Scholar] [CrossRef]
- Yang, N.; Horn, R. Evidence for voltage-dependent S4 movement in sodium channels. Neuron 1995, 15, 213–218. [Google Scholar] [CrossRef]
- Yang, N.; George, A.L., Jr.; Horn, R. Molecular basis of charge movement in voltage-gated sodium channels. Neuron 1996, 16, 113–122. [Google Scholar] [CrossRef]
- Chanda, B.; Bezanilla, F. Tracking voltage-dependent conformational changes in skeletal muscle sodium channel during activation. J. Gen. Physiol. 2002, 120, 629–645. [Google Scholar] [CrossRef]
- Catterall, W.A.; Cestele, S.; Yarov-Yarovoy, V.; Yu, F.H.; Konoki, K.; Scheuer, T. Voltage-gated ion channels and gating modifier toxins. Toxicon 2007, 49, 124–141. [Google Scholar] [CrossRef] [Green Version]
- Bosmans, F.; Tytgat, J. Voltage-gated sodium channel modulation by scorpion alpha-toxins. Toxicon 2007, 49, 142–158. [Google Scholar] [CrossRef]
- Chen, H.; Gordon, D.; Heinemann, S.H. Modulation of cloned skeletal muscle sodium channels by the scorpion toxins Lqh II, Lqh III, and LqhalphaIT. Pflugers Arch. 2000, 439, 423–432. [Google Scholar] [CrossRef]
- Leipold, E.; Lu, S.; Gordon, D.; Hansel, A.; Heinemann, S.H. Combinatorial interaction of scorpion toxins Lqh-2, Lqh-3, and LqhalphaIT with sodium channel receptor sites-3. Mol. Pharmacol. 2004, 65, 685–691. [Google Scholar] [CrossRef]
- Gilles, N.; Blanchet, C.; Shichor, I.; Zaninetti, M.; Lotan, I.; Bertrand, D.; Gordon, D. A scorpion alpha-like toxin that is active on insects and mammals reveals an unexpected specificity and distribution of sodium channel subtypes in rat brain neurons. J. Neurosci. 1999, 19, 8730–8739. [Google Scholar]
- Krimm, I.; Gilles, N.; Sautiere, P.; Stankiewicz, M.; Pelhate, M.; Gordon, D.; Lancelin, J.M. NMR structures and activity of a novel alpha-like toxin from the scorpion Leiurus quinquestriatus hebraeus. J. Mol. Biol. 1999, 285, 1749–1763. [Google Scholar] [CrossRef]
- Zilberberg, N.; Froy, O.; Loret, E.; Cestele, S.; Arad, D.; Gordon, D.; Gurevitz, M. Identification of structural elements of a scorpion alpha-neurotoxin important for receptor site recognition. J. Biol. Chem. 1997, 272, 14810–14816. [Google Scholar] [CrossRef]
- Chen, H.; Heinemann, S.H. Interaction of scorpion alpha-toxins with cardiac sodium channels: Binding properties and enhancement of slow inactivation. J. Gen. Physiol. 2001, 117, 505–518. [Google Scholar] [CrossRef]
- Kahn, R.; Karbat, I.; Ilan, N.; Cohen, L.; Sokolov, S.; Catterall, W.A.; Gordon, D.; Gurevitz, M. Molecular requirements for recognition of brain voltage-gated sodium channels by scorpion alpha-toxins. J. Biol. Chem. 2009, 284, 20684–20691. [Google Scholar] [CrossRef]
- Gur, M.; Kahn, R.; Karbat, I.; Regev, N.; Wang, J.; Catterall, W.A.; Gordon, D.; Gurevitz, M. Elucidation of the molecular basis of selective recognition uncovers the interaction site for the core domain of scorpion alpha-toxins on sodium channels. J. Biol. Chem. 2011, 286, 35209–35217. [Google Scholar]
- Wang, J.; Yarov-Yarovoy, V.; Kahn, R.; Gordon, D.; Gurevitz, M.; Scheuer, T.; Catterall, W.A. Mapping the receptor site for alpha-scorpion toxins on a Na+ channel voltage sensor. Proc. Natl. Acad. Sci. USA 2011, 108, 15426–15431. [Google Scholar]
- Sun, Y.M.; Bosmans, F.; Zhu, R.H.; Goudet, C.; Xiong, Y.M.; Tytgat, J.; Wang, D.C. Importance of the conserved aromatic residues in the scorpion alpha-like toxin BmK M1: The hydrophobic surface region revisited. J. Biol. Chem. 2003, 278, 24125–24131. [Google Scholar]
- Wang, C.G.; Gilles, N.; Hamon, A.; Le Gall, F.; Stankiewicz, M.; Pelhate, M.; Xiong, Y.M.; Wang, D.C.; Chi, C.W. Exploration of the functional site of a scorpion alpha-like toxin by site-directed mutagenesis. Biochemistry 2003, 42, 4699–4708. [Google Scholar]
- Dai, H.; Yin, S.; Li, T.; Cao, Z.; Ji, Y.; Wu, Y.; Li, W. Recombinant expression, purification, and characterization of scorpion toxin BmalphaTX14. Protein Expr. Purif. 2012, 82, 325–331. [Google Scholar] [CrossRef]
- Han, S.; Yi, H.; Yin, S.J.; Chen, Z.Y.; Liu, H.; Cao, Z.J.; Wu, Y.L.; Li, W.X. Structural basis of a potent peptide inhibitor designed for Kv1.3 channel, a therapeutic target of autoimmune disease. J. Biol. Chem. 2008, 283, 19058–19065. [Google Scholar]
- Chen, Z.; Hu, Y.; Han, S.; Yin, S.; He, Y.; Wu, Y.; Cao, Z.; Li, W. ImKTx1, a new Kv1.3 channel blocker with a unique primary structure. J. Biochem. Mol. Toxicol. 2011, 25, 244–251. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Zeng, D.Y.; Hu, Y.T.; He, Y.W.; Pan, N.; Ding, J.P.; Cao, Z.J.; Liu, M.L.; Li, W.X.; Yi, H.; et al. Structural and functional diversity of acidic scorpion potassium channel toxins. PLoS One 2012, 7, e35154. [Google Scholar] [CrossRef]
- Karbat, I.; Kahn, R.; Cohen, L.; Ilan, N.; Gilles, N.; Corzo, G.; Froy, O.; Gur, M.; Albrecht, G.; Heinemann, S.H.; et al. The unique pharmacology of the scorpion alpha-like toxin Lqh3 is associated with its flexible C-tail. FEBS J. 2007, 274, 1918–1931. [Google Scholar] [CrossRef]
- Li, S.; Ma, Y.; Jang, S.; Wu, Y.; Liu, H.; Cao, Z.; Li, W. A HindIII BAC library construction of MesobuthusmartensiiKarsch (Scorpiones:Buthidae): An important genetic resource for comparative genomics and phylogenetic analysis. Genes Genet. Syst. 2009, 84, 417–424. [Google Scholar] [CrossRef]
- Ma, Y.; Zhao, R.; He, Y.; Li, S.; Liu, J.; Wu, Y.; Cao, Z.; Li, W. Transcriptome analysis of the venom gland of the scorpion Scorpiopsjendeki: Implication for the evolution of the scorpion venom arsenal. BMC Genomics 2009, 10, 290. [Google Scholar] [CrossRef]
- He, Y.; Zhao, R.; Di, Z.; Li, Z.; Xu, X.; Hong, W.; Wu, Y.; Zhao, H.; Li, W.; Cao, Z. Molecular diversity of Chaerilidae venom peptides reveals the dynamic evolution of scorpion venom components from Buthidae to non-Buthidae. J. Proteomics 2013, 89, 1–14. [Google Scholar] [CrossRef]
- Gong, J.; Kini, R.M.; Gwee, M.C.; Gopalakrishnakone, P.; Chung, M.C. Makatoxin I, a novel toxin isolated from the venom of the scorpion ButhusmartensiKarsch, exhibits nitrergic actions. J. Biol. Chem. 1997, 272, 8320–8324. [Google Scholar]
- Chen, H.; Lu, S.; Leipold, E.; Gordon, D.; Hansel, A.; Heinemann, S.H. Differential sensitivity of sodium channels from the central and peripheral nervous system to the scorpion toxins Lqh-2 and Lqh-3. Eur. J. Neurosci. 2002, 16, 767–770. [Google Scholar] [CrossRef]
- Liu, L.H.; Bosmans, F.; Maertens, C.; Zhu, R.H.; Wang, D.C.; Tytgat, J. Molecular basis of the mammalian potency of the scorpion alpha-like toxin, BmK M1. FASEB J. 2005, 19, 594–596. [Google Scholar]
- Ye, X.; Bosmans, F.; Li, C.; Zhang, Y.; Wang, D.C.; Tytgat, J. Structural basis for the voltage-gated Na+ channel selectivity of the scorpion alpha-like toxin BmK M1. J. Mol. Biol. 2005, 353, 788–803. [Google Scholar] [CrossRef]
- Weinberger, H.; Moran, Y.; Gordon, D.; Turkov, M.; Kahn, R.; Gurevitz, M. Positions under positive selection--key for selectivity and potency of scorpion alpha-toxins. Mol. Biol. Evol. 2010, 27, 1025–1034. [Google Scholar] [CrossRef]
- Chen, R.; Chung, S.H. Binding modes and functional surface of anti-mammalian scorpion alpha-toxins to sodium channels. Biochemistry 2012, 51, 7775–7782. [Google Scholar] [CrossRef]
- Durek, T.; Vetter, I.; Wang, C.I.; Motin, L.; Knapp, O.; Adams, D.J.; Lewis, R.J.; Alewood, P.F. Chemical engineering and structural and pharmacological characterization of the alpha-scorpion toxin OD1. ACS Chem. Biol. 2013, 8, 1215–1222. [Google Scholar] [CrossRef]
- Wang, K.; Yin, S.J.; Lu, M.; Yi, H.; Dai, C.; Xu, X.J.; Cao, Z.J.; Wu, Y.L.; Li, W.X. Functional analysis of the alpha-neurotoxin, BmalphaTX14, derived from the Chinese scorpion, Buthus martensii Kars. Biotechnol. Lett. 2006, 28, 1767–1772. [Google Scholar] [CrossRef]
- Zhu, S.; Peigneur, S.; Gao, B.; Lu, X.; Cao, C.; Tytgat, J. Evolutionary diversification of Mesobuthus alpha-scorpion toxins affecting sodium channels. Mol. Cell. Proteomics 2012, 11. [Google Scholar] [CrossRef]
- Zimmer, T.; Bollensdorff, C.; Haufe, V.; Birch-Hirschfeld, E.; Benndorf, K. Mouse heart Na+ channels: Primary structure and function of two isoforms and alternatively spliced variants. Am. J. Physiol.-Heart Circ. Physiol. 2002, 282, H1007–H1017. [Google Scholar]
- Yu, J.B.; Ji, P.; Zha, X.M.; Shen, W.D.; Wu, X.F. Cloning and expression of ButhusmartensiiKarsch scorpion toxin gene (BmK IT3) in Escherichia coli. Sheng Wu Gong Cheng Xue Bao 2002, 18, 106–108. [Google Scholar]
- Zhu, L.; Peigneur, S.; Gao, B.; Tytgat, J.; Zhu, S. Two recombinant alpha-like scorpion toxins from Mesobuthuseupeus with differential affinity toward insect and mammalian Na+ channels. Biochimie 2013, 95, 1732–1740. [Google Scholar] [CrossRef]
- Zilberberg, N.; Gordon, D.; Pelhate, M.; Adams, M.E.; Norris, T.M.; Zlotkin, E.; Gurevitz, M. Functional expression and genetic alteration of an alpha scorpion neurotoxin. Biochemistry 1996, 35, 10215–10222. [Google Scholar] [CrossRef]
- Karbat, I.; Frolow, F.; Froy, O.; Gilles, N.; Cohen, L.; Turkov, M.; Gordon, D.; Gurevitz, M. Molecular basis of the high insecticidal potency of scorpion alpha-toxins. J. Biol. Chem. 2004, 279, 31679–31686. [Google Scholar]
- Karbat, I.; Turkov, M.; Cohen, L.; Kahn, R.; Gordon, D.; Gurevitz, M.; Frolow, F. X-ray structure and mutagenesis of the scorpion depressant toxin LqhIT2 reveals key determinants crucial for activity and anti-insect selectivity. J. Mol. Biol. 2007, 366, 586–601. [Google Scholar] [CrossRef]
- Sun, Y.M.; Liu, W.; Zhu, R.H.; Goudet, C.; Tytgat, J.; Wang, D.C. Roles of disulfide bridges in scorpion toxin BmK M1 analyzed by mutagenesis. J. Pept. Res. 2002, 60, 247–256. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Hu, Y.T.; Yang, W.S.; He, Y.W.; Feng, J.; Wang, B.; Zhao, R.M.; Ding, J.P.; Cao, Z.J.; Li, W.X.; et al. Hg1, novel peptide inhibitor specific for Kv1.3 channels from first scorpion Kunitz-type potassium channel toxin family. J. Biol. Chem. 2012, 287, 13813–13821. [Google Scholar] [CrossRef]
- Zhao, R.; Dai, H.; Qiu, S.; Li, T.; He, Y.; Ma, Y.; Chen, Z.; Wu, Y.; Li, W.; Cao, Z. SdPI, the first functionally characterized Kunitz-type trypsin inhibitor from scorpion venom. PLoS One 2011, 6, e27548. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, T.; Xu, L.; Liu, H.; He, Y.; Liang, S.; Li, W.; Wu, Y. Characterization of a Novel BmαTX47 Toxin Modulating Sodium Channels: The Crucial Role of Expression Vectors in Toxin Pharmacological Activity. Toxins 2014, 6, 816-829. https://doi.org/10.3390/toxins6030816
Li T, Xu L, Liu H, He Y, Liang S, Li W, Wu Y. Characterization of a Novel BmαTX47 Toxin Modulating Sodium Channels: The Crucial Role of Expression Vectors in Toxin Pharmacological Activity. Toxins. 2014; 6(3):816-829. https://doi.org/10.3390/toxins6030816
Chicago/Turabian StyleLi, Tian, Lingna Xu, Honglian Liu, Yawen He, Songping Liang, Wenxin Li, and Yingliang Wu. 2014. "Characterization of a Novel BmαTX47 Toxin Modulating Sodium Channels: The Crucial Role of Expression Vectors in Toxin Pharmacological Activity" Toxins 6, no. 3: 816-829. https://doi.org/10.3390/toxins6030816
APA StyleLi, T., Xu, L., Liu, H., He, Y., Liang, S., Li, W., & Wu, Y. (2014). Characterization of a Novel BmαTX47 Toxin Modulating Sodium Channels: The Crucial Role of Expression Vectors in Toxin Pharmacological Activity. Toxins, 6(3), 816-829. https://doi.org/10.3390/toxins6030816