
Dose-Dependent Effects on Sphingoid Bases and Cytokines in Chickens Fed Diets Prepared with Fusarium Verticillioides Culture Material Containing Fumonisins

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Effect of FB on the Accumulation of Free Sphingoid Bases and the Sa/So Ratio: Sensitivity of Tissues and Dose-Response Effect

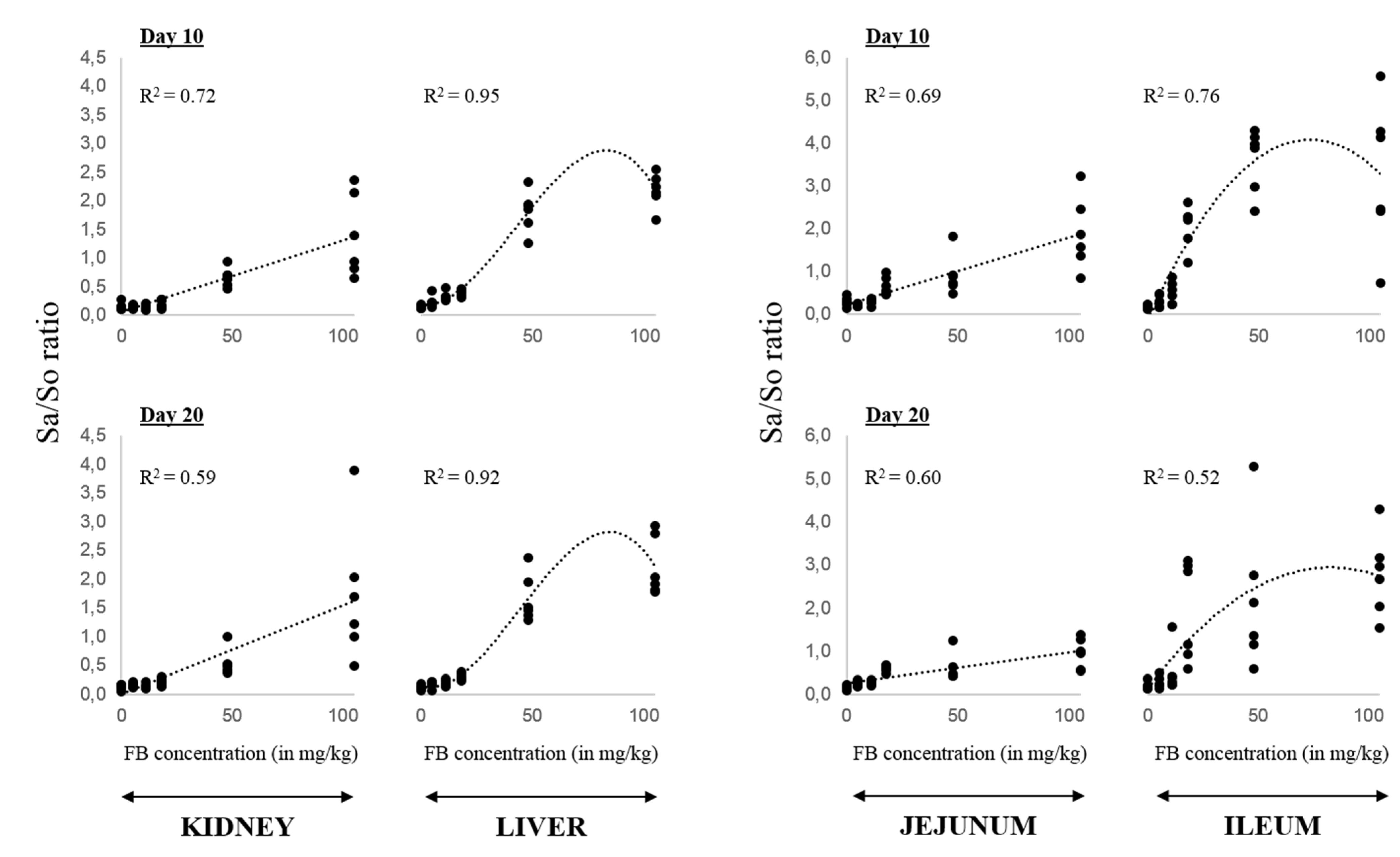{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sa/So ratios | Liver | Kidney | Jejunum | Ileum | Cecum | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| FB in Diet (mg/kg) | d10 | d20 | d10 | d20 | d10 | d20 | d10 | d20 | d10 | d20 |
| 5.6 mg/kg | 0.23 ± 0.04 | 0.17 ± 0.02 | 0.14 ± 0.01 | 0.16 ± 0.02 | 0.22 ± 0.01 | 0.25 ± 0.02 | 0.31 ± 0.06 | 0.27 ± 0.05 | 0.14 ± 0.02 | 0.19 ± 0.04 |
| 11.3 mg/kg | 0.33 ± 0.03 ** | 0.19 ± 0.02 | 0.14 ± 0.02 | 0.14 ± 0.02 | 0.29 ± 0.03 | 0.28 ± 0.02 * | 0.51 ± 0.11 | 0.52 ± 0.21 | 0.23 ± 0.01 | 0.13 ± 0.01 * |
| 17.5 mg/kg | 0.40 ± 0.02 *** | 0.31 ± 0.02 ** | 0.20 ± 0.03 | 0.21 ± 0.03 | 0.67 ± 0.08 * | 0.60 ± 0.03 *** | 2.06 ± 0.20 ** | 1.93 ± 0.47 * | 0.42 ± 0.03 ** | 0.35 ± 0.03 ** |
| 47.8 mg/kg | 1.82 ± 0.15 ** | 1.66 ± 0.17 ** | 0.66 ± 0.07 ** | 0.55 ± 0.09 * | 0.93 ± 0.19 | 1.12 ± 0.49 | 3.61 ± 0.30 ** | 2.37 ± 0.80 | 0.75 ± 0.18 | 0.38 ± 0.02 ** |
| 104.8 mg/kg | 2.18 ± 0.12 *** | 2.22 ± 0.21 ** | 1.39 ± 0.29 * | 1.72 ± 0.48 | 1.89 ± 0.34 * | 0.95 ± 0.14 * | 4.08 ± 1.00 * | 2.88 ± 0.48 * | 0.62 ± 0.08 * | 0.44 ± 0.04 *** |
| Probability of diet effect | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.011 | <0.001 | <0.001 | <0.001 | <0.001 |

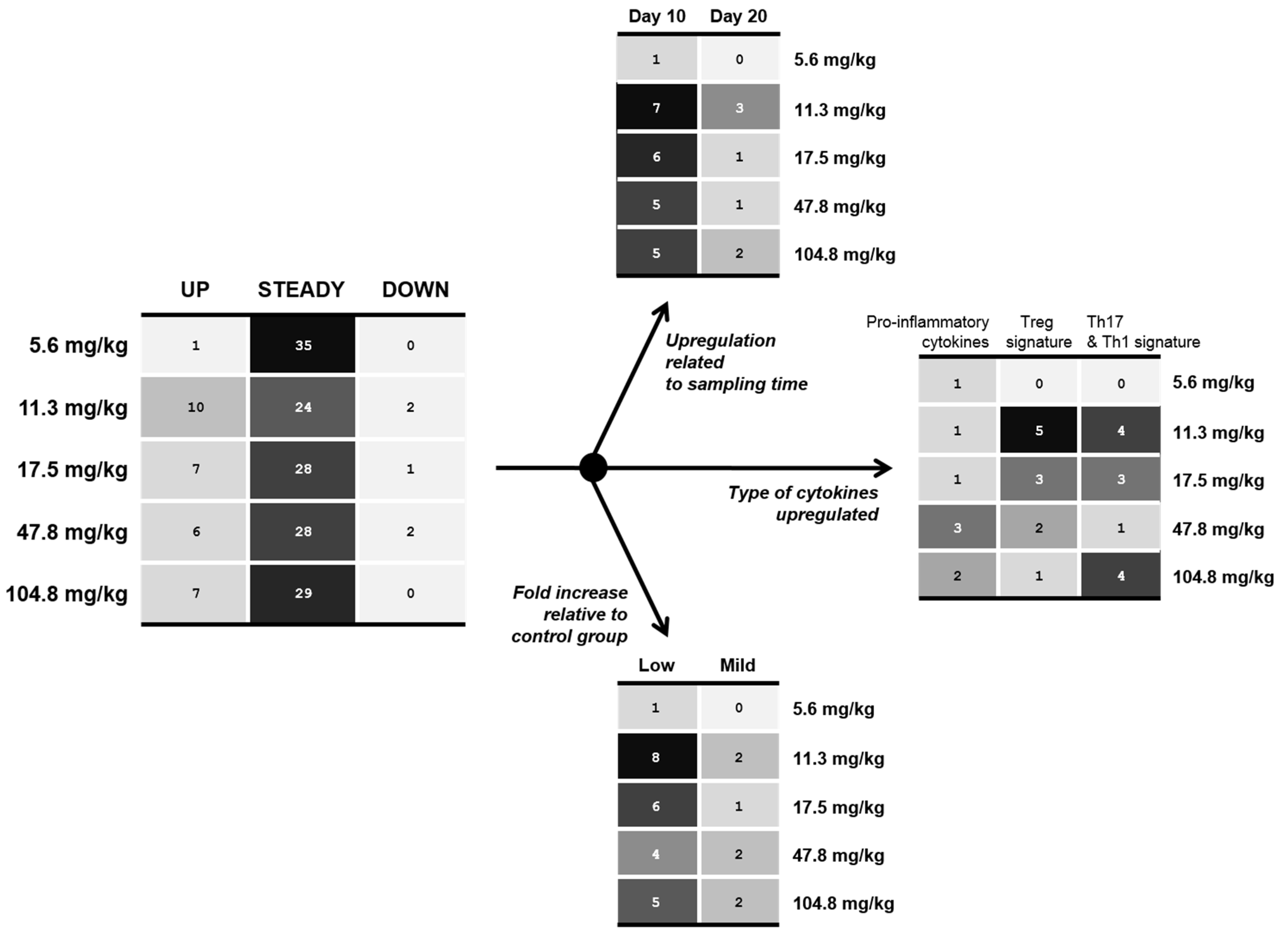
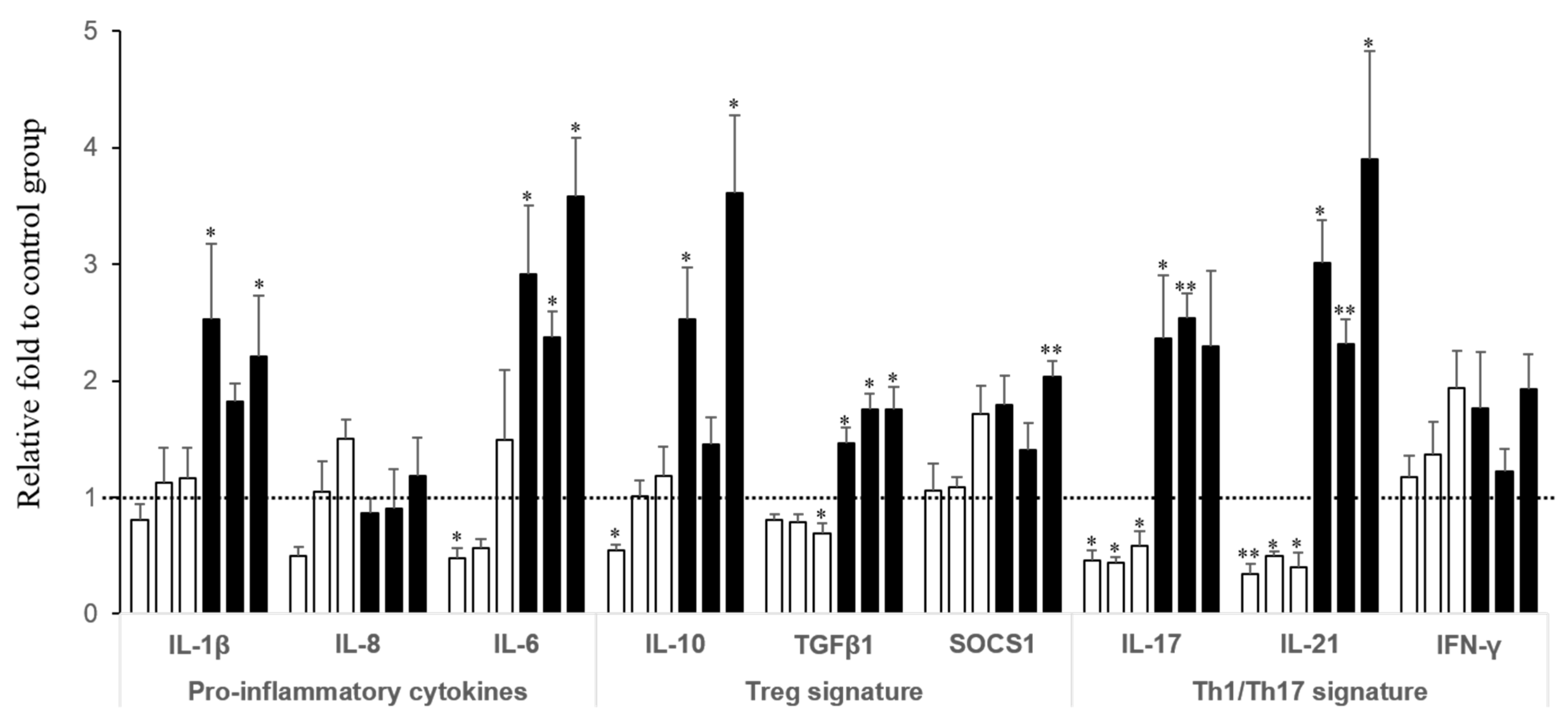
2.2. Effect of FB on the Intestinal Immune Defense: Gene Expression in Local and Specific Tissues and Dose-Response Effect
| FB in diet (mg/kg) | Pro-inflammatory cytokines | Treg signature | Th1 & Th17 signature | ||||||
|---|---|---|---|---|---|---|---|---|---|
| IL-1β | IL-8 | IL-6 | IL-10 | TGFβ1 | SOCS1 | IL-17 | IL-21 | IFN-γ | |
| Jejunum d10 | |||||||||
| 5.6 | 2.11 ± 0.35 ** | 1.02 ± 0.14 | 1.15 ± 0.25 | 1.40 ± 0.30 | 1.12 ± 0.14 | 1.20 ± 0.11 | 1.20 ± 0.31 | 1.53 ± 0.30 | 1.29 ± 0.16 |
| 11.3 | 2.39 ± 0.26 *** | 1.74 ± 0.40 | 0.95 ± 0.13 | 3.76 ± 0.65 ** | 0.94 ± 0.07 | 1.29 ± 0.13 | 2.32 ± 0.22 * | 1.69 ± 0.20 | 1.61 ± 0.15 |
| 17.5 | 1.96 ± 0.14 ** | 1.29 ± 0.23 | 1.52 ± 0.14 | 3.45 ± 0.82 * | 1.12 ± 0.11 | 1.32 ± 0.19 | 1.58 ± 0.50 | 2.40 ± 0.63 * | 2.17 ± 0.48 ** |
| 47.8 | 2.08 ± 0.20 ** | 1.63 ± 0.18 | 1.82 ± 0.36 | 2.07 ± 0.59 | 0.98 ± 0.10 | 1.39 ± 0.31 | 1.09 ± 0.17 | 1.78 ± 0.19 | 1.61 ± 0.24 |
| 104.8 | 1.80 ± 0.16 * | 2.54 ± 0.36 *** | 2.37 ± 0.96 | 2.40 ± 0.61 | 1.06 ± 0.12 | 1.31 ± 0.11 | 0.82 ± 0.09 | 2.19 ± 0.33 * | 1.60 ± 0.25 |
| Probability of the diet effect | 0.004 | 0.004 | 0.199 | 0.014 | 0.765 | 0.677 | 0.021 | 0.101 | 0.116 |
| Jejunum d20 | |||||||||
| 5.6 | 1.00 ± 0.30 | 0.94 ± 0.39 | 0.88 ± 0.10 | 0.50 ± 0.14 | 1.09 ± 0.13 | 0.90 ± 0.18 | 1.56 ± 0.77 | 1.10 ± 0.29 | 0.65 ± 0.13 |
| 11.3 | 1.66 ± 0.47 | 2.20 ± 0.81 | 1.85 ± 0.42 | 0.86 ± 0.09 | 1.54 ± 0.12 * | 2.03 ± 0.50 * | 4.47 ± 1.36 * | 1.52 ± 0.20 | 1.51 ± 0.09 |
| 17.5 | 0.69 ± 0.09 | 1.17 ± 0.32 | 0.70 ± 0.09 | 0.82 ± 0.27 | 1.83 ± 0.20 ** | 1.18 ± 0.28 | 0.59 ± 0.11 | 1.27 ± 0.33 | 1.23 ± 0.27 |
| 47.8 | 0.55 ± 0.05 * | 1.47 ± 0.71 | 2.24 ± 1.07 | 0.84 ± 0.43 | 1.17 ± 0.09 | 0.89 ± 0.14 | 0.77 ± 0.23 | 1.49 ± 0.73 | 1.11 ± 0.48 |
| 104.8 | 1.13 ± 0.15 | 2.14 ± 0.61 | 1.13 ± 0.24 | 0.57 ± 0.08 | 1.73 ± 0.18 ** | 1.60 ± 0.22 | 3.41 ± 1.57 | 1.26 ± 0.25 | 1.04 ± 0.09 |
| Probability of the diet effect | 0.069 | 0.470 | 0.258 | 0.792 | 0.003 | 0.052 | 0.050 | 0.919 | 0.269 |
| Ileum d10 | |||||||||
| 5.6 | 1.55 ± 0.27 | 1.81 ± 0.57 | 0.59 ± 0.09 | 1.25 ± 0.20 | 1.08 ± 0.11 | 1.67 ± 0.24 | 1.72 ± 0.41 | 1.32 ± 0.19 | 1.90 ± 0.46 |
| 11.3 | 1.51 ± 0.08 | 1.89 ± 0.23 | 0.16 ± 0.04 * | 1.32 ± 0.30 | 1.63 ± 0.15 * | 2.13 ± 0.12 * | 2.07 ± 0.25 * | 1.00 ± 0.17 | 1.85 ± 0.19 * |
| 17.5 | 1.49 ± 0.27 | 2.58 ± 0.50 | 0.18 ± 0.03 * | 1.16 ± 0.21 | 1.43 ± 0.21 | 2.18 ± 0.22 * | 1.18 ± 0.31 | 0.56 ± 0.13 | 1.84 ± 0.16 * |
| 47.8 | 1.85 ± 0.20 * | 2.70 ± 0.39 * | 0.27 ± 0.04 * | 1.10 ± 0.23 | 1.27 ± 0.09 | 2.33 ± 0.49 * | 0.96 ± 0.16 | 0.66 ± 0.17 | 2.24 ± 0.22 * |
| 104.8 | 1.48 ± 0.33 | 1.45 ± 0.21 | 0.54 ± 0.07 | 1.19 ± 0.26 | 1.55 ± 0.42 | 1.94 ± 0.50 | 0.92 ± 0.22 | 1.65 ± 0.14 * | 2.11 ± 0.55 * |
| Probability of the diet effect | 0.241 | 0.046 | <0.001 | 0.947 | 0.331 | 0.093 | 0.035 | 0.008 | 0.221 |
| Ileum d20 | |||||||||
| 5.6 | 0.85 ± 0.18 | 0.76 ± 0.23 | 0.82 ± 0.13 | 0.58 ± 0.14 | 0.94 ± 0.10 | 0.67 ± 0.11 | 1.78 ± 0.45 | 0.76 ± 0.16 | 0.80 ± 0.18 |
| 11.3 | 0.74 ± 0.11 | 0.41 ± 0.08 * | 0.81 ± 0.06 | 0.61 ± 0.15 | 1.17 ± 0.13 | 0.70 ± 0.06 | 1.46 ± 0.23 | 0.92 ± 0.23 | 0.75 ± 0.15 |
| 17.5 | 0.79 ± 0.10 | 0.49 ± 0.19 | 0.90 ± 0.10 | 1.27 ± 0.34 | 1.11 ± 0.24 | 0.81 ± 0.15 | 1.62 ± 0.47 | 0.81 ± 0.14 | 1.23 ± 0.31 |
| 47.8 | 1.35 ± 0.34 | 0.66 ± 0.19 | 0.84 ± 0.07 | 2.85 ± 1.24 * | 1.40 ± 0.13 | 1.22 ± 0.29 | 2.09 ± 0.69 | 2.11 ± 0.36 | 1.38 ± 0.29 |
| 104.8 | 1.40 ± 0.26 | 1.53 ± 0.37 | 0.74 ± 0.09 | 2.19 ± 0.37 | 0.93 ± 0.15 | 1.10 ± 0.15 | 3.69 ± 0.74 ** | 1.14 ± 0.11 | 1.53 ± 0.38 |
| Probability of the diet effect | 0.135 | 0.026 | 0.772 | 0.091 | 0.300 | 0.215 | 0.025 | 0.001 | 0.257 |


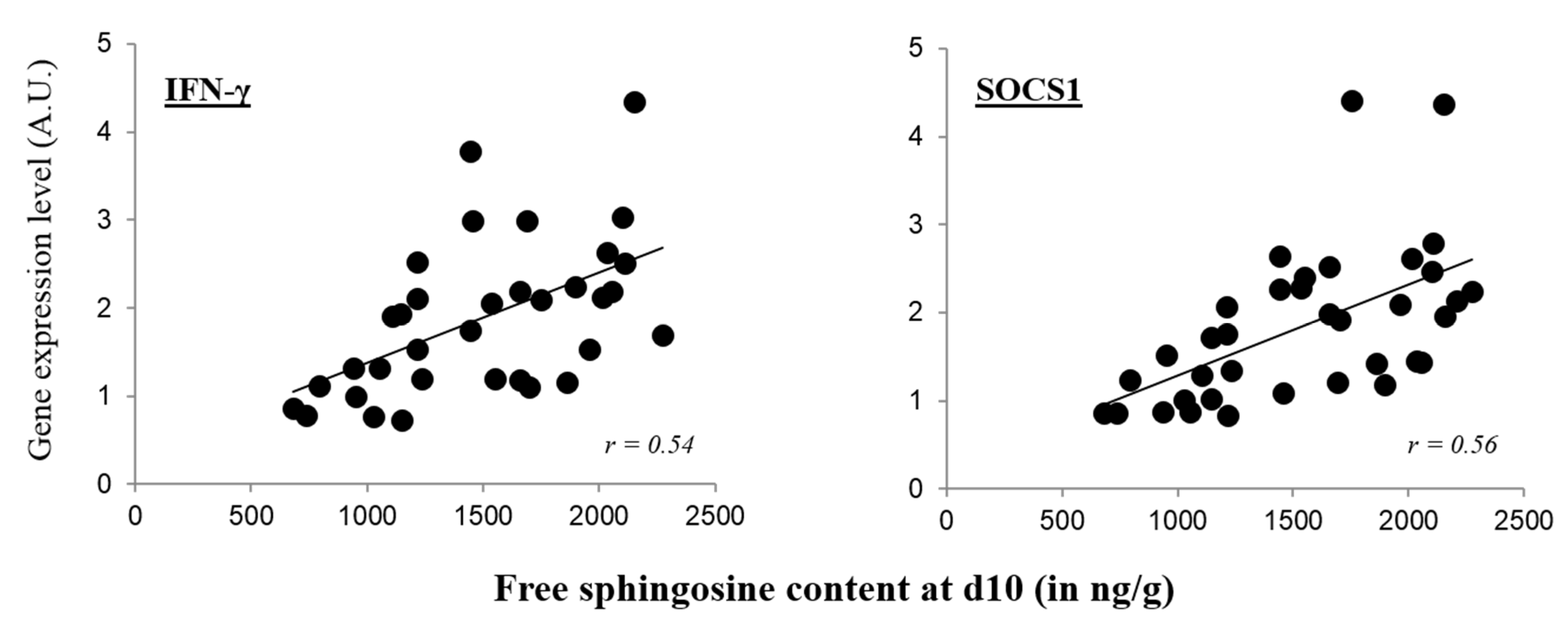
2.3. Correlation of Free Sphingosine and Immune Factors

3. Discussion
4. Experimental Section
4.1. Experimental Birds, Housing, Diet Formulation and Sampling
| Item | Starter Diet | |||||
|---|---|---|---|---|---|---|
| Ingredient (% of diet) | ||||||
| Corn | 54.18 | |||||
| Soybean meal (48% CP) | 38.05 | |||||
| Soy oil | 3.52 | |||||
| Sodium chloride | 0.47 | |||||
| DL-Methionine | 0.25 | |||||
| Threonine | 0.07 | |||||
| l-Lysine, HCl | 0.10 | |||||
| Limestone | 1.68 | |||||
| Monocalcium phosphate | 1.33 | |||||
| Vitamin and mineral premix 1 | 0.35 | |||||
| Nutrient composition (calculated) | ||||||
| ME, kcal/kg | 3066 | |||||
| CP, % | 22.43 | |||||
| Ca, % | 1.01 | |||||
| Non-phytate phosphorus, % | 0.43 | |||||
| Met, % | 0.59 | |||||
| Thr, % | 0.92 | |||||
| Lys, % | 1.34 | |||||
| Analyzed composition | Target and actual concentration of mycotoxins in the formulated diets (in mg/kg of feed) | |||||
| 0 | 5 | 10 | 20 | 50 | 100 | |
| Fumonisin B1 | 0.26 | 3.9 | 7.1 | 11.0 | 31.7 | 66.9 |
| Fumonisin B2 | 0.09 | 1.4 | 3.4 | 5.1 | 12.4 | 27.5 |
| Fumonisin B3 | 0.02 | 0.3 | 0.8 | 1.4 | 3.7 | 10.4 |
| Total Fumonisins | 0.37 | 5.6 | 11.3 | 17.5 | 47.8 | 104.8 |
| Deoxynivalenol | Range from 0.236 to 0.344 mg/kg | |||||
| Zearalenone | Range from 0.015 to 0.029 mg/kg | |||||
| Aflatoxin B1 | <LOD; LOD = 0.3 μg/kg | |||||
| Ochratoxin A | <LOD; LOD = 0.2 μg/kg | |||||
| T-2 toxin | <LOD; LOD = 25 μg/kg | |||||
4.2. Experimental Parameter Measures
4.2.1. Sphingolipid Analysis (day 10 & 20)
4.2.2. Gene Expression (day 10 & 20)
| Gene | Primer sequence | Amplicon | Ensembl access | References | |
|---|---|---|---|---|---|
| Size | Intron 1 | ||||
| Housekeeping genes | |||||
| GAPDH | F (300 nM) TCCTAGGATACACAGAGGACCA R (300 nM) CGGTTGCTATATCCAAACTCA | 151 bp | 2(499) | ENSGALG00000014442 | Present study |
| RPL4 | F (300 nM) TTATGCCATCTGTTCTGCC R (300 nM) GCGATTCCTCATCTTACCCT | 235 bp | 2(893) | ENSGALG00000007711 | Present study |
| Pro-inflammatory cytokines | |||||
| IL-1β | F (300 nM) GCATCAAGGGCTACAAGCTC R (300 nM) CAGGCGGTAGAAGATGAAGC | 131 bp | 1(87) | ENSGALG00000000534 | [48] |
| IL-6 | F (300 nM) GAATGTTTTAGTTCGGGCACA R (300 nM) TTCCTAGAAGGAAATGAGAATGC | 130 bp | 0 | ENSGALG00000010915 | Present study |
| IL-8 | F (300 nM) GCGGCCCCCACTGCAAGAAT R (300 nM) TCACAGTGGTGCATCAGAATTGAGC | 146 bp | 2(1210) | ENSGALG00000011670 | Present study |
| Treg signature | |||||
| IL-10 | F (300 nM) GCTGAGGGTGAAGTTTGAGG R (300 nM) AGACTGGCAGCCAAAGGTC | 121 bp | 2(1127) | ENSGALG00000000892 | Present study |
| TGFβ1 | F (300 nM) CGGGACGGATGAGAAGAAC R (300 nM) CGGCCCACGTAGTAAATGAT | 258 bp | no data | [49] | |
| SOCS1 | F (300 nM) CAAGCGGATTTCAGTAGCATC R (300 nM) GGCTCAGACTTCAGCTTCTCA | 110 bp | no intron | ENSGALG00000007158 | Present study |
| Th17 & Th1 signature | |||||
| IL-17 | F (300 nM) TATCAGCAAACGCTCACTGG R (300 nM) AGTTCACGCACCTGGAATG | 110 bp | 1(666) | ENSGALG00000016678 | Present study |
| IL-21 | F (300 nM) GCTTTCAAAGACAATTGACCATC R (300 nM) TACAGCTGTGAGCAGGCATC | 106 bp | 2(3765) | ENSGALG00000011844 | Present study |
| IFN-γ | F (300 nM) AGCTGACGGTGGACCTATTATT R (300 nM) GGCTTTGCGCTGGATTC | 259 bp | 2(998) | ENSGALG00000009903 | [49] |
4.3. Statistics
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Abbreviations
| CT | Cecal Tonsils |
| FB | Fumonisins |
| GIT | Gastrointestinal Tract |
| Sa | Sphinganine |
| So | Sphingosine |
| So-1-P | Sphingosine-1-Phosphate |
Conflicts of Interest
References
- Bryden, W.L. Mycotoxin contamination of the feed supply chain: Implications for animal productivity and feed security. Anim. Feed Sci. Technol. 2012, 173, 134–158. [Google Scholar] [CrossRef]
- Rodrigues, I.; Naehrer, K. A three-year survey on the worldwide occurrence of mycotoxins in feedstuffs and feed. Toxins 2012, 4, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Streit, E.; Naehrer, K.; Rodrigues, I.; Schatzmayr, G. Mycotoxin occurrence in feed and feed raw materials worldwide: Long-term analysis with special focus on europe and asia. J. Sci. Food Agric. 2013, 93, 2892–2899. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Munkvold, G.P. Mycotoxins in ethanol co-products: Modeling economic impacts on the livestock industry and management strategies. J. Agric. Food Chem. 2008, 56, 3900–3911. [Google Scholar] [CrossRef] [PubMed]
- Grenier, B.; Applegate, T. Modulation of intestinal functions following mycotoxin ingestion: Meta-analysis of published experiments in animals. Toxins 2013, 5, 396–430. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.; Norred, W.P.; Bacon, C.W.; Riley, R.T.; Merrill, A.H. Inhibition of sphingolipid biosynthesis by fumonisins. Implications for diseases associated with fusarium moniliforme. J. Biol. Chem. 1991, 266, 14486–14490. [Google Scholar] [PubMed]
- Merrill, A.H.; Sullards, M.C.; Wang, E.; Voss, K.A.; Riley, R.T. Sphingolipid metabolism: Roles in signal transduction and disruption by fumonisins. Environ. Health Perspect. 2001, 109, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Enongene, E.N.; Sharma, R.P.; Bhandari, N.; Voss, K.A.; Riley, R.T. Disruption of sphingolipid metabolism in small intestines, liver and kidney of mice dosed subcutaneously with fumonisin B1. Food Chem. Toxicol. 2000, 38, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Enongene, E.N.; Sharma, R.P.; Bhandari, N.; Miller, J.D.; Meredith, F.I.; Voss, K.A.; Riley, R.T. Persistence and reversibility of the elevation in free sphingoid bases induced by fumonisin inhibition of ceramide synthase. Toxicol. Sci. 2002, 67, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Loiseau, N.; Debrauwer, L.; Sambou, T.; Bouhet, S.; Miller, J.D.; Martin, P.G.; Viadère, J.-L.; Pinton, P.; Puel, O.; Pineau, T.; et al. Fumonisin B1 exposure and its selective effect on porcine jejunal segment: Sphingolipids, glycolipids and trans-epithelial passage disturbance. Biochem. Pharmacol. 2007, 74, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Henry, M.H.; Wyatt, R.D.; Fletchert, O.J. The toxicity of purified fumonisin B1 in broiler chicks. Poult. Sci. 2000, 79, 1378–1384. [Google Scholar] [CrossRef] [PubMed]
- Broomhead, J.N.; Ledoux, D.R.; Bermudez, A.J.; Rottinghaus, G.E. Chronic effects of fumonisin B1 in broilers and turkeys fed dietary treatments to market age. Poult. Sci. 2002, 81, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Haschek, W.M.; Gumprecht, L.A.; Smith, G.; Tumbleson, M.E.; Constable, P.D. Fumonisin toxicosis in swine: An overview of porcine pulmonary edema and current perspectives. Environ. Health Perspect. 2001, 109, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Tran, S.T.; Auvergne, A.; Benard, G.; Bailly, J.D.; Tardieu, D.; Babile, R.; Guerre, P. Chronic effects of fumonisin B1 on ducks. Poult. Sci. 2005, 84, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Benlasher, E.; Geng, X.; Xuan Nguyen, N.T.; Tardieu, D.; Bailly, J.D.; Auvergne, A.; Guerre, P. Comparative effects of fumonisins on sphingolipid metabolism and toxicity in ducks and turkeys. Avian Dis. 2012, 56, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Voss, K.A.; Smith, G.W.; Haschek, W.M. Fumonisins: Toxicokinetics, mechanism of action and toxicity. Anim. Feed Sci. Technol. 2007, 137, 299–325. [Google Scholar] [CrossRef]
- Soriano, J.M.; González, L.; Catalá, A.I. Mechanism of action of sphingolipids and their metabolites in the toxicity of fumonisin B1. Prog. Lipid Res. 2005, 44, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Yoo, H.-S.; Norred, W.P.; Showker, J.; Riley, R.T. Elevated sphingoid bases and complex sphingolipid depletion as contributing factors in fumonisin-induced cytotoxicity. Toxicol. Appl. Pharmacol. 1996, 138, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Schmelz, E.M.; Dombrink-Kurtzman, M.A.; Roberts, P.C.; Kozutsumi, Y.; Kawasaki, T.; Merrill, A.H., Jr. Induction of apoptosis by fumonisin B1 in HT29 cells is mediated by the accumulation of endogenous free sphingoid bases. Toxicol. Appl. Pharmacol. 1998, 148, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Tardieu, D.; Tran, S.T.; Auvergne, A.; Babilé, R.; Benard, G.; Bailly, J.D.; Guerre, P. Effects of fumonisins on liver and kidney sphinganine and the sphinganine to sphingosine ratio during chronic exposure in ducks. Chem. Biol. Interact. 2006, 160, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Mahfoud, R.; Maresca, M.; Santelli, M.; Pfohl-Leszkowicz, A.; Puigserver, A.; Fantini, J. PH-dependent interaction of fumonisin B1 with cholesterol: Physicochemical and molecular modeling studies at the air-water interface. J. Agric. Food Chem. 2001, 50, 327–331. [Google Scholar] [CrossRef]
- Bouhet, S.; Oswald, I.P. The intestine as a possible target for fumonisin toxicity. Mol. Nutr. Food Res. 2007, 51, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Minervini, F.; Garbetta, A.; D’Antuono, I.; Cardinali, A.; Martino, N.; Debellis, L.; Visconti, A. Toxic mechanisms induced by fumonisin B1 mycotoxin on human intestinal cell line. Arch. Environ. Contam. Toxicol. 2014, 67, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, L.N.; Colborn, T.; Hayes, T.B.; Heindel, J.J.; Jacobs, D.R.; Lee, D.H.; Shioda, T.; Soto, A.M.; vom Saal, F.S.; Welshons, W.V.; et al. Hormones and endocrine-disrupting chemicals: Low-dose effects and nonmonotonic dose responses. Endocr. Rev. 2012, 33, 378–455. [Google Scholar] [CrossRef] [PubMed]
- Diaz, G.J.; Calabrese, E.; Blain, R. Aflatoxicosis in chickens (Gallus gallus): An example of hormesis? Poult. Sci. 2008, 87, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Applegate, T.J.; Schatzmayr, G.; Pricket, K.; Troche, C.; Jiang, Z. Effect of aflatoxin culture on intestinal function and nutrient loss in laying hens. Poult. Sci. 2009, 88, 1235–1241. [Google Scholar] [CrossRef] [PubMed]
- Riley, R.T.; Enongene, E.; Voss, K.A.; Norred, W.P.; Meredith, F.I.; Sharma, R.P.; Spitsbergen, J.; Williams, D.E.; Carlson, D.B.; Merrill, A.H. Sphingolipid perturbations as mechanisms for fumonisin carcinogenesis. Environ. Health Perspect. 2001, 109, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Riley, R.T.; Voss, K.A. Differential sensitivity of rat kidney and liver to fumonisin toxicity: Organ-specific differences in toxin accumulation and sphingoid base metabolism. Toxicol. Sci. 2006, 92, 335–345. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Suzuki, H.; Sharma, N.; Sharma, R.P. Ceramide synthase inhibition by fumonisin B1 treatment activates sphingolipid-metabolizing systems in mouse liver. Toxicol. Sci. 2006, 94, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Kunisawa, J.; Kiyono, H. Immunological function of sphingosine 1-phosphate in the intestine. Nutrients 2012, 4, 154–166. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Nagahashi, M.; Kim, E.Y.; Harikumar, K.B.; Yamada, A.; Huang, W.-C.; Hait, N.C.; Allegood, J.C.; Price, M.M.; Avni, D.; et al. Sphingosine-1-phosphate links persistent STAT3 activation, chronic intestinal inflammation, and development of colitis-associated cancer. Cancer Cell 2013, 23, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Oswald, I.P.; Desautels, C.; Laffitte, J.; Fournout, S.; Peres, S.Y.; Odin, M.; Le Bars, P.; Le Bars, J.; Fairbrother, J.M. Mycotoxin fumonisin B1 increases intestinal colonization by pathogenic escherichia coli in pigs. Appl. Environ. Microbiol. 2003, 69, 5870–5874. [Google Scholar] [CrossRef] [PubMed]
- Maresca, M.; Fantini, J. Some food-associated mycotoxins as potential risk factors in humans predisposed to chronic intestinal inflammatory diseases. Toxicon 2010, 56, 282–294. [Google Scholar] [CrossRef] [PubMed]
- Cano, P.M.; Seeboth, J.; Meurens, F.; Cognie, J.; Abrami, R.; Oswald, I.P.; Guzylack-Piriou, L. Deoxynivalenol as a new factor in the persistence of intestinal inflammatory diseases: An emerging hypothesis through possible modulation of th17-mediated response. PLoS One 2013, 8, e53647. [Google Scholar] [CrossRef] [PubMed]
- Bracarense, A.-P.F.L.; Lucioli, J.; Grenier, B.; Drociunas Pacheco, G.; Moll, W.-D.; Schatzmayr, G.; Oswald, I.P. Chronic ingestion of deoxynivalenol and fumonisin, alone or in interaction, induces morphological and immunological changes in the intestine of piglets. Br. J. Nutr. 2012, 107, 1776–1786. [Google Scholar] [CrossRef] [PubMed]
- Monteleone, I.; Pallone, F.; Monteleone, G. Th17-related cytokines: New players in the control of chronic intestinal inflammation. BMC Med. 2011, 9, 122. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, R.; Nishimoto, S.; Muto, G.; Sekiya, T.; Tamiya, T.; Kimura, A.; Morita, R.; Asakawa, M.; Chinen, T.; Yoshimura, A. Socs1 is essential for regulatory t cell functions by preventing loss of foxp3 expression as well as ifn-γ and il-17a production. J. Exp. Med. 2011, 208, 2055–2067. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, A.; Suzuki, M.; Sakaguchi, R.; Hanada, T.; Yasukawa, H. Socs, inflammation and autoimmunity. Front. Immunol. 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Bar-Shira, E.; Sklan, D.; Friedman, A. Establishment of immune competence in the avian galt during the immediate post-hatch period. Dev. Comp. Immunol. 2003, 27, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Friedman, A.; Bar-shira, E.; Sklan, D. Ontogeny of gut associated immune competence in the chick. World's Poult. Sci. J. 2003, 59, 209–219. [Google Scholar] [CrossRef]
- Dombrink-Kurtzman, M.; Javed, T.; Bennett, G.; Richard, J.; Cote, L.M.; Buck, W. Lymphocyte cytotoxicity and erythrocytic abnormalities induced in broiler chicks by fumonisins B1 and B2 and moniliformin fromfusarium proliferatum. Mycopathologia 1993, 124, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Dombrink-Kurtzman, M. Fumonisin and beauvericin induce apoptosis in turkey peripheral blood lymphocytes. Mycopathologia 2003, 156, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Shanmugasundaram, R.; Selvaraj, R.K. Regulatory t cell properties of chicken CD4+CD25+ cells. J. Immunol. 2011, 186, 1997–2002. [Google Scholar] [CrossRef] [PubMed]
- Shanmugasundaram, R.; Selvaraj, R.K. CD4+CD25+ regulatory T cell ontogeny and preferential migration to the cecal tonsils in chickens. PLoS One 2012, 7, e33970. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, A.E.; Plattner, R.D.; Shackelford, D.D.; Leslie, J.F.; Nelson, P.E. Heritability of fumonisin B1 production in gibberella fujikuroi mating population a. Appl. Environ. Microbiol. 1992, 58, 2799–2805. [Google Scholar] [PubMed]
- Streit, E.; Schwab, C.; Sulyok, M.; Naehrer, K.; Krska, R.; Schatzmayr, G. Multi-mycotoxin screening reveals the occurrence of 139 different secondary metabolites in feed and feed ingredients. Toxins 2013, 5, 504–523. [Google Scholar] [CrossRef] [PubMed]
- Peirson, S.N.; Butler, J.N.; Foster, R.G. Experimental validation of novel and conventional approaches to quantitative real-time pcr data analysis. Nucleic Acids Res. 2003, 31, e73. [Google Scholar] [CrossRef] [PubMed]
- Adedokun, S.A.; Ajuwon, K.M.; Romero, L.F.; Adeola, O. Ileal endogenous amino acid losses: Response of broiler chickens to fiber and mild coccidial vaccine challenge. Poult. Sci. 2012, 91, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.H.; Lillehoj, H.S.; Lillehoj, E.P.; Lee, S.H. Changes in immune-related gene expression and intestinal lymphocyte subpopulations following eimeria maxima infection of chickens. Vet. Immunol. Immunopathol. 2006, 114, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Antonissen, G.; Van Immerseel, F.; Pasmans, F.; Ducatelle, R.; Haesebrouck, F.; Timbermont, L.; Verlinden, M.; Janssens, G.P.J.; Eeckhaut, V.; Eeckhout, M.; et al. The mycotoxin deoxynivalenol predisposes for the development of clostridium perfringens-induced necrotic enteritis in broiler chickens. PLoS One 2014, 9, e108775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grenier, B.; Schwartz-Zimmermann, H.E.; Caha, S.; Moll, W.D.; Schatzmayr, G.; Applegate, T.J. Dose-Dependent Effects on Sphingoid Bases and Cytokines in Chickens Fed Diets Prepared with Fusarium Verticillioides Culture Material Containing Fumonisins. Toxins 2015, 7, 1253-1272. https://doi.org/10.3390/toxins7041253
Grenier B, Schwartz-Zimmermann HE, Caha S, Moll WD, Schatzmayr G, Applegate TJ. Dose-Dependent Effects on Sphingoid Bases and Cytokines in Chickens Fed Diets Prepared with Fusarium Verticillioides Culture Material Containing Fumonisins. Toxins. 2015; 7(4):1253-1272. https://doi.org/10.3390/toxins7041253
Chicago/Turabian StyleGrenier, Bertrand, Heidi E. Schwartz-Zimmermann, Sylvia Caha, Wulf Dieter Moll, Gerd Schatzmayr, and Todd J. Applegate. 2015. "Dose-Dependent Effects on Sphingoid Bases and Cytokines in Chickens Fed Diets Prepared with Fusarium Verticillioides Culture Material Containing Fumonisins" Toxins 7, no. 4: 1253-1272. https://doi.org/10.3390/toxins7041253
APA StyleGrenier, B., Schwartz-Zimmermann, H. E., Caha, S., Moll, W. D., Schatzmayr, G., & Applegate, T. J. (2015). Dose-Dependent Effects on Sphingoid Bases and Cytokines in Chickens Fed Diets Prepared with Fusarium Verticillioides Culture Material Containing Fumonisins. Toxins, 7(4), 1253-1272. https://doi.org/10.3390/toxins7041253