
Toxicity of the Anti-ribosomal Lectin Ebulin f in Lungs and Intestines in Elderly Mice

 ,
, 
Abstract
:1. Introduction
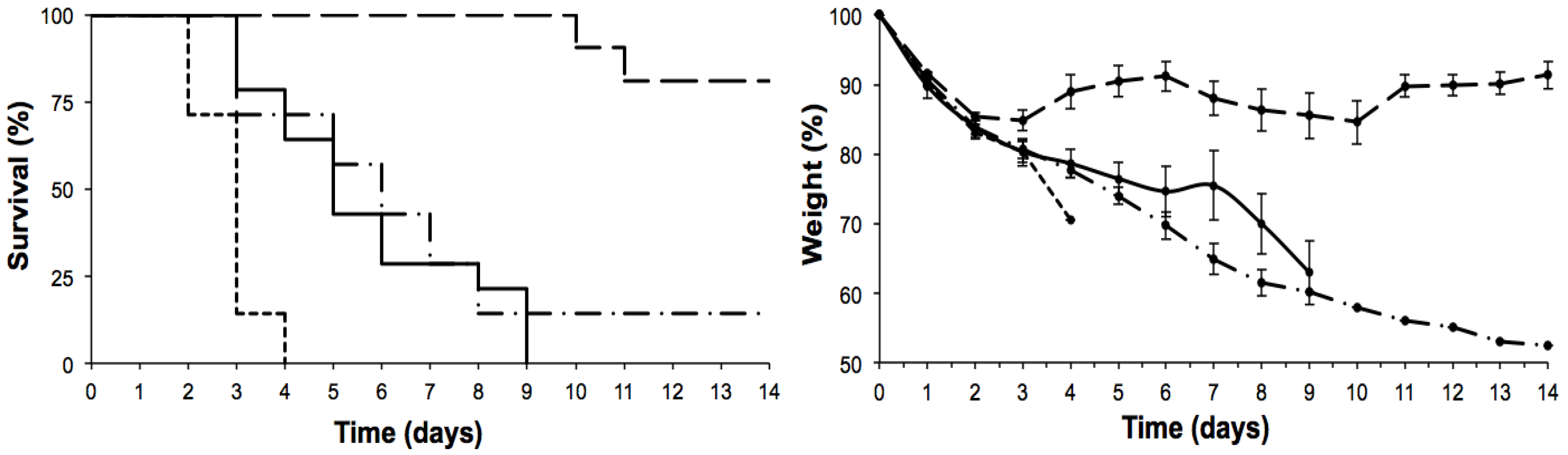
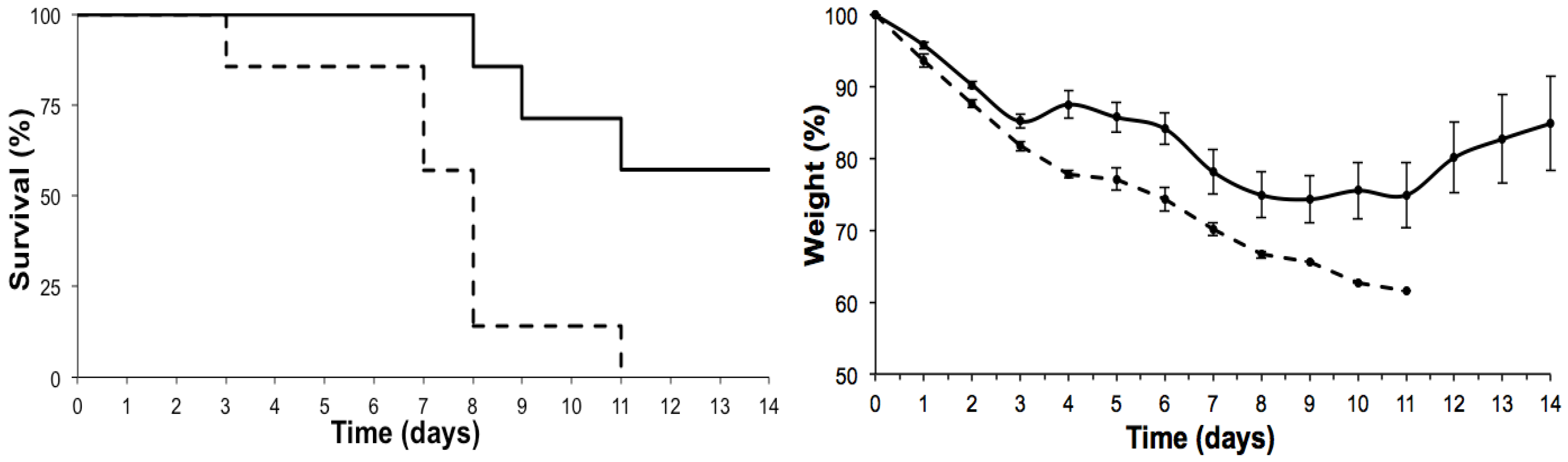
2. Results



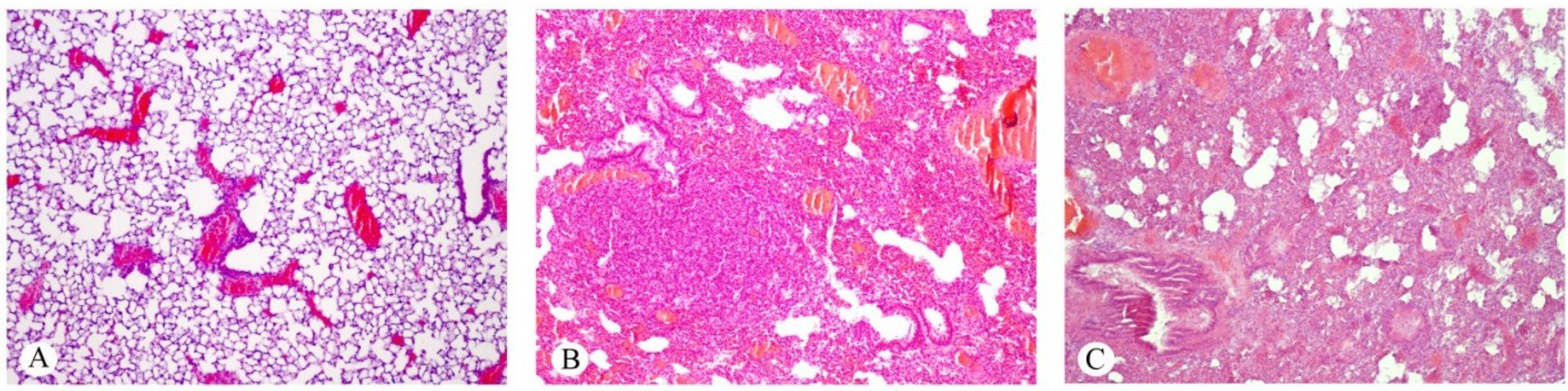
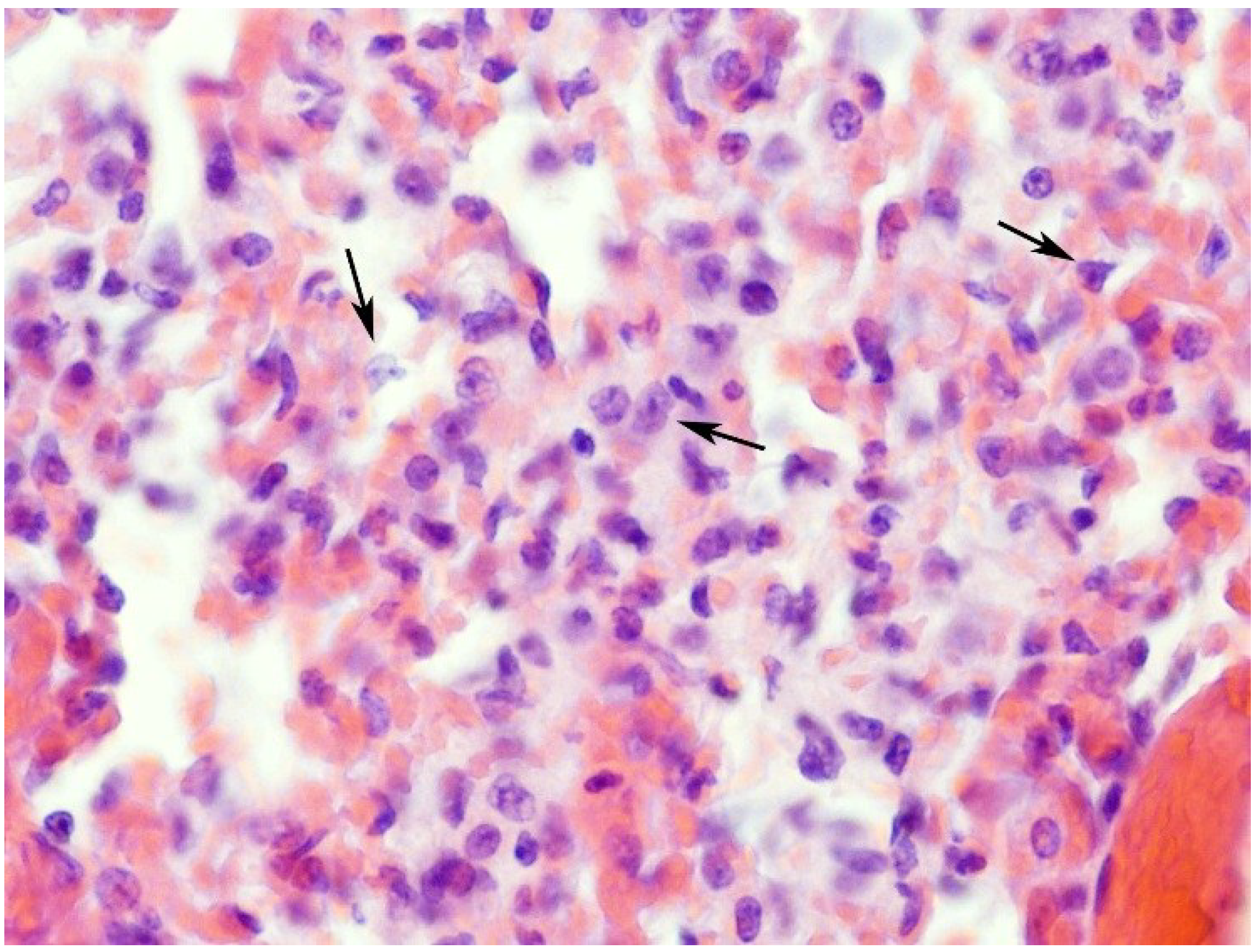
2.1. Effects on the Lung



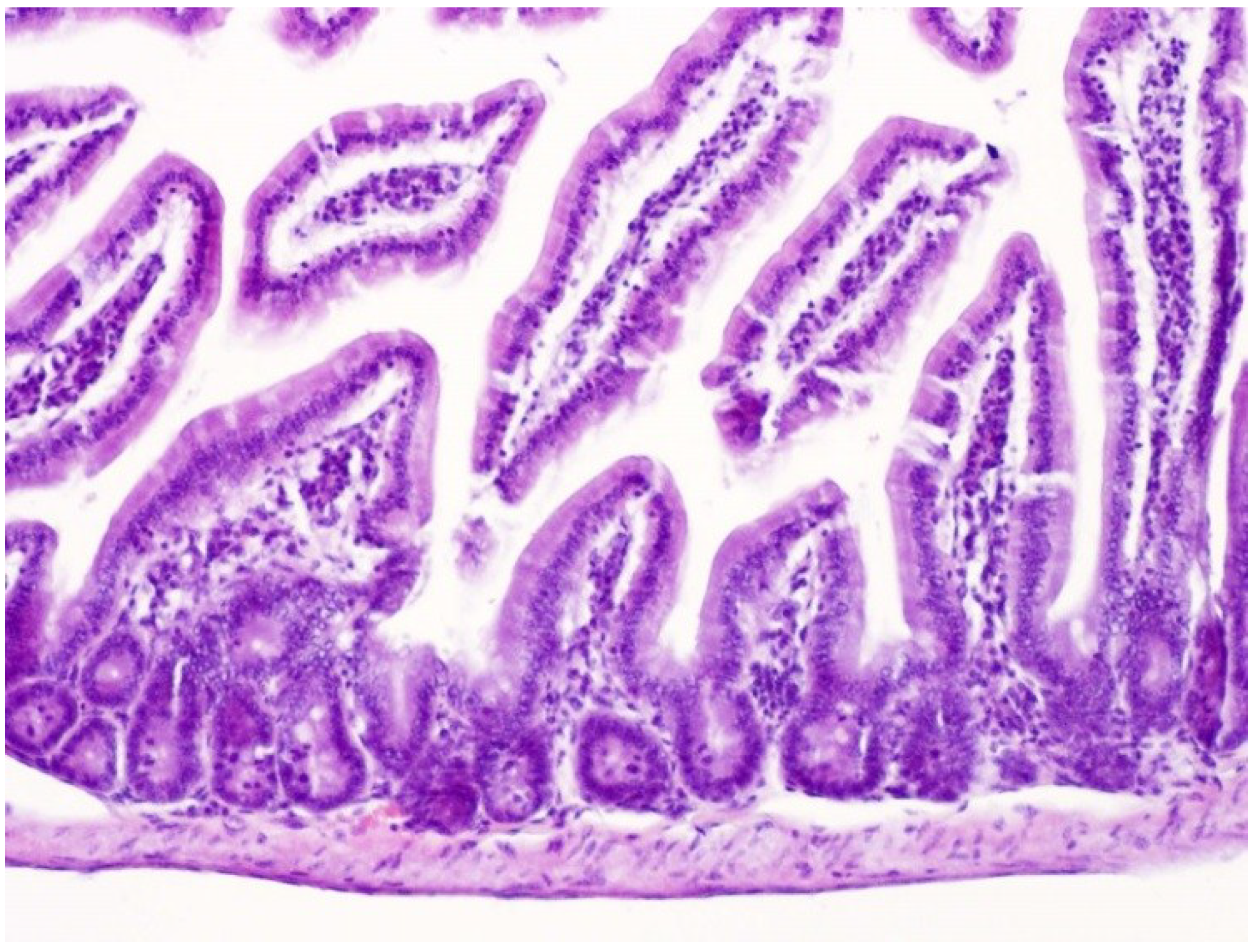
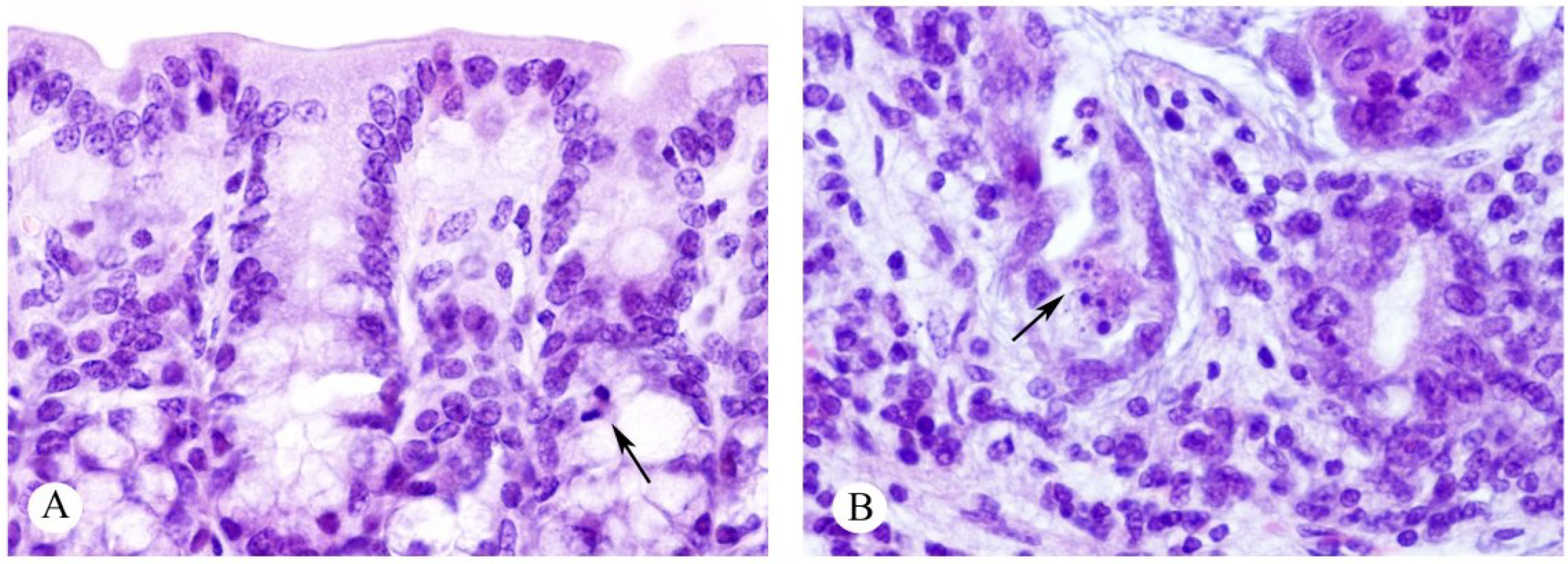
2.2. Effects on the Intestines



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organ | Control 6 m | Control 12 m | 6 m | 12 m |
|---|---|---|---|---|
| Small intestine | 32.20 ± 3.63 * | 34.10 ± 1.20 | 28.00 ± 2.40 * | 29.75 ± 1.70 |
| Large intestine | 9.42 ± 0.57 | 11.01 ± 0.70 # | 8.75 ± 0.84 | 8.75 ± 0.86 # |
3. Discussion
4. Experimental Section
4.1. Materials
4.2. Isolation of Ebulin f from S. ebulus Fruits
4.3. Animals and Experimental Groups
4.4. Treatment with Ebulin f
4.5. Histological Analysis
4.6. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Takahashi, T. Flow behavior of digesta and the absorption of nutrients in the gastro intestine. J. Nutr. Sci. Vitaminol. 2011, 57, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Bornhorst, G.M.; Paul Singh, R. Gastric digestion in vivo and in vitro: How the structural aspects of food influence the digestion process. Annu. Rev. Food Sci. Technol. 2014, 5, 111–132. [Google Scholar] [CrossRef] [PubMed]
- Cervantes, S. Cellular and molecular mechanisms of intestinal elongation in mammals: The long and short of it. Histol. Histopathol. 2013, 28, 427–436. [Google Scholar] [PubMed]
- Ramachandran, A.; Madesh, M.; Balasubramanian, K.A. Apoptosis in the intestinal epithelium: Its relevance in normal and pathophysiological conditions. J. Gastroenterol. Hepatol. 2000, 15, 109–200. [Google Scholar] [CrossRef] [PubMed]
- Ciccocioppo, R.; Di Sabatino, A.; Luinetti, O.; Rossi, M.; Cifone, M.G.; Corazza, G.R. Small bowel enterocyte apoptosis and proliferation are increased in the elderly. Gerontology 2002, 48, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, J.; Li, Q.; Zhang, J.; Duan, X.L. Morphological changes of cell proliferation and apoptosis in rat jejunal mucosa at different ages. World J. Gastroenterol. 2003, 9, 2060–2064. [Google Scholar] [PubMed]
- Drozdowski, L.; Thomson, A.B. Aging and the intestine. World J. Gastroenterol. 2006, 12, 7578–7584. [Google Scholar] [PubMed]
- Feibusch, J.M.; Holt, P.R. Impaired absorptive capacity for carbohydrate in the aging human. Dig. Dis. Sci. 1982, 27, 1095–1100. [Google Scholar] [CrossRef] [PubMed]
- Coudray, C.; Feillet-Coudray, C.; Rambeau, M.; Tressol, J.C.; Gueux, E.; Mazur, A.; Rayssiguier, Y. The effect of aging on intestinal absorption and status of calcium, magnesium, zinc, and copper in rats: A stable isotope study. J. Trace Elem. Med. Biol. 2006, 20, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, P.; Tejero, J.; Cabrero, P.; Cordoba-Diaz, D.; Girbes, T. Differential sensitivity of d-galactose-binding lectins from fruits of dwarf elder (Sambucus ebulus L.) to a simulated gastric fluid. Food Chem. 2013, 136, 794–802. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, P.; Gayoso, M.J.; Tejero, J.; Cabrero, P.; Cordoba-Diaz, D.; Basterrechea, J.E.; Girbés, T. Toxicity in mice of lectin ebulin f present in dwarf elderberry (Sambucus ebulus L.). Toxicon 2013, 61, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Kiselova, Y.; Ivanova, D.; Chervenkov, T.; Gerova, D.; Galunska, B.; Yankova, T. Correlation between the in vitro antioxidant activity and polyphenol content of aqueous extracts from bulgarian herbs. Phytother. Res. 2006, 20, 961–965. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, D.; Tasinov, O.; Kiselova-Kaneva, Y. Improved lipid profile and increased serum antioxidant capacity in healthy volunteers after Sambucus ebulus L. fruit infusion consumption. Int. J. Food Sci. Nutr. 2014, 65, 740–744. [Google Scholar] [CrossRef] [PubMed]
- Yesilada, E.; Gürbüz, I.; Toker, G. Anti-ulcerogenic activity and isolation of the active principles from Sambucus ebulus L. leaves. J. Ethnopharmacol. 2014, 153, 478–483. [Google Scholar] [CrossRef] [PubMed]
- Rahimi-Esboei, B.; Ebrahimzadeh, M.A.; Gholami, Sh.; Falah-Omrani, V. Anti-giardial activity of Sambucus ebulus. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 2047–2050. [Google Scholar] [PubMed]
- Roschek, B.; Fink, R.C.; McMichael, M.D.; Li, D.; Alberte, R.S. Elderberry flavonoids bind to and prevent H1N1 infection in vitro. Phytochemistry 2009, 70, 1255–1261. [Google Scholar] [CrossRef] [PubMed]
- Suntar, I.P.; Akkol, E.K.; Yalcin, F.N.; Koca, U.; Keles, H.; Yesilada, E. Wound healing potential of Sambucus ebulus L. leaves and isolation of an active component, quercetin 3-O-glucoside. J. Ethnopharmacol. 2010, 129, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, P.; Cabrero, P.; Basterrechea, J.E.; Tejero, J.; Cordoba-Diaz, D.; Girbes, T. Isolation and molecular characterization of two lectins from dwarf elder (Sambucus ebulus L.) blossoms related to the Sam n1 allergen. Toxins 2013, 5, 1767–1779. [Google Scholar] [CrossRef] [PubMed]
- Girbes, T.; Ferreras, J.M.; Arias, F.J.; Stirpe, F. Description, distribution, activity and phylogenetic relationship of ribosome-inactivating proteins in plants, fungi and bacteria. Mini Rev. Med. Chem. 2004, 4, 461–476. [Google Scholar] [CrossRef] [PubMed]
- Stirpe, F. Ribosome-inactivating proteins. Toxicon 2004, 44, 371–383. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.B.; Wong, J.H.; Wang, H. Recent progress in research on ribosome inactivating proteins. Curr. Protein Pept. Sci. 2010, 11, 37–53. [Google Scholar] [CrossRef] [PubMed]
- Lord, J.M.; Spooner, R.A. Ricin trafficking in plant and mammalian cells. Toxins 2011, 3, 787–801. [Google Scholar] [CrossRef] [PubMed]
- Pascal, J.M.; Day, P.J.; Monzingo, A.F.; Ernst, S.R.; Robertus, J.D.; Iglesias, R.; Pérez, Y.; Ferreras, J.M.; Citores, L.; Girbes, T. 2.8-A Crystal structure of a nontoxic type-II ribosome-inactivating protein, ebulin l. Proteins 2001, 43, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Citores, L.; de Benito, F.M.; Iglesias, R.; Miguel Ferreras, J.; Argüeso, P.; Jimenez, P.; Méndez, E.; Girbes, T. Presence of polymerized and free forms of the non-toxic type 2 ribosome-inactivating protein ebulin and a structurally related new homodimeric lectin in fruits of Sambucus ebulus L. Planta 1998, 204, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, P.; Gayoso, M.J.; Garrosa, M.; Córdoba-Díaz, D.; Cabrero, P.; Tejero, J.; Aracil, M.; Girbés, T. Paneth cells are also target of the ribotoxic lectin nigrin b. Histol. Histopathol. 2014, 29, 1057–1063. [Google Scholar] [PubMed]
- Walsh, M.J.; Dodd, J.E.; Hautbergue, G.M. Ribosome-inactivating proteins: Potent poisons and molecular tools. Virulence 2013, 4, 774–784. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M.; Rasooly, R. Review of the inhibition of biological activities of food-related selected toxins by natural compounds. Toxins 2013, 5, 743–775. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, G.D. Understanding ricin from a defensive viewpoint. Toxins 2011, 3, 1373–1392. [Google Scholar] [CrossRef] [PubMed]
- Lord, M.J.; Jolliffe, N.A.; Marsden, C.J.; Pateman, C.S.; Smith, D.C.; Spooner, R.A.; Watson, P.D.; Roberts, L.M. Ricin. Toxicol. Rev. 2003, 22, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Battelli, M.G.; Musiani, S.; Buonamici, L.; Santi, S.; Riccio, M.; Maraldi, N.M.; Girbés, T.; Stirpe, F. Interaction of volkensin with HeLa cells: Binding, uptake, intracellular localization, degradation and exocytosis. Cell Mol. Life Sci. 2004, 61, 1975–1984. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, R.; Arias, Y.; Ferreras, J.M.; Jiménez, P.; Rojo, M.A.; Girbés, T. Sensitivity of cancer cell lines to the novel non-toxic type 2 ribosome-inactivating protein nigrin b. Cancer Lett. 2001, 167, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Citores, L.; Ferreras, J.M.; Muñoz, R.; Benítez, J.; Jiménez, P.; Girbés, T. Targeting cancer cells with transferrin conjugates containing the non-toxic type 2 ribosome-inactivating proteins nigrin b or ebulin l. Cancer Lett. 2002, 184, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Benítez, J.; Ferreras, J.M.; Muñoz, R.; Arias, Y.; Iglesias, R.; Córdoba-Díaz, M.; del Villar, R.; Girbés, T. Cytotoxicity of an ebulin l-anti-human CD105 immunotoxin on mouse fibroblasts (L929) and rat myoblasts (L6E9) cells expressing human CD105. Med. Chem. 2005, 1, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, T.; Takeyama, Y.; Ueda, T.; Shinzeki, M.; Sawa, H.; Nakajima, T.; Kuroda, Y. Breakdown of intestinal mucosa via accelerated apoptosis increases intestinal permeability in experimental severe acute pancreatitis. J. Surg. Res. 2006, 135, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Horrix, C.; Raviv, Z.; Flescher, E.; Voss, C.; Berger, M.R. Plant ribosome-inactivating proteins type II induce the unfolded protein response in human cancer cells. Cell Mol. Life Sci. 2011, 68, 1269–1281. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Han, C.; Du, J.; Zhao, S.; Fu, Y.; Zheng, G.; Sun, Y.; Zhang, Y.; Liu, W.; Wan, J.; et al. Proteomic study of differential protein expression in mouse lung tissues after aerosolized ricin poisoning. Int. J. Mol. Sci. 2014, 15, 7281–7292. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, P.; Cabrero, P.; Basterrechea, J.E.; Tejero, J.; Cordoba-Diaz, D.; Cordoba-Diaz, M.; Girbes, T. Effects of short-term heating on total polyphenols, anthocyanins, antioxidant activity and lectins of different parts of dwarf elder (Sambucus ebulus L.). Plant Foods Hum. Nutr. 2014, 69, 168–174. [Google Scholar] [CrossRef] [PubMed]
- European Parliament and the Council. Available online: http://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex:32010L0063 (accessed on 27 November 2014).
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garrosa, M.; Jiménez, P.; Tejero, J.; Cabrero, P.; Cordoba-Diaz, D.; Quinto, E.J.; Gayoso, M.J.; Girbés, T. Toxicity of the Anti-ribosomal Lectin Ebulin f in Lungs and Intestines in Elderly Mice. Toxins 2015, 7, 367-379. https://doi.org/10.3390/toxins7020367
Garrosa M, Jiménez P, Tejero J, Cabrero P, Cordoba-Diaz D, Quinto EJ, Gayoso MJ, Girbés T. Toxicity of the Anti-ribosomal Lectin Ebulin f in Lungs and Intestines in Elderly Mice. Toxins. 2015; 7(2):367-379. https://doi.org/10.3390/toxins7020367
Chicago/Turabian StyleGarrosa, Manuel, Pilar Jiménez, Jesús Tejero, Patricia Cabrero, Damián Cordoba-Diaz, Emiliano J. Quinto, Manuel J. Gayoso, and Tomás Girbés. 2015. "Toxicity of the Anti-ribosomal Lectin Ebulin f in Lungs and Intestines in Elderly Mice" Toxins 7, no. 2: 367-379. https://doi.org/10.3390/toxins7020367
APA StyleGarrosa, M., Jiménez, P., Tejero, J., Cabrero, P., Cordoba-Diaz, D., Quinto, E. J., Gayoso, M. J., & Girbés, T. (2015). Toxicity of the Anti-ribosomal Lectin Ebulin f in Lungs and Intestines in Elderly Mice. Toxins, 7(2), 367-379. https://doi.org/10.3390/toxins7020367