
Aerobic De-Epoxydation of Trichothecene Mycotoxins by a Soil Bacterial Consortium Isolated Using In Situ Soil Enrichment



 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
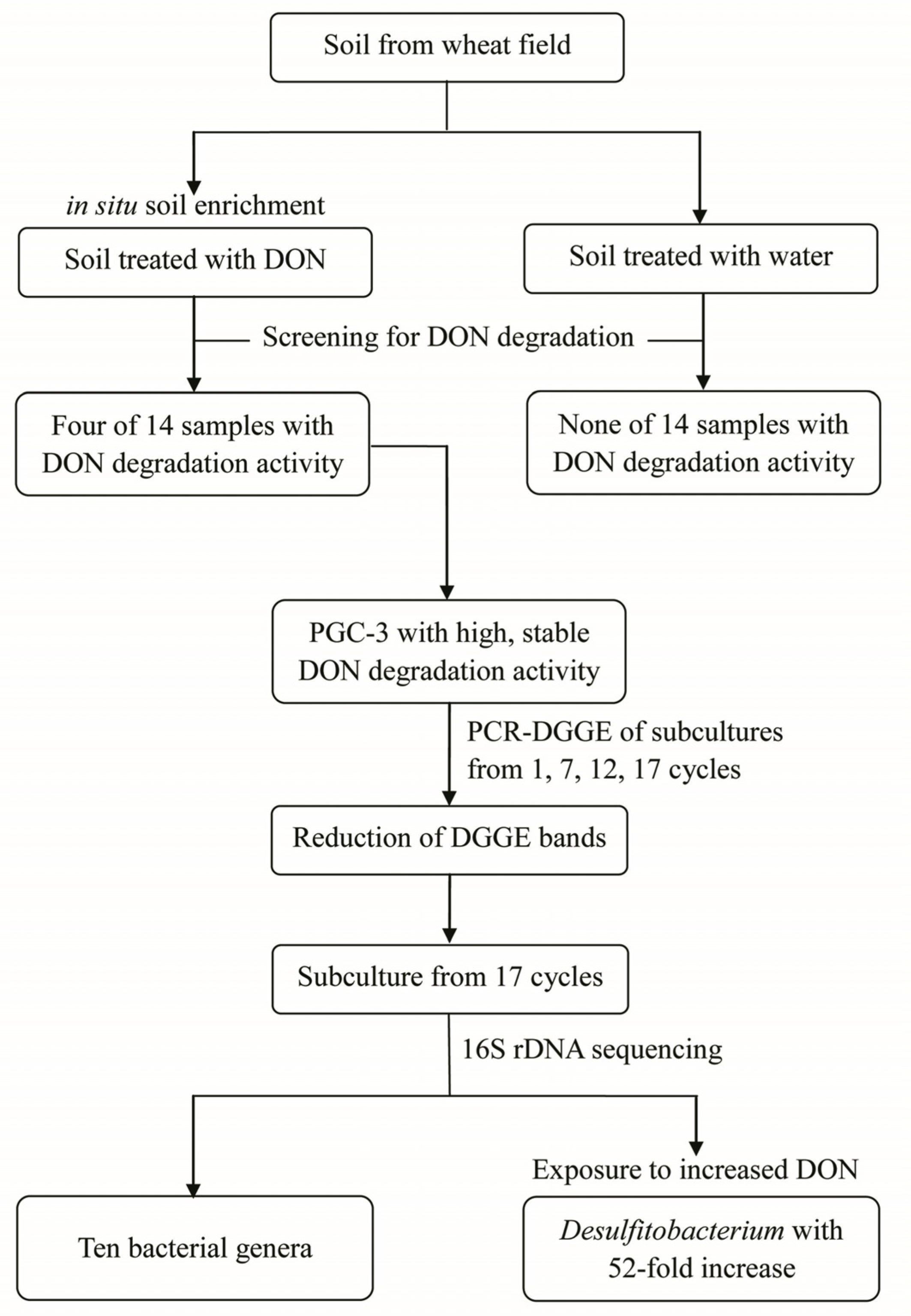
2.1. Isolation of DON-Degrading Culture from Soil
2.2. Analysis of the New DON-Derived Bacterial Metabolite
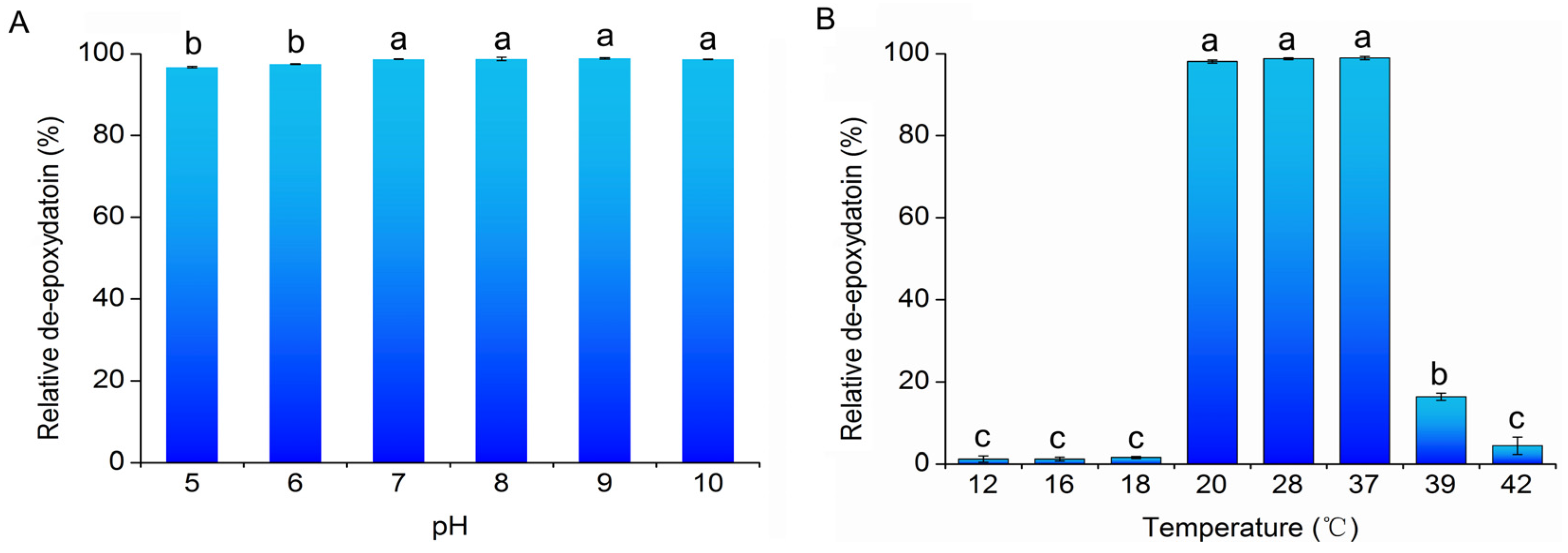
2.3. Effects of Temperature and pH on De-Epoxydation Activity
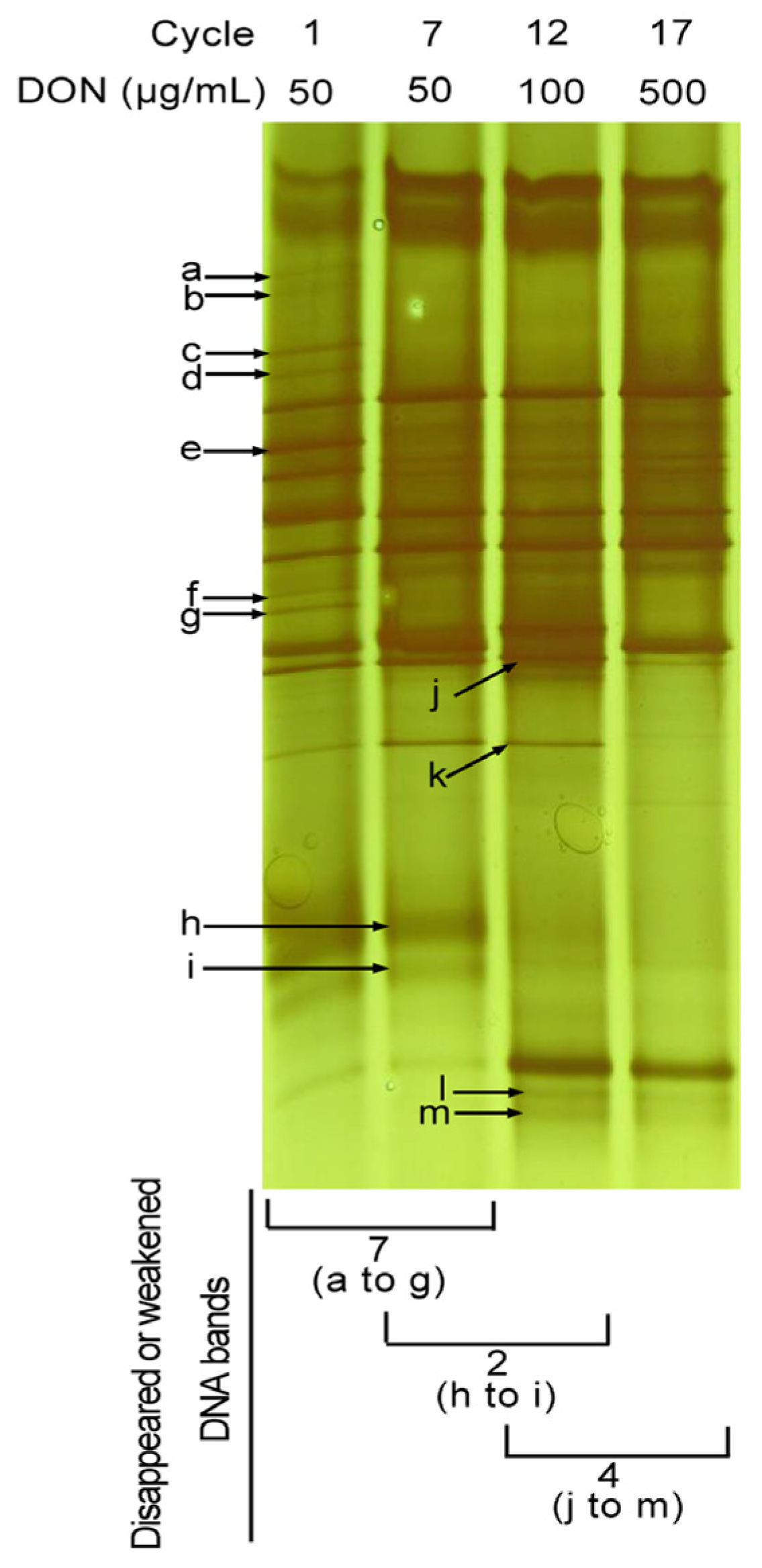
2.4. Bacterial Culture Complexity of PGC-3 after Subculturing under Different DON Concentrations
2.5. Bacterial Culture Compositions and the Dynamics of Present Genera
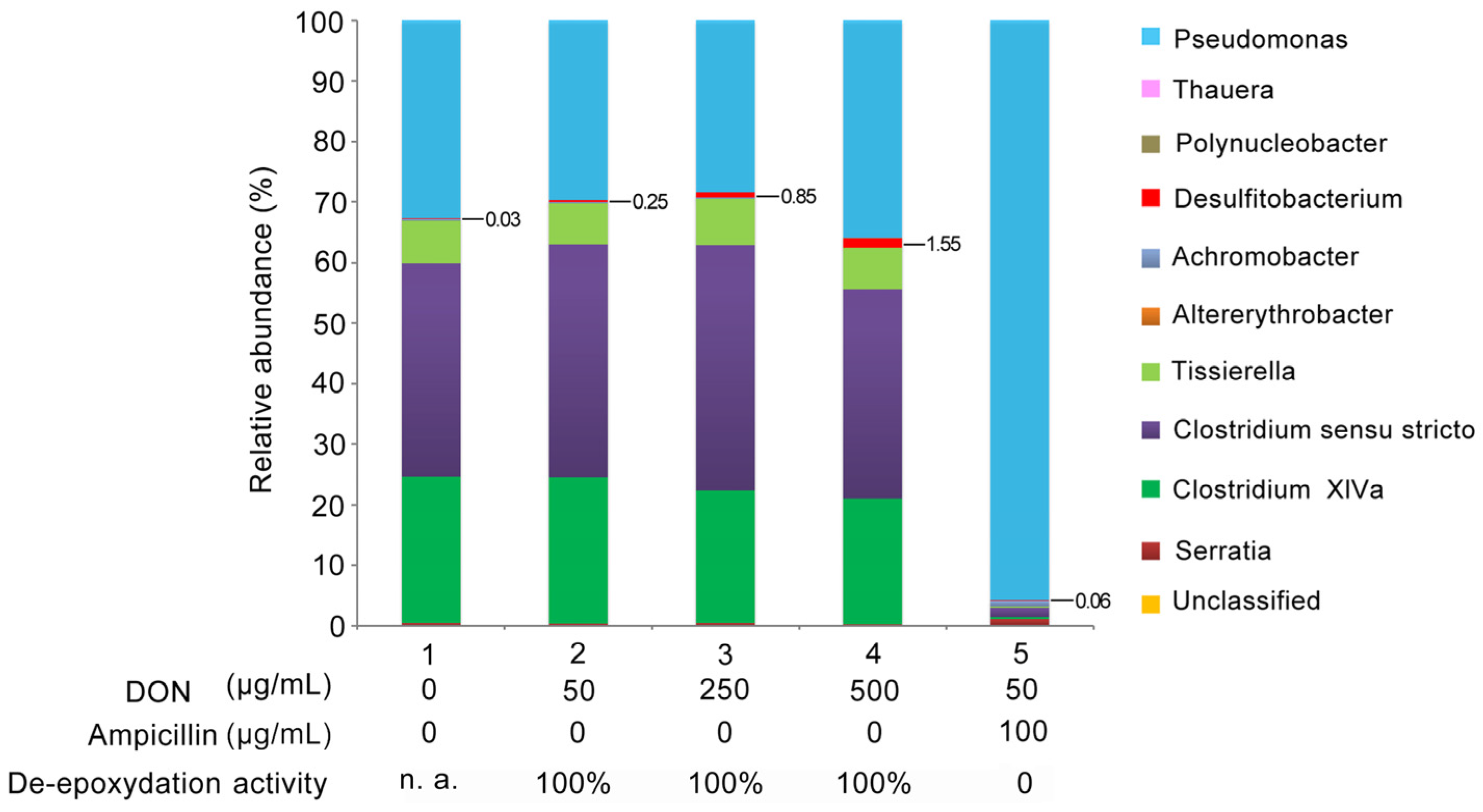
2.6. The Variation in Bacterial Abundance in Response to Increased DON Selective Pressure
3. Discussion
4. Materials and Methods
4.1. Trichothecenes and Culture Media
4.2. Soil Samples
4.3. Screening of Soil Samples
4.4. Extraction and Analysis of Trichothecenes and Their Metabolites
4.5. Temperature and pH Conditions Used for DON De-Epoxydation
4.6. Genomic DNA Extraction
4.7. PCR-DGGE Analysis
4.8. Analysis of Bacterial Population and Their Representing Species
4.9. Bacterial Sub-Culturing under Diverse Growth Conditions
4.10. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bai, G.H.; Shaner, G. Management and resistance in wheat and barley to Fusarium head blight. Annu. Rev. Phytopathol. 2004, 42, 135–161. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.M.; Nicholson, P. Community ecology of fungal pathogens causing wheat head blight. Annu. Rev. Phytopathol. 2009, 47, 83–103. [Google Scholar] [CrossRef] [PubMed]
- Starkey, D.E.; Ward, T.J.; Aoki, T.; Gale, L.R.; Kistler, H.C.; Geiser, D.M.; Suga, H.; Toth, B.; Varga, J.; O’Donnell, K. Global molecular surveillance reveals novel Fusarium head blight species and trichothecene toxin diversity. Fungal Genet. Biol. 2007, 44, 1191–1204. [Google Scholar] [CrossRef] [PubMed]
- Foroud, N.A.; Eudes, F. Trichothecenes in cereal grains. Int. J. Mol. Sci. 2009, 10, 147–173. [Google Scholar] [CrossRef] [PubMed]
- Ndoye, M.; Zhang, J.B.; Wang, J.H.; Gong, A.D.; Li, H.P.; Qu, B.; Li, S.J.; Liao, Y.C. Nivalenol and 15-acetyldeoxynivalenol chemotypes of Fusarium graminearum clade species are prevalent on maize throughout China. J. Phytopathol. 2012, 160, 519–524. [Google Scholar] [CrossRef]
- Zhang, J.B.; Li, H.P.; Dang, F.J.; Qu, B.; Xu, Y.B.; Zhao, C.S.; Liao, Y.C. Determination of the trichothecene mycotoxin chemotypes and associated geographical distribution and phylogenetic species of the Fusarium graminearum clade from China. Mycol. Res. 2007, 111, 967–975. [Google Scholar] [CrossRef] [PubMed]
- Placinta, C.M.; D’Mello, J.P.F.; Macdonald, A.M.C. A review of worldwide contamination of cereal grains and animal feed with Fusarium mycotoxins. Anim. Feed Sci. Technol. 1999, 78, 21–37. [Google Scholar] [CrossRef]
- Streit, E.; Schatzmayr, G.; Tassis, P.; Tzika, E.; Marin, D.; Taranu, I.; Tabuc, C.; Nicolau, A.; Aprodu, I.; Puel, O. Current situation of mycotoxin contamination and co-occurrence in animal feed—Focus on Europe. Toxins 2012, 4, 788–809. [Google Scholar] [CrossRef] [PubMed]
- Canady, R.A.; Coker, R.D.; Rgan, S.K.; Krska, R.; Kuiper-Goodman, T.; Olsen, M.; Pestka, J.J.; Resnik, S.; Schlatter, J. Safety evaluation of certain mycotoxins in food: Prepared by the fifty-sixth meeting of the Joint FAO/WHO Expert Committee on Food Additives. In Proceedings of the World Health Organization International Programme on Chemical Safety, Geneva, Switzerland, 6–15 February 2001; pp. 420–555.
- Zhang, J.B.; Wang, J.H.; Gong, A.D.; Chen, F.F.; Song, B.; Li, X.; Li, H.P.; Peng, C.H.; Liao, Y.C. Natural occurrence of Fusarium head blight, mycotoxins and mycotoxin-producing isolates of Fusarium in commercial fields of wheat in Hubei. Plant Pathol. 2013, 62, 92–102. [Google Scholar] [CrossRef]
- Parry, D.W.; Jenkinson, P.; Mcleod, L. Fusarium ear blight (scab) in small grain cereals—A review. Plant Pathol. 1995, 44, 207–238. [Google Scholar] [CrossRef]
- Nakajima, T. Progress and outlook for the control of nivalenol and deoxynivalenol contamination due to Fusarium head blight in wheat. Mycotoxins 2007, 57, 129–134. [Google Scholar] [CrossRef]
- Ueno, Y.; Nakayama, K.; Ishii, K.; Tashiro, F.; Minoda, Y.; Omori, T.; Komagata, K. Metabolism of T-2 toxin in Curtobacterium sp. strain 114-2. Appl. Environ. Microbiol. 1983, 46, 120–127. [Google Scholar] [PubMed]
- Rotter, B.A.; Prelusky, D.B.; Pestka, J.J. Toxicology of deoxynivalenol (vomitoxin). J. Toxicol. Environ. Health 1996, 48, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Pestka, J.J. Deoxynivalenol: Mechanisms of action, human exposure, and toxicological relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, A.E.; Proctor, R.H.; Bai, G.H.; McCormick, S.P.; Shaner, G.; Buechley, G.; Hohn, T.M. Reduced virulence of trichothecene-nonproducing mutants of Gibberella zeae in wheat field tests. Mol. Plant-Microbe Interact. 1996, 9, 775–781. [Google Scholar] [CrossRef]
- Bai, G.H.; Desjardins, A.E.; Plattner, R.D. Deoxynivalenol-nonproducing Fusarium graminearum causes initial Infection, but does not cause disease spread in wheat spikes. Mycopathologia 2002, 153, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Kris, A.; Adriaan, V.; Monica, H.; Geert, H. Deoxynivalenol: A major player in the multifaceted response of Fusarium to its environment. Toxins 2013, 6, 1–19. [Google Scholar]
- Zhou, T.; He, J.; Gong, J. Microbial transformation of trichothecene mycotoxins. World Mycotoxin J. 2008, 1, 23–30. [Google Scholar] [CrossRef]
- He, J.W.; Zhou, T.; Young, J.C.; Boland, G.J.; Scott, P.M. Chemical and biological transformations for detoxification of trichothecene mycotoxins in human and animal food chains: A review. Trends Food Sci. Technol. 2010, 21, 67–76. [Google Scholar] [CrossRef]
- Karlovsky, P. Biological detoxification of the mycotoxin deoxynivalenol and its use in genetically engineered crops and feed additives. Appl. Microbiol. Biotechnol. 2011, 91, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Eriksen, G.S.; Pettersson, H.; Lindberg, J.E. Comparative cytotoxicity of deoxynivalenol, nivalenol, their acetylated derivatives and de-epoxy metabolites. Food Chem. Toxicol. 2004, 42, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Pierron, A.; Mimoun, S.; Murate, L.S.; Loiseau, N.; Lippi, Y.; Bracarense, A.P.F.L.; Schatzmayr, G.; He, J.W.; Zhou, T.; Moll, W.D. Microbial biotransformation of Don: Molecular basis for reduced toxicity. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- King, R.R.; Mcqueen, R.E.; Levesque, D.; Greenhalgh, R. Transformation of deoxynivalenol (vomitoxin) by rumen microorganisms. J. Agric. Food Chem. 1984, 32, 1181–1183. [Google Scholar] [CrossRef]
- Swanson, S.P.; Rood, H.D.; Behrens, J.C.; Sanders, P.E. Preparation and characterization of the deepoxy trichothecenes: Deepoxy HT-2, deepoxy T-2 triol, deepoxy T-2 tetraol, deepoxy 15-monoacetoxyscirpenol, and deepoxy scirpentriol. Appl. Environ. Microbiol. 1987, 53, 2821–2826. [Google Scholar] [PubMed]
- He, P.; Young, L.G.; Forsberg, C. Microbial transformation of deoxynivalenol (vomitoxin). Appl. Environ. Microbiol. 1992, 58, 3857–3863. [Google Scholar] [PubMed]
- Young, J.C.; Zhou, T.; Yu, H.; Zhu, H.; Gong, J. Degradation of trichothecene mycotoxins by chicken intestinal microbes. Food Chem. Toxicol. 2007, 45, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Guan, S.; He, J.W.; Young, J.C.; Zhu, H.; Li, X.; Ji, C.; Zhou, T. Transformation of trichothecene mycotoxins by microorganisms from fish digesta. Aquaculture 2009, 290, 290–295. [Google Scholar] [CrossRef]
- Yu, H.; Zhou, T.; Gong, J.; Young, C.; Su, X.; Li, X.Z.; Zhu, H.; Tsao, R.; Yang, R. Isolation of deoxynivalenol-transforming bacteria from the chicken intestines using the approach of PCR-DGGE guided microbial selection. BMC Microbiol. 2010, 10. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, E.; Binder, E.M.; Heidler, D.; Krska, R. Structural characterization of metabolites after the microbial degradation of type A trichothecenes by the bacterial strain BBSH 797. Food Addit. Contam. 2002, 19, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Sato, I.; Koitabashi, M.; Yoshida, S.; Imai, M.; Tsushima, S. A novel actinomycete derived from wheat heads degrades deoxynivalenol in the grain of wheat and barley affected by Fusarium head blight. Appl. Microbiol. Biotechnol. 2012, 96, 1059–1070. [Google Scholar] [CrossRef] [PubMed]
- Islam, R.; Zhou, T.; Young, J.C.; Goodwin, P.H.; Pauls, K.P. Aerobic and anaerobic de-epoxydation of mycotoxin deoxynivalenol by bacteria originating from agricultural soil. World J. Microbiol. Biotechnol. 2012, 28, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Ding, N.; Xing, F.; Liu, X.; Selvaraj, J.N.; Wang, L.; Zhao, Y.; Wang, Y.; Guo, W.; Dai, X.; Liu, Y. Variation in fungal microbiome (mycobiome) and aflatoxin in stored in-shell peanuts at four different areas of china. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Völkl, A.; Vogler, B.; Schollenberger, M.; Karlovsky, P. Microbial detoxification of mycotoxin deoxynivalenol. J. Basic Microbiol. 2004, 44, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Utkin, I.; Woese, C.; Wiegel, J. Isolation and characterization of Desulfitobacterium dehalogenans gen. nov., sp. nov., an anaerobic bacterium which reductively dechlorinates chlorophenolic compounds. Int. J. Syst. Bacteriol. 1994, 44, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Beaudet, R.; Lévesque, M.J.; Villemur, R.; Lanthier, M.; Chénier, M.; Lépine, F.; Bisaillon, J.G. Anaerobic biodegradation of pentachlorophenol in a contaminated soil inoculated with a methanogenic consortium or with Desulfitobacterium frappieri strain PCP-1. Appl. Microbiol. Biotechnol. 1998, 50, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Kaksonen, A.H.; Plumb, J.J.; Robertson, W.J.; Franzmann, P.D. Culturable diversity and community fatty acid profiling of sulfate-reducing fluidized-bed reactors treating acidic, metal-containing wastewater. Geomicrobiol. J. 2004, 21, 469–480. [Google Scholar] [CrossRef]
- Villemur, R.; Lanthier, M.; Beaudet, R.; Lepine, F. The Desulfitobacterium genus. FEMS Microbiol. Rev. 2006, 30, 706–733. [Google Scholar] [CrossRef] [PubMed]
- Theisen, S.; Berger, S. Screening of epoxide hydrolase producing microorganisms for biotransformation of deoxynivalenol. Mycotoxin Res. 2005, 21, 71–73. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Ohta, M.; Ueno, Y. Reactivity of 12,13-epoxytrichothecenes with epoxide hydrolase, glutathione-s-transferase and glutathione. Chem. Pharm. Bull. 1977, 25, 3410–3414. [Google Scholar] [CrossRef]
- Goodstadt, L.; Ponting, C.P. Vitamin k epoxide reductase: Homology, active site and catalytic mechanism. Trends Biochem. Sci. 2004, 29, 289–292. [Google Scholar] [CrossRef] [PubMed]
- Rupnik, M.; Wilcox, M.H.; Gerding, D.N. Clostridium difficile infection: New developments in epidemiology and pathogenesis. Nat. Rev. Microbiol. 2009, 7, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Nuyts, S.; Van, M.L.; Theys, J.; Landuyt, W.; Lambin, P.; Anné, J. Clostridium spores for tumor-specific drug delivery. Anti-Cancer Drugs 2002, 13, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Stover, C.K.; Pham, X.Q.; Erwin, A.L.; Mizoguchi, S.D.; Warrener, P.; Hickey, M.J.; Brinkman, F.S.L.; Hufnagle, W.O.; Kowalik, D.J.; Lagrou, M. Complete genome sequence of pseudomonas aeruginosa pao1, an opportunistic pathogen. Nature 2000, 406, 959–964. [Google Scholar] [PubMed]
- Buell, C.R.; Joardar, V.; Lindeberg, M.; Selengut, J.; Paulsen, I.T.; Gwinn, M.L.; Dodson, R.J.; Deboy, R.T.; Durkin, A.S.; Kolonay, J.F. The complete sequence of the Arabidopsis and Tomato pathogen pseudomonas syringae pv. Tomato dc3000. Proc. Natl. Acad. Sci. USA 2003, 100, 10181–10186. [Google Scholar] [CrossRef] [PubMed]
- O’Mahony, M.M.; Dobson, A.; Barnes, J.D.; Singleton, I. The use of ozone in the remediation of polycyclic aromatic hydrocarbon contaminated soil. Chemosphere 2006, 63, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Feinberg, B.; McLaughlin, C.S. Biochemical mechanism of action of trichothecene mycotoxins. In Trichothecene Mycotoxicosis Pathophysiologic Effects; Beasley, V.R., Ed.; CRC-Press: Boca Rata, FL, USA, 1989; Volume 1, pp. 27–35. [Google Scholar]
- Swanson, S.P.; Corley, R.A. The distribution, metabolism, and excretion of trichothecene mycotoxins. In Trichothecene Mycotoxicosis Pathophysiologic Effects; Beasley, V.R., Ed.; CRC-Press: Boca Rata, FL, USA, 1989; Volume 1, pp. 37–61. [Google Scholar]
- Swanson, S.P.; Helaszek, C.; Buck, W.B.; Rood, H.D., Jr.; Haschek, W.M. The role of intestinal microflora in the metabolism of trichothecene mycotoxins. Food Chem. Toxicol. 1988, 26, 823–829. [Google Scholar] [CrossRef]
- Song, X.S.; Xing, S.; Li, H.P.; Zhang, J.B.; Qu, B.; Jiang, J.H.; Fan, C.; Yang, P.; Liu, J.L.; Hu, Z.Q.; et al. An antibody that confers plant disease resistance targets a membrane-bound glyoxal oxidase in Fusarium. New Phytol. 2016, 210, 997–1010. [Google Scholar] [CrossRef] [PubMed]
- Clifford, L.J.; Jia, Q.S.; Pestka, J.J. An improved method for the purification of the trichothecene deoxynivalenol (vomitoxin) from Fusarium graminearum culture. J. Agric. Food Chem. 2003, 51, 521–523. [Google Scholar] [CrossRef] [PubMed]
- Ikunaga, Y.; Sato, I.; Grond, S.; Numaziri, N.; Yoshida, S.; Yamaya, H.; Hiradate, S.; Hasegawa, M.; Toshima, H.; Koitabashi, M.; et al. Nocardioides sp. strain WSN05–2, isolated from a wheat field, degrades deoxynivalenol, producing the novel intermediate 3-epi-deoxynivalenol. Appl. Microbiol. Biotechnol. 2011, 89, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Li, H.P.; Qu, B.; Zhang, J.B.; Huang, T.; Chen, F.F.; Liao, Y.C. Development of a generic PCR detection of 3-acetyldeoxynivalenol-, 15-acetyldeoxynivalenol- and nivalenol-chemotypes of Fusarium graminearum clade. Int. J. Mol. Sci. 2008, 9, 2495–2504. [Google Scholar] [CrossRef] [PubMed]
- Nübel, U.; Engelen, B.; Felske, A.; Snaidr, J.; Wieshuber, A.; Amann, R.I.; Ludwig, W.; Backhaus, H. Sequence heterogeneities of genes encoding 16S rRNAs in Paenibacillus polymyxa detected by temperature gradient gel electrophoresis. J. Bacteriol. 1996, 178, 5636–5643. [Google Scholar] [PubMed]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glockner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2012, 41. [Google Scholar] [CrossRef] [PubMed]
- FASTX Toolkit. Available online: http://hannonlab.cshl.edu/fastx_toolkit/index.html (accessed on 22 September 2016).
- Niu, Q.; Li, P.; Hao, S.; Zhang, Y.; Kim, S.W.; Li, H.; Ma, X.; Gao, S.; He, L.; Wu, W.; et al. Dynamic distribution of the gut microbiota and the relationship with apparent crude fiber digestibility and growth stages in pigs. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]








© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, W.-J.; Yuan, Q.-S.; Zhang, Y.-B.; Guo, M.-W.; Gong, A.-D.; Zhang, J.-B.; Wu, A.-B.; Huang, T.; Qu, B.; Li, H.-P.; et al. Aerobic De-Epoxydation of Trichothecene Mycotoxins by a Soil Bacterial Consortium Isolated Using In Situ Soil Enrichment. Toxins 2016, 8, 277. https://doi.org/10.3390/toxins8100277
He W-J, Yuan Q-S, Zhang Y-B, Guo M-W, Gong A-D, Zhang J-B, Wu A-B, Huang T, Qu B, Li H-P, et al. Aerobic De-Epoxydation of Trichothecene Mycotoxins by a Soil Bacterial Consortium Isolated Using In Situ Soil Enrichment. Toxins. 2016; 8(10):277. https://doi.org/10.3390/toxins8100277
Chicago/Turabian StyleHe, Wei-Jie, Qing-Song Yuan, You-Bing Zhang, Mao-Wei Guo, An-Dong Gong, Jing-Bo Zhang, Ai-Bo Wu, Tao Huang, Bo Qu, He-Ping Li, and et al. 2016. "Aerobic De-Epoxydation of Trichothecene Mycotoxins by a Soil Bacterial Consortium Isolated Using In Situ Soil Enrichment" Toxins 8, no. 10: 277. https://doi.org/10.3390/toxins8100277
APA StyleHe, W. -J., Yuan, Q. -S., Zhang, Y. -B., Guo, M. -W., Gong, A. -D., Zhang, J. -B., Wu, A. -B., Huang, T., Qu, B., Li, H. -P., & Liao, Y. -C. (2016). Aerobic De-Epoxydation of Trichothecene Mycotoxins by a Soil Bacterial Consortium Isolated Using In Situ Soil Enrichment. Toxins, 8(10), 277. https://doi.org/10.3390/toxins8100277