
Toxicity and Growth Assessments of Three Thermophilic Benthic Dinoflagellates (Ostreopsis cf. ovata, Prorocentrum lima and Coolia monotis) Developing in the Southern Mediterranean Basin

 ,
,  ,
,
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Culture Observations
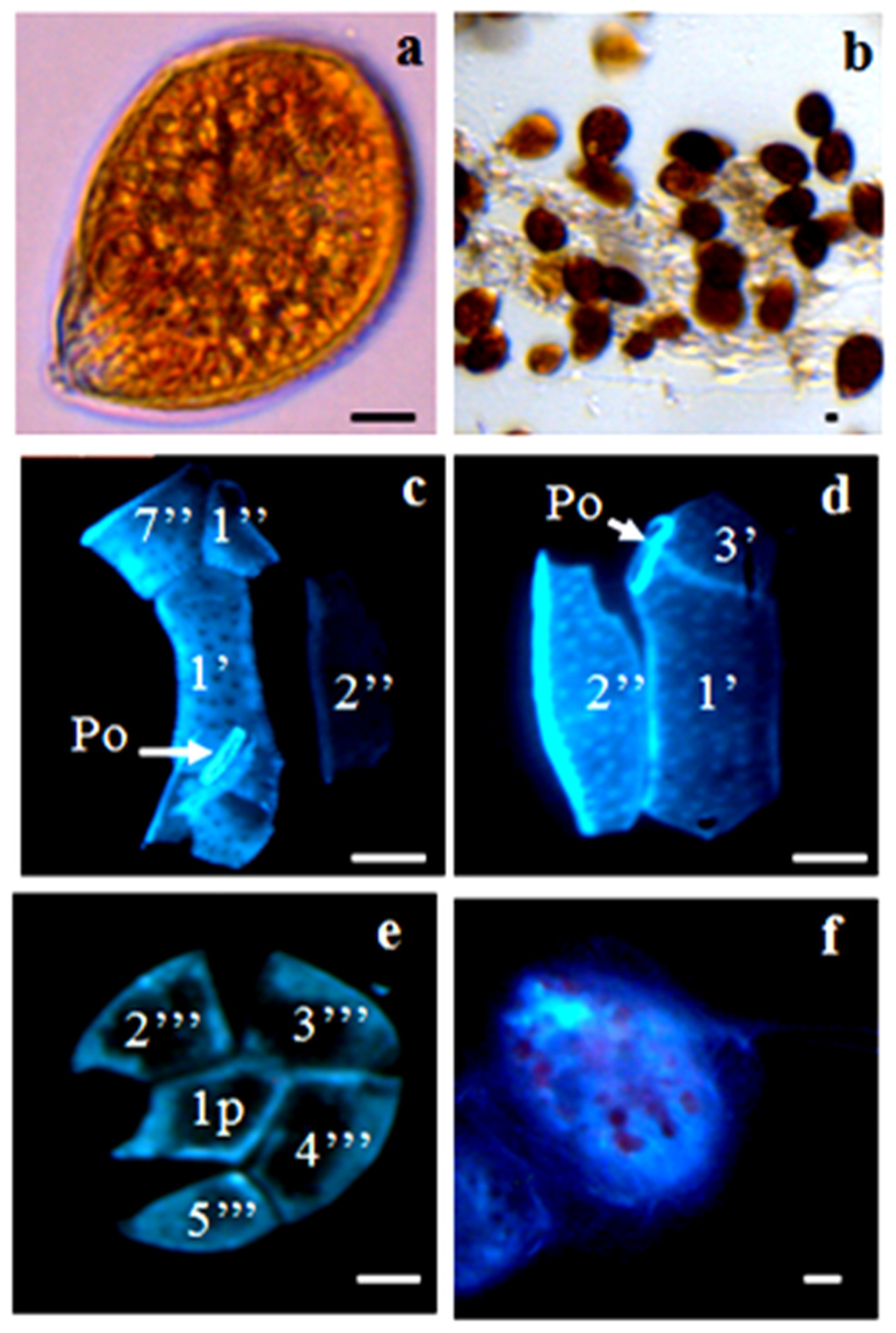
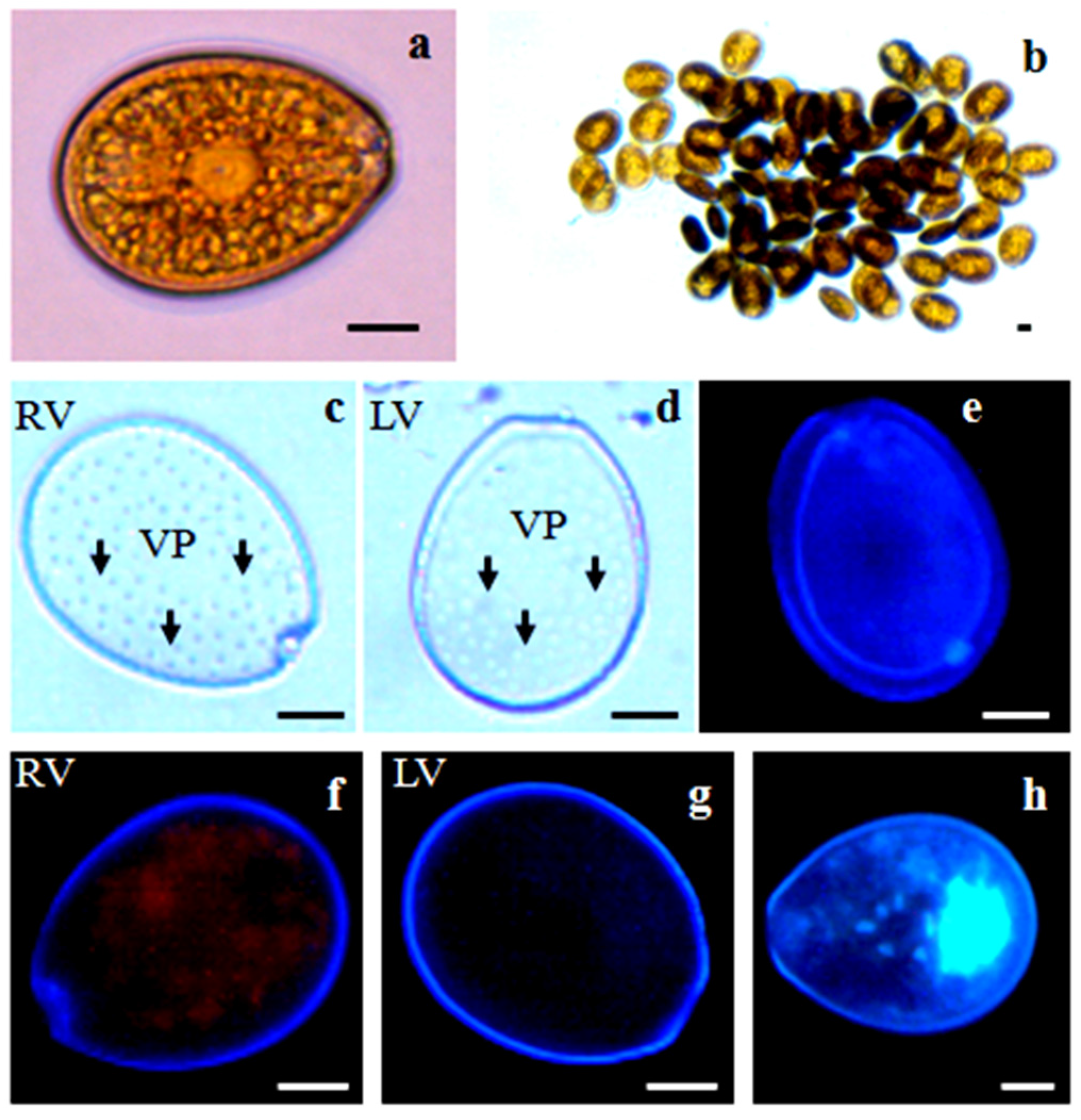
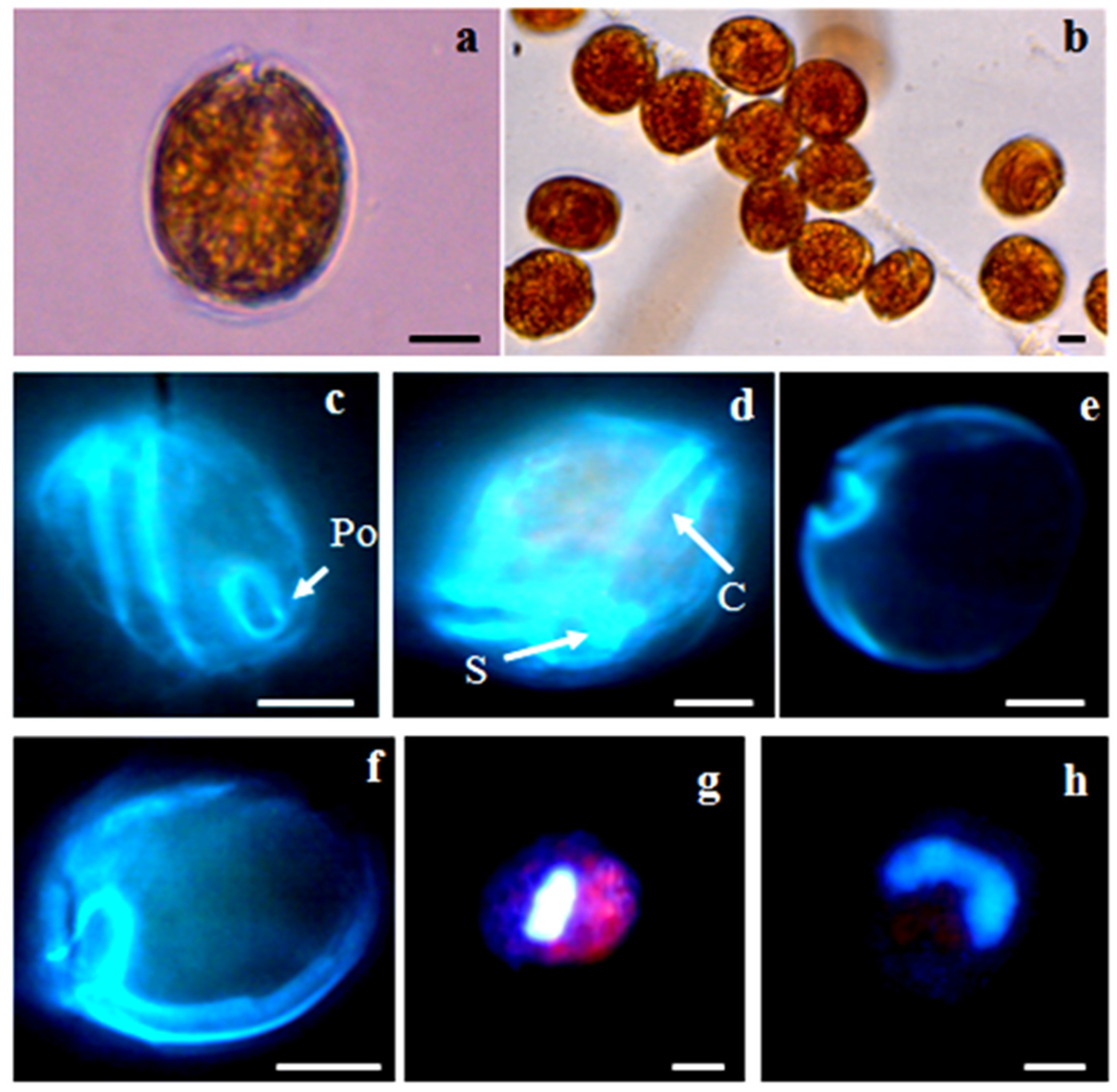
2.2. Morphology
2.2.1. Ostreopsis cf. ovata
2.2.2. Prorocentrum lima
2.2.3. Coolia monotis
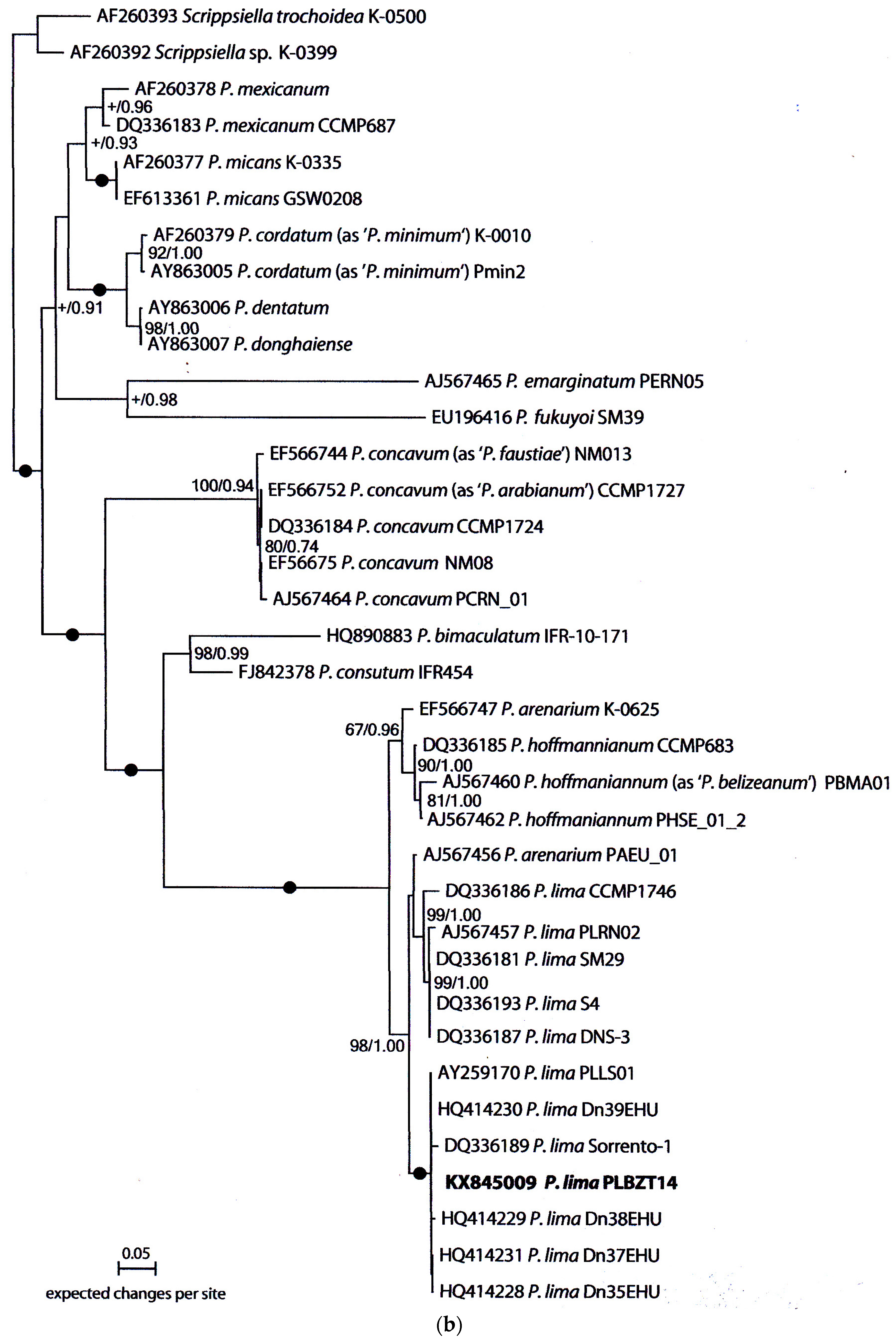
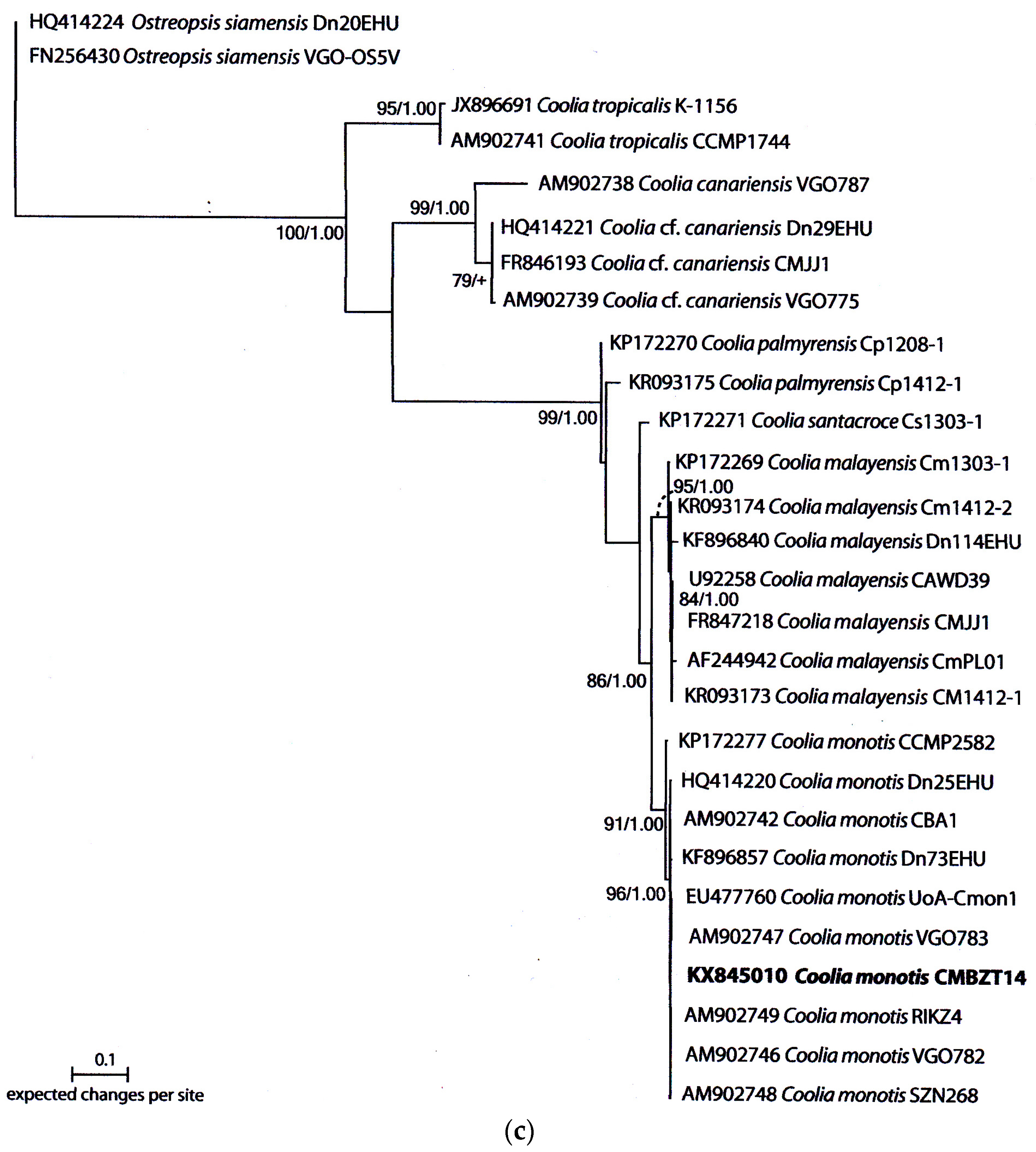
2.3. Molecular Analysis and Phylogeny
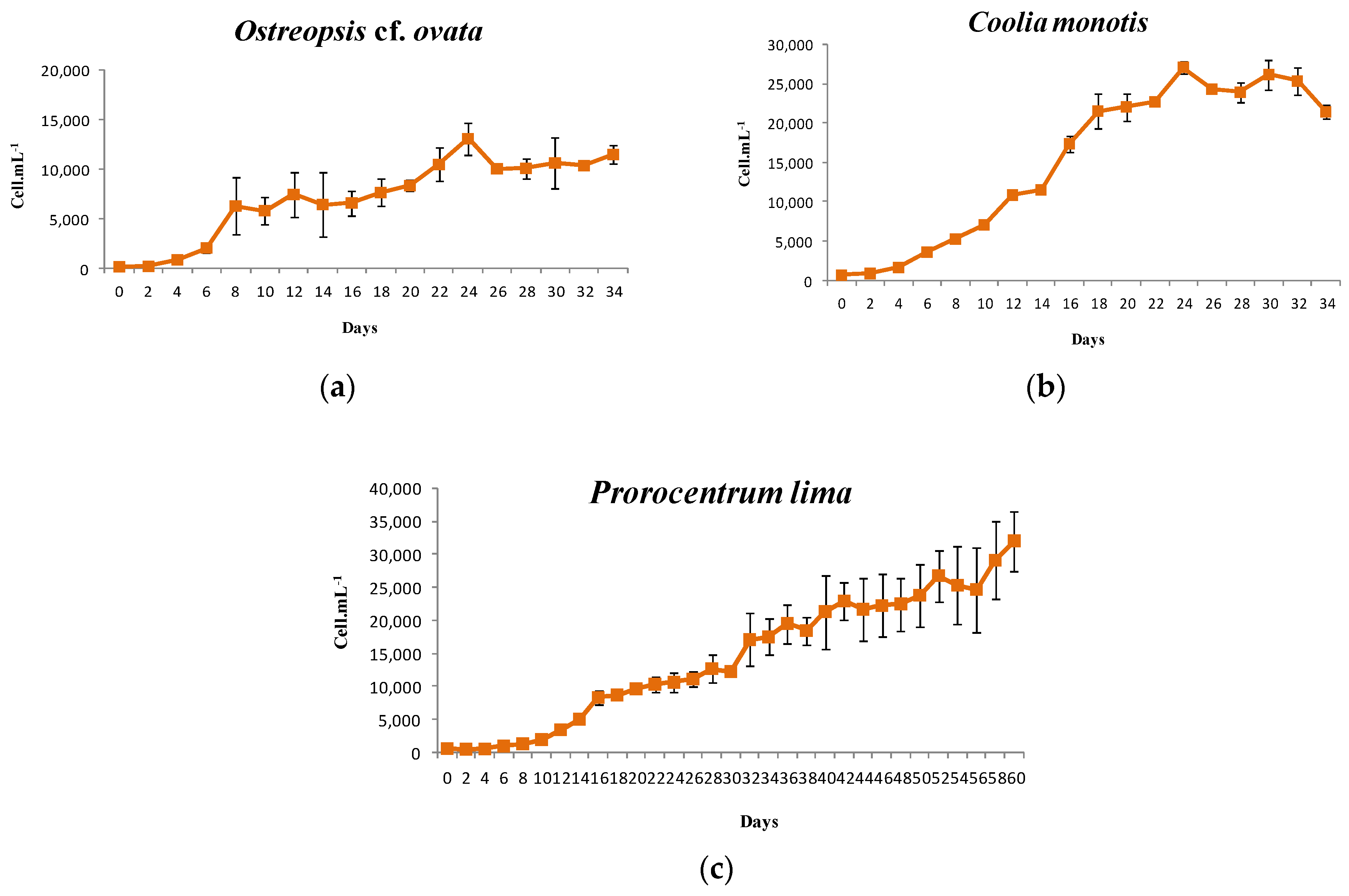
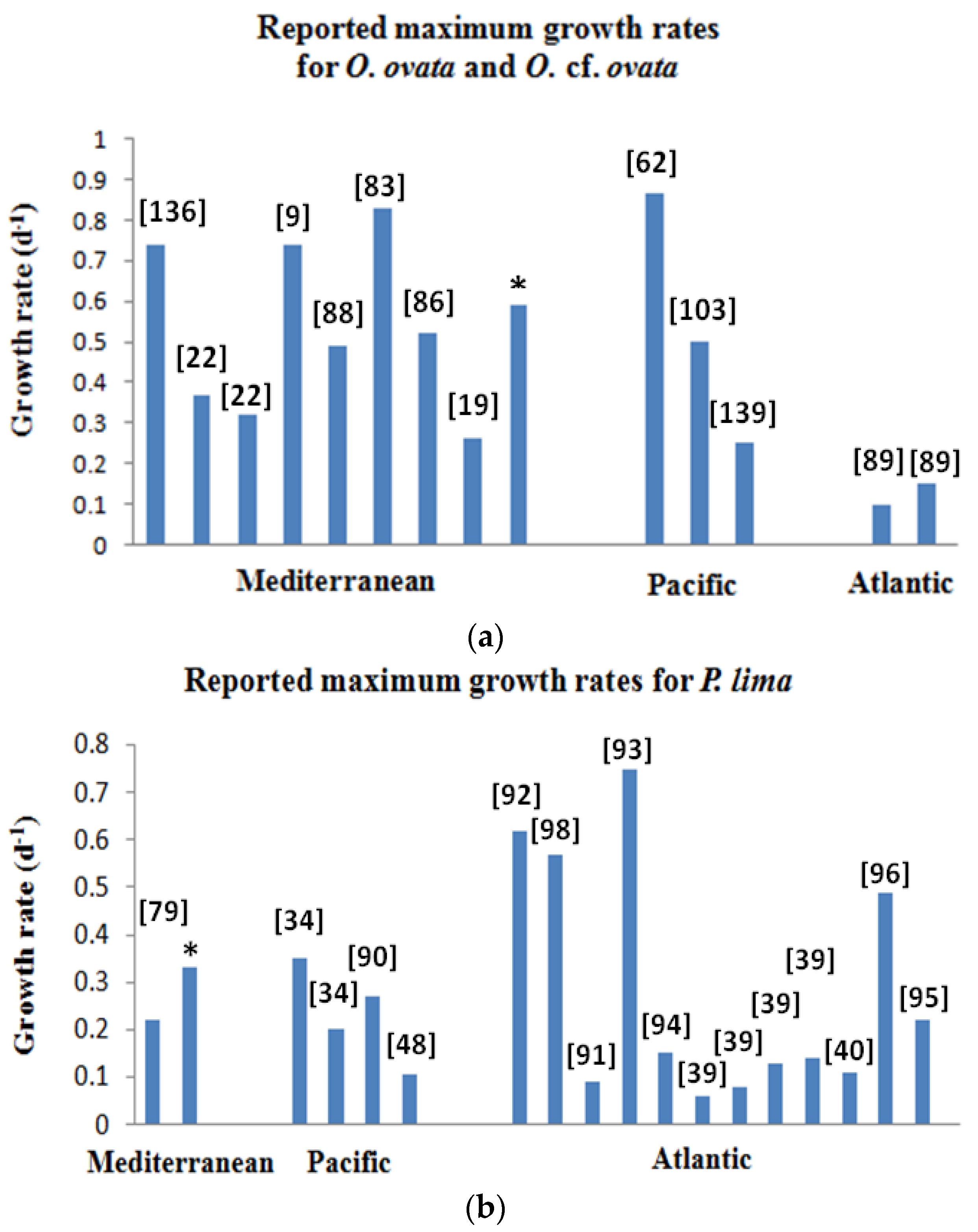
2.4. Growth Characteristics
2.4.1. Ostreopsis cf. ovata
2.4.2. Prorocentrum lima
2.4.3. Coolia monotis
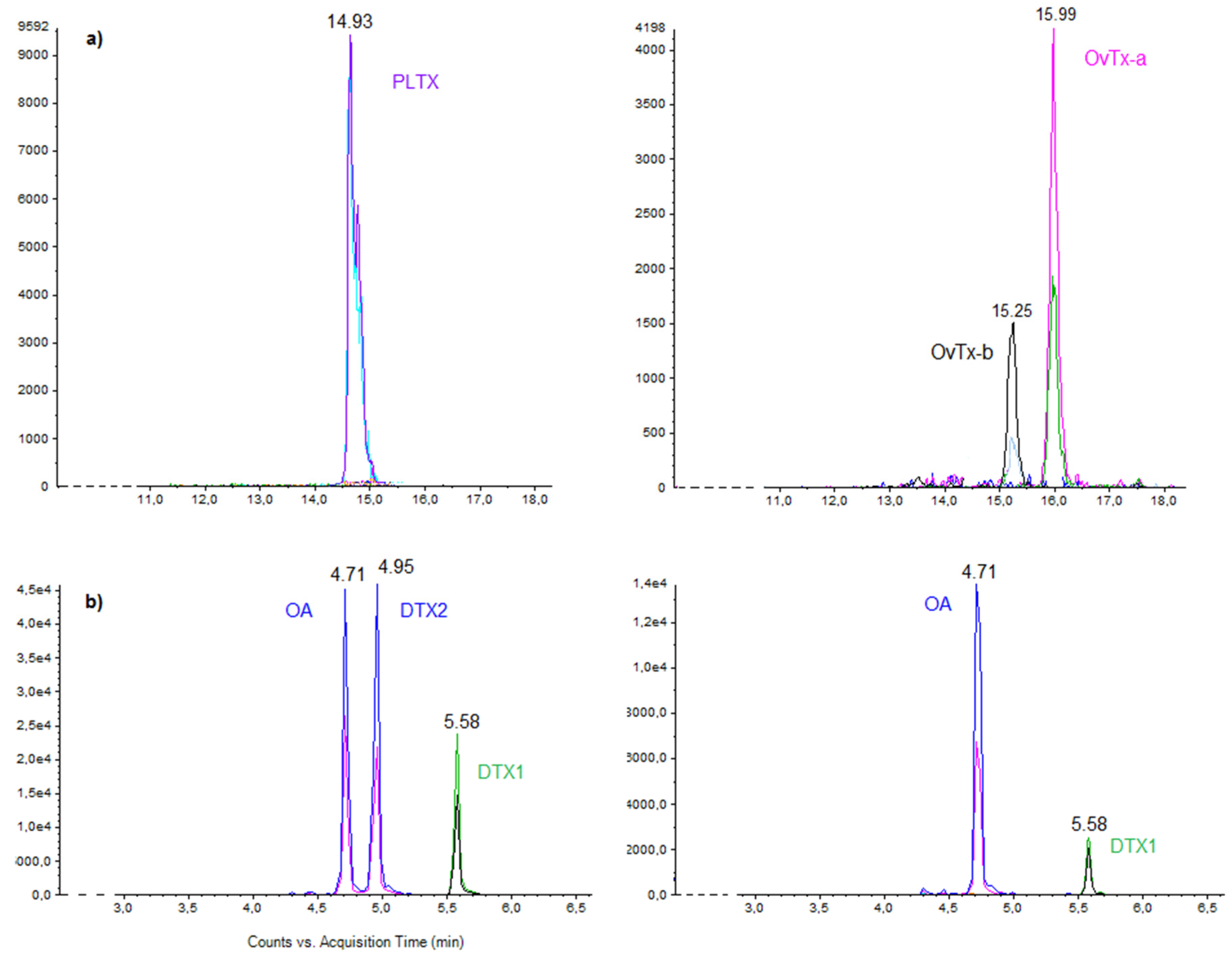
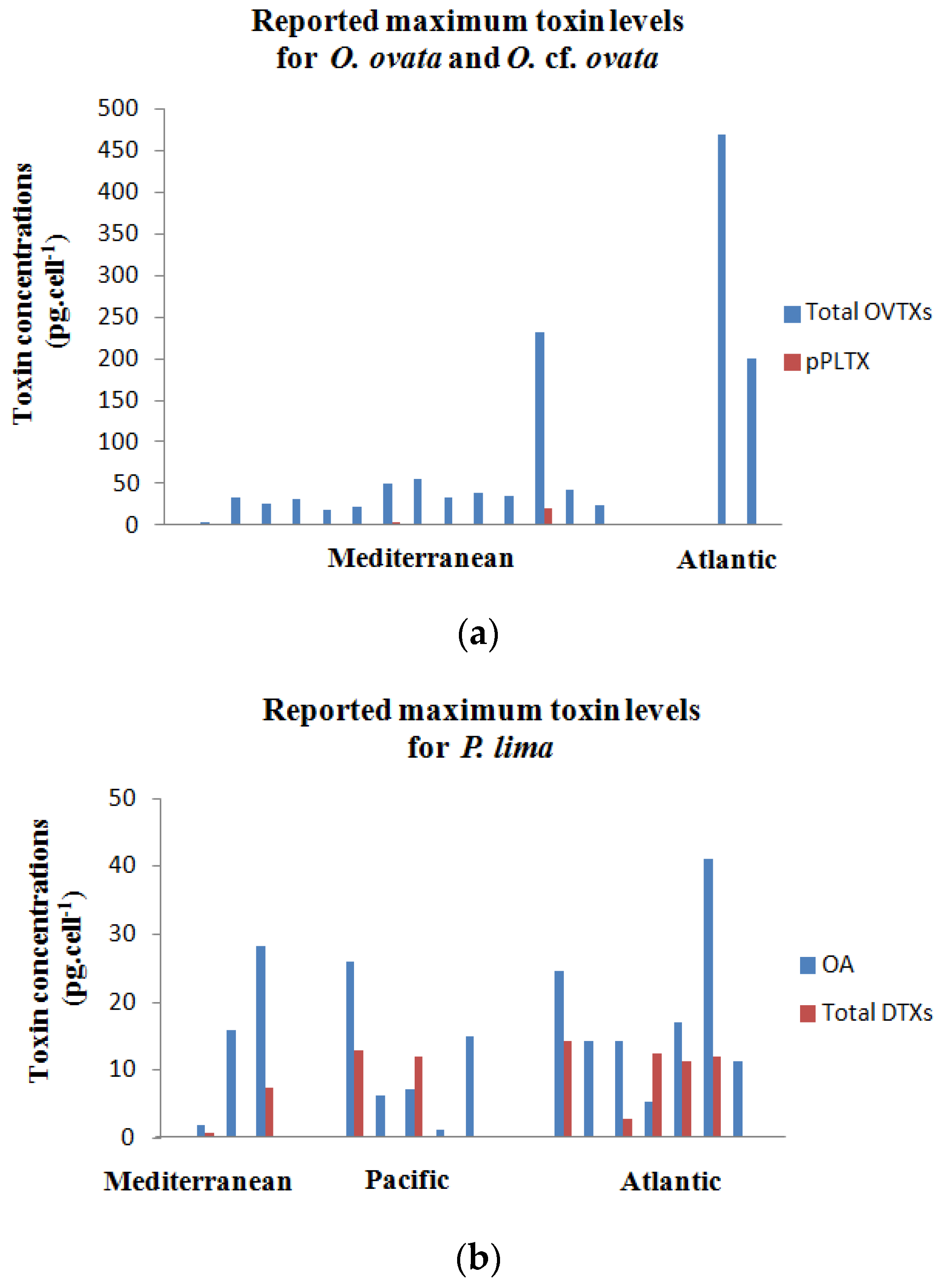
2.5. Toxin Profiles
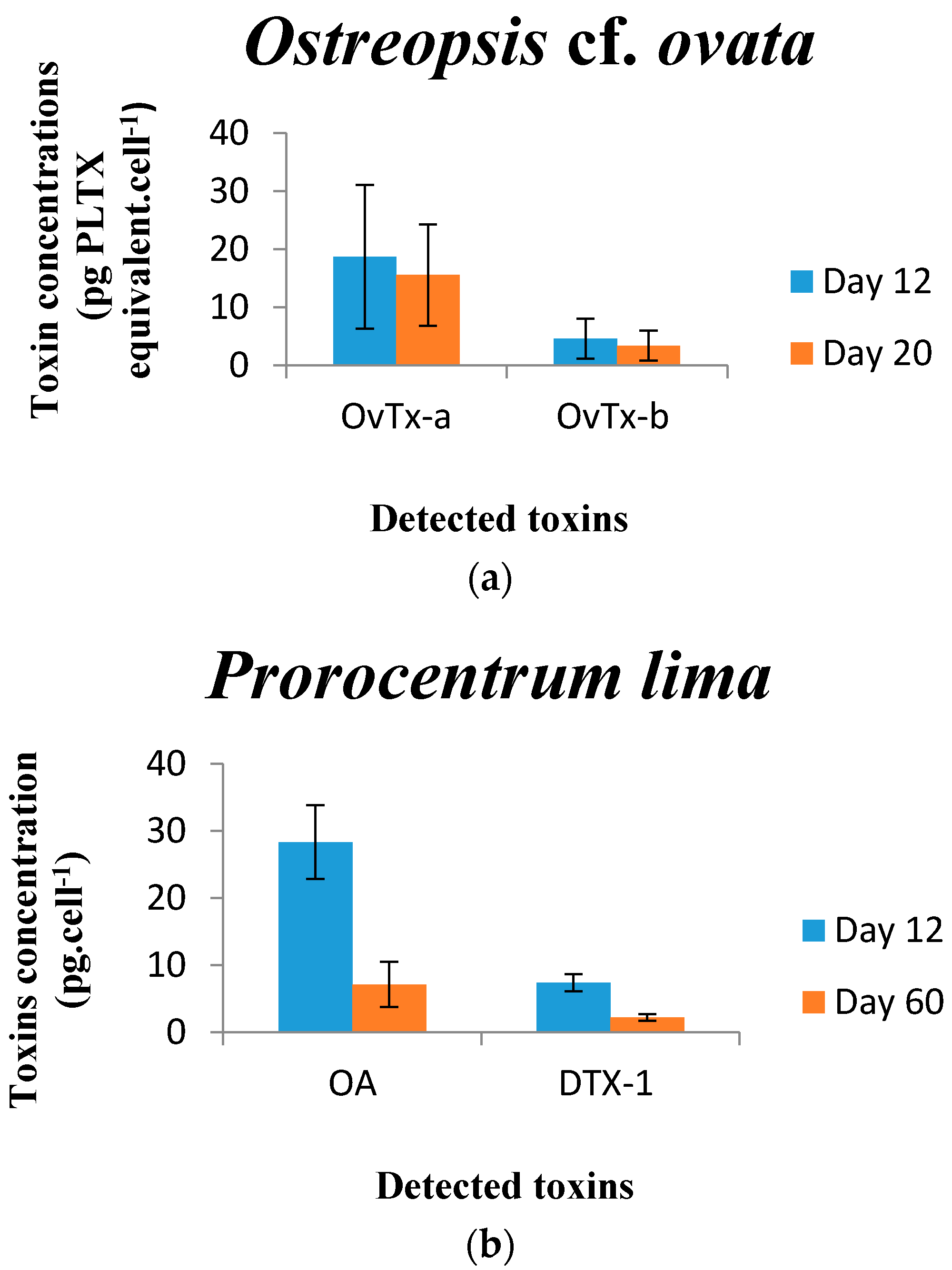
2.5.1. Ostreopsis cf. ovata
2.5.2. Prorocentrum lima
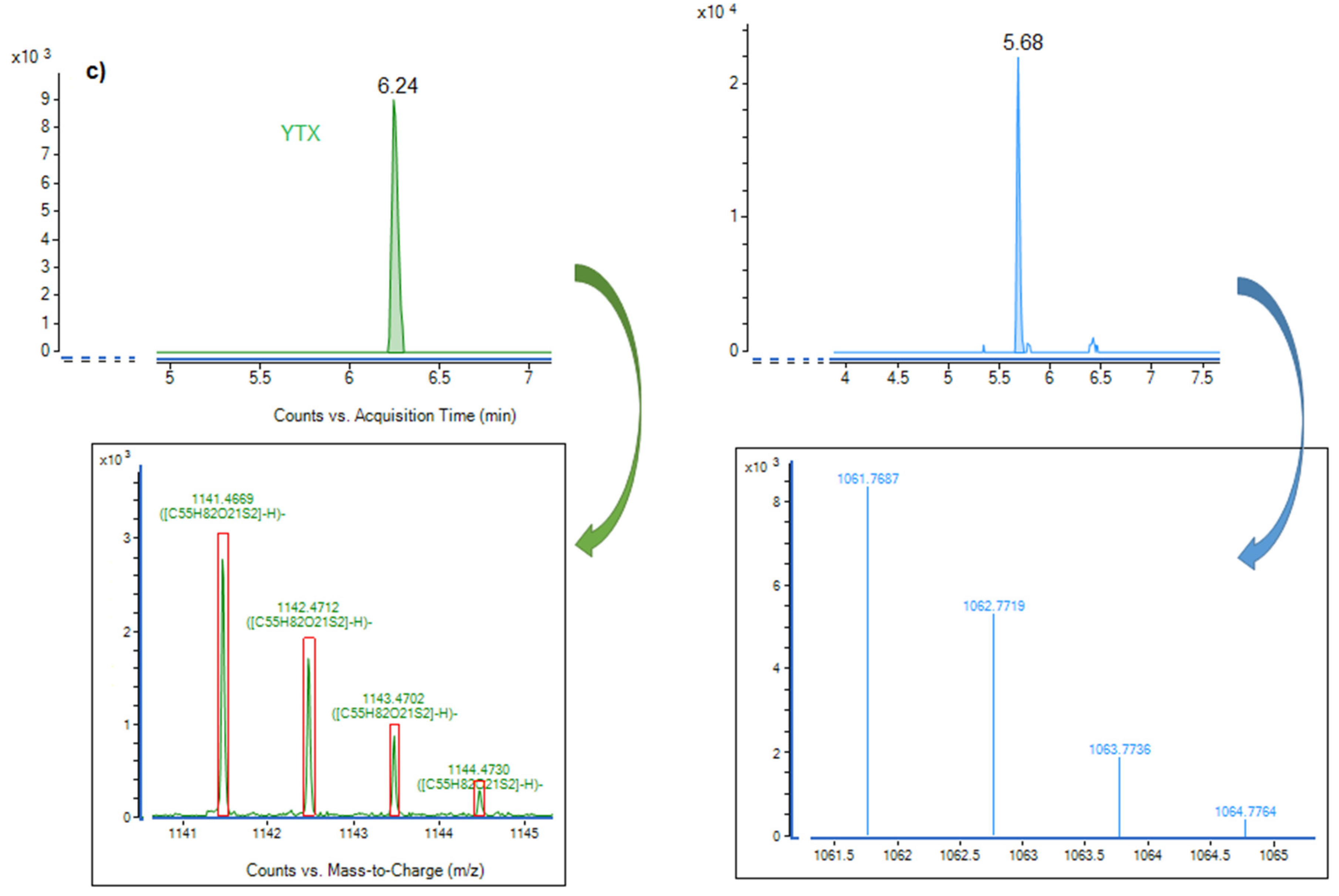
2.5.3. Coolia monotis
3. Conclusions
4. Experimental Section
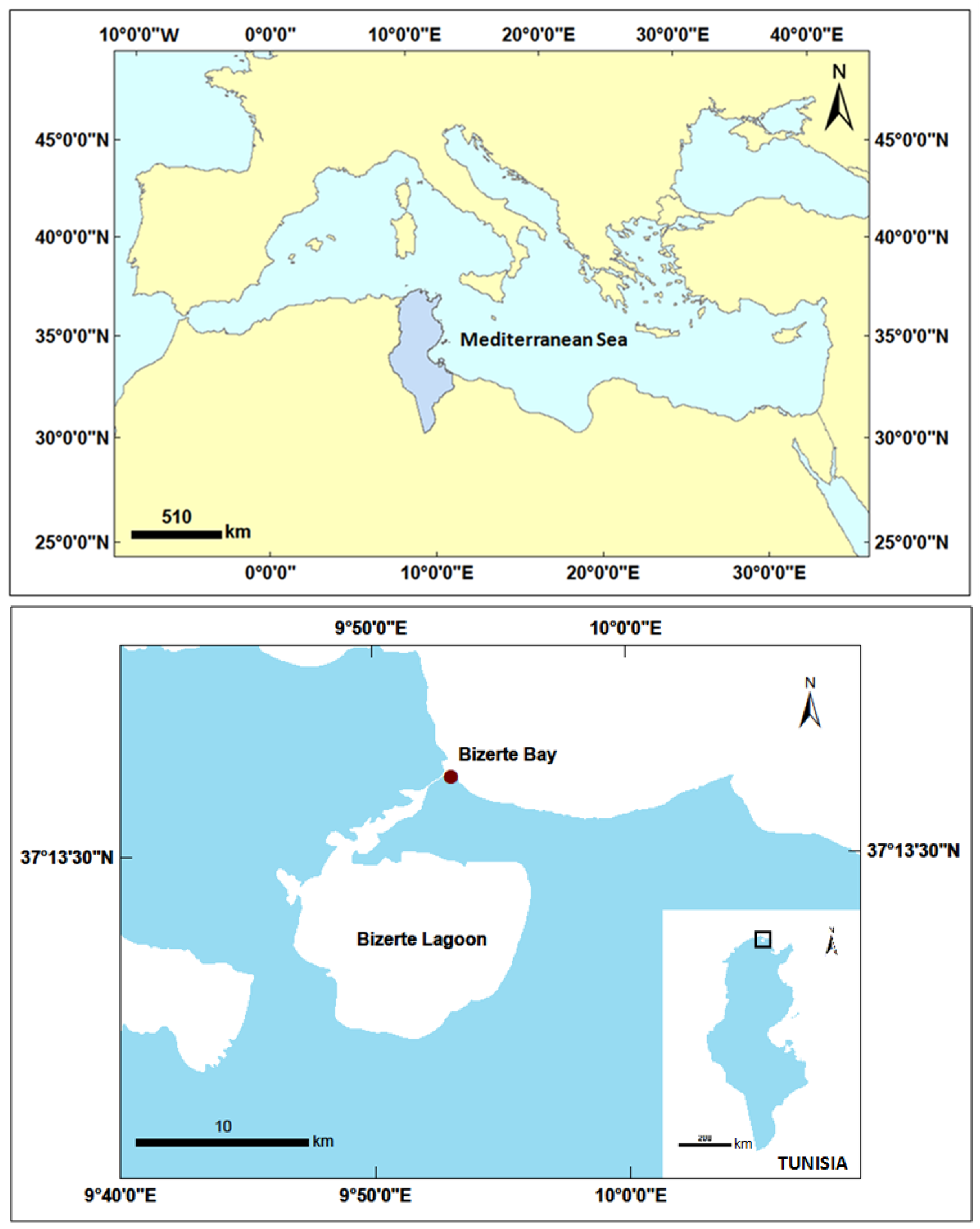
4.1. Sampling Site
4.2. Isolation and Culture Conditions
4.3. Morphology
4.4. Molecular Analysis and Phylogeny
4.4.1. DNA Extraction and PCR
4.4.2. Phylogeny
4.5. Growth Characteristics
4.6. Toxin Analysis
4.6.1. Sample Preparation
4.6.2. Instrumentation: LC-MS/MS Systems
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviation
| Opt | Optimum |
| Dt | Doubling time |
| µmax | Maximum growth rate |
| S-PES | Seawater with Provasoli’s ES supplement |
| S-ES-1 | Seawater with ES-1supplements |
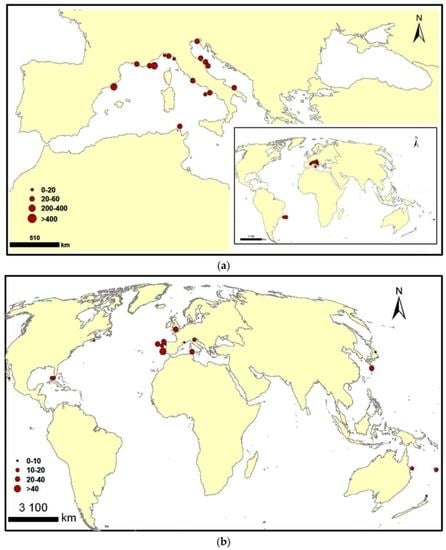
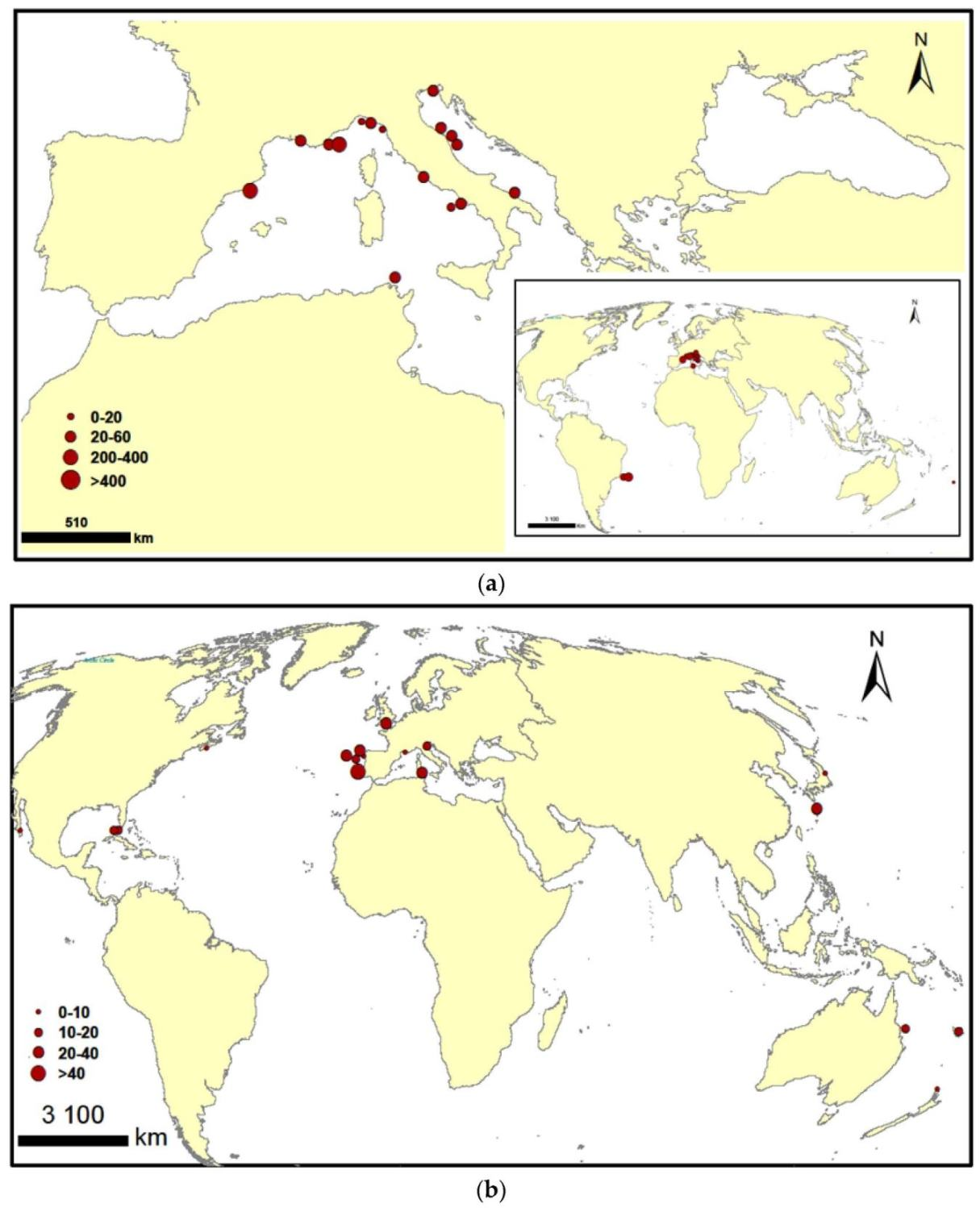
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain and Sampling Location | Temperature (°C) | Salinity | Irradiance (µmol photons.m−2·s−1) and L:D Cycle (h) | Culture Medium | Growth Rate (d−1) | Toxicity: Detected Toxins (pg·cell−1), Hemolytic Activity, Toxicity to Mice and Other Organisms | Isolated from | Reference |
|---|---|---|---|---|---|---|---|---|
| MEDITERRANEAN WATERS | ||||||||
| CNR-A1 (Italy, Tyrrhenian Sea, Gioia Tauro) | 17 ± 1 | ** | 100 (14L:10D) | K, F/20, F/2 | ** | Presence of palytoxin Hemolytic activity on human erythrocytes | Seawater Rhodophyceae Phaeophyceae | Penna et al. (2005) b [20] |
| CNR-D1 (Italy, Tyrrhenian Sea, La Spezia) | ||||||||
| CNR-Z1 (Spain, Balearic Sea, Paguera) | ||||||||
| ** (Greece, North Aegean coasts) | 19 ± 1 | ** | 70 (14L:10D) | F/2, K | ** | ** | Macrophytes | Aligizaki and Nikolaidis (2006) b [77] |
| ** (Italy, Ligurian coast, Genoa) | 25 | ** | 2000 lX (16L:8D) | K-Keller | ** | OVTX-a = 3.11/3.85, pPLTX = 0.40/0.55 | Rhodophyta, Chlorophyta, Phaeophyta | Ciminiello et al. (2008) b [24] |
| D483 (Italy, Gulf of Naples, Gaiola) | 18 | ** | 50 (12L:12D) | K/2 | ** | OVTX-a = 3.67–9.41, OVTX-b = 1.69–3.43, OVTX-c = 2.51–4.12, OVTX-d = 0.08–0.74, Mascarenotoxin-a = ND-0.47, Mascarenotoxin-c = ND-0.32 | Asparagopsis taxiformis | Rossi et al. (2010) a [28] |
| VGO820, VGO1049 (Spain, Catalonian coast, Llavaneres) | 20 | ** | 174.4 (10L:14D) | K/2, K, L1, Schreiber | 0.49–0.74 | ** | Seawater | Bravo et al. (2010) b [136] |
| OOAN0601 (Italy, Adriatic coast, Marche region, Numana) | 20 | 32 | 90 (16L:8D) | F/2 | ** | OVTX-a = 18, OVTX-b = 9, OVTX-c = 2, OVTX-d+e = 4, pPLTX = 0.2 | Seawater (proximity of Cystoseira sp. and Alsidium corallinum) | Ciminiello et al. (2010) b [25] |
| OOTL0602 (Italy, Tyrrhenian Sea, Lazio region, Porto Romano) | 20 | 35 | 90 (16L:8D) | F/2 | 0.32 | ≈OVTX-a = 14–25, pPLTX = 0.7–1.1 (in cells) | Seawater (proximity of Cystoseira sp. and Alsidium corallinum) | Guerrini et al. (2010) b [22] |
| OOAN0601 (Italy, Adriatic Sea, Marche region, Numana) | 20 | 35 | 90 (16L:8D) | F/2 | 0.37 | ≈OVTX-a = 18.5–31, pPLTX = 1.3–2.5 (in cells) | Seawater (proximity of Cystoseira sp. and Alsidium corallinum) | Guerrini et al. (2010) b [22] |
| KAC85 (Italy, Tyrrhenian Sea, Monte Argentario) | 16–24, 26,28,30 | 38 | 140 (16L:8D) | F/10 | 0.1–0.74 | Hemolytic activity on horse blood cells 20 °C = 18.1 ng·SnE·cell−1. 22 °C = 11.57 ng·SnE·cell−1. (saponin nano-equivalent per cell) | Seaweeds | Granéli et al. (2011) b [9] |
| ** (Italy, Northern Ionian Sea, Mar Piccolo, Mar Grande and Lido Bruno) | 24 ± 2 | 37 | 100 (12L:12D) | F/2 | ** | Live cells: strongly affected P. lividus embryonic development.Cell lysate: inhibited P. lividus embryonic development, were toxic to Artemia salina nauplii and induced hemolysis on human erythrocytes | Seawater Rocks scraping | Pagliara and Caroppo (2012) c [54] |
| OOAN0601 (Italy, Adriatic coast, Marche region, Numana) | 20,25,30 | 26,32,36,40 | 90,100–110 (16L:8D) | F/2 | 0.34–0.49 | OVTX-a, -b, -c, -d, -e, pPLTX. Total toxin content in cell pellets = 57–155 (µg·L−1) Hemolytic activity on sheep erythrocytes Toxic to Artemia sp. nauplii and to D. labrax | Seawater (proximity of Cystoseira sp. and Alcidium corallinum) | Pezzolesi et al. (2012) a [88] |
| D483 (Italy, Gulf of Naples) CBA-T (Italy, Portonovo) OS2T (Italy, Gulf of Trieste) | 18,22,26,30 | 36 | 50–200 (9L:15D, 12L:12D, 15L:9D) | K/2 | 0.18–0.83 | Strain D483: OVTX-a = 2.1–9.81, OVTX-b = 0.7–5.1, OVTX-c = 0.005–1.2, OVTX-d+e = 0.22–6.8, McTX-a = 0.006–0.47, McTX-c = ND-0.32 | ** | Scalco et al. (2012) a [83] |
| IFR-OST-0.1M (France, Marseille, Frioul Island, Morgiret) | 22 | 35 | 460 (16L:8D) | K, L1 | ** | OVTX-a = 50, pPLTX = 3.7 | Seawater (proximity of Dictyota sp. and Haliptilon virgatum) | Sechet et al. (2012) a [61] |
| IFR-OST-0.1V (France, Villefranche-sur-Mer Bay) | 22 | 35 | 460 (16L:8D) | K, L1 | ** | OVTX-a = 55, pPLTX = 2.5 | Seawater (dominant algae Halopteris scoparium) | Sechet et al. (2012) a [61] |
| CBA2-122 (Italy, Adriatic Sea, Portonovo) | 23 ± 1 | ** | 100 (14L:10D) | F/4 | ** | OVTX-f = 17, OVTX-a = 8, OVTX-b = 6, OVTX-c = 0.8, OVTX-d+e = 2, pPLTX = 0.1 | ** | Ciminiello et al. (2012) a [27] |
| OOAN0918 (Italy, Adriatic Sea, Passetto) | 20 | 36 | 90 (16L:8D) | F/2 | ** | ≈ OVTX-a = 8.5–19, OVTX-b = 5–11, OVTX-c = 1–2, OVTX-d+e = 3–6, pPLTX = 0.5–1 | Seawater | Vanucci et al. (2012) a [85] |
| C5 (Italy, Adriatic Sea, Gulf of Trieste, Canovella de’ Zoppoli) | 23 ± 1 | ** | 100 (14L:10D) | F/4 | ** | OVTX-a = 7.5–20, OVTX-b = 3.6–9.3, OVTX-c = 0.6–1.5, OVTX-d+e = 1.6–4.4, pPLTX = 0.03–0.08 | Seawater | Honsell et al. (2013) a [23] |
| OOAB0801 (Italy, Adriatic Sea, Puglia region) | 20 ± 1 | 36 | 110–120 (16L:8D) | F/2 | 0.52 | OVTX-a = 52%–55%, OVTX-b = 25%–29%, OVTX-c = 4%–7%, OVTX-d+e = 11%–16%, pPLTX = 1%–2%. Maximum PLTXs content = 21.5 | ** | Pezzolesi et al. (2014) a [86] |
| IFR-OST-03V (France, Villefranche sur-Mer) | 22 | 35 | 420 (16L:8D) | L1 | 0.26 | OVTX-a = 50%, OVTX-b = 25%, OVTX c = 9%, pPLTX = 8%, OVTX-d = 4%, OVTX-e = 3%, OVTX-f = 1%. Toxin content = 70–251 | Seawater (proximity of Stypocaulon sp. and Acetabularia sp.) | Brissard et al. (2014) a [19] |
| IFR-OST-03V (France, Villefranche-sur-Mer) | 22 | 38 | 420 (16L:8D) | L1 | ** | Ovatoxin-h, OVTX-a,-b, -c, -d, -e,-f, pPLTX | Seawater (proximity of Stypocaulon sp. and Acetabularia sp.) | Brissard et al. (2015) a [65] |
| 6 strains including IRTA-SMM-12-62 (Spain, South Catalonia, Ebro River Delta) | 24 | 36 | 100 (12L:12D) | ** | ** | OVTX-a,-b, -c, -d, -e,-g, IsobPLTX Total toxin content = 50–250 | Jania rubens | García-Altares et al. (2015) a [99] |
| CBA29-2012 (Italy, Quarto dei Mille Genoa) | 20 ± 0.5 | ** | 85–135 (16L:8D) | F/2 | ** | OVTX-a = 33.5, OVTX-d+e = 9, pPLTX = 1.5. Total toxin content in cell pellets = 44Toxic to Artemia salina nauplii | ** | Giussani et al. (2015) a [82] |
| OOBZT14 (Tunisia, Bizerte Bay) | 25 | 36 | 80 (12L:12D) | ENSW | 0.59 | OVTX-a = 15.56–18.7, OVTX-b = 3.4–4.6 | Cymodocea nodosa | This study a |
| ATLANTIC WATERS | ||||||||
| Isolate 538 (Carribean Sea, Leeward Islands, IIe St. Barthelemy, Port de Gustavia) | 28 | 35 | 300 ft-c (12L:12D) | GPM | ** | Mouse bioassay: lack of water or lipid soluble ciguatera toxins in O. ovata extracts | Tide-pool | Besada et al. (1982) b [137] |
| IEO-OS06BR, IEO-OS15BR (Brazil, Rio de Janeiro) | 17 ± 1 | ** | 100 (14L:10D) | K, F/20, F/2 | ** | Presence of palytoxin Hemolytic activity on human erythrocytes | Rhodophyceae Phaeophyceae | Penna et al. (2005) b [20] |
| LCA-B7 (Brazil, Rio de Janeiro, Armação dos Búzios) | 24 ± 2 | ** | 60 (12L:12D) | L2/2 | 0.1 | OVTX-a = 78–171, OVTX-b = 87–205, OVTX-c = 3–37, OVTX-d+e = 5–55 Hemolytic activity on rabbit erythrocytes | Sargassum vulgare bed | Nascimento et al. (2012) a [89] |
| LCA-E7 (Brazil, Rio de Janeiro, Armação dos Búzios) | 24 ± 2 | ** | 60 (12L:12D) | L2/2 | 0.15 | OvTx-a = 20–71, OvTx-b = 23–77, OvTx-c = 4–30, OvTx-d+e = 3–80 and pPLTX = ND-0.62 Hemolytic activity on rabbit erythrocytes | Sargassum vulgare bed | Nascimento et al. (2012) a [89] |
| Dn145EHU, Dn146EHU, Dn147EHU (Portugal, Lagos) | 20 | ** | 80 (12L:12D) | F/4 | ** | ** | Seawater Macroalgae | David et al. (2013) a [138] |
| PACIFIC WATERS | ||||||||
| ** (Japan, Okinawa, Ishigaki Island) | 25 | ** | 4000–8000 lx (18L:6D) | S-PES | ** | Butanol soluble fraction toxic to mice Hemolytic activity on mouse blood cells No effects on killifish | Turbinaria ornata Amphiroa sp. | Nakajima et al. (1981) b [53] |
| CAWD174 (Cook Islands, south coast of Rarotonga) | ** | ** | ** | F/2 | ** | Palytoxin-equivalents = 1.18 Not Toxic to miceNegatif haemolysis neutralisation assay | Halimeda sp. | Rhodes et al. (2010) b [105] |
| s0715, s0662 (Japan, Kochi, Subogata and Tei) | 25 | ** | 100 (12L:12D) | PES, F/2, IMK | ** | Toxic to mice | Tricleocarpa Pterocladiella, Dictyota | Sato et al. (2011) a [102] |
| s0662 (Japan, Kochi, Tei) | 25 | 30.8 | 90–100 (12L:12D) | F/2, IMK, PES, SWM3 | 0.181–0.866 | ** | ** | Yamaguchi et al. (2012) a [62] |
| s0662 (Japan, Kochi, Tei) | 24–30 | 31 | 140 (12L:12D) | F/10 | ≈0.22–0.5 | Haemolytic activity on horse blood cells only during the decaying phases 25 °C = 0.70 ± 0.15 SnE cell−1 27 °C = 0.46 ± 0.01 SnE cell−1 (ng saponin equivalent per cell) | ** | Vidyarathna and Granéli (2012) b [103] |
| JHAOS5, JHWOS13 (Korea, Jeju Island) | 20 | 30 | 180 (12L:12D) | IMK, F/2 | 0.15–0.25 | Strain JHAOS5: Supressed the growth of HL-60 cells (=human promyelocytic leukemia tumor cell line) | Sand, Macroalgae | Shah et al. (2014) b [139] |
| INDIAN WATERS | ||||||||
| P-0117, P-0128 (Reunion Island, East coast, West Indian Ocean) | 26 | ** | 20–40 (12L:12D) | F/2 | ** | Haemolytic analysis (sheep blood):no palytoxin-like activity | Actinotrichia fragilis, Turbinaria conoides, Jania sp., Galaxaura sp | Carnicer et al. (2015) a [106] |
| INDO-PACIFIC WATERS | ||||||||
| 20 strains (Peninsula and East of Malaysia) | 26–27 | 32 | 30 (12L:12D) | ES-DK | ** | 4 strains/20 toxic to Artemia franciscana, 16 strains not toxic | Sargassum spp. Padina spp. Seagrasses Dead corals | Mohammad Noor et al. (2007) b [101] |
| TD7OS, TF5OS (Gulf of Thailand and Andaman Sea) | 25 | 31 ± 1 | ** (12L:12D) | IMK/2 | ** | Both strains did not cause the death of mice, abnormal behavior observed. | Padina spp. Sargassum spp | Tawong et al. (2014) a [104] |
| Strain and Sampling Location | Temperature (°C) | Salinity | Irradiance (µmol photons.m−2·s−1) and L:D Cycle (h) | Culture Medium | Growth Rate (d−1) | Toxicity: Detected Toxins (pg·cell−1), Hemolytic Activity, Toxicity to Mice and Other Organisms | Isolated from | Reference |
|---|---|---|---|---|---|---|---|---|
| MEDITERRANEAN WATERS | ||||||||
| MARS1 (France, Marseille) | 20 | ** | 40 (12L:12D) | F/2 | ** | OA = 1.9, DTX-1 = 0.8 | ** | Barbier et al. (1999) [35] |
| Several strains including KC2, KC6, KC45, KC49, KC60 (Greece, North Aegean coasts) | 19 ± 1 | ** | 70 (14L:10D) | F/2 | ** | PP2AIA (Protein phosphatase type 2A inhibition assay):Estimated OA equivalents > 0.50–10.23 KC2, KC6: Toxic to Artemia nauplii | Macrophytes | Aligizaki et al. (2009) [87] |
| ** (Italy, Adriatic Sea, lagoon of Goro) | 20 | 25 | 90 (16L:8D) | F/2 | 0.22,0.23 | OA = 6.69–12.50/15.8, DTX-1 = 0.12–0.39 | ** | Vanucci et al. (2010) [79] |
| PLBZT14 (Tunisia, Bizerte Bay) | 25 | 36 | 80 (12L:12D) | ENSW | 0.33 | OA = 7.13–28.33, DTX-1 = 2.23–7.4 | Cymodocea nodosa | This study |
| ATLANTIC WATERS | ||||||||
| 5 strains (Spain, Vigo) | ** | ** | ** | ** | ** | OA = 5–24.5, DTX-1 = 6–14.3 | ** | Lee et al. (1989) [31] |
| PL100A (USA, Florida, Knight Key) | ≈20–40 Opt = 26 | ≈20–40 Opt = 30 | 120–4400 µW·cm−2 Opt = 4000 (14L:10D) | K | 0.3,0.47,0.62 | ** | Heterosiphonia gibbesii | Morton and Norris (1990) [92] |
| PL1V (Spain, Atlantic region) | ** | ** | ** | ** | ** | Toxic (unique indication) | ** | Faust (1991) [140] |
| ** (Canada, Nova Scotia, Mahone Bay) | 20 | ** | 150 (16L:8D) | F/2 | ** | OA and DTX-1 = equal proportions = 25 ng·mL−1 of culture | Seawater | Marr et al. (1992) [32] |
| PL2V (Spain, Vigo) | 20 | 35.5 ± 0.5 | 24 (12L:12D) | K | 0.092 | OA, DTX-1 = 10–15% Total toxin content = 4.35–7.67 | ** | Morlaix and Lassus (1992) [91] |
| PL100A (USA, Florida, Knight Key) | ≈19–35 Opt = 27 | ≈20–43 Opt = 30 | 1500–5500 µW·cm−2 Opt = 4500 (14L:10D) | K | µmax≈0.3–0.56 | ** | Heterosiphonia gibbesii | Morton et al. (1992) [98] |
| ** (Canada, Nova Scotia, Mahone Bay ) | 5,10,15,20,25 | ** | 150 (16L:8D) | F/2 | 0.1–0.7 | OA+DTX-1 = 1.4–8.0 OA:DTX-1 = 1.37 ± 0.23 | Seawater | Jackson et al. (1993) [44] |
| ** (USA, Florida, Dry Tortugas) | 26 | ** | 150 (16L:8D) | K | 0.16–0.75 | OA = 7.5–14.2 DTX-1 = trace concentrations | ** | Tomas and Baden (1993) [93] |
| PL2V (Spain, Vigo) | 20 | ** | 40 (12L:12D) | F/2 | ** | OA = 14.3, DTX-1 = 2.7 | ** | Barbier et al. (1999) [35] |
| Isolate 712 (Spain, Vigo) | 20 | ** | 10 nmol photons·m−2·s−1 (12L:12D) | PES | ** | ** | ** | Zhou and Fritz (1994) [141] |
| ** (Canada, Nova Scotia, Mahone Bay) | 18 ± 1 | 32 | 90 ± 5 (14L:10D) | L1 | 0.1–0.15 | DTX-4 = 1.8–7.8, OA = 0.37–6.6, DTX-1 = 0.04–2.6, OA-D8 = 0.02–1.5 fmol cell−1 | ** | Pan et al. (1999) [94] |
| 19 strains (Spain, Ria of Vigo and Pontevedra) | 19 ± 1 | ** | 60–70 (14L:10D) | K | 0.06–0.14 | OA = 0.19–12.87, OA ester = 0.77–17.51, DTX-1 = 0–12.45, DTX-2 = 0–1.14, DTX-2 ester = 0–1.60 | Macroalgae | Bravo et al. (2001) [39] |
| 20 strains (United Kingdom, Fleet lagoon) | 15,17 | ** | 70,90 (12L:12D; 16L:8D) | L-2 | Strain 2.9a: 0.11 | OA = 0.42–17.13, DTX-1 = 0.41–11.29; DTX-4, DTX-4+O and DTX-4+ CH2+2O detected | Seawater, seaweeds, eelgrass | Nascimento et al. (2005) [40] |
| ** (Cuba, NW Havana city) | 22 ± 1 | ** | Fluorescent lamp of 40 W (12L:12D) | K | ** | Cultured cells: DTX-1 = 7.15 Natural cells: DTX-1 = 4.2 | Padina sp | Delgado et al. (2005) [107] |
| IO66-01 (Portugal, Lisbon Bay) | 19 ± 1 | 35 | 40 (14L:10D) | F/2-Si | 0.49 | Total OA = 8.8–41.0 and DTX-1 = 2.5–12.0 OA-D6, OA-D8, OA-D9 esters detected | Seawater | Vale et al. (2009) [96] |
| CCAP1136/11 (Spain, Ria de Vigo) | 20 | 38 | 35 (16L: 8D) | F/2 | 0.11–0.22 | OA = 0.10–1.25 (Day1–15), Maximum OA = 11.27 ± 3.30 (Day 34) | ** | Varkitzi et al. (2010) [95] |
| Dn35EHU, Dn37EHU, Dn38EHU (Spain, S-E Bay of Biscay) | 17-,22 | 30,35 | 60 (12L:12D) | F/2 | ** | Toxic to Artemia franciscana nauplii (mortality of 86.9% after 24 h) | Macroalgae Seawater | Laza-Martinez et al. (2011) [58] |
| PACIFIC WATERS | ||||||||
| ** (Japan, Okinawa, Ishigaki Island) | 25 | ** | 4000–8000 lx (18L:6D) | S-PES | ** | Ether and Butanol soluble fractions toxic to mice Hemolytic activity on mouse blood cells. No effects on killifish | Turbinaria ornata and Amphiroa sp | Nakajima et al. (1981) [53] |
| ** (Tahiti Island) | 25 | ** | 4000–8000 lx (18L:6D) | S-ES-1 | ** | OA (= PLT2) = 40 mg·10−10·cells Toxic to mice (minimum lethal dose = 200 µg·kg−1) | ** | Murakami et al. (1982) [30] |
| ** (Japan, Okinawa) | ** | ** | ** | ** | ** | OA = 26, DTX-1 = 13 | ** | Lee et al. (1989) [31] |
| OK-8510, OK-8603A, OK-8603B (Japan, Okinawa) | ** | ** | ** | ** | ** | Toxic (unique indication) Toxic (unique indication) | ** | Faust (1991) [140] Faust (1991) [140] |
| SP-8708A, SP-8708D (Saipan Island) | ||||||||
| 17 clones (Australia, N/S/S-E Heron Island) | 28 | ** | 52 (16L:8D) | K | 0.2–0.35 | OA = 1.31–5.88 ≈ Methyl-okadaic acid = 4.0–12.0 | Macroalgae (Phaeophytes, Rhodophytes) | Morton and Tindall (1995) [34] |
| ** (New Zealand, Northland, Rangaunu Harbour ) | 18 ± 1 | ** | 100 (14L:10D) | GP | ** | OA = 6.3 ± 1 Toxic to Artemia salina (50 cells = 50% death response in 24 h, 200 cells = 50% death response in 20 h) | Sediments | Rhodes and Syhre (1995) [33] |
| ** (Japan, Sanriku coast) | 15,20,25 | ** | 170 (14L:10D) | T1 | >0.2 | OA = 0.3 to 1.3 | Sargassum confusum Carpopeltis flabellate | Koike et al. (1998) [142] |
| P6 (New Caledonia) | 25–29 | 30–34 | 50–90 (12L:12D) | F10k | 0.27 | OA = 1.1–15, 7-deoxy-okadaic acid = 0.2–1.5 Inhibition of PP2A activity | ** | Holmes et al. (2001) [90] |
| PL01 (Taiwan) | 25 | ** | ** (14L:10D) | K –ES | ** | 4-hydroxyprorocentrolide, 14-O-acetyl-4-hydroxyprorocentrolide | Seaweeds | Lu et al. (2001) [110] |
| PRL-1 (Mexico, Gulf of California, El Pardito) | 22 ± 1 | ** | 4 × 20 W fluorescent lamps (12L:12D) | ES-SI | 0.107 | OA, DTX-1, (OA:DTX1) = (1:2) OA+DTX-1 = 5.2 (HPLC-MS) Total toxin content = 19 (mouse bioassay) Toxic to mice, to Artemia franciscana larvae and to the yeast Debaryomyces hansenii | Rocky substrate | Heredia-Tapia et al. (2002) [48] |
| PL021117001 (Taiwan, Northern coast) | 25 | ** | ** (16L:8D) | K-ES | ** | Prorocentin, OA Inhibitory activity of Prorocentin against human colon adenocarcinoma and human malignant melanoma | ** | Lu et al. (2005) [42] |
| ** (Southern China, Hainan Island, Coast of Sanya) | 25 | ** | 70 (12L:12D) | K | ** | OA, two diol esters (OA-D10a and OA-D10b) | Macrophytes | Li et al. (2012) [111] |
| INDIAN WATERS | ||||||||
| 8 strains (La Reunion, Mayotte, Europa, and Mauritius Islands) | 26 | ** | 90 (12L:12D) | PPES | ** | OA = 128.3–6261.3 ng·mg−1 crude extract Inhibitory effect on PP2A Cytotoxic activity on FR3T3 fibroblasts | ** | Bouaicha et al. (2001) [36] |
| INDO- PACIFIC WATERS | ||||||||
| 3 strains (Peninsula and East of Malaysia) | 26–27 | 32 | 30 (12L:12D) | ES-DK | ** | High toxicity to Artemia franciscana larvae | Sargassum spp. Padina spp. Dead corals | Mohammad Noor et al. (2007) [101] |
| Strain and Sampling Location | Temperature (°C) | Salinity | Irradiance (µmol photons·m−2·s−1) and L:D Cycle (h) | Culture Medium | Growth Rate (d−1) | Toxicity: Detected Compounds, Hemolytic Activity, Toxicity to Mice and Other Organisms | Isolated from | Reference |
|---|---|---|---|---|---|---|---|---|
| MEDITERRANEAN SEA | ||||||||
| CNR-CMA4 (Italy, Ionian Sea, Taranto) | 17 ± 1 | ** | 100 (14L:10D) | K, F/20, F/2 | ** | ** | Seawater Rhodophyceae Phaeophyceae | Penna et al. (2005) a [20] (MI) |
| CNR-CMB2 (Italy, Tyrrhenian Sea, Ganzirri) | - | - | - | - | - | - | - | - |
| IEO-CM6V (Spain, Almeria) | - | - | - | - | - | - | - | - |
| SZN-CM43 (Italy, Tyrrhenian Sea, Napoli) | - | - | - | - | - | - | - | - |
| ** (Greece, North Aegean Sea, Thermaikos Gulf) | 23 | ≈33 | 60–70 (14L:10D) | L1 | ** | ** | Seawater | Dolapsakis et al. (2006) a [143] (MI) |
| ** (Greece, North Aegean coasts) | 19 ± 1 | ** | 70 (14L:10D) | F/2, K | ** | ** | Macrophytes | Aligizaki and Nikolaidis (2006) a [77] |
| ** (Italy, Northern Ionian Sea, Mar Piccolo, Mar Grande and Lido Bruno) | 24 ± 2 | 37 | 100 (12L:12D) | F/2 | ** | Live cells: no effects on P. lividus embryonic development Cell lysate: inhibition of P. lividus embryos development, low hemolytic activity on human erythrocytes, not toxic to Artemia salina nauplii | Seawater Rocks scraping | Pagliara and Caroppo (2012) b [54] |
| CMBZT14 (Tunisia, Bizerte Bay) | 25 | 36 | 80 (12L:12D) | ENSW | 0.35 | Unknown molecule: Chromatographic peak at 5.6 min with a mass m/z = 1061.768 | Cymodocea nodosa | This study a (MI) |
| ATLANTIC WATERS | ||||||||
| Clones 542, 543 (Carribean Sea, Leeward Islands, IIe St. Barthelemy, Port de Gustavia) | 28 | 35 | 300 ft-c (12L:12D) | GPM | ** | Mouse bioassay: lack of water or lipid soluble ciguatera toxins in C. monotis extracts | Tide-pool | Besada et al. (1982) a [137] |
| ** (Central America, Twin Cays, Belize) | 23 ± 0.5 | 36 | 30–90 (12L:12D) | Erdschreiber | Dt = 3–4 days | ** | Floating detritus Surface sediment | Faust (1992) a [97] |
| CM300A (USA, Florida, Knight Key) | ≈21–35, Opt = 29 °C | ≈23–43, Opt = 33 | 1500–5500, Opt = 5300 µW·cm−2 (14L:10D) | K | µmax ≈ 0.2–0.6 | ** | Heterosiphonia gibbesii | Morton et al. (1992) c [98] |
| CCMP304 (Spain, Ria de Vigo) | 5,10,15,20,25,30,35 | 18–37 | 100 (14L:10D) | GP | ** | Not toxic to mice | Macroalgae Sediments | Rhodes et al. (2000) a [55] |
| IEO-CM2V (Spain, Vigo) CCMP1345 (USA, Florida) | 17 ± 1 | ** | 100 (14L:10D) | K, F/20, F/2 | ** | No hemolytic activity on human erythrocytes | Seawater Rhodophyceae Phaeophyceae | Penna et al. (2005) a [20] (MI) |
| IEO-CM3V (Spain, Vigo) NICMM-RIKZ3, NICMM-RIKZ4 (North Sea, Netherlands, Yerseke) | 17 ± 1 | ** | 100 (14L:10D) | K, F/20, F/2 | ** | ** | Seawater Rhodophyceae Phaeophyceae | Penna et al. (2005) a [20] (MI) |
| Dn23EHU,Dn24EHU,Dn25EHU,Dn26EHU (Spain, S-E Bay of Biscay) | 17–22 | 30–35 | 60 (12L:12D) | F/2 | ** | Not toxic to Artemia franciscana nauplii | Seawater Macroalgae | Laza-Martinez et al. (2011) a [58] (MI) |
| 32 strains (Atlantic coast of the Iberian Peninsula) | 20 | ** | 80 (12L:12D) | F/2 | ** | ** | Seawater Macroalgae | David et al. (2014) a [144] (MI) |
| PACIFIC WATERS | ||||||||
| ** (Japan, Okinawa, Ishigaki Island) | 25 | ** | 4000–8000 lx (18L:6D) | S-PES | ** | Not toxic to mice Hemolytic activity on mouse blood cells No effects on killifish | Turbinaria ornata Amphiroa sp | Nakajima et al. (1981) a [53] |
| ** (Japan, Okinawa, Coast of Motobu) | 23–28 | ** | 1500–3000 Lx (18L:6D) | S-ES-1 | ** | Ceramide with a 2-hydroxy-15-methyl-3-octadecenoyl moiety | ** | Tanaka et al. (1998) a [116] |
| ** (Australia, Queensland, Platypus Bay) | 25 | ** | 50–60 (12L:12D) | F10K | ** | Cooliatoxin: a monosulfated polyether toxin (m/z = 1061.5) Butanol soluble fraction lethal to mice (LD50 = 1 mg·kg−1 in mice) | Cladophora sp | Holmes et al. (1995) d [52] |
| CAWD39 (New Zealand, Northland, Ninety Mile Beach) | 20,25/18 Opt = 25 | 15–43 Opt>28 | 100 (14L:10D) | GP | 25 °C:Dt = 4 days | Toxic to larvae of Artemia salina and Haliotis virginea | Foliose red Landsburgia quercifolia | Rhodes and Thomas (1997) c [56] |
| CAWD39 (New Zealand, Northland, Ninety Mile Beach) | 5,10,15,20,25,30,35 | 18–37 | 100 (14L:10D) | GP | ** | Not Toxic to mice Two analogs of unknown polyether compounds detected | Macroalgae Sediments | Rhodes et al. (2000) c [55] |
| CAWD77 (New Zealand, Northland, Rangiputa) | 5,10,15,20,25,30,35 | 18–37 | 100 (14L:10D) | GP | ** | Toxic to mice Cytotoxic | Macroalgae Sediments | Rhodes et al. (2000) c [55] |
| CMLHT01 (South China Sea, Hainan island) | ** | ** | ** | ** | ** | Cooliatin = dioxocyclononane (C15H22O5) | Seaweeds | Liang et al. (2009) a [117] |
| CAWD151 (Cook Islands, Rarotongan lagoons) | 25 | ** | 80 (14L:10D) | F/2 | ** | Low toxicity to mice | Halimeda sp. | Rhodes et al. (2010) c [114] (MI) |
| ** (Coast of Vietnam) | 26 | 32 | 25 (12L:12D) | T | ** | ** | Macroalgae Seagrasses | Ho and Nguyen (2014) a [145] |
References
- Hallegraeff, G.M. Ocean climate change, phytoplankton community responses, and harmful algal blooms: A formidable predictive challenge. J. Phycol. 2010, 46, 220–235. [Google Scholar] [CrossRef]
- Aissaoui, A.; Turki, S.; Ben Hassine, O.K. Occurrence of harmful dinoflagellates in the Punic harbors of Carthage (Gulf of Tunis, Tunisia) and their correlations with the physicochemical parameters. Bull. Inst. Natn. Scien. Tech. Merde Salammbô 2012, 39, 127–140. [Google Scholar]
- Accoroni, S.; Romagnoli, T.; Colombo, F.; Pennesi, C.; Di Camillo, C.G.; Marini, M.; Battocchi, C.; Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; et al. Ostreopsis cf. ovata bloom in the northern Adriatic Sea during summer 2009: Ecology, molecular characterization and toxin profile. Mar. Pollut. Bull. 2011, 62, 2512–2519. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.M.; Andersen, P.; Bricelj, V.M.; Cullen, J.J.; Rensel, J.E. Monitoring and Management Strategies for Harmful Algal Blooms in Coastal Waters; Asia Pacific Economic Program: Singapore; Intergovernmental Oceanographic Commission: Paris, France, 2001; p. 268. [Google Scholar]
- Burkholder, J.M. Implications of harmful microalgae and heterotrophic dinoflagellates in management of sustainable marine fisheries. Ecol. Appl. 1998, 8, S37–S62. [Google Scholar] [CrossRef]
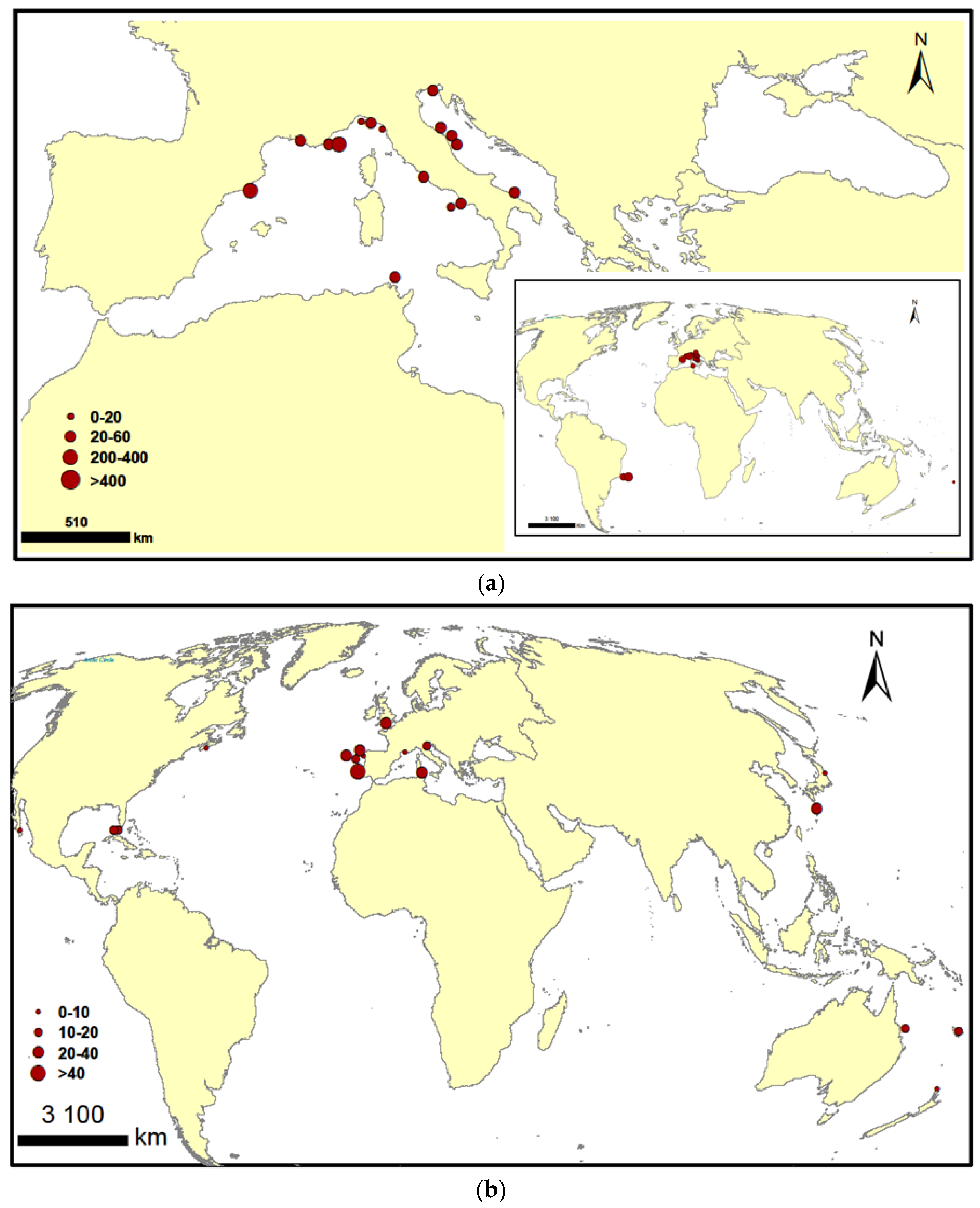
- Aligizaki, K. Spread of potentially toxic benthic dinoflagellates in the Mediterranean Sea: A response to climate change? In Phytoplankton Responses to Mediterranean Environmental Changes; CIESM Workshop Monographs N°40; CIESM Publisher: Monaco; Tunis, Tunisia, 2009; pp. 57–61. [Google Scholar]
- Shears, N.T.; Ross, P.M. Blooms of benthic dinoflagellates of the genus Ostreopsis; an increasing and ecologically important phenomenon on temperate reefs in New Zealand and worldwide. Harmful Algae 2009, 8, 916–925. [Google Scholar] [CrossRef]
- Ingarao, C.; Pagliani, T. Harmful Algae along Abruzzo coast from 2007 to 2010: Correlations with environmental factors and new reports. In Proceedings of the 14th International Conference on Harmful Algae, Crete, Greece, 1–5 November 2010.
- Granéli, E.; Vidyarathna, N.K.; Funari, E.; Cumaranatunga, P.R.T.; Scenati, R. Can increases in temperature stimulate blooms of the toxic benthic dinoflagellate Ostreopsis ovata? Harmful Algae 2011, 10, 98–115. [Google Scholar] [CrossRef]
- Wells, M.L.; Trainer, V.L.; Smayda, T.J.; Karlson, B.S.O.; Trick, C.G.; Kudela, R.M.; Ishikawa, A.; Bernard, S.; Wulff, A.; Anderson, D.M.; et al. Harmful algal blooms and climate change: Learning from the past and present to forecast the future. Harmful Algae 2015, 49, 68–93. [Google Scholar] [CrossRef] [PubMed]
- Laabir, M.; Collos, Y.; Masseret, E.; Grzebyk, D.; Abadie, E.; Savart, V.; Sibat, M.; Amzil, Z. Influence of Environmental Factors on the Paralytic Shellfish Toxin Content and Profile of Alexandrium catenella (Dinophyceae) Isolated from the Mediterranean Sea. Mar. Drugs 2013, 11, 1583–1601. [Google Scholar] [CrossRef] [PubMed]
- Blanfuné, A.; Boudouresque, C.F.; Grossel, H.; Thibaut, T. Distribution and abundance of Ostreopsis spp. and associated species (Dinophyceae) in the northwestern Mediterranean: The region and the macroalgal substrate matter. Environ. Sci. Pollut. Res. 2015, 22, 12332–12346. [Google Scholar] [CrossRef] [PubMed]
- Fukuyo, Y. Taxonomical study on benthic dinoflagellates collected in coral reefs. Nippon Suisan Gakkaishi 1981, 47, 967–978. [Google Scholar] [CrossRef]
- Mangialajo, L.; Ganzin, N.; Accoroni, S.; Asnaghi, V.; Blanfuné, A.; Cabrini, M.; Cattaneo-Vietti, R.; Chavanon, F.; Chiantore, M.; Cohu, S.; et al. Trends in Ostreopsis proliferation along the Northern Mediterranean coasts. Toxicon 2011, 57, 408–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bravo, I.; Vila, M.; Casabianca, S.; Rodriguez, F.; Rial, P.; Riobó, P.; Penna, A. Life cycle stages of the benthic palytoxin-producing dinoflagellate Ostreopsis cf. ovata (Dinophyceae). Harmful Algae 2012, 18, 24–34. [Google Scholar] [CrossRef] [Green Version]
- Brescianini, C.; Grillo, C.; Melchiorre, N.; Bertolotto, R.; Ferrari, A.; Vivaldi, B.; Icardi, G.; Gramaccioni, L.; Funari, E.; Scardala, S. Ostreopsis ovata algal blooms affecting human health in Genova, Italy, 2005 and 2006. Eurosurveillance 2006, 11, e060907.3. [Google Scholar] [PubMed]
- Durando, P.; Ansaldi, F.; Oreste, P.; Moscatelli, P.; Marensi, L.; Grillo, C.; Gasparini, R.; Icardi, G. Ostreopsis ovata and human health: Epidemiological and clinical features of respiratory syndrome outbreaks from a two year syndromic surveillance, 2005−2006, in northwest Italy. Eurosurveillance 2007, 12, e070607. [Google Scholar] [PubMed]
- Ciminiello, P.; Dell’Aversano, C.; Iacovo, ED.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Benedettini, G.; Onorari, M.; Serena, F.; Battocchi, C.; et al. First finding of Ostreopsis cf. ovata toxins in marine aerosols. Environ. Sci. Technol. 2014, 48, 3532–3540. [Google Scholar] [CrossRef] [PubMed]
- Brissard, C.; Herrenknecht, C.; Sechet, V.; Herve, F.; Pisapia, F.; Harcouet, J.; Lemee, R.; Chomerat, N.; Hess, P.; Amzil, Z. Complex toxin profile of French Mediterranean Ostreopsis cf. ovata strains, seafood accumulation and ovatoxins prepurification. Mar. Drugs 2014, 12, 2851–2876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penna, A.; Vila, M.; Fraga, S.; Giacobbe, M.G.; Andreoni, F.; Riobó, P.; Vernesi, C. Characterization of Ostreopsis and Coolia (Dinophyceae) isolates in the Western Mediterranean Sea based on morphology, toxicity and internal transcribed spacer 5.8S rDNA sequences. J. Phycol. 2005, 41, 212–225. [Google Scholar] [CrossRef]
- Riobó, P.; Paz, B.; Franco, J.M. Analysis of palytoxin- like in Ostreopsis cultures by liquid chromatography with precolumn derivatization and fluorescence detection. Anal. Chim. Acta 2006, 566, 217–223. [Google Scholar] [CrossRef]
- Guerrini, F.; Pezzolesi, L.; Feller, A.; Riccardi, M.; Ciminiello, P.; Dell’Aversano, C.; Tartaglione, L.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; et al. Comparative growth and toxin profile of cultured Ostreopsis ovata from the Tyrrhenian and Adriatic Seas. Toxicon 2010, 55, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Honsell, G.; Bonifacio, A.; de Bortoli, M.; Penna, A.; Battocchi, C.; Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Sosa, S.; Yasumoto, T.; et al. New insights on cytological and metabolic features of Ostreopsis cf. ovata Fukuyo (Dinophyceae): A multidisciplinary approach. PLoS ONE 2013, 8, e57291. [Google Scholar] [CrossRef] [PubMed]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Grillo, C.; Melchiorre, N. Putative palytoxin and its new analogue, ovatoxin-a, in Ostreopsis ovata collected along the Ligurian coasts during the 2006 toxic outbreak. J. Am. Soc. Mass Spectrom. 2008, 19, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Grauso, L.; Tartaglione, L.; Guerrini, F.; Pistocchi, R. Complex palytoxin-like profile of Ostreopsis ovata. Identification of four new ovatoxins by high-resolution liquid chromatography/mass spectrometry. Rapid Commun. Mass Spectrom. 2010, 24, 2735–2744. [Google Scholar] [CrossRef] [PubMed]
- Honsell, G.; de Bortoli, M.; Boscolo, S.; Dell’Aversano, C.; Tartaglione, L.; Battocchi, C.; Penna, A.; Berti, F.; Fontanive, G.; Poli, M.; et al. Ostreopsis cf. ovata (Dinophyceae): Toxin content, immunocytochemistry and ultrastructure. Eur. J. Phycol. 2011, 46, 70. [Google Scholar] [CrossRef]
- Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Battocchi, C.; Crinelli, R.; Carloni, E.; Magnani, M.; et al. Unique toxin profile of a Mediterranean Ostreopsis cf. ovata strain: HR LC-MSn characterization of ovatoxin-f, a new palytoxin congener. Chem. Res. Toxicol. 2012, 25, 1243–1252. [Google Scholar] [CrossRef] [PubMed]
- Rossi, R.; Castellano, V.; Scalco, E.; Serpe, L.; Zingone, A.; Soprano, V. New palytoxin-like molecules in Mediterranean Ostreopsis cf. ovata (dinoflagellates) and in Palythoa tuberculosa detected by liquid chromatography–electrospray ionization time-of-flight mass spectrometry. Toxicon 2010, 56, 1381–1387. [Google Scholar] [CrossRef] [PubMed]
- Ehrenberg, C. Über das leuchten und über neue mikroskopische leuchtthiere des mittelmeeres. In Monatsberichte der Königlichen Preuß; Akademie der Wissenschaften zu Berlin: Berlin, Germany, 1860; pp. 791–793. [Google Scholar]
- Murakami, Y.; Oshima, Y.; Yasumoto, T. Identification of okadaic acid as a toxic component of a marine dinoflagellate Prorocentrum lima. Nippon Suisan Gakkaishi 1982, 48, 69–72. [Google Scholar] [CrossRef]
- Lee, J.S.; lgarashi, T.; Fraga, S.; Dahl, E.; Hovgaard, P.; Yasumoto, T. Determination of diarrhetic shellfish toxins in various dinoflagellate species. J. Appl. Phycol. 1989, 1, 147–152. [Google Scholar] [CrossRef]
- Marr, J.C.; Jackson, A.E.; McLachlan, J.L. Occurence of Prorocentrum lima, a DSP toxin-producing species the Atlantic coast of Canada. J. Appl. Phycol. 1992, 4, 17–24. [Google Scholar] [CrossRef]
- Rhodes, L.L.; Syhre, M. Okadaic acid production by a New Zealand Prorocentrum lima isolate. N. Z. J. Mar. Freshw. Res. 1995, 29, 367–370. [Google Scholar] [CrossRef]
- Morton, S.L.; Tindall, D.R. Morphological and biochemical variability of the toxic dinoflagellate Prorocentrum lima isolated from three sites at Heron Island, Australia. J. Phycol. 1995, 31, 914–921. [Google Scholar] [CrossRef]
- Barbier, M.; Amzil, Z.; Mondeguer, F.; Bhaud, Y.; Soyer-Gobillard, M.-O.; Lassus, P. Okadaic acid and PP2A cellular immunolocalization in Prorocentrum lima (Dinophyceae). Phycologia 1999, 38, 41–46. [Google Scholar] [CrossRef]
- Bouaicha, N.; Chezeau, A.; Turquet, J.; Quod, J.P.; Puiseux-Dao, S. Morphological and toxicological variability of Prorocentrum lima clones isolated from four locations in the South-West Indian Ocean. Toxicon 2001, 39, 1195–1202. [Google Scholar] [CrossRef]
- Hu, T.; de Freitas, A.S.W.; Doyle, J.; Jackson, D.; Marr, J.; Nixon, E.; Pleasance, S.; Quiliam, M.A.; Walter, J.A.; Wright, J.L.C. New DSP toxins derivatives isolated from toxic mussels and the dinoflagellate P. lima and P. concavum. In Toxic Phytoplankton Blooms in the Sea; Smayda, T., Shimizu, Y., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 507–512. [Google Scholar]
- Hu, T.; Curtis, J.M.; Walter, J.A.; Wright, J.C. Identification of DTX-4, a new water-soluble phosphatase inhibitor from the toxic dinoflagellate Prorocentrum lima. J. Chem. Soc. Chem. Commun. 1995, 597–599. [Google Scholar] [CrossRef]
- Bravo, I.; Fernandez, M.L.; Ramilo, I.; Martinez, A. Toxin composition of the toxic dinoflagellate Prorocentrum lima isolated from different locations along the Galician coast (NW Spain). Toxicon 2001, 39, 1537–1545. [Google Scholar] [CrossRef]
- Nascimento, S.M.; Purdie, D.A.; Morris, S. Morphology, toxin composition and pigment content of Prorocentrum lima strains isolated from a coastal lagoon in Southern UK. Toxicon 2005, 45, 633–649. [Google Scholar] [CrossRef] [PubMed]
- Torigoe, K.; Murata, M.; Yasumoto, T. Prorocentrolide, a toxic nitrogenous macrocycle from a marine dinoflagellate, Prorocentrum lima. J. Am. Chem. Soc. 1988, 110, 7876–7877. [Google Scholar] [CrossRef]
- Lu, C.K.; Chou, H.N.; Lee, C.K.; Lee, T.H. Prorocentin, a new polyketide from the marine dinoflagellate Prorocentrum lima. Org. Lett. 2005, 7, 3893–3896. [Google Scholar] [CrossRef] [PubMed]
- Fattorusso, E.; Ciminiello, P.; Costantino, V.; Magno, S.; Mangoni, A.; Milandri, A.; Poletti, R.; Pompei, M.; Viviani, R. Okadaic acid in mussels of Adriatic Sea. Mar. Pollut. Bull. 1992, 24, 234–237. [Google Scholar] [CrossRef]
- Jackson, A.E.; Marr, J.C.; McLachalan, J.L. Production of diarrhetic shellfish toxins by an isolate of Prorocentrum lima from Nova Scotia, Canada. In Toxic Phytoplankton Blooms in the Sea; Smayda, T., Shimizu, Y., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 513–518. [Google Scholar]
- Draisci, R.; Croci, L.; Giannetti, L.; Cozzi, L.; Lucentini, L.; de Medici, D.; Stacchini, A. Comparison of mouse bioassay, HPLC and enzyme immunoassay methods for determining diarrhetic shellfish poisoning toxins in mussels. Toxicon 1994, 32, 1379–1384. [Google Scholar] [CrossRef]
- Lawrence, J.E.; Bauder, A.G.; Quilliam, M.A.; Cembella, A.D. Prorocentrum lima: A putative link to diarrhetic shellfish poisoning in Nova Scotia, Canada. In Harmful Algae; Reguera, B., Blanco, J., Fernandez, M.L., Wyatt, T., Eds.; Xunta de Galicia and Intergovernmental Oceanographic Commission of UNESCO: Santiago de Compostela, Spain, 1998; pp. 78–79. [Google Scholar]
- Pavela-Vrančič, M.; Meštrović, V.; Marasović, I.; Gillman, M.; Furey, A.; James, K.J. DSP toxin profile in the coastal waters of the central Adriatic Sea. Toxicon 2002, 40, 1601–1607. [Google Scholar] [CrossRef]
- Heredia-Tapia, A.; Arredondo-Vega, B.O.; Nuñez-Vázquez, E.J.; Yasumoto, T.; Yasuda, M.; Ochoa, J.L. Isolation of Prorocentrum lima (Syn Exuviella lima) and diarrhetic shellfish poisoning (DSP) risk assessment in the Gulf of California, Mexico. Toxicon 2002, 40, 1121–1127. [Google Scholar] [CrossRef]
- Alexander, J.; Auðunsson, G.A.; Benford, D.; Cockburn, A.; Cravedi, J.; Dogliotti, E.; di Domenico, A.; Fernández-Cruz, M.L.; Fink-Gremmels, J.; Fürst, P.; et al. Marine biotoxins in shellfish—Okadaic acid and analogues. Scientific Opinion of the Panel on Contaminants in the Food chain. EFSA J. 2008, 589, 1–62. [Google Scholar]
- Meunier, A. Microplankton de la Mer Flamande Pt. 3 Les Péridiniens; Mémoires Musée Royal d’Histoire Naturelle de Belgique; Bruxelles Hayez: Sint-Jans-Molenbeek, Belgium, 1919; Volume 8, pp. 3–116. [Google Scholar]
- Mohammad-Noor, N.; Moestrup, Ø.; Lundholm, N.; Fraga, S.; Adam, A.; Holmes, M.J.; Saleh, E. Autecology and phylogeny of Coolia tropicalis and Coolia malayensis (Dinophyceae), with emphasis on taxonomy of C. tropicalis based on light microscopy, scanning electron microscopy, and LSU rDNA. J. Phycol. 2013, 49, 536–545. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.J.; Lewis, R.J.; Jones, A.; Hokama, A.M.; Hoy, W. Cooliatoxin, the first toxin from Coolia monotis (Dinophyceae). Nat. Toxins 1995, 3, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, I.; Oshima, Y.; Yasumoto, T. Toxicity of benthic dinoflagellates in Okinawa. Nippon Suisan Gakkaishi 1981, 47, 1029–1033. [Google Scholar] [CrossRef]
- Pagliara, P.; Caroppo, C. Toxicity assessment of Amphidinium carterae, Coolia cfr. monotis and Ostreopsis cfr. ovata (Dinophyta) isolated from the northern Ionian Sea (Mediterranean Sea). Toxicon 2012, 60, 1203–1214. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, L.L.; Adamson, J.; Suzuki, T.; Briggs, L.; Garthwaite, I. Toxic epiphytic marine dinoflagellates Ostreopsis siamensis and Coolia monotis (Dinophyceae) in New Zealand. N. Z. J. Mar. Freshw. Res. 2000, 34, 371–384. [Google Scholar] [CrossRef]
- Rhodes, L.L.; Thomas, A.E. Coolia monotis (Dinophyceae): A toxic epiphytic microalgal species found in New Zealand. N. Z. J. Mar. Freshw. Res. 1997, 31, 139–141. [Google Scholar] [CrossRef]
- Rhodes, L.; Smith, K.; Gimenez Papiol, G.; Adamson, J.; Harwood, T.; Munday, R. Epiphytic dinoflagellates in sub-tropical New Zealand, in particular the genus Coolia Meunier. Harmful Algae 2014, 34, 36–41. [Google Scholar] [CrossRef]
- Laza-Martinez, A.; Orive, E.; Miguel, I. Morphological and genetic characterization of benthic dinoflagellates of the genera Coolia, Ostreopsis, and Prorocentrum from the south eastern Bay of Biscay. Eur. J. Phycol. 2011, 46, 45–65. [Google Scholar] [CrossRef]
- Nagahama, Y.; Murray, S.; Tomaru, A.; Fukuyo, Y. Species boundaries in the toxic dinoflagellate Prorocentrum lima (Dinophyceae, Prorocentrales), based on morphological and phylogenetic characters. J. Phycol. 2011, 47, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Parsons, M.L.; Aligizaki, K.; Bottein, M.Y.D.; Fraga, S.; Morton, S.L.; Penna, A.; Rhodes, L. Gambierdiscus and Ostreopsis: Reassessment of the state of knowledge of their taxonomy, geography, ecophysiology, and toxicology. Harmful Algae 2012, 14, 107–129. [Google Scholar] [CrossRef]
- Sechet, V.; Sibat, M.; Chomerat, N.; Nezan, E.; Grossel, H.; Lehebel-Peron, J.B.; Jauffrais, T.; Ganzin, N.; Marco-Miralles, F.; Lemee, R.; et al. Ostreopsis cf. ovata in the French Mediterranean coast: Molecular characterisation and toxin profile. Cryptogam. Algol. 2012, 33, 89–98. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Tanimoto, Y.; Yoshimatsu, T.; Sato, S.; Nishimura, T.; Uehara, K.; Adachi, M. Culture method and growth characteristics of marine benthic dinoflagellate Ostreopsis spp. isolated from Japanese coastal waters. Fish. Sci. 2012, 78, 993–1000. [Google Scholar] [CrossRef]
- Hoppenrath, M.; Chomérat, N.; Horiguchi, T.; Schweikert, M.; Nagahama, Y.; Murray, S. Taxonomy and phylogeny of the benthic Prorocentrum species (Dinophyceae)—A proposal and review. Harmful Algae 2013, 27, 1–28. [Google Scholar] [CrossRef]
- Karafas, S.; York, R.; Tomas, C. Morphological and genetic analysis of the Coolia monotis species complex with the introduction of two new species, Coolia santacroce sp. nov. and Coolia palmyrensis sp. nov. (Dinophyceae). Harmful Algae 2015, 46, 18–33. [Google Scholar] [CrossRef]
- Brissard, C.; Hervé, F.; Sibat, M.; Séchet, V.; Hess, P.; Amzil, Z.; Herrenknecht, C. Characterization of ovatoxin-h, a new ovatoxin analog, and evaluation of chromatographic columns for ovatoxin analysis and purification. J. Chromatogr. A 2015, 1388, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Turki, S.; El Abed, A. On the presence of potentially toxic algae in the lagoons of Tunisia. Harmful Algae News 2001, 22, 10. [Google Scholar]
- Turki, S. Distribution of toxic dinoflagellates along the leaves of seagrass Posidonia oceanica and Cymodocea nodosa from the Gulf of Tunis. Cah. Biol. Mar. 2005, 46, 29–34. [Google Scholar]
- Armi, Z.; Turki, S.; Trabelsi, E.; Maiz, N.B. First recorded proliferation of Coolia monotis (Meunier, 1919) in the North Lake of Tunis (Tunisia) correlation with environmental factors. Environ. Monit. Assess. 2010, 164, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Sahraoui, I.; Bouchouicha, D.; Mabrouk, H.H.; Hlaili, A.S. Driving factors of the potentially toxic and harmful species of Prorocentrum Ehrenberg in a semi-enclosed Mediterranean lagoon (Tunisia, SW Mediterranean). Mediterr. Mar. Sci. 2013, 14, 353–362. [Google Scholar] [CrossRef]
- Aissaoui, A.; Dhib, A.; Reguera, B.; Ben Hassine, O.K.; Turki, S.; Aleya, L. First evidence of cell deformation occurrence during a Dinophysis bloom along the shores of the Gulf of Tunis (SW Mediterranean Sea). Harmful Algae 2014, 39, 191–201. [Google Scholar] [CrossRef]
- Mabrouk, L.; Ben Brahim, M.; Hamza, A.; Mahfoudhi, M.; Bradai, M.N. A comparison of abundance and diversity of epiphytic microalgal assemblages on the leaves of the seagrasses Posidonia oceanica (L.) and Cymodocea nodosa (Ucria) Asch in Eastern Tunisia. J. Mar. Biol. 2014, 2014. [Google Scholar] [CrossRef]
- Ben Brahim, M.; Feki, M.; Bouain, A. Occurrences of the toxic dinoflagellate Ostreopsis ovata in relation with environmental factors in Kerkennah Island (Southern coast of Tunisia). J. Coast. Life Med. 2015, 3, 596–599. [Google Scholar] [CrossRef]
- Dhib, A.; Fertouna-Bellakhal, M.; Turki, S.; Aleya, L. Harmful planktonic and epiphytic microalgae in a Mediterranean Lagoon: The contribution of the macrophyte Ruppia cirrhosa to microalgae dissemination. Harmful Algae 2015, 45, 1–13. [Google Scholar] [CrossRef]
- Ismael, A.; Halim, Y. Potentially harmful Ostreopsis spp. in the coastal waters of Alexandria—Egypt. Mediterr. Mar. Sci. 2012, 13, 208–212. [Google Scholar] [CrossRef]
- Ismeal, A.A. First record of Coolia monotis Meunier along the Alexandrium Coast—Egypt. Egypt. J. Aquat. Res. 2014, 40, 19–25. [Google Scholar] [CrossRef]
- Illoul, H.; Rodríguez, F.; Vila, M.; Adjas, N.; Aït Younes, A.; Bournissa, M.; Moroghli, A.; Marouf, N.; Rabia, S.; Lala, F.; et al. The genus Ostreopsis along the Algerian coastal waters (SW Mediterranean Sea) associated with a human respiratory intoxication episode. Cryptogam. Algol. 2012, 33, 209–216. [Google Scholar] [CrossRef]
- Aligizaki, K.; Nikolaidis, G. The presence of the potentially toxic genera Ostreopsis and Coolia (Dinophyceae) in the North Aegean Sea, Greece. Harmful Algae 2006, 5, 717–730. [Google Scholar] [CrossRef]
- Accoroni, S.; Romagnoli, T.; Pichierri, S.; Totti, C. New insights on the life cycle stages of the toxic benthic dinoflagellate Ostreopsis cf. ovata. Harmful Algae 2014, 34, 7–16. [Google Scholar] [CrossRef]
- Vanucci, S.; Guerrini, F.; Milandri, A.; Pistocchi, R. Effects of different levels of N- and P-deficiency on cell yield, okadaic acid, DTX-1, protein and carbohydrate dynamics in the benthic dinoflagellate Prorocentrum lima. Harmful Algae 2010, 9, 590–599. [Google Scholar] [CrossRef]
- Reynolds, C.S. Variability in the provision and function of mucilage in phytoplankton: Facultative responses to the environment. Hydrobiologia 2007, 578, 37–45. [Google Scholar] [CrossRef]
- Barone, R. Behavioural trait of Ostreopsis ovata (Dinophyceae) in Mediterranean rock pools: The spider’s strategy. Harmful Algae News 2007, 33, 1–3. [Google Scholar]
- Giussani, V.; Sbrana, F.; Asnaghi, V.; Vassalli, M.; Faimali, M.; Casabianca, S.; Penna, A.; Ciminiello, P.; Dell’Aversano, C.; Tartaglione, L.; et al. Active role of the mucilage in the toxicity mechanism of the harmful benthic dinoflagellate Ostreopsis cf. ovata. Harmful Algae 2015, 44, 46–53. [Google Scholar] [CrossRef]
- Scalco, E.; Brunet, C.; Marino, F.; Rossi, R.; Soprano, V.; Zingone, A.; Montresor, M. Growth and toxicity responses of Mediterranean Ostreopsis cf. ovata to seasonal irradiance and temperature conditions. Harmful Algae 2012, 17, 25–34. [Google Scholar] [CrossRef]
- Accoroni, S.; Romagnoli, T.; Pichierri, S.; Colombo, F.; Totti, C. Morphometric analysis of Ostreopsis cf. ovata cells in relation to environmental conditions and bloom phases. Harmful Algae 2012, 19, 15–22. [Google Scholar] [CrossRef]
- Vanucci, S.; Pezzolesi, L.; Pistocchi, R.; Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Tartaglione, L.; Guerrini, F. Nitrogen and phosphorus limitation effects on cell growth, biovolume, and toxin production in Ostreopsis cf. ovata. Harmful Algae 2012, 15, 78–90. [Google Scholar] [CrossRef]
- Pezzolesi, L.; Pistocchi, R.; Fratangeli, F.; Dell’Aversano, C.; Dello Iacovo, E.; Tartaglione, L. Growth dynamics in relation to the production of the main cellular components in the toxic dinoflagellate Ostreopsis cf. ovata. Harmful Algae 2014, 36, 1–10. [Google Scholar] [CrossRef]
- Aligizaki, K.; Nikolaidis, G.; Katikou, P.; Baxevanis, A.D.; Abatzopoulos, T.J. Potentially toxic epiphytic Prorocentrum (Dinophyceae) species in Greek coastal waters. Harmful Algae 2009, 8, 299–311. [Google Scholar] [CrossRef]
- Pezzolesi, L.; Guerrini, F.; Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Pistocchi, R. Influence of temperature and salinity on Ostreopsis cf. ovata growth and evaluation of toxin content through HR LC-MS and biological assay. Water Res. 2012, 46, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, S.M.; Corrêa, E.V.; Menezes, M.; Varela, D.; Paredes, J.; Morris, S. Growth and toxin profile of Ostreopsis cf. ovata (Dinophyta) from Rio de Janeiro, Brazil. Harmful Algae 2012, 13, 1–9. [Google Scholar] [CrossRef]
- Holmes, M.J.; Lee, F.C.; Khoo, H.W.; Teo, S.L.M. Production of 7-deoxy-okadaic acid by a New Caledonian strain of Prorocentrum lima (Dinophyceae). J. Phycol. 2001, 37, 280–288. [Google Scholar] [CrossRef]
- Morlaix, M.; Lassus, P. Influence de l’azote et du phosphore sur la croissance et la toxicité de Prorocentrum lima (Ehrenberg) Dodge. Cryptogam. Algol. 1992, 13, 187–195. [Google Scholar]
- Morton, S.; Norris, D.R. Role of temperature, salinity, and light intensity on the seasonality of Prorocentrum lima (Ehrenberg) Dodge. In Toxic Marine Phytoplankton; Granéli, E., Sundstrom, B., Edler, L., Anderson, D.M., Eds.; Elsevier: New York, NY, USA, 1990; pp. 201–205. [Google Scholar]
- Tomas, C.R.; Baden, D.G. The influence of phosphorus source on the growth and cellular toxin content of the benthic dinoflagellate Prorocentrum lima. In Toxic Phytoplankton Blooms in the Sea; Smayda, T.J., Shimizu, Y., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 565–570. [Google Scholar]
- Pan, Y.; Cembella, A.D.; Quilliam, M.A. Cell cycle and toxin production in the benthic dinoflagellate Prorocentrum lima. Mar. Biol. 1999, 134, 541–549. [Google Scholar] [CrossRef]
- Varkitzi, I.; Pagou, K.; Granéli, E.; Hatzianestis, I.; Pyrgaki, C.; Pavlidou, A.; Montesanto, B.; Economou-Amilli, A. Unbalanced N:P ratios and nutrient stress controlling growth and toxin production of the harmful dinoflagellate Prorocentrum lima. Harmful Algae 2010, 9, 304–311. [Google Scholar] [CrossRef]
- Vale, P.; Veloso, V.; Amorim, A. Toxin composition of Prorocentrum lima strain isolated from the Portuguese coast. Toxicon 2009, 54, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Faust, M.A. Observation on the morphology and sexual reproduction of Coolia monotis (Dinophyceae). J. Phycol. 1992, 28, 94–104. [Google Scholar]
- Morton, S.L.; Norris, D.R.; Bomber, J.W. Effect of temperature, salinity and light-intensity on the growth and seasonality of toxic dinoflagellates associated with ciguatera. J. Exp. Mar. Biol. Ecol. 1992, 157, 79–90. [Google Scholar] [CrossRef]
- García-Altares, M.; Tartaglione, L.; Dell’Aversano, C.; Carnicer, O.; de la Iglesia, P.; Forino, M.; Diogène, J.; Ciminiello, P. The novel ovatoxin-g and isobaric palytoxin (so far referred to as putative palytoxin) from Ostreopsis cf. ovata (NW Mediterranean Sea): Structural insights by LC-high resolution MSn. Anal. Bioanal. Chem. 2015, 407, 1191–1204. [Google Scholar]
- Pistocchi, R.; Pezzolesi, L.; Guerrini, F.; Vanucci, S.; Dell’Aversano, C.; Fattorusso, E. A review on the effects of environmental conditions on growth and toxin production of Ostreopsis ovata. Toxicon 2011, 57, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Mohammad-Noor, N.; Daugbjerg, N.; Moestrup, Ø.; Anton, A. Marine epibenthic dinoflagellates from Malaysia—A study of live cultures and preserved samples based on light and scanning electron microscopy. Nord. J. Bot. 2007, 24, 629–690. [Google Scholar] [CrossRef]
- Sato, S.; Nishimura, T.; Uehara, K.; Sakanari, H.; Tawong, W.; Hariganeya, N.; Smith, K.; Rhodes, L.; Yasumoto, T.; Taira, Y.; et al. Phylogeography of Ostreopsis along West Pacific Coast, with special reference to a novel clade from Japan. PLoS ONE 2011, 6, e27983. [Google Scholar] [CrossRef] [PubMed]
- Vidyarathna, N.K.; Granéli, E. Influence of temperature on growth, toxicity and carbohydrate production of a Japanese Ostreopsis ovata strain, a toxic-bloom-forming dinoflagellate. Aquat. Microb. Ecol. 2012, 65, 261–270. [Google Scholar] [CrossRef]
- Tawong, W.; Nishimura, T.; Sakanari, H.; Sato, S.; Yamaguchi, H.; Adachi, M. Distribution and molecular phylogeny of the dinoflagellate genus Ostreopsis in Thailand. Harmful Algae 2014, 37, 160–171. [Google Scholar] [CrossRef]
- Rhodes, L.; Smith, K.; Munday, R.; Briggs, L.; Selwood, A.; Adamson, J.; Holland, P. Ostreopsis isolates from the Pacific region. In Proceedings of the 14th International Conference on Harmful Algae, Crete, Greece, 1–5 November 2010; Pagou, P., Hallegraeff, G., Eds.; International Society for the Study of Harmful Algae and IOC of UNESCO: Paris, France, 2013. [Google Scholar]
- Carnicer, O.; Tunin-Ley, A.; Andree, K.B.; Turquet, J.; Diogène, J.; Fernández-Tejedor, M. Contribution to the genus Ostreopsis in Reunion Island (Indian Ocean): Molecular, morphologic and toxicity characterization. Cryptogam. Algol. 2015, 36, 101–119. [Google Scholar] [CrossRef]
- Delgado, G.; Popowski, G.; García, C.; Lagos, N.; Lechuga-Devéze, C. Presence of DSP-toxins in Prorocentrum lima (Ehrenberg) Dodge in Cuba. Rev. Investig. Mar. 2005, 26, 229–234. [Google Scholar]
- McLachlan, J.L.; Marr, J.C.; Colon-Kelly, A.; Adamson, A. Effect of nitrogen concentrations and cold temperatures on DSPtoxin concentrations in the dinofagellate Prorocentrum lima (Prorocentrales, Dinophyceae). Nat. Toxins 1994, 2, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Quilliam, M.A.; Hardstaff, W.R.; Ishida, N.; McLachlan, J.L.; Reeves, A.R.; Rose, N.W.; Windust, A.J. Production of Diarrhetic Shellfish Poisoning (DSP) toxins by Prorocentrum lima in culture and development of analytical methods. In Harmful and Toxic Algal Blooms; Yasumoto, T., Oshima, Y., Fukuyo, Y., Eds.; IOC of UNESCO: Paris, France, 1996; pp. 289–292. [Google Scholar]
- Lu, C.K.; Lee, G.H.; Huang, R.; Chou, H.N. Spiro-prorocentrimine, a novel macrocyclic lactone from a benthic Prorocentrum sp. of Taiwan. Tetrahedron Lett. 2001, 42, 1713–1716. [Google Scholar] [CrossRef]
- Li, J.; Li, M.; Pan, J.; Liang, J.; Zhou, Y.; Wu, J. Identification of the okadaic acid-based toxin profile of a marine dinoflagellate strain Prorocentrum lima by LC–MS/MS and NMR spectroscopic data. J. Sep. Sci. 2012, 35, 782–789. [Google Scholar] [CrossRef] [PubMed]
- Fraga, S.; Penna, A.; Bianconi, I.; Paz, B.; Zapata, M. Coolia canariensis sp. nov. (Dinophyceae), a new nontoxic epiphytic benthic dinoflagellate from the Canary Islands. J. Phycol. 2008, 44, 1060–1070. [Google Scholar] [CrossRef] [PubMed]
- Aligizaki, K.; Katikou, P.; Nikolaidis, G. Toxic benthic dinoflagellates and potential risk in the Mediterranean Sea. In Proceedings of the 7th International Conference on Molluscan Shellfish Safety (ICMSS09), Nantes, France, 14–19 June 2009.
- Rhodes, L.L.; Smith, K.F.; Munday, R.; Selwood, A.I.; McNabb, P.S.; Holland, P.T.; Bottein, M.Y. Toxic dinoflagellates (Dinophyceae) from Rarotonga, Cook Islands. Toxicon 2010, 56, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Momigliano, P.; Sparrow, L.; Blair, D.; Heimann, K. The Diversity of Coolia spp. (Dinophyceae: Ostreopsidaceae) in the Central Great Barrier Reef Region. PLoS ONE 2013, 8, e79278. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, I.; Matsuoka, S.; Murata, M.; Tachibana, K.A. New ceramide with a novel branched-chain fatty acid isolated from the epiphytic dinoflagellate Coolia monotis. J. Nat. Prod. 1998, 61, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.L.; Long, L.J.; Zhang, S.; Wu, J. Cooliatin, an unprecedented natural dioxocyclononane from dinoflagellate Coolia monotis from South China Sea. Chem. Pharm. Bull. 2009, 57, 888–889. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.M.; Alpermann, T.J.; Cembella, A.D.; Collos, Y.; Masseret, E.; Montresor, M. The globally distributed Alexandrium tamarense complex (Dinophyceae) in a Mediterranean lagoon facilitated by semi-multiplex PCR. J. Plankton Res. 2012, 33, 405–414. [Google Scholar]
- Parkhill, J.P.; Cembella, A.D. Effects of salinity, light and inorganic nitrogen on growth and toxigenity of the marine dinoflagellate Alexandrium tamarense from northeastern Canada. J. Plankton Res. 1999, 21, 939–955. [Google Scholar] [CrossRef]
- Grzebyk, D.; Béchemin, C.; Ward, C.; Vérité, C.; Codd, G.; Maestrini, S.Y. Effects of salinity and two coastal waters on the growth and toxin content of the dinoflagellate Alexandrium minutum. J. Plankton Res. 2003, 25, 1185–1199. [Google Scholar] [CrossRef]
- Kim, D.I.; Matsuyama, Y.; Nagasoe, S.; Yamaguch, M.; Yoon, Y.H.; Oshima, Y.; Imada, N.; Honjo, T. Effects of temperature, salinity and irradiance on the growth of the harmful red tide dinoflagellate Cochlodinium polykrikoides Margalef (Dinophyceae). J. Plankton Res. 2004, 26, 61–66. [Google Scholar] [CrossRef]
- Wang, D.Z.; Hsieh, D.P.H. Dynamics of C2 toxin and chlorophyll a formation in the dinoflagellate Alexandrium tamarense during large scale cultivation. Toxicon 2005, 39, 1533–1536. [Google Scholar] [CrossRef]
- Laabir, M.; Jauzein, C.; Genovesi, B.; Masseret, E.; Grzebyk, D.; Cecchi, P.; Vaquer, A.; Perrin, Y.; Collos, Y. Influence of temperature, salinity and irradiance on the growth and cell yield for the harmful red tide dinoflagellate Alexandrium catenella colonising Mediterranean waters. J. Plankton Res. 2011, 33, 1550–1563. [Google Scholar] [CrossRef]
- Band-Schmidt, C.J.; Morquecho, L.; Lechuga-Deveze, C.H.; Anderson, D.M. Effects of growth medium, temperature, salinity and seawater source on the growth of Gymnodinium catenatum (Dinophyceae) from Bahia Concepcíon, Gulf of California, Mexico. J. Plankton Res. 2004, 26, 1459–1470. [Google Scholar] [CrossRef]
- Turki, S.; Dhib, A.; Fertouna-Bellakhal, M.; Frossard, V.; Balti, N.; Kharrat, R.; Aleya, L. Harmful algal blooms (HABs) associated with phycotoxins in shellfish: What can be learned from five years of monitoring in Bizerte Lagoon (Southern Mediterranean Sea)? Ecol. Eng. 2014, 67, 39–47. [Google Scholar] [CrossRef]
- Harrison, P.J.; Waters, R.E.; Taylor, F.J.R. A broad spectrum artificial seawater medium for coastal and open ocean phytoplankton. J. Phycol. 1980, 16, 28–35. [Google Scholar] [CrossRef]
- Fritz, L.; Triemer, R.E. A rapid and simple technique utilizing Calcofluor White M2R for the visualization of dinoflagellate thecal plates. J. Phycol. 1985, 21, 662–664. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritschi, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989; pp. 11–71. [Google Scholar]
- Nézan, E.; Siano, R.; Boulben, S.; Six, C.; Bilien, G.; Cheze, K.; Duval, A.; Le Panse, S.; Quere, J.; Chomerat, N. Genetic diversity of the harmful family Kareniaceae (Gymnodiniales, Dinophyceae) in France, with the description of Karlodinium gentienii sp. nov.: A new potentially toxic dinoflagellate. Harmful Algae 2014, 40, 75–91. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Gascuel, O. A Simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Guillard, R.L.R. Division rates. In Handbook of Phycological Methods: Cultures Methods and Growth Measurements; Stein, J.R., Ed.; Cambridge University Press: Cambridge, UK, 1973; pp. 290–311. [Google Scholar]
- Bravo, I.; Vila, M.; Magadán, S.; Rial, P.; Rodriguez, F.; Fraga, S.; Franco, J.M.; Riobó, P.; Sala, M.M. Studies on cultures of Ostreopsis cf. ovata:life cycle observations. In Proceedings of the 14th International Conference on Harmful Algae (ICHA14), Crete, Greece, 1–5 November 2010.
- Besada, E.G.; Loeblich, L.A.; Loeblich, A.R., III. Observations on tropical, benthic dinoflagellates from ciguatera-endemic areas: Coolia, Gambierdiscus, and Ostreopsis. Bull. Mar. Sci. 1982, 32, 723–725. [Google Scholar]
- David, H.; Laza-Martínez, A.; Miguel, I.; Orive, E. Ostreopsis cf. siamensis and Ostreopsis cf. ovata from the Atlantic Iberian Peninsula: Morphological and phylogenetic characterization. Harmful Algae 2013, 30, 44–55. [Google Scholar]
- Shah, M.M.R.; Samarakoon, K.W.; Ko, J.Y.; Lakmal, H.H.C.; Lee, J.H.; An, S.J.; Jeon, Y.J.; Lee, J.B. Potentiality of benthic dinoflagellate cultures and screening of their bioactivities in Jeju Island, Korea. Afr. J. Biotechnol. 2014, 13, 792–805. [Google Scholar]
- Faust, M.A. Morphology of ciguatera-causing Prorocentrum lima (Pyrrophyta) from widely differing sites. J. Phycol. 1991, 27, 642–648. [Google Scholar] [CrossRef]
- Zhou, J.; Fritz, L. The PAS/Accumulation Bodies in Prorocentrum lima and Prorocentrum maculosum (Dinophyceae) are Dinoflagellate Lysosomes. J. Phycol. 1994, 30, 39–44. [Google Scholar] [CrossRef]
- Koike, K.; Sato, S.; Yamaji, M.; Nagahama, Y.; Kotaki, Y.; Ogata, T.; Kodama, M. Occurrence of okadaic acidproducing Prorocentrum lima on the Sanriku coast, northern Japan. Toxicon 1998, 36, 2039–2042. [Google Scholar] [CrossRef]
- Dolapsakis, N.P.; Kilpatrick, M.W.; Economou-amilli, A.; Tafas, T. Morphology and rDNA phylogeny of a Mediterranean Coolia monotis (Dinophyceae) strain from Greece. Sci. Mar. 2006, 70, 67–76. [Google Scholar] [CrossRef]
- David, H.; Laza-Martínez, A.; Miguel, I.; Orive, E. Broad distribution of Coolia monotis and restricted distribution of Coolia cf. canariensis (Dinophyceae) in the Atlantic coast of the Iberian Peninsula. Phycologia 2014, 53, 342–352. [Google Scholar] [CrossRef]
- Ho, T.V.; Nguyen, N. Morphology and distribution of the three epiphytic dinoflagellate species Coolia monotis, C. tropicalis, and C. canariensis (Ostreopsidaceae, Gonyaulacales, Dinophyceae) from Vietnamese coastal waters. Ocean Sci. J. 2014, 49, 211–221. [Google Scholar] [CrossRef]














| Growth Phase Measures | Exponential Phase | Stationary Phase | |||
|---|---|---|---|---|---|
| Length (µm) | Width (µm) | Length (µm) | Width (µm) | ||
| O. cf. ovata | Mean | 50.38 | 36.80 | 55.18 | 39.58 |
| SD | 4.36 | 3.33 | 5.25 | 3.61 | |
| Min | 41.85 | 32.92 | 42.90 | 36.12 | |
| Max | 58.51 | 45.28 | 65.01 | 45.88 | |
| P. lima | Mean | 45.69 | 36.00 | 45.45 | 36.04 |
| SD | 1.66 | 1.35 | 1.88 | 1.66 | |
| Min | 42.98 | 34.94 | 42.02 | 33.61 | |
| Max | 48.80 | 37.95 | 49.13 | 37.34 | |
| C. monotis | Mean | 30.65 | 29.23 | 30.66 | 29.35 |
| SD | 1.32 | 1.44 | 2.77 | 3.27 | |
| Min | 28.14 | 27.63 | 23.13 | 22.07 | |
| Max | 33.37 | 32.62 | 35.58 | 36.96 | |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ben-Gharbia, H.; Yahia, O.K.-D.; Amzil, Z.; Chomérat, N.; Abadie, E.; Masseret, E.; Sibat, M.; Zmerli Triki, H.; Nouri, H.; Laabir, M. Toxicity and Growth Assessments of Three Thermophilic Benthic Dinoflagellates (Ostreopsis cf. ovata, Prorocentrum lima and Coolia monotis) Developing in the Southern Mediterranean Basin. Toxins 2016, 8, 297. https://doi.org/10.3390/toxins8100297
Ben-Gharbia H, Yahia OK-D, Amzil Z, Chomérat N, Abadie E, Masseret E, Sibat M, Zmerli Triki H, Nouri H, Laabir M. Toxicity and Growth Assessments of Three Thermophilic Benthic Dinoflagellates (Ostreopsis cf. ovata, Prorocentrum lima and Coolia monotis) Developing in the Southern Mediterranean Basin. Toxins. 2016; 8(10):297. https://doi.org/10.3390/toxins8100297
Chicago/Turabian StyleBen-Gharbia, Hela, Ons Kéfi-Daly Yahia, Zouher Amzil, Nicolas Chomérat, Eric Abadie, Estelle Masseret, Manoella Sibat, Habiba Zmerli Triki, Habiba Nouri, and Mohamed Laabir. 2016. "Toxicity and Growth Assessments of Three Thermophilic Benthic Dinoflagellates (Ostreopsis cf. ovata, Prorocentrum lima and Coolia monotis) Developing in the Southern Mediterranean Basin" Toxins 8, no. 10: 297. https://doi.org/10.3390/toxins8100297
APA StyleBen-Gharbia, H., Yahia, O. K. -D., Amzil, Z., Chomérat, N., Abadie, E., Masseret, E., Sibat, M., Zmerli Triki, H., Nouri, H., & Laabir, M. (2016). Toxicity and Growth Assessments of Three Thermophilic Benthic Dinoflagellates (Ostreopsis cf. ovata, Prorocentrum lima and Coolia monotis) Developing in the Southern Mediterranean Basin. Toxins, 8(10), 297. https://doi.org/10.3390/toxins8100297