
Presence of Multiple Mycotoxins and Other Fungal Metabolites in Native Grasses from a Wetland Ecosystem in Argentina Intended for Grazing Cattle

 , ,
, , 

Abstract
:
1. Introduction
2. Results
2.1. Performance of the Applied Analytical Method

{kind=link}
{kind=link}
| Analyte | Apparent Recovery (%) | LOD a (µg/kg) |
|---|---|---|
| 3-Nitropropionic acid | 63.3 ± 9.5 | 0.40 |
| Agroclavine | 59.5 ± 5.1 | 0.08 |
| Altenuene | 157.8 ± 40.0 | 2.00 |
| Alternariol | 100.0 ± 9.5 | 0.50 |
| Alternariol methyl ether | 96.6 ± 8.8 | 0.05 |
| Altertoxin-I | 77.9 ± 6.5 | 0.80 |
| Aspinolid B | 75.5 ± 13.2 | 0.80 |
| Asterric acid | 140.0 ± 9.6 | 3.00 |
| Aurofusarin | 75.3 ± 4.7 | 4.00 |
| Averantin | 76.5 ± 0.9 | 0.04 |
| Averufanin b | n.d. d | - |
| Averufin | 54.50 ± 2.50 | 0.03 |
| Beauvericin | 41.8 ± 6.0 | 0.04 |
| Brefeldin A | 66.5 ± 6.2 | 2.00 |
| Brevianamid F | 76.2 ± 2.1 | 0.40 |
| Chanoclavin | 73.5 ± 31.2 | 0.05 |
| Chrysophanol | 114.8 ± 11.8 | 1.50 |
| Clonostachydiol | 80.0 ± 21.0 | 0.80 |
| Curvularin | 127.6 ± 15.8 | 0.40 |
| Cytochalasin B | 66.8 ± 7.1 | 1.50 |
| Cytochalasin C | 66.8 ± 6.1 | 0.40 |
| Cytochalasin D | 62.5 ± 4.1 | 0.20 |
| Cytochalasin H | 57.2 ± 5.6 | 15.00 |
| Cytochalasin J | 59.2 ± 7.9 | 2.00 |
| Dechlorogriseofulvin | 73.7 ± 9.1 | 1.50 |
| Diacetoxyscirpenol | 61.0 ± 9.8 | 0.25 |
| Dihydrogriseofulvin | 69.7 ± 8.8 | 1.00 |
| Emodin | 131.3 ± 28.2 | 0.08 |
| Enniatin B | 77.7 ± 42.9 | 0.01 |
| Equisetin | 215.0 ± 38.7 | 0.08 |
| Festuclavine | 73.1 ± 17.3 | 0.03 |
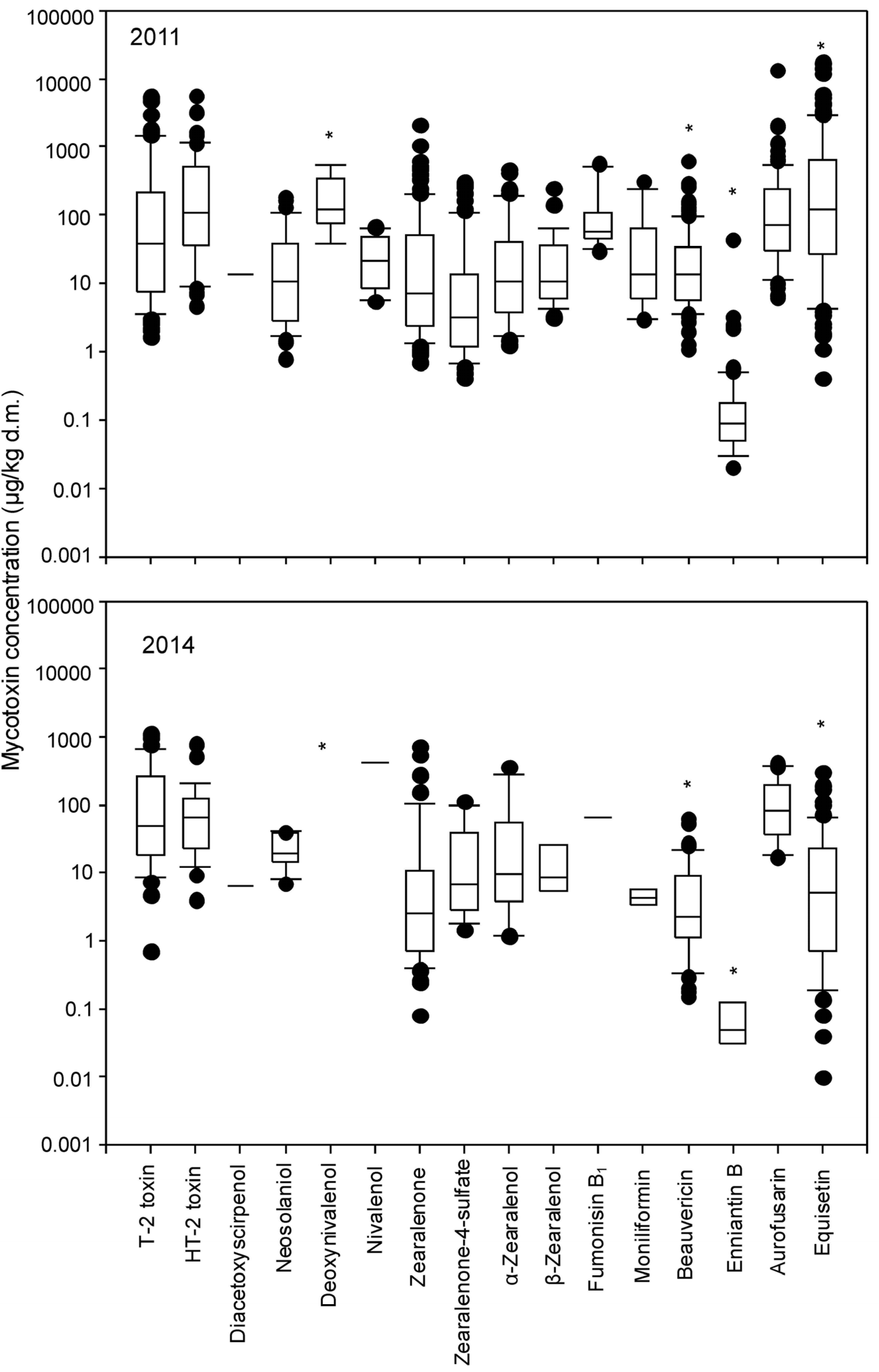
| Fumonisin B1 | 59.8 ± 8.1 | 3.00 |
| Griseofulvin | 69.7 ± 4.8 | 0.80 |
| HT-2 toxin | 68.3 ± 1.0 | 4.00 |
| Kojic acid | 83.4 ± 12.2 | 20.00 |
| Macrosporin | 125.8 ± 5.2 | 0.20 |
| Moniliformin | 113.7 ± 6.7 | 1.00 |
| Monocerin | 73.8 ± 3.0 | 0.40 |
| Neosolaniol | 79.2 ± 3.5 | 0.50 |
| Nidurufin b | n.d. | - |
| Nivalenol | 41.3 ± 12.0 | 0.80 |
| Norsolorinic acid | 47.0 ± 15.1 | 0.40 |
| Penicillide | 110.9 ± 16.3 | 1.50 |
| Physcion | 114.3 ± 16.6 | 8.00 |
| Pseurotin A | 73.7 ± 48.7 | 0.50 |
| Radicicol | 117.0 ± 12.0 | 0.40 |
| Secalonic acid D | 79.1 ± 8.6 | 4.00 |
| Skyrin | 76.1 ± 4.6 | 0.20 |
| Sterigmatocystin | 69.6 ± 4.0 | 0.30 |
| Sulochrin | 89.7 ± 14.9 | 1.50 |
| T-2 toxin | 64.1 ± 3.6 | 0.80 |
| Tentoxin | 56.2 ± 11.9 | 0.20 |
| Tenuazonic acid | 407.4 ± 108.9 | 3.00 |
| Tryptophol | 79.7 ± 6.4 | 15.0 |
| Versicolorin A | 103.7 ± 3.6 | 0.40 |
| Versicolorin C c | n.d. | - |
| Zearalenone | 124.6 ± 10.9 | 0.30 |
| Zearalenone-4-sulfate | 143.1 ± 16.1 | 0.40 |
| α-zearalenol | 120.6 ± 17.2 | 0.80 |
| β-zearalenol | 109.3 ± 15.6 | 0.80 |
2.2. Occurrence of Fungal Metabolites in Natural Grass Samples

| Analyte | N° of Positives | Concentration of Positive Samples from 2011 (µg/kg d.m.) | N° of Positives | Concentration in Positive Samples from 2014 (µg/kg d.m.) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Median | 75 Percentile | 90 Percentile | Max | Median | 75 Percentile | 90 Percentile | Max | |||
| T-2 toxin | 67/106 | 37.8 | 218 | 1375 | 5438 | 39/69 | 47.2 | 243 | 559 | 1095 |
| HT-2 toxin | 52/106 | 110 | 474 | 1149 | 5651 | 29/60 | 66.5 | 117 | 205 | 783 |
| Diacetoxyscirpenol | 2/106 | 13.6 | 21.5 | 21.5 | 21.5 | 2/69 | 6.49 | 10.5 | 10.5 | 10.5 |
| Neosolaniol | 36/106 | 10.4 | 39.0 | 94.5 | 187 | 11/69 | 19.4 | 38.1 | 39.6 | 39.9 |
| Nivalenol | 15/106 | 21.0 | 44.3 | 63.6 | 66.9 | 2/69 | 422 | 832 | 832 | 831 |
| Zearalenone | 95/106 | 7.20 | 50.3 | 203 | 2120 | 56/69 | 2.50 | 10.0 | 80.2 | 712 |
| Zearalenone-4-sulfate | 67/106 | 3.10 | 13.0 | 99.4 | 303 | 17/69 | 6.81 | 35.7 | 87.4 | 111 |
| α-zearalenol | 53/106 | 10.7 | 39.4 | 181 | 464 | 12/69 | 9.37 | 45.1 | 154 | 360 |
| β-zearalenol | 28/106 | 10.9 | 34.9 | 53.9 | 245 | 7/69 | 8.35 | 24.2 | 47.5 | 52.8 |
| Fumonisin B1 | 11/106 | 57.2 | 97.0 | 442 | 566 | 2/69 | 64.0 | 98.7 | 98.7 | 98.7 |
| Moniliformin | 12/106 | 13.5 | 56.2 | 152 | 313 | 3/69 | 4.38 | 5.3 | 5.64 | 5.64 |
| Beauvericin | 106/106 | 13.5 | 33.2 | 95.1 | 624 | 43/69 | 2.23 | 9.1 | 18.5 | 63.4 |
| Enniatin B | 74/106 | 0.09 | 0.17 | 0.47 | 43.2 | 4/69 | 0.05 | 0.10 | 0.14 | 0.14 |
| Aurofusarin | 96/106 | 71.4 | 231 | 508 | 13238 | 25/69 | 80.7 | 195 | 365 | 409 |
| Equisetin | 106/106 | 118 | 624 | 2841 | 17264 | 63/69 | 2.21 | 12.8 | 54.8 | 297 |
| Sterigmatocystin | 96/106 | 4.15 | 16.5 | 42.6 | 733 | 41/69 | 6.78 | 15.1 | 53.8 | 147 |
| Averantin | 85/106 | 1.60 | 5.07 | 19.4 | 499 | 65/69 | 1.16 | 2.60 | 25.3 | 319 |
| Averufanin | 51/106 | 0.70 | 1.55 | 4.44 | 17.5 | 21/69 | 0.20 | 0.54 | 3.73 | 4.65 |
| Averufin | 67/106 | 3.80 | 8.57 | 20.4 | 173 | 67/69 | 2.78 | 8.00 | 49.3 | 401 |
| Versicolorin A | 20/106 | 0.65 | 1.35 | 27.4 | 46.5 | 38/69 | 3.08 | 11.9 | 103 | 719 |
| Versicolorin C | 33/106 | 1.50 | 2.42 | 4.22 | 25.3 | 47/69 | 3.36 | 10.1 | 43.9 | 209 |
| Nidurufin | 41/106 | 0.60 | 41.2 | 91.1 | 228 | 30/69 | 0.87 | 3.58 | 10.1 | 28.3 |
| Norsolorinic acid | 59/106 | 3.60 | 23.4 | 93.6 | 257 | 44/69 | 3.55 | 6.76 | 23.7 | 63.1 |
| Kojic acid | 39/106 | 206 | 329 | 404 | 522 | 66/69 | 103 | 127 | 139 | 187 |
| 3-nitropropionic acid | 32/106 | 2.70 | 5.70 | 9.94 | 28.8 | 37/69 | 3.77 | 1.91 | 28.0 | 102 |
| Aspinolid B | 40/106 | 39.2 | 82.6 | 186 | 602 | 27/69 | 10.0 | 25.0 | 43.7 | 423 |
| Asterric acid | 44/106 | 33.6 | 58.8 | 141 | 346 | 26/69 | 22.9 | 58.3 | 140.2 | 223 |
| Sulochrin | 42/106 | 23.7 | 62.6 | 90.1 | 198 | 30/69 | 18.3 | 52.7 | 283 | 172 |
| Pseurotin A | 31/106 | 4.70 | 8.37 | 13.4 | 15.2 | 10/69 | 7.88 | 23.4 | 148 | 29.5 |
| Agroclavine | 2/106 | 11.2 | 14.0 | 14.0 | 14.0 | 1/69 | ||||
| Chanoclavin | 69/106 | 2.60 | 49.1 | 180.6 | 2259 | 31/69 | 2.44 | 45.1 | 439 | 815 |
| Festuclavine | 5/106 | 32.1 | 93.6 | 98.4 | 98.4 | 5/69 | 7.63 | 75.6 | 215 | 215 |
| Secalonic acid | 28/106 | 72.9 | 116 | 229 | 1431 | 7/69 | 36.5 | 202 | 230 | 236 |
| Griseofulvin | 30/106 | 18.9 | 150 | 594 | 5880 | 4/69 | 23.4 | 75.0 | 123 | 123 |
| Dechlorogriseofulvin | 18/106 | 16.2 | 124 | 270 | 3904 | 2/69 | 20.0 | 32.5 | 32.5 | 32.5 |
| Dihydrogriseofulvin | 20/106 | 17.4 | 119 | 333 | 5720 | 3/69 | 14.2 | 55.6 | 69.4 | 69.4 |
| Curvularin | 73/106 | 9.30 | 39.7 | 358 | 5362 | 14/69 | 5.10 | 8.62 | 121 | 934 |
| Brefeldin A | 2/106 | 1648 | 2988 | 2988 | 2988 | 2/69 | 745 | 874 | 874 | 875 |
| Penicillide | 7/106 | 5.70 | 10.1 | 14.1 | 14.9 | 1/69 | 15.4 | |||
| Tenuazonic acid | 28/106 | 31.4 | 59.4 | 141 | 222 | 27/69 | 2.43 | 4.67 | 11.3 | 178 |
| Alternariol | 105/106 | 65.3 | 160. | 349 | 1036 | 61/69 | 17.4 | 58.0 | 136 | 1021 |
| Alternariol methyl ether | 67/106 | 12.6 | 33.3 | 69.2 | 377 | 67/69 | 10.5 | 20.2 | 39.2 | 160 |
| Tentoxin | 90/106 | 1.60 | 4.30 | 12.8 | 324 | 21/69 | 3.76 | 21.3 | 136 | 252 |
| Altenuene | 23/106 | 10.3 | 15.5 | 19.8 | 28.4 | 2/69 | 14.3 | 18.7 | 18.7 | 18.7 |
| Altertoxin-I | 75/106 | 5.00 | 9.40 | 15.6 | 383 | 5/69 | 1.43 | 1.86 | 2.72 | 2.72 |
| Macrosporin | 60/106 | 2.85 | 8.90 | 21.8 | 50.4 | 17/69 | 1.95 | 4.53 | 19.2 | 76.2 |
| Clonostachydiol | 13/106 | 11.5 | 21.4 | 53.0 | 121 | 3/69 | 9.97 | 71.9 | 92.6 | 92.6 |
| Cytochalasin B | 10/106 | 90.0 | 136 | 925 | 1616 | 6/69 | 32.6 | 42.6 | 91.6 | 97.0 |
| Cytochalasin C | 14/106 | 27.9 | 64.6 | 264 | 412 | 3/69 | 33.6 | 38.1 | 39.5 | 39.5 |
| Cytochalasin D | 10/106 | 24.6 | 67.2 | 87.6 | 106 | 6/69 | 9.28 | 10.3 | 24.1 | 25.6 |
| Cytochalasin H | 9/106 | 368 | 1152 | 1802 | 2003 | 4/69 | 163 | 638 | 1064 | 1064 |
| Cytochalasin J | 20/106 | 40.8 | 133 | 215 | 434 | 4/69 | 122 | 436 | 677 | 677 |
| Monocerin | 106/106 | 66.7 | 215 | 1916 | 6745 | 65/69 | 20.7 | 46.9 | 642 | 7289 |
| Brevianamid F | 33/106 | 8.00 | 11.2 | 17.1 | 34.3 | 18/69 | 32.3 | 36.1 | 49.0 | 62.5 |
| Tryptophol | 75/106 | 73.1 | 113 | 217 | 466 | 67/69 | 67.3 | 226.8 | 1070 | 2513 |
| Emodin | 106/106 | 177 | 471 | 1551 | 3725 | 67/69 | 62.1 | 95.4 | 282 | 5401 |
| Chrysophanol | 104/106 | 41.1 | 77.8 | 142 | 15952 | 63/69 | 15.5 | 24.8 | 47.6 | 120 |
| Physcion | 74/106 | 69.9 | 166 | 391 | 17752 | 7/69 | 31.7 | 35.4 | 45.9 | 48.4 |
| Skyrin | 105/106 | 5.90 | 14.2 | 38.3 | 241 | 55/69 | 1.93 | 2.67 | 5.42 | 13.7 |
| Radicicol | 50/106 | 33.2 | 95.0 | 182 | 380 | 20/69 | 17.5 | 56.5 | 75.0 | 210 |
3. Discussion
4. Experimental Section
4.1. Sampling
4.2. Mycotoxin Analysis
4.2.1. Chemicals and Reagents
4.2.2. Extraction and Estimation of Apparent Recoveries
4.2.3. LC-MS/MS Parameters
4.3. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Arelovich, H.M.; Bravo, R.D.; Martinez, M.F. Development, characteristic, and trends for beef cattle production in Argentina. Anim. Front. 2011, 1, 37–45. [Google Scholar] [CrossRef]
- Garcia, P.T.; Pensela, N.A.; Sancho, A.M.; Latimori, N.J.; Kloster, A.M.; Amigone, M.A.; Casal, J.J. Beef lipids in relation to animal breed and nutrition in Argentina. Meat Sci. 2008, 79, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Rotolo, G.C.; Rydberg, T.; Lieblein, G.; Francis, C. Emergy evaluation of grazing cattle in Argentina’s Pampas. Agric. Ecosyst. Environ. 2007, 119, 383–395. [Google Scholar] [CrossRef]
- Mostrom, M.; Jacobsen, B. Ruminant mycotoxicosis. Vet. Clin. N. Am. Food Anim. Pract. 2011, 27, 315–344. [Google Scholar] [CrossRef] [PubMed]
- Fink-Gremmels, J. Mycotoxins in cattle feeds and carry-over to dairy milk: A review. Food Addit. Contam. Part A. 2008, 25, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Zain, M. Impact of mycotoxins on humans and animals. J. Saudi Chem. Soc. 2011, 15, 129–144. [Google Scholar] [CrossRef]
- Mathews, K.H.; Vanderveer, M. Beef Production, Markets, and Trade in Argentina and Uruguay: An Overview. Economic Research Service, USDA. Available online: http://www.ers.usda.gov/publications/ldpm-livestock,-dairy,-and-poultry-outlook/ldpm-15901.aspx (accessed on 20 February 2015).
- Alonso, V.A.; Pereyra, C.M.; Keller, L.A.M.; Dalcero, A.M.; Rosa, C.A.R.; Chiacchiera, S.M.; Cavaglieri, L.R. Fungi and mycotoxins in silage: An overview (Review). J. Appl. Microbiol. 2013, 115, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.G.; Hewitt, S.A.; McEvoy, J.D.; Currie, J.W.; Cannavan, A.; Blanchflower, W.J.; Elliot, C.T. Zeranol is formed from Fusarium sp. toxins in cattle in vivo. Food Adit. Contam. 1998, 15, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.F.; Morris, C.A. Review of zearalenone studies with sheep in New Zealand. Proc. N. Z. Soc. Anim. Prod. 2006, 66, 306–310. [Google Scholar]
- Ramsar Convention Secretariat. The Ramsar Convention Manual: A Guide to the Convention on Wetlands (Ramsar, Iran, 1971), 6th ed.; Ramsar Convention Secretariat: Gland, Switzerland, 2013. [Google Scholar]
- Malachova, A.; Sulyok, M.; Beltrán, E.; Berthiller, F.; Krska, R. Optimization and validation of a quantitative liquid chromatography — Tandem mass spectrometric method covering 295 bacterial and fungal metabolites including all relevant mycotoxins in four model food matrices. J. Chromatogr. A 2014, 1362, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Sulyok, M.; Berthiller, F.; Kraska, R.; Shhuhmacher, R. Development and validation of a liquid chromatography/tandem mass spectrometric method for the determination of 39 mycotoxins in wheat and maize. Rapid Commun. Mass Spectrom. 2006, 20, 2649–2659. [Google Scholar] [CrossRef] [PubMed]
- Hult, K.; Telling, A.; Gatenbeck, S. Degradation of ochratoxin A in ruminants. Appl. Environ. Microbiol. 1976, 32, 443–444. [Google Scholar] [PubMed]
- Allcroft, R.; Robert, B.A.; Lloyd, M.K. Excretion of aflatoxin in a lactating cow. Food Cosmet. Toxicol. 1968, 6, 619–625. [Google Scholar] [CrossRef]
- Engel, V.G.; Hagemeister, H. Untersuchungenueber den verblieb von aflatoxin B1 im Verdaaundtarkt von Kuehen. Milchwissenschaft 1978, 33, 21–23. [Google Scholar]
- Kiessling, K.H.; Pettersson, H.; Sandholm, K. Metabolism of aflatoxin, ochratoxin, zearalenone, and three trichothecenes by intact rumen fluid, rumen protozoa, and rumen bacteria. Appl. Environ. Microbiol. 1984, 47, 1070–1073. [Google Scholar] [PubMed]
- Seeling, K.; Lebzien, P.; Danicke, S.; Spilke, J.; Sudekum, K.H.; Flachowsky, G. Effects of level of feed intake and Fusarium toxin contaminated wheat on rumen fermentation as well as on blood and milk parameters in cows. J. Anim. Physiol. Anim. Nutr. 2006, 90, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Kleinova, M.; Zöllner, P.; Kahlbacher, H.; Hochsteiner, W.; Lindner, W. Metabolic profiles of the mycotoxin zearalenone and of the growth promoter zeranol in urine, liver, and muscle of heifers. J. Agric. Food Chem. 2002, 50, 4769–4776. [Google Scholar] [CrossRef] [PubMed]
- Berthiller, F.; Werner, U.; Sulyok, M.; Krska, R.; Hauser, M.T.; Schuhmacher, R. Liquid chromatography coupled to tandem mass spectrometry (LC-MS/MS) determination of phase II metabolites of the mycotoxin zearalenone in the model plant Arabidopsis thaliana. Food Addit. Contam. 2006, 23, 1194–1200. [Google Scholar] [CrossRef] [PubMed]
- Plasencia, J.; Mirocha, C.J. Isolation and characterization of zearalenone sulfate produced by Fusarium spp. Appl. Environ. Microbiol. 1991, 57, 146–150. [Google Scholar] [PubMed]
- Zhou, T.; He, J.; Gong, J. Microbial transformation of trichothecene mycotoxins. World Mycotoxin J. 2008, 1, 23–30. [Google Scholar] [CrossRef]
- Nichea, M.J.; Cendoya, E.; Zachetti, V.G.L.; Chiacchiera, S.M.; Sulyok, M.; Krska, R.; Torres, A.M.; Chulze, S.N.; Ramirez, M.L. Mycotoxin profile of Fusarium armeniacum isolated from natural grasses intended for cattle feed. World Mycotoxin J. 2015, 8, 451–457. [Google Scholar] [CrossRef]
- Osweiler, G.D. Mycotoxins-contemporary issues of food animal health and productivity. Vet. Clin. N. Am. Food Anim. Pract. 2000, 16, 511–530. [Google Scholar]
- Eckard, S.; Wettstein, F.E.; Forrer, H.R.Y.; Vogelsang, S. Incidence of Fusarium species and mycotoxins in silage maize. Toxins 2011, 3, 949–967. [Google Scholar] [CrossRef] [PubMed]
- Schollenberger, M.; Müller, H.M.; Rüfle, M.; Suchy, S.; Plank, S.; Drochner, W. Natural occurrence of 16 Fusarium toxins in grains and feedstuffs of plant origin from Germany. Mycopathologia 2006, 161, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Streit, E.; Schatzmayr, G.; Tassis, P.; Tzika, E.; Marin, D.; Taranu, I.; Tabuc, C.; Nicolau, A.; Aprodu, I.; Puel, O.; et al. Current situation of mycotoxin contamination and co-occurrence in animal feed-focus on Europe. Toxins 2012, 4, 788–809. [Google Scholar] [CrossRef] [PubMed]
- Sulyok, M.; Krska, R.; Schuhmacher, R. A liquid chromatography/tandem mass spectrometric multi-mycotoxin method for the quantification of 87 analytes and its application to semiquantitative screening of moldy food samples. Anal. Bioanal. Chem. 2007, 389, 1505–1523. [Google Scholar] [CrossRef] [PubMed]
- Van Asselt, E.D.; Azambuja, W.; Moretti, A.; Kastelein, P.; De Rijk, T.C.; Stratakou, I.; van der Fels-Klerx, H.J. A Dutch field study on fungal infection and mycotoxin concentrations in maize. Food Addit. Contam. 2012, 29, 1556–1565. [Google Scholar] [CrossRef] [PubMed]
- Hadley, G.; Wolf, C.; Harsh, S. Dairy cattle culling patterns, explanations, and implications. J. Dairy Sci. 2006, 89, 2286–2296. [Google Scholar] [CrossRef]
- Wenz, J.; Jensen, S.; Lombard, J.; Wagner, B.; Dinsmore, R. Herd management practices and their association with bulk tank somatic cell count on United States dairy operations. J. Dairy Sci. 2007, 90, 3652–3659. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xu, L. Beauvericin, a bioactive compound produced by fungi: A short review. Molecules 2012, 17, 2367–2377. [Google Scholar] [CrossRef] [PubMed]
- Sulyok, M.; Krska, R.; Schuhmacher, R. Application of an LC-MS/MS based multi-mycotoxin method for the semi-quantitative determination of mycotoxins occurring in different types of food infected by moulds. Food Chem. 2010, 119, 408–416. [Google Scholar] [CrossRef]
- Warth, B.; Parich, A.; Atehnkeng, J.; Bandyopadhyay, R.; Schuhmacher, R.; Sulyok, M.; Krska, R. Quantitation of mycotoxins in food and feed from Burkina Faso and Mozambique using a modern LC-MS/MS multitoxin method. J. Agric. Food Chem. 2012, 60, 9352–9363. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority. Scientific Opinion on the risks for animal and public health related to the presence of Alternaria toxins in feed and food. EFSA J. 2011, 9, 2407–2497. [Google Scholar]
- Shimshoni, J.A.; Cuneah, O.; Sulyok, M.; Krska, R.; Galon, N.; Sharir, B.; Shlosberg, A. Mycotoxins in corn and wheat silage in Israel. Food Addit. Contam. Part A 2013, 30, 1614–1625. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, A. Fusarium Mycotoxins: Chemistry, Genetics and Biology; APS Press: St. Paul, MN, USA, 2006; pp. 79–108. [Google Scholar]
- Mogensen, J.M.; Sørensen, S.M.; Sulyok, M.; van der Westhuizen, L.; Shephard, G.S.; Frisvad, J.C.; Thrane, U.; Krska, R.; Nielsen, K.F. Single kernel analysis of fumonisins and other fungal metabolites in maize from South African subsistence farmers. Food Addit. Contam. Part A 2011, 28, 1724–1734. [Google Scholar] [CrossRef] [PubMed]
- Axford, L.; Simpson, T.; Willis, C. Synthesis and incorporation of the first polyketide synthase free intermediate in monocerin biosynthesis. Angew. Chem. Int. Ed. 2004, 43, 727–730. [Google Scholar] [CrossRef] [PubMed]
- Yu, J. Current understanding on aflatoxin biosynthesis and future perspective in reducing aflatoxin contamination. Toxins 2012, 4, 1024–1057. [Google Scholar] [CrossRef] [PubMed]
- Versilovskis, A.; de Saeger, S. Sterigmatocystin: Occurrence in foodstuffs and analytical methods-an overview. Mol. Nutr. Food Res. 2010, 54, 136–147. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority. Scientific Opinion on the risk for public and animal health related to the presence of sterigmatocystin in food and feed. EFSA J. 2013, 11. [Google Scholar] [CrossRef]
- European Parliament. Directive 2002/32/EC of the European Parliament and of the Council of 7 May 2002 on undesirable substances in animal feed. Off. J. L 2002, 140, 10–21. [Google Scholar]
- O’Brien, M.; Nielsen, K.F.; O’Kiely, P.; Forristal, P.D.; Fuller, H.T.; Frisvad, J.C. Mycotoxins and other secondary metabolites produced in vitro by Penicillium paneum Frisvad and Penicillium roqueforti Thom isolated from baled grass silage in Ireland. J. Agric. Food Chem. 2006, 54, 9268–9276. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.-C.; Chung, J.-G. Anticancer potential of emodin. Biomedicine 2012, 2, 108–116. [Google Scholar] [CrossRef]
- Jizba, J.; Sedmera, P.; Zima, J.; Beran, M.; Blumauerová, M.; Kandybin, N.V.; Samoukina, G.V. Macrotetrolide antibiotics produced by Streptomyces globisporus. Folia Microbiol. 1991, 36, 437–443. [Google Scholar] [CrossRef]
- Riet-Correa, F.; Rivero, R.; Odriozola, E.; Adrien, M.; Medeiros, R.M.T.; Schild, A.L. Mycotoxicoses of ruminants and horses. J. Vet. Diagn. Investig. 2013, 25, 692–708. [Google Scholar] [CrossRef] [PubMed]
- Driehuis, F.; Spanjer, M.C.; Scholten, J.M.; Te Giffel, M.C. Occurrence of mycotoxins in maize, grass and wheat silage for dairy cattle in the Netherlands. Food Addit. Contam. Part B Surveill. 2008, 1, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Reed, K.F.M.; Moore, D.D. A preliminary survey of zearalenone and other mycotoxins in Australian silage and pasture. Anim. Prod. Sci. 2009, 49, 696–703. [Google Scholar] [CrossRef]
- Skládanka, J.; Nedělník, J.; Adam, V.; Doležal, P.; Moravcová, H.; Dohnal, V. Forage as a primary source of mycotoxins in animal diets. Int. J. Environ. Res. Public Health 2011, 8, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Canevari, P.; Blanco, D.E.; Bucher, E.H.; Castro, G.; Davison, I. (Eds.) Los Humedales de la Argentina. Clasificación, situación actual, conservación y legislación; Wetlands International Publication: Buenos Aires, Argentina, 1998; Volume 46, p. 208.
- Vishwanath, V.; Sulyok, M.; Labuda, R.; Bicker, W.; Krska, R. Simultaneous determination of 186 fungal and bacterial metabolites in indoor matrices by liquid chromatography/tandem mass spectrometry. Anal. Bioanal. Chem. 2009, 395, 1355–1372. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Commission Decision of 12 August 2002 implementing Council Directive 96/23/EC concerning the performance of analytical methods and the interpretation of results. Off. J. Eur. Union 2002, 29, 10–32. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nichea, M.J.; Palacios, S.A.; Chiacchiera, S.M.; Sulyok, M.; Krska, R.; Chulze, S.N.; Torres, A.M.; Ramirez, M.L. Presence of Multiple Mycotoxins and Other Fungal Metabolites in Native Grasses from a Wetland Ecosystem in Argentina Intended for Grazing Cattle. Toxins 2015, 7, 3309-3329. https://doi.org/10.3390/toxins7083309
Nichea MJ, Palacios SA, Chiacchiera SM, Sulyok M, Krska R, Chulze SN, Torres AM, Ramirez ML. Presence of Multiple Mycotoxins and Other Fungal Metabolites in Native Grasses from a Wetland Ecosystem in Argentina Intended for Grazing Cattle. Toxins. 2015; 7(8):3309-3329. https://doi.org/10.3390/toxins7083309
Chicago/Turabian StyleNichea, María J., Sofia A. Palacios, Stella M. Chiacchiera, Michael Sulyok, Rudolf Krska, Sofia N. Chulze, Adriana M. Torres, and María L. Ramirez. 2015. "Presence of Multiple Mycotoxins and Other Fungal Metabolites in Native Grasses from a Wetland Ecosystem in Argentina Intended for Grazing Cattle" Toxins 7, no. 8: 3309-3329. https://doi.org/10.3390/toxins7083309
APA StyleNichea, M. J., Palacios, S. A., Chiacchiera, S. M., Sulyok, M., Krska, R., Chulze, S. N., Torres, A. M., & Ramirez, M. L. (2015). Presence of Multiple Mycotoxins and Other Fungal Metabolites in Native Grasses from a Wetland Ecosystem in Argentina Intended for Grazing Cattle. Toxins, 7(8), 3309-3329. https://doi.org/10.3390/toxins7083309