
Natural Co-Occurrence of Mycotoxins in Foods and Feeds and Their in vitro Combined Toxicological Effects



Abstract
:1. Introduction
2. Mycotoxin Regulations
2.1. Aflatoxins
2.2. Ochratoxin A
2.3. Fumonisins
2.4. Zearalenone
2.5. Trichothecenes
2.6. Other Regulated Mycotoxins
3. Natural Co-Occurrence of Mycotoxins in Foods and Feeds
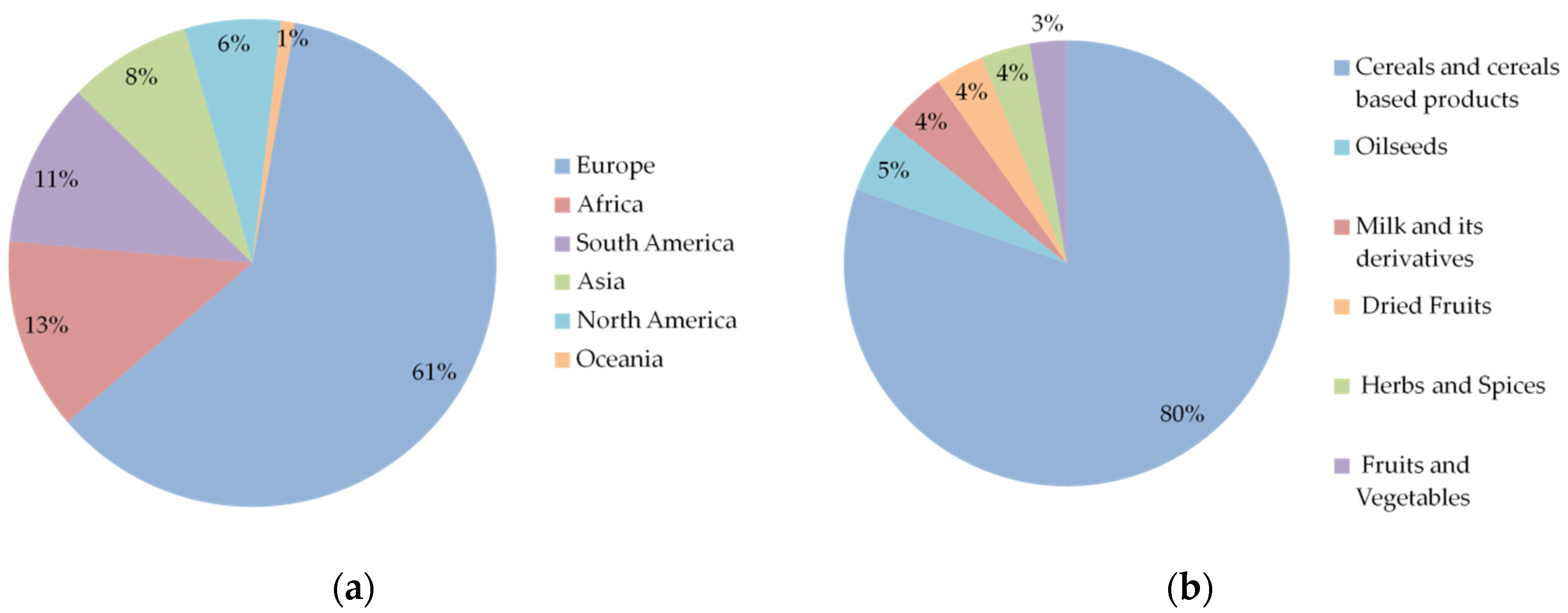
3.1. Results by Commodity Type
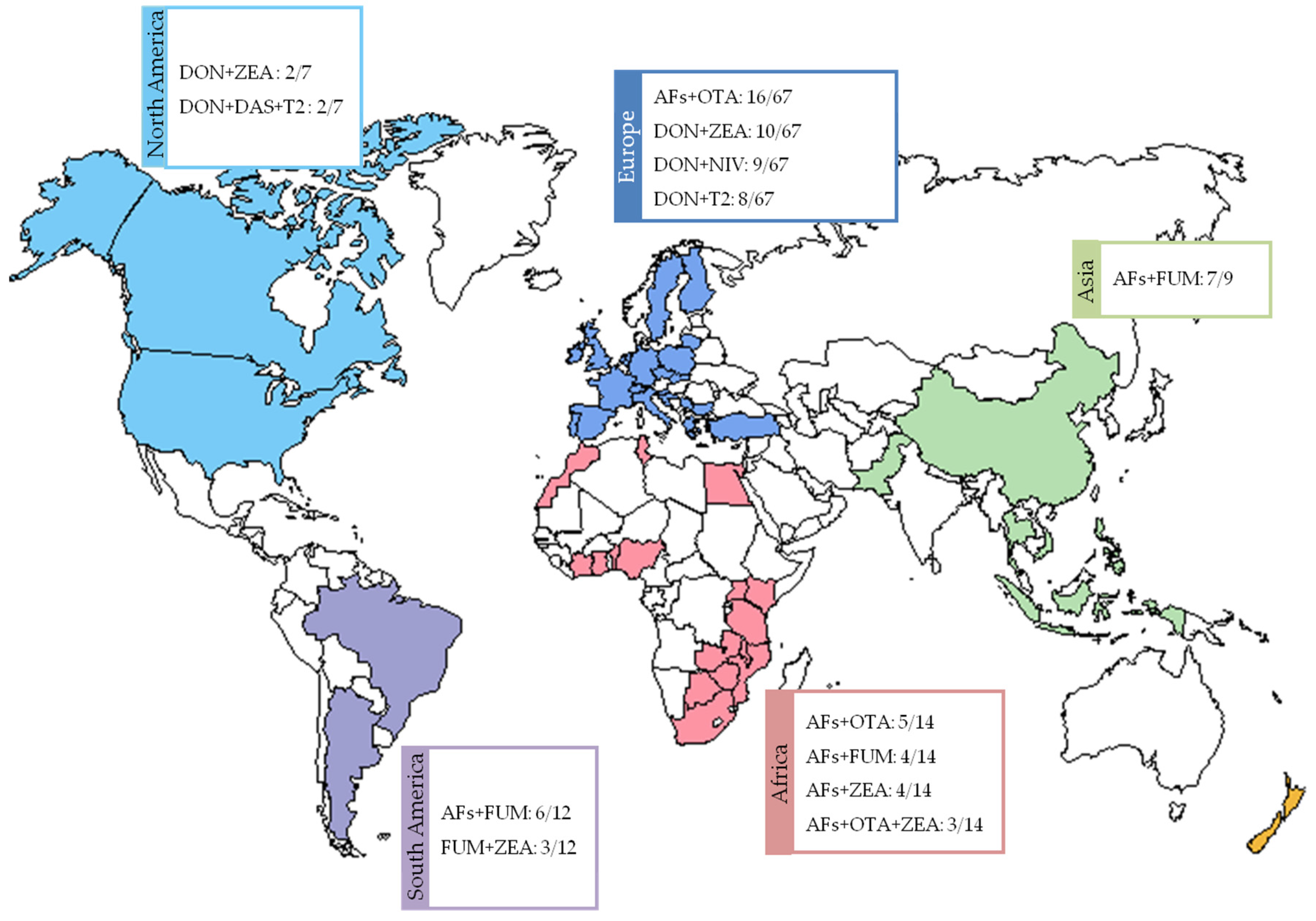
3.2. Results by Region
| 1. AFs FUM | 41. DAS HT2 | 81. BEA ENNs MON |
| 2. DON ZEA | 42. T2 NIV | 82. AFs OTA DON |
| 3. AFs OTA | 43. T2 ZEA | 83. AFs OTA T2 |
| 4. FUM ZEA | 44. NIV BEA | 84. AFs OTA NIV |
| 5. DON NIV | 45. ENNs FUS | 85. AFs OTA FUM |
| 6. DON T2 | 46. AFs DON | 86. AFs FUM T2 |
| 7. DON HT2 | 47. FUM BEA | 87. AFs DON ZEA |
| 8. AFs ZEA | 48. FUM MON | 88. FUM DON NIV |
| 9. FUM DON | 49. OTA NIV | 89. FUM T2 HT2 |
| 10. FUM OTA | 50. DON ADON T2 | 90. FUM BEA OTA |
| 11. DON T2 ZEA | 51. DON DAS HT2 | 91. FUM BEA FUS |
| 12. T2 HT2 | 52. DON T2 NIV | 92. FUM OTA CIT |
| 13. BEA ENNs | 53. DON HT2 FUS-X | 93. DON ADON T2 HT2 |
| 14. AFs OTA ZEA | 54. DON OTA ZEA | 94. DON ADON HT2 ZEA |
| 15. DON ADON NIV | 55. AFs FUM NIV | 95. DON ADON ZEA αZOL |
| 16. DON ADON ZEA | 56. AFs BEA NIV | 96. DON MAS NIV ZEA |
| 17. AFs FUM ZEA | 57. DON ADON T2 NIV | 97. DON DAS T2 HT2 |
| 18. FUM DON ZEA | 58. DON ADON T2 ZEA | 98. DON T2 ZEA αZOL |
| 19. DON ADON | 59. DON T2 HT2 NIV | 99. DON HT2 NIV ZEA |
| 20. DON OTA | 60. DON T2 NIV ZEA | 100. DAS T2 HT2 ZEA |
| 21. FUM NIV | 61. DON ADON T2 HT2 ZEA | 101. T2 HT2 MAS ZEA |
| 22. OTA ZEA | 62. DON DAS | 102. T2 HT2 NIV BEA |
| 23. OTA CIT | 63. DON αZOL | 103. NIV ZEA BEA ENNs |
| 24. DON T2 HT2 | 64. DON ENNs | 104. AFs OTA DON ZEA |
| 25. DON HT2 NIV | 65. NIV HT2 | 105. AFs OTA T2 ZEA |
| 26. DON HT2 ZEA | 66. NIV FUS-X | 106. FUM DON NIV ZEA |
| 27. DON NIV ZEA | 67. NIV ZEA | 107. DON ADON MAS HT2 ZEA |
| 28. FUM ZEA OTA | 68. ZEA ENNs | 108. DON ADON HT2 NIV ZEA |
| 29. DON ADON HT2 NIV | 69. BEA FUS | 109. DON ADON HT2 NIV FUS-X |
| 30. DON T2 HT2 ZEA | 70. FUM T2 | 110. DON DAS T2 HT2 ZEA |
| 31. DON FUS-X | 71. FUM FUS | 111. DON MAS T2 HT2 ZEA |
| 32. ADON ZEA | 72. MYC-A ROQ-C | 112. DON T2 HT2 NIV ZEA |
| 33. AFs NIV | 73. DON DAS T2 | 113. MAS T2 HT2 NIV ZEA |
| 34. DON ADON HT2 | 74. DON NIV FUS-X | 114. AFs FUM OTA DON ZEA |
| 35. DAS T2 HT2 | 75. DON NIV MAS | 115. DON ADON T2 NIV ZEA αZOL |
| 36. AFs FUM DON | 76. DON ZEA αZOL | 116. DON ADON NIV ZEA αZOL βZOL |
| 37. DON ADON NIV ZEA | 77. T2 T2tetraol HT2 | |
| 38. AFs FUM OTA ZEA | 78. T2 HT2 ZEA | |
| 39. DON ADON T2 HT2 NIV | 79. NIV FUS-X BEA | |
| 40. DAS T2 | 80. BEA ENNs FUS |
4. Toxicological Impact of Mycotoxin Interactions
4.1. Characterization of the Different Interactions Between Mycotoxins
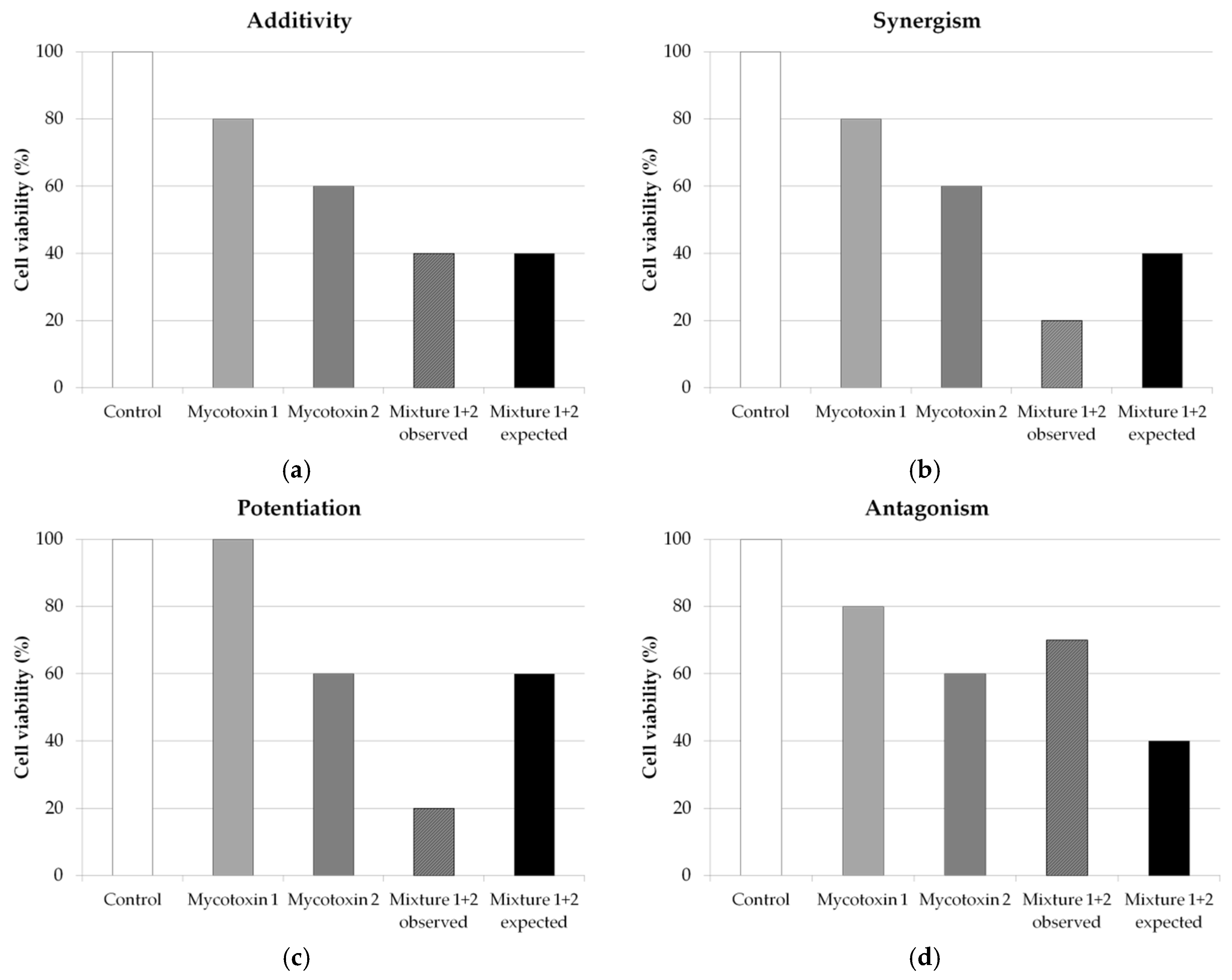
- Additivity is mentioned when the effect of the combination could be calculated as the sum of the individual effects of the two studied toxins (Figure 5a). Thus, additivity is a priori an absence of interaction.
- Synergism is observed when the effect of the mycotoxin combination is greater than expected in comparison to the sum of the individual effects of the two studied mycotoxins (Figure 5b). In the case when one or both of the mycotoxins does not induce effect whereas the combination induces a significant effect, one can speak of potentiation (Figure 5c). However, very few studies use this term to categorize the effect, and most of them use synergism.
- Antagonism is cited when the effect of the mycotoxin combination is lower than expected from the sum of the individual effects of the two studied mycotoxins (Figure 5d). If the effect of the mycotoxin combination mainly reflected the effect of the most toxic mycotoxin, without additional effect of the other mycotoxin, the term “less-than-additive” may be used.
4.2. Main Experimental Designs for Studying Mycotoxin Interactions
4.3. In Vitro Interactions Between Fusariotoxins
5. Conclusion
Acknowledgments
Conflicts of Interest
Abbreviations
| 3-ADON | 3-acetyldeoxynivalenol |
| 15-ADON | 15-acetyldeoxynivalenol |
| α-ZOL | α-zearalenol |
| β-ZOL | β-zearalenol |
| AFs | aflatoxins |
| BEA | beauvericine |
| DAS | diacetoxyscirpenol |
| DON | deoxynivalenol |
| EC | European Commission |
| EFSA | European Food Safety Authority |
| ENA, ENA1,ENB, ENB1 | enniatins A, A1, B, B1 |
| ENs | enniatins |
| FAO | Food and Agriculture Organization |
| FDA | Food and Drug Administration |
| FB1, FB2, FB3 | fumonisin B1, B2, B3 |
| FUM | fumonisins |
| FUS-X | fusarenone-X |
| IC | inhibitory concentration |
| HT-2 | HT-2 toxin |
| MON | moniliformin |
| NIV | nivalenol |
| OTA | ochratoxin A |
| TCTs | trichothecenes |
| T-2 | T-2 toxin |
| ZEA | zearalenone |
| WHO | World Health Organization |
References
- Bryden, W.L. Mycotoxin contamination of the feed supply chain: Implications for animal productivity and feed security. Anim. Feed Sci. Technol. 2012, 173, 134–158. [Google Scholar] [CrossRef]
- Marin, S.; Ramos, A.J.; Cano-Sancho, G.; Sanchis, V. Mycotoxins: Occurrence, toxicology, and exposure assessment. Food Chem. Toxicol. 2013, 60, 218–237. [Google Scholar] [CrossRef] [PubMed]
- Milani, J.M. Ecological conditions affecting mycotoxin production in cereals: A review. Vet. Med. Czech Repub. 2013, 58, 405–411. [Google Scholar]
- Da Rocha, M.E.B.; da Freire, F.C.O.; Maia, F.E.F.; Guedes, M.I.F.; Rondina, D. Mycotoxins and their effects on human and animal health. Food Control 2014, 36, 159–165. [Google Scholar] [CrossRef]
- Bullerman, L.B.; Bianchini, A. Stability of mycotoxins during food processing. Int. J. Food Microbiol. 2007, 119, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Grenier, B.; Oswald, I. Mycotoxin co-contamination of food and feed: meta-analysis of publications describing toxicological interactions. World Mycotoxin J. 2011, 4, 285–313. [Google Scholar] [CrossRef]
- CAST Report. Task Force Report N 139. In Mycotoxins: Risks in plant, animal, and human systems; Richard, J.L.; Payne, G.A. (Eds.) Council for Agricultural Science and Technology: Ames, IA, USA, 2003.
- Binder, E.M.; Tan, L.M.; Chin, L.J.; Handl, J.; Richard, J. Worldwide occurrence of mycotoxins in commodities, feeds and feed ingredients. Anim. Feed Sci. Technol. 2007, 137, 265–282. [Google Scholar] [CrossRef]
- Clarke, R.; Connolly, L.; Frizzell, C.; Elliott, C.T. Cytotoxic assessment of the regulated, co-existing mycotoxins aflatoxin B1, fumonisin B1 and ochratoxin, in single, binary and tertiary mixtures. Toxicon 2014, 90, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Streit, E.; Naehrer, K.; Rodrigues, I.; Schatzmayr, G. Mycotoxin occurrence in feed and feed raw materials worldwide: Long-term analysis with special focus on Europe and Asia. J. Sci. Food Agric. 2013, 93, 2892–2899. [Google Scholar] [CrossRef] [PubMed]
- Jestoi, M. Emerging Fusarium—Mycotoxins fusaproliferin, beauvericin, enniatins, and moniliformin—A review. Crit. Rev. Food Sci. Nutr. 2008, 48, 21–49. [Google Scholar] [CrossRef] [PubMed]
- Grenier, B.; Applegate, T.J. Modulation of intestinal functions following mycotoxin ingestion: Meta-analysis of published experiments in animals. Toxins 2013, 5, 396–430. [Google Scholar] [CrossRef] [PubMed]
- Kadota, T.; Furusawa, H.; Hirano, S.; Tajima, O.; Kamata, Y.; Sugita-Konishi, Y. Comparative study of deoxynivalenol, 3-acetyldeoxynivalenol, and 15-acetyldeoxynivalenol on intestinal transport and IL-8 secretion in the human cell line Caco-2. Toxicol. In Vitro 2013, 27, 1888–1895. [Google Scholar] [CrossRef] [PubMed]
- EC, (European Commission). Commission regulation (EC) No 1881/2006 of 19 December 2006 (consolidated version 2014-07-01) setting maximum levels for certain contaminants in foodstuffs. Available online: http://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2006:364:0005:0024:EN:PDF (accessed on 30 November 2015).
- Van Egmond, H.P.; Jonker, M.A. Worldwide regulations for mycotoxins in food and feed in 2003; Food and Agriculture Organization of the United Nations, 2004. [Google Scholar]
- Barug, D.; van Egmond, H.P.; López-García, R.; van Osenbruggen, W.A.; Visconti, A. (Eds.) Mycotoxins worldwide: Current issues in Africa. In Meeting the mycotoxin menace, Proceedings of the 2nd World Mycotoxin Forum held in Nordwijk, Nordwijk, the Netherlands, 17–18 February 2003; pp. 81–88.
- Berthiller, F.; Crews, C.; Dall’Asta, C.; Saeger, S.D.; Haesaert, G.; Karlovsky, P.; Oswald, I.P.; Seefelder, W.; Speijers, G.; Stroka, J. Masked mycotoxins: A review. Mol. Nutr. Food Res. 2013, 57, 165–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cirlini, M.; Dall’Asta, C.; Galaverna, G. Hyphenated chromatographic techniques for structural characterization and determination of masked mycotoxins. J. Chromatogr. A 2012, 1255, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, I.; Naehrer, K. A three-year survey on the worldwide occurrence of mycotoxins in feedstuffs and feed. Toxins 2012, 4, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Giuberti, G.; Frisvad, J.C.; Bertuzzi, T.; Nielsen, K.F. Review on mycotoxin issues in ruminants: Occurrence in forages, effects of mycotoxin ingestion on health status and animal performance and ractical strategies to counteract their negative effects. Toxins 2015, 7, 3057–3111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speijers, G.J.A.; Speijers, M.H.M. Combined toxic effects of mycotoxins. Toxicol. Lett. 2004, 153, 91–98. [Google Scholar] [CrossRef] [PubMed]
- FDA, (U.S. Food and Drug Administration). Guidance for industry: Action levels for poisonous or deleterious substances in human food and animal feed—Aflatoxin. Available online: http://www.fda.gov/Food/GuidanceRegulation/GuidanceDocumentsRegulatoryInformation/ChemicalContaminantsMetalsNaturalToxinsPesticides/ucm077969.htm#afla (accessed on 3 December 2015).
- EC, (European Commission). Directive 2002/32/EC of the European Parliament and of the Council of 7 May 2002 (consolidated version 2013-12-27) on undesirable substances in animal feed. Available online: http://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:02002L0032-20131227 (accessed on 30 November 2015).
- Bosco, F.; Molle, C. Mycotoxins in Food. In Food Industrial Processes - Methods and Equipment; Valdez, B., Ed.; InTech: Torino, Italy, 2012. [Google Scholar]
- Creppy, E.E. Update of survey, regulation and toxic effects of mycotoxins in Europe. Toxicol. Lett. 2002, 127, 19–28. [Google Scholar] [CrossRef]
- Ficheux, A.S.; Sibiril, Y.; le Garrec, R.; Parent-Massin, D. In vitro myelotoxicity assessment of the emerging mycotoxins Beauvericin, Enniatin b and Moniliformin on human hematopoietic progenitors. Toxicon 2012, 59, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Filazi, A.; Sireli, U.T. Occurrence of aflatoxins in food. In Aflatoxins—Recent Advances and Future Prospects; Razzaghi-Abyaneh, M., Ed.; InTech: Ankara, Turkey, 2013. [Google Scholar]
- Glenn, A.E. Mycotoxigenic Fusarium species in animal feed. Anim. Feed Sci. Technol. 2007, 137, 213–240. [Google Scholar] [CrossRef]
- Logrieco, A.; Moretti, A.; Castella, G.; Kostecki, M.; Golinski, P.; Ritieni, A.; Chelkowski, J. Beauvericin production by Fusarium species. Appl. Environ. Microbiol. 1998, 64, 3084–3088. [Google Scholar] [PubMed]
- Oliveira, P.M.; Zannini, E.; Arendt, E.K. Cereal fungal infection, mycotoxins, and lactic acid bacteria mediated bioprotection: From crop farming to cereal products. Food Microbiol. 2014, 37, 78–95. [Google Scholar] [CrossRef] [PubMed]
- Pitt, J.I. Toxigenic fungi and mycotoxins. Br. Med. Bull. 2000, 56, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Yazar, S.; Omurtag, G.Z. Fumonisins, trichothecenes and zearalenone in cereals. Int. J. Mol. Sci. 2008, 9, 2062–2090. [Google Scholar] [CrossRef] [PubMed]
- EC, (European Commission). Commission recommandation of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding. Available online: http://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32006H0576&from=EN (accessed on 30 November 2015).
- FDA, (U.S. Food and Drug Administration). Guidance for industry: Fumonisin levels in human foods and animal feeds; final guidance (June 6, 2000; revised November 9, 2001). Available online: http://www.fda.gov/Food/GuidanceRegulation/GuidanceDocumentsRegulatoryInformation/ChemicalContaminantsMetalsNaturalToxinsPesticides/ucm109231.htm (accessed on 3 December 2015).
- FDA, (U.S. Food and Drug Administration). Guidance for industry and FDA: Advisory levels for deoxynivalenol (DON) in finished wheat products for human consumption and grains and grain by-products used for animal feed (June 29, 2010; Revised July 7, 2010). Available online: http://www.fda.gov/downloads/Food/GuidanceRegulation/UCM217558.pdf (accessed on 3 December 2015).
- FDA, (U.S. Food and Drug Administration). Compliance Policy Guides - CPG Sec.510.150 Apple juice, apple juice concentrates, and apple juice products - adulteration with patulin (October 22, 2001; revised November 29, 2005). Available online: http://www.fda.gov/iceci/compliancemanuals/compliancepolicyguidancemanual/ucm074427.htm (accessed on 3 December 2015).
- Abbas, H.K.; Mirocha, C.J.; Meronuck, R.A.; Pokorny, J.D.; Gould, S.L.; Kommedahl, T. Mycotoxins and Fusarium spp. associated with infected ears of corn in Minnesota. Appl. Environ. Microbiol. 1988, 54, 1930–1933. [Google Scholar] [PubMed]
- Abramson, D.; Mills, J.T.; Marquardt, R.R.; Frohlich, A.A. Mycotoxins in fungal contaminated samples of animal feed from western Canada, 1982–1994. Can. J. Vet. Res. 1997, 61, 49–52. [Google Scholar] [PubMed]
- Abramson, D.; Clear, R.M.; Nowicki, T.W. Fusarium species and trichothecene mycotoxins in suspect samples of 1985 manitoba wheat. Can. J. Plant Sci. 1987, 67, 611–619. [Google Scholar] [CrossRef]
- Ali, N.; Sardjono; Yamashita, A.; Yoshizawa, T. Natural co-occurrence of aflatoxins and Fusarium mycotoxins (fumonisins, deoxynivalenol, nivalenol and zearalenone) in corn from Indonesia. Food Addit. Contam. 1998, 15, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Almeida, I.; Martins, H.M.; Santos, S.; Costa, J.M.; Bernardo, F. Co-occurrence of mycotoxins in swine feed produced in Portugal. Mycotoxin Res. 2011, 27, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Alvito, P.C.; Sizoo, E.A.; Almeida, C.M.M.; van Egmond, H.P. Occurrence of aflatoxins and ochratoxin A in baby foods in Portugal. Food Anal. Methods 2010, 3, 22–30. [Google Scholar] [CrossRef]
- Bankole, S.A.; Mabekoje, O.O. Occurrence of aflatoxins and fumonisins in preharvest maize from south-western Nigeria. Food Addit. Contam. 2004, 21, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Baydar, T.; Engin, A.B.; Girgin, G.; Aydin, S.; Sahin, G. Aflatoxin and ochratoxin in various types of commonly consumed retail ground samples in Ankara, Turkey. Ann. Agric. Environ. Med. AAEM 2005, 12, 193–197. [Google Scholar] [PubMed]
- Bircan, C. Incidence of ochratoxin A in dried fruits and co-occurrence with aflatoxins in dried figs. Food Chem. Toxicol. 2009, 47, 1996–2001. [Google Scholar] [CrossRef] [PubMed]
- Błajet-Kosicka, A.; Twarużek, M.; Kosicki, R.; Sibiorowska, E.; Grajewski, J. Co-occurrence and evaluation of mycotoxins in organic and conventional rye grain and products. Food Control 2014, 38, 61–66. [Google Scholar] [CrossRef]
- Camargos, S.M.; Machinski, M.; Soares, R. Co-occurrence of fumonisins and aflatoxins in freshly harvested Brazilian maize. Trop. Sci. 2001, 41, 182–184. [Google Scholar]
- Cano-Sancho, G.; Ramos, A.J.; Marín, S.; Sanchis, V. Presence and co-occurrence of aflatoxins, deoxynivalenol, fumonisins and zearalenone in gluten-free and ethnic foods. Food Control 2012, 26, 282–286. [Google Scholar] [CrossRef]
- Cano-Sancho, G.; Valle-Algarra, F.M.; Jiménez, M.; Burdaspal, P.; Legarda, T.M.; Ramos, A.J.; Sanchis, V.; Marín, S. Presence of trichothecenes and co-occurrence in cereal-based food from Catalonia (Spain). Food Control 2011, 22, 490–495. [Google Scholar] [CrossRef]
- Castillo, M.-Á.; Montes, R.; Navarro, A.; Segarra, R.; Cuesta, G.; Hernández, E. Occurrence of deoxynivalenol and nivalenol in Spanish corn-based food products. J. Food Compos. Anal. 2008, 21, 423–427. [Google Scholar] [CrossRef]
- Cavaliere, C.; D’Ascenzo, G.; Foglia, P.; Pastorini, E.; Samperi, R.; Laganà, A. Determination of type B trichothecenes and macrocyclic lactone mycotoxins in field contaminated maize. Food Chem. 2005, 92, 559–568. [Google Scholar] [CrossRef]
- Cerveró, M.C.; Castillo, M.A.; Montes, R.; Hernández, E. Determination of trichothecenes, zearalenone and zearalenols in commercially available corn-based foods in Spain. Rev. Iberoam. Micol. 2007, 24, 52–55. [Google Scholar] [CrossRef]
- Chamberlain, W.J.; Bacon, C.W.; Norred, W.P.; Voss, K.A. Levels of fumonisin B1 in corn naturally contaminated with aflatoxins. Food Chem. Toxicol. 1993, 31, 995–998. [Google Scholar] [CrossRef]
- Cunha, S.C.; Fernandes, J.O. Development and validation of a method based on a QuEChERS procedure and heart-cutting GC-MS for determination of five mycotoxins in cereal products. J. Sep. Sci. 2010, 33, 600–609. [Google Scholar] [CrossRef] [PubMed]
- Doko, M.B.; Canet, C.; Brown, N.; Sydenham, E.W.; Mpuchane, S.; Siame, B.A. Natural co-occurrence of fumonisins and zearalenone in cereals and cereal-based foods from Eastern and Southern Africa. J. Agric. Food Chem. 1996, 44, 3240–3243. [Google Scholar] [CrossRef]
- Domijan, A.-M.; Peraica, M.; Cvjetković, B.; Turcin, S.; Jurjević, Z.; Ivić, D. Mould contamination and co-occurrence of mycotoxins in maize grain in Croatia. Acta Pharm. Zagreb Croat. 2005, 55, 349–356. [Google Scholar]
- Domijan, A.-M.; Peraica, M.; Jurjević, Ž.; Ivić, D.; Cvjetković, B. Fumonisin B1, fumonisin B2, zearalenone and ochratoxin A contamination of maize in Croatia. Food Addit. Contam. 2005, 22, 677–680. [Google Scholar] [CrossRef] [PubMed]
- Driehuis, F.; Spanjer, M.C.; Scholten, J.M.; te Giffel, M.C. Occurrence of mycotoxins in feedstuffs of dairy cows and estimation of total dietary intakes. J. Dairy Sci. 2008, 91, 4261–4271. [Google Scholar] [CrossRef] [PubMed]
- Eckard, S.; Wettstein, F.E.; Forrer, H.-R.; Vogelgsang, S. Incidence of Fusarium species and mycotoxins in silage maize. Toxins 2011, 3, 949–967. [Google Scholar] [CrossRef] [PubMed]
- Ediage, E.N.; di Mavungu, J.D.; Monbaliu, S.; Van Peteghem, C.; De Saeger, S. A validated multianalyte LC-MS/MS method for quantification of 25 mycotoxins in cassava flour, peanut cake and maize samples. J. Agric. Food Chem. 2011, 59, 5173–5180. [Google Scholar] [CrossRef] [PubMed]
- El Adlouni, C.; Tozlovanu, M.; Naman, F.; Faid, M.; Pfohl-Leszkowicz, A. Preliminary data on the presence of mycotoxins (ochratoxin A, citrinin and aflatoxin B1) in black table olives “Greek style” of Moroccan origin. Mol. Nutr. Food Res. 2006, 50, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Eskola, M.; Parikka, P.; Rizzo, A. Trichothecenes, ochratoxin A and zearalenone contamination and Fusarium infection in Finnish cereal samples in 1998. Food Addit. Contam. 2001, 18, 707–718. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Tao, B.; Pang, M.; Liu, Y.; Dong, J. Occurrence of major mycotoxins in maize from Hebei Province, China. Front. Agric. China 2011, 5, 497–503. [Google Scholar] [CrossRef]
- Fernández Pinto, V.; Patriarca, A.; Locani, O.; Vaamonde, G. Natural co-occurrence of aflatoxin and cyclopiazonic acid in peanuts grown in Argentina. Food Addit. Contam. 2001, 18, 1017–1020. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, I.; Fernandes, J.O.; Cunha, S.C. Optimization and validation of a method based in a QuEChERS procedure and gas chromatography–mass spectrometry for the determination of multi-mycotoxins in popcorn. Food Control 2012, 27, 188–193. [Google Scholar] [CrossRef]
- Fontaine, K.; Passeró, E.; Vallone, L.; Hymery, N.; Coton, M.; Jany, J.-L.; Mounier, J.; Coton, E. Occurrence of roquefortine C, mycophenolic acid and aflatoxin M1 mycotoxins in blue-veined cheeses. Food Control 2015, 47, 634–640. [Google Scholar] [CrossRef]
- Garrido, C.E.; Hernández Pezzani, C.; Pacin, A. Mycotoxins occurrence in Argentina’s maize (Zea mays L.), from 1999 to 2010. Food Control 2012, 25, 660–665. [Google Scholar] [CrossRef]
- Ghali, R.; Hmaissia-khlifa, K.; Ghorbel, H.; Maaroufi, K.; Hedili, A. Incidence of aflatoxins, ochratoxin A and zearalenone in tunisian foods. Food Control 2008, 19, 921–924. [Google Scholar] [CrossRef]
- Gonzalez, H.H. L.; Martinez, E.J.; Pacin, A.M.; Resnik, S.L.; Sydenham, E.W. Natural co-occurrence of fumonisins, deoxynivalenol, zearalenone and aflatoxins in field trial corn in Argentina. Food Addit. Contam. 1999, 16, 565–569. [Google Scholar] [CrossRef] [PubMed]
- González Pereyra, M.L.; Chiacchiera, S.M.; Rosa, C.A.; Sager, R.; Dalcero, A.M.; Cavaglieri, L. Comparative analysis of the mycobiota and mycotoxins contaminating corn trench silos and silo bags. J. Sci. Food Agric. 2011, 91, 1474–1481. [Google Scholar] [CrossRef] [PubMed]
- Gutema, T.; Munimbazi, C.; Bullerman, L.B. Occurrence of fumonisins and moniliformin in corn and corn-based food products of U.S. origin. J. Food Prot. 2000, 63, 1732–1737. [Google Scholar] [PubMed]
- Hajslova, J.; Lancova, K.; Sehnalova, M.; Krplova, A.; Zachariasova, M.; Moravcova, H.; Nedelnik, J.; Markova, J.; Ehrenbergerova, J. Occurrence of trichothecene mycotoxins in cereals harvested in the Czech Republic. Czech J. Food Sci. - UZPI Czech Repub. 2007, 25, 339–350. [Google Scholar]
- Hasan, H.A.H. Patulin and aflatoxin in brown rot lesion of apple fruits and their regulation. World J. Microbiol. Biotechnol. 2000, 16, 607–612. [Google Scholar] [CrossRef]
- Heperkan, D.; Somuncuoglu, S.; Karbancioglu-Güler, F.; Mecik, N. Natural contamination of cyclopiazonic acid in dried figs and co-occurrence of aflatoxin. Food Control 2012, 23, 82–86. [Google Scholar] [CrossRef]
- Hernández Hierro, J.M.; Garcia-Villanova, R.J.; Rodríguez Torrero, P.; Toruño Fonseca, I.M. Aflatoxins and ochratoxin A in red paprika for retail sale in Spain: Occurrence and evaluation of a simultaneous analytical method. J. Agric. Food Chem. 2008, 56, 751–756. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez-Vea, M.; González-Peñas, E.; Lizarraga, E.; López de Cerain, A. Co-occurrence of aflatoxins, ochratoxin A and zearalenone in barley from a northern region of Spain. Food Chem. 2012, 132, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez-Vea, M.; Martínez, R.; González-Peñas, E.; Lizarraga, E.; López de Cerain, A. Co-occurrence of aflatoxins, ochratoxin A and zearalenone in breakfast cereals from spanish market. Food Control 2011, 22, 1949–1955. [Google Scholar] [CrossRef]
- Jaimez, J.; Fente, C.A.; Franco, C.M.; Cepeda, A.; Vázquez, B.I. A survey of the fungal contamination and presence of ochratoxin A and zearalenone on Spanish feed and raw materials. J. Sci. Food Agric. 2004, 84, 832–840. [Google Scholar] [CrossRef]
- Jakšić, S.; Abramović, B.; Jajić, I.; Baloš, M.Ž.; Mihaljev, Ž.; Despotović, V.; Šojić, D. Co-occurrence of fumonisins and deoxynivalenol in wheat and maize harvested in Serbia. Bull. Environ. Contam. Toxicol. 2012, 89, 615–619. [Google Scholar] [CrossRef] [PubMed]
- Jakšić, S.; Prunic, B.; Milanov, D.; Jajić, I.; Bjelica, L.; Abramovic, B. Fumonisins and co-occurring mycotoxins in north Serbian corn. Zb. Matice Srp. Za Prir. Nauke 2011, 49–59. [Google Scholar] [CrossRef]
- Jestoi, M.; Rokka, M.; Yli-Mattila, T.; Parikka, P.; Rizzo, A.; Peltonen, K. Presence and concentrations of the Fusarium-related mycotoxins beauvericin, enniatins and moniliformin in finnish grain samples. Food Addit. Contam. 2004, 21, 794–802. [Google Scholar] [CrossRef] [PubMed]
- Juan, C.; Mañes, J.; Raiola, A.; Ritieni, A. Evaluation of beauvericin and enniatins in Italian cereal products and multicereal food by liquid chromatography coupled to triple quadrupole mass spectrometry. Food Chem. 2013, 140, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Juan, C.; Ritieni, A.; Mañes, J. Occurrence of Fusarium mycotoxins in Italian cereal and cereal products from organic farming. Food Chem. 2013, 141, 1747–1755. [Google Scholar] [CrossRef] [PubMed]
- Jurjević, Z.; Solfrizzo, M.; Cvjetković, B.; de Girolamo, A.; Visconti, A. Occurrence of beauvericin in corn from Croatia. Food Technol Biotechnol 2002, 40, 91–94. [Google Scholar]
- Jurjević, Ž.; Solfrizzo, M.; Cvjetković, B.; Avantaggiato, G.; Visconti, A. Occurrence of beauvericin in Croatia corn survey from 1996 to 1997. Phytopathology 2000, 90, 40. [Google Scholar]
- Kabak, B. Aflatoxin M1 and ochratoxin A in baby formulae in Turkey: Occurrence and safety evaluation. Food Control 2012, 26, 182–187. [Google Scholar] [CrossRef]
- Kabak, B. Determination of aflatoxins and ochratoxin A in retail cereal products from Turkey by high performance liquid chromatography with fluorescence detection. Food Control 2012, 28, 1–6. [Google Scholar] [CrossRef]
- Kimanya, M.E.; De Meulenaer, B.; Tiisekwa, B.; Ndomondo-Sigonda, M.; Devlieghere, F.; Van Camp, J.; Kolsteren, P. Co-occurrence of fumonisins with aflatoxins in home-stored maize for human consumption in rural villages of Tanzania. Food Addit. Contam. Part Chem. Anal. Control Expo. Risk Assess. 2008, 25, 1353–1364. [Google Scholar] [CrossRef] [PubMed]
- Kirinčič, S.; Škrjanc, B.; Kos, N.; Kozolc, B.; Pirnat, N.; Tavčar-Kalcher, G. Mycotoxins in cereals and cereal products in Slovenia—Official control of foods in the years 2008–2012. Food Control 2015, 50, 157–165. [Google Scholar] [CrossRef]
- Klarić, M.S.; Cvetnić, Z.; Pepeljnjak, S.; Kosalec, I. Co-occurrence of aflatoxins, ochratoxin A, fumonisins, and zearalenone in cereals and feed, determined by competitive direct enzyme-linked immunosorbent assay and thin-layer chromatography. Arh. Hig. Rada Toksikol. 2009, 60, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Kokkonen, M.; Jestoi, M.; Rizzo, A. Determination of selected mycotoxins in mould cheeses with liquid chromatography coupled to tandem with mass spectrometry. Food Addit. Contam. 2005, 22, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Kpodo, K.; Thrane, U.; Hald, B. Fusaria and fumonisins in maize from Ghana and their co-occurrence with aflatoxins. Int. J. Food Microbiol. 2000, 61, 147–157. [Google Scholar] [CrossRef]
- Labuda, R.; Parich, A.; Berthiller, F.; Tančinová, D. Incidence of trichothecenes and zearalenone in poultry feed mixtures from Slovakia. Int. J. Food Microbiol. 2005, 105, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Labuda, R.; Parich, A.; Vekiru, E.; Tancinová, D. Incidence of fumonisins, moniliformin and Fusarium species in poultry feed mixtures from Slovakia. Ann. Agric. Environ. Med. AAEM 2005, 12, 81–86. [Google Scholar] [PubMed]
- Li, F.Q.; Yoshizawa, T.; Kawamura, O.; Luo, X.Y.; Li, Y.W. Aflatoxins and fumonisins in corn from the high-incidence area for human hepatocellular carcinoma in Guangxi, China. J. Agric. Food Chem. 2001, 49, 4122–4126. [Google Scholar] [CrossRef] [PubMed]
- Logrieco, A.; Rizzo, A.; Ferracane, R.; Ritieni, A. Occurrence of beauvericin and enniatins in wheat affected by Fusarium avenaceum head blight. Appl. Environ. Microbiol. 2002, 68, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Mahnine, N.; Meca, G.; Elabidi, A.; Fekhaoui, M.; Saoiabi, A.; Font, G.; Mañes, J.; Zinedine, A. Further data on the levels of emerging Fusarium mycotoxins enniatins (A, A1, B, B1), beauvericin and fusaproliferin in breakfast and infant cereals from Morocco. Food Chem. 2011, 124, 481–485. [Google Scholar] [CrossRef]
- Makun, H.A.; Dutton, M.F.; Njobeh, P.B.; Mwanza, M.; Kabiru, A.Y. Natural multi-occurrence of mycotoxins in rice from Niger State, Nigeria. Mycotoxin Res. 2011, 27, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Mankeviciene, A.; Butkute, B.; Dabkevicius, Z. Peculiarities of cereal grain co-contamination with Fusarium mycotoxins. Zemdirb.-Agric. 2011, 98, 415–420. [Google Scholar]
- Martins, H.M.; Guerra, M.M.; Bernardo, F. Zearalenone, deoxynivalenol and fumonisins in mixed-feed for laying hens. Mycotoxin Res. 2006, 22, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.L.; Gimeno, A.; Martins, H.M.; Bernardo, F. Co-occurrence of patulin and citrinin in Portuguese apples with rotten spots. Food Addit. Contam. 2002, 19, 568–574. [Google Scholar] [CrossRef] [PubMed]
- Molinié, A.; Faucet, V.; Castegnaro, M.; Pfohl-Leszkowicz, A. Analysis of some breakfast cereals on the French market for their contents of ochratoxin A, citrinin and fumonisin B1: Development of a method for simultaneous extraction of ochratoxin A and citrinin. Food Chem. 2005, 92, 391–400. [Google Scholar] [CrossRef]
- Monge, M.P.; Dalcero, A.M.; Magnoli, C.E.; Chiacchiera, S.M. Natural co-occurrence of fungi and mycotoxins in poultry feeds from Entre Ríos, Argentina. Food Addit. Contam. Part B Surveill. 2013, 6, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Montes, R.; Segarra, R.; Castillo, M.-Á. Trichothecenes in breakfast cereals from the Spanish retail market. J. Food Compos. Anal. 2012, 27, 38–44. [Google Scholar] [CrossRef]
- Moreno, E.C.; Garcia, G.T.; Ono, M.A.; Vizoni, É.; Kawamura, O.; Hirooka, E.Y.; Ono, E.Y.S. Co-occurrence of mycotoxins in corn samples from the Northern region of Paraná State, Brazil. Food Chem. 2009, 116, 220–226. [Google Scholar] [CrossRef]
- Müller, H.-M.; Schwadorf, K. Natural occurrence of Fusarium toxins in barley grown in a southwestern area of Germany. Bull. Environ. Contam. Toxicol. 1993, 51, 532–537. [Google Scholar] [CrossRef] [PubMed]
- Müller, H.-M.; Schwadorf, K. A survey of the natural occurrence of Fusarium toxins in wheat grown ina southwestern area of Germany. Mycopathologia 1993, 121, 115–121. [Google Scholar] [CrossRef]
- Oliveira, C.A.; Rosmaninho, J.; Rosim, R. Aflatoxin M 1 and cyclopiazonic acid in fluid milk traded in São Paulo, Brazil. Food Addit. Contam. 2006, 23, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Ono, E.Y.S.; Ono, M.A.; Funo, F.Y.; Medina, A.E.; Oliveira IV, T.C.; Kawamura, O.; Ueno, Y.; Hirooka, E.Y. Evaluation of fumonisin-aflatoxin co-occurrence in Brazilian corn hybrids by ELISA. Food Addit. Contam. 2001, 18, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Ozbey, F.; Kabak, B. Natural co-occurrence of aflatoxins and ochratoxin A in spices. Food Control 2012, 28, 354–361. [Google Scholar] [CrossRef]
- Perkowski, J.; Jeleń, H.; Kiecana, I.; Goliński, P. Natural contamination of spring barley with group A trichothecene mycotoxins in south-eastern Poland. Food Addit. Contam. 1997, 14, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Perkowski, J.; Basiński, T. Natural contamination of oat with group A trichothecene mycotoxins in Poland. Food Addit. Contam. 2002, 19, 478–482. [Google Scholar] [CrossRef] [PubMed]
- Pettersson, H.; Brown, C.; Hauk, J.; Hoth, S.; Meyer, J.; Wessels, D. Survey of T-2 and HT-2 toxins by LC-MS/MS in oats and oat products from European oat mills in 2005-2009. Food Addit. Contam. Part B Surveill. Commun. 2011, 4, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Pfohl-Leszkowicz, A.; Tozlovanu, M.; Manderville, R.; Peraica, M.; Castegnaro, M.; Stefanovic, V. New molecular and field evidences for the implication of mycotoxins but not aristolochic acid in human nephropathy and urinary tract tumor. Mol. Nutr. Food Res. 2007, 51, 1131–1146. [Google Scholar] [CrossRef] [PubMed]
- Raiola, A.; Meca, G.; Mañes, J.; Ritieni, A. Bioaccessibility of deoxynivalenol and its natural co-occurrence with ochratoxin A and aflatoxin B1 in Italian commercial pasta. Food Chem. Toxicol. 2012, 50, 280–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritieni, A.; Moretti, A.; Logrieco, A.; Bottalico, A.; Randazzo, G.; Monti, S.M.; Ferracane, R.; Fogliano, V. Occurrence of Fusaproliferin, Fumonisin B1, and Beauvericin in Maize from Italy. J. Agric. Food Chem. 1997, 45, 4011–4016. [Google Scholar] [CrossRef]
- Rocha, L.O.; Nakai, V.K.; Braghini, R.; Reis, T.A.; Kobashigawa, E.; Corrêa, B. Mycoflora and co-occurrence of fumonisins and aflatoxins in freshly harvested corn in different regions of Brazil. Int. J. Mol. Sci. 2009, 10, 5090–5103. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Carrasco, Y.; Moltó, J.C.; Berrada, H.; Mañes, J. A survey of trichothecenes, zearalenone and patulin in milled grain-based products using GC–MS/MS. Food Chem. 2014, 146, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Carrasco, Y.; Ruiz, M.J.; Font, G.; Berrada, H. Exposure estimates to Fusarium mycotoxins through cereals intake. Chemosphere 2013, 93, 2297–2303. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Carrasco, Y.; Berrada, H.; Font, G.; Mañes, J. Multi-mycotoxin analysis in wheat semolina using an acetonitrile-based extraction procedure and gas chromatography–tandem mass spectrometry. J. Chromatogr. A 2012, 1270, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Sangare-Tigori, B.; Moukha, S.; Kouadio, H.J.; Betbeder, A.-M.; Dano, D.S.; Creppy, E.E. Co-occurrence of aflatoxin B1, fumonisin B1, ochratoxin A and zearalenone in cereals and peanuts from Côte d’Ivoire. Food Addit. Contam. 2006, 23, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.; Marín, S.; Sanchis, V.; Ramos, A.J. Co-occurrence of aflatoxins, ochratoxin A and zearalenone in Capsicum powder samples available on the Spanish market. Food Chem. 2010, 122, 826–830. [Google Scholar] [CrossRef]
- Schollenberger, M.; Müller, H.-M.; Rüfle, M.; Terry-Jara, H.; Suchy, S.; Plank, S.; Drochner, W. Natural occurrence of Fusarium toxins in soy food marketed in Germany. Int. J. Food Microbiol. 2007, 113, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Schollenberger, M.; Müller, H.-M.; Rüfle, M.; Suchy, S.; Planck, S.; Drochner, W. Survey of Fusarium toxins in foodstuffs of plant origin marketed in Germany. Int. J. Food Microbiol. 2005, 97, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Schollenberger, M.; Suchy, S.; Jara, H.T.; Drochner, W.; MÜller, H.-M. A survey of Fusarium toxins in cereal-based foods marketed in an area of southwest Germany. Mycopathologia 1999, 147, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Sedmikova, M.; Reisnerova, H.; Dufkova, Z.; Jilek, F.; Barta, I. Potential hazard of simultaneous occurrence of aflatoxin B1 and ochratoxin A. Vet. Med.-UZPI Czech Repub. 2001, 46, 169–174. [Google Scholar]
- Senyuva, H.Z.; Gilbert, J.; Ozcan, S.; Ulken, U. Survey for co-occurrence of ochratoxin A and aflatoxin B1 in dried figs in Turkey by using a single laboratory-validated alkaline extraction method for ochratoxin A. J. Food Prot. 2005, 68, 1512–1515. [Google Scholar] [PubMed]
- Serrano, A.B.; Font, G.; Mañes, J.; Ferrer, E. Emerging Fusarium mycotoxins in organic and conventional pasta collected in Spain. Food Chem. Toxicol. 2013, 51, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Serrano, A.B.; Font, G.; Ruiz, M.J.; Ferrer, E. Co-occurrence and risk assessment of mycotoxins in food and diet from Mediterranean area. Food Chem. 2012, 135, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Sifou, A.; Meca, G.; Serrano, A.B.; Mahnine, N.; El Abidi, A.; Mañes, J.; El Azzouzi, M.; Zinedine, A. First report on the presence of emerging Fusarium mycotoxins enniatins (A, A1, B, B1), beauvericin and fusaproliferin in rice on the Moroccan retail markets. Food Control 2011, 22, 1826–1830. [Google Scholar] [CrossRef]
- Sultana, N.; Rashid, A.; Tahira, I.; Hanif, H.U.; Hanif, N.Q. Distribution of various mycotoxins in compound feed, total mix ration and silage. Pak. Vet. J. Pak. 2013. [Google Scholar]
- Sun, G.; Wang, S.; Hu, X.; Su, J.; Zhang, Y.; Xie, Y.; Zhang, H.; Tang, L.; Wang, J.-S. Co-contamination of aflatoxin B1 and fumonisin B1 in food and human dietary exposure in three areas of China. Food Addit. Contam. Part Chem. Anal. Control Expo. Risk Assess. 2011, 28, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Yamamoto, S.; Hasegawa, A.; Aoki, N.; Besling, J.R.; Sugiura, Y.; Ueno, Y. A survey of the natural occurrence of Fusarium mycotoxins, deoxynivalenol, nivalenol and zearalenone, in cereals harvested in the Netherlands. Mycopathologia 1990, 110, 19–22. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Hasegawa, A.; Yamamoto, S.; Sugiura, Y.; Ueno, Y. A case report on a minor contamination of nivalenol in cereals harvested in Canada. Mycopathologia 1988, 101, 157–160. [Google Scholar] [CrossRef] [PubMed]
- Ueno, Y.; Iijima, K.; Wang, S.-D.; Sugiura, Y.; Sekijima, M.; Tanaka, T.; Chen, C.; Yu, S.-Z. Fumonisins as a possible contributory risk factor for primary liver cancer: A 3-year study of corn harvested in Haimen, China, by HPLC and ELISA. Food Chem. Toxicol. 1997, 35, 1143–1150. [Google Scholar] [CrossRef]
- Varga, E.; Glauner, T.; Berthiller, F.; Krska, R.; Schuhmacher, R.; Sulyok, M. Development and validation of a (semi-)quantitative UHPLC-MS/MS method for the determination of 191 mycotoxins and other fungal metabolites in almonds, hazelnuts, peanuts and pistachios. Anal. Bioanal. Chem. 2013, 405, 5087–5104. [Google Scholar] [CrossRef] [PubMed]
- Vargas, E.A.; Preis, R.A.; Castro, L.; Silva, C.M. Co-occurrence of aflatoxins B1, B2, G1, G2, zearalenone and fumonisin B1 in Brazilian corn. Food Addit. Contam. 2001, 18, 981–986. [Google Scholar] [CrossRef] [PubMed]
- Villa, P.; Markaki, P. Aflatoxin B1 and ochratoxin A in breakfast cereals from athens market: Occurrence and risk assessment. Food Control 2009, 20, 455–461. [Google Scholar] [CrossRef]
- Vrabcheva, T.; Usleber, E.; Dietrich, R.; Märtlbauer, E. Co-occurrence of ochratoxin A and citrinin in cereals from Bulgarian villages with a history of Balkan endemic nephropathy. J. Agric. Food Chem. 2000, 48, 2483–2488. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.-S.; Liang, Y.-X.; Chau, N.T.; Dien, L.D.; Tanaka, T.; Ueno, Y. Natural co-occurrence of Fusarium toxins and aflatoxin B1 in com for feed in North Vietnam. Nat. Toxins 1995, 3, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, A.; Yoshizawa, T.; Aiura, Y.; Sanchez, P.C.; Dizon, E.I.; Arim, R.H.; Sardjono. Fusarium mycotoxins (fumonisins, nivalenol, and zearalenone) and aflatoxins in corn from Southeast Asia. Biosci. Biotechnol. Biochem. 1995, 59, 1804–1807. [Google Scholar] [CrossRef] [PubMed]
- Yoshizawa, T.; Yamashita, A.; Chokethaworn, N. Occurrence of fumonisins and aflatoxins in corn from Thailand. Food Addit. Contam. 1996, 13, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Zinedine, A.; Brera, C.; Elakhdari, S.; Catano, C.; Debegnach, F.; Angelini, S.; De Santis, B.; Faid, M.; Benlemlih, M.; Minardi, V.; Miraglia, M. Natural occurrence of mycotoxins in cereals and spices commercialized in Morocco. Food Control 2006, 17, 868–874. [Google Scholar] [CrossRef]
- BIOMIN. Science & Solutions; BIOMIN Holding GmbH: Herzogenburg, Austria, 2015. [Google Scholar]
- Merhej, J.; Richard-Forget, F.; Barreau, C. Regulation of trichothecene biosynthesis in Fusarium: Recent advances and new insights. Appl. Microbiol. Biotechnol. 2011, 91, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Gareis, M.; Zimmerman, C.; Schothorst, R.; Paulsch, W.; Vidnes, A.; Bergsten, C.; Paulsen, B.; Brera, C.; Maraglia, M.; Grossi, S.; et al. Collection of occurrence data of Fusarium toxins in food and assessment of dietary intake by the population of EU member states; Directorate-General Health and Consumer Protection: Kulmbach, Germany; Berlin, Germany; Bilthoven, The Netherlands; Oslo, Norway; Rome, Italy, 2003; p. 606. [Google Scholar]
- Miraglia, M.; Brera, C. Assessment of dietary intake of ochratoxin A by the population of EU Member States; Directorate-General Health and Consumer Protection: Rome, Italy, 2002; p. 153. [Google Scholar]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed]
- Pereira, V.L.; Fernandes, J.O.; Cunha, S.C. Mycotoxins in cereals and related foodstuffs: A review on occurrence and recent methods of analysis. Trends Food Sci. Technol. 2014, 36, 96–136. [Google Scholar] [CrossRef]
- Paterson, R.R.M.; Lima, N. How will climate change affect mycotoxins in food? Food Res. Int. 2010, 43, 1902–1914. [Google Scholar] [CrossRef] [Green Version]
- Paterson, R.R.M.; Lima, N. Further mycotoxin effects from climate change. Food Res. Int. 2011, 44, 2555–2566. [Google Scholar] [CrossRef] [Green Version]
- Klarić, M.Š. Adverse effects of combined mycotoxins/Štetni učinci kombiniranih mikotoksina. Arch. Ind. Hyg. Toxicol. 2012, 63, 519–530. [Google Scholar]
- Richard, J.L. Some major mycotoxins and their mycotoxicoses—An overview. Int. J. Food Microbiol. 2007, 119, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Alassane-Kpembi, I.; Puel, O.; Oswald, I.P. Toxicological interactions between the mycotoxins deoxynivalenol, nivalenol and their acetylated derivatives in intestinal epithelial cells. Arch. Toxicol. 2015, 89, 1337–1346. [Google Scholar] [CrossRef] [PubMed]
- Alassane-Kpembi, I.; Kolf-Clauw, M.; Gauthier, T.; Abrami, R.; Abiola, F.A.; Oswald, I.P.; Puel, O. New insights into mycotoxin mixtures: The toxicity of low doses of Type B trichothecenes on intestinal epithelial cells is synergistic. Toxicol. Appl. Pharmacol. 2013, 272, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Bensassi, F.; Gallerne, C.; Sharaf el dein, O.; Hajlaoui, M.R.; Lemaire, C.; Bacha, H. In vitro investigation of toxicological interactions between the fusariotoxins deoxynivalenol and zearalenone. Toxicon 2014, 84, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Bernhoft, A.; Keblys, M.; Morrison, E.; Larsen, H.J.S.; Flåøyen, A. Combined effects of selected Penicillium mycotoxins on in vitro proliferation of porcine lymphocytes. Mycopathologia 2004, 158, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Bouaziz, C.; Bouslimi, A.; Kadri, R.; Zaied, C.; Bacha, H.; Abid-Essefi, S. The in vitro effects of zearalenone and T-2 toxins on Vero cells. Exp. Toxicol. Pathol. 2013, 65, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Bouslimi, A.; Bouaziz, C.; Ayed-Boussema, I.; Hassen, W.; Bacha, H. Individual and combined effects of ochratoxin A and citrinin on viability and DNA fragmentation in cultured Vero cells and on chromosome aberrations in mice bone marrow cells. Toxicology 2008, 251, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bouslimi, A.; Ouannes, Z.; Golli, E.E.; Bouaziz, C.; Hassen, W.; Bacha, H. Cytotoxicity and oxidative damage in kidney cells exposed to the mycotoxins ochratoxin A and citrinin: Individual and combined effects. Toxicol. Mech. Methods 2008, 18, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Braunberg, R.C.; Barton, C.N.; Gantt, O.O.; Friedman, L. Interaction of citrinin and ochratoxin A. Nat. Toxins 1994, 2, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Corcuera, L.A.; Arbillaga, L.; Vettorazzi, A.; Azqueta, A.; López de Cerain, A. Ochratoxin A reduces aflatoxin B1 induced DNA damage detected by the comet assay in Hep G2 cells. Food Chem. Toxicol. 2011, 49, 2883–2889. [Google Scholar] [CrossRef] [PubMed]
- Creppy, E.E.; Chiarappa, P.; Baudrimont, I.; Borracci, P.; Moukha, S.; Carratù, M.R. Synergistic effects of fumonisin B1 and ochratoxin A: Are in vitro cytotoxicity data predictive of in vivo acute toxicity? Toxicology 2004, 201, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Creppy, E.-E.; Lorkowski, G.; Beck, G.; Röschenthaler, R.; Dirheimer, G. Combined action of citrinin and ochratoxin A on hepatoma tissue culture cells. Toxicol. Lett. 1980, 5, 375–380. [Google Scholar] [CrossRef]
- Dombrink-Kurtzman, M.A. Fumonisin and beauvericin induce apoptosis in turkey peripheral blood lymphocytes. Mycopathologia 2003, 156, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Ficheux, A.S.; Sibiril, Y.; Parent-Massin, D. Co-exposure of Fusarium mycotoxins: in vitro myelotoxicity assessment on human hematopoietic progenitors. Toxicon 2012, 60, 1171–1179. [Google Scholar] [CrossRef] [PubMed]
- Föllmann, W.; Behm, C.; Degen, G.H. Toxicity of the mycotoxin citrinin and its metabolite dihydrocitrinone and of mixtures of citrinin and ochratoxin A in vitro. Arch. Toxicol. 2014, 88, 1097–1107. [Google Scholar] [CrossRef] [PubMed]
- Föllmann, W.; Lebrun, S.; Kullik, B.; Koch, M.; Römer, H.C.; Golka, K. Cytotoxicity of ochratoxin A and citrinin in different cell types in vitro. Mycotoxin Res. 2000, 16, 123–126. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, K.; Mounier, J.; Coton, E.; Hymery, N. Individual and combined effects of roquefortine C and mycophenolic acid on human monocytic and intestinal cells. World Mycotoxin J. 2015, 1–11. [Google Scholar] [CrossRef]
- Gayathri, L.; Dhivya, R.; Dhanasekaran, D.; Periasamy, V.S.; Alshatwi, A.A.; Akbarsha, M.A. Hepatotoxic effect of ochratoxin A and citrinin, alone and in combination, and protective effect of vitamin E: In vitro study in HepG2 cell. Food Chem. Toxicol. 2015, 83, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Golli-Bennour, E.E.; Kouidhi, B.; Bouslimi, A.; Abid-Essefi, S.; Hassen, W.; Bacha, H. Cytotoxicity and genotoxicity induced by aflatoxin B1, ochratoxin A, and their combination in cultured Vero cells. J. Biochem. Mol. Toxicol. 2010, 24, 42–50. [Google Scholar] [CrossRef] [PubMed]
- He, C.-H.; Fan, Y.-H.; Wang, Y.; Huang, C.-Y.; Wang, X.-C.; Zhang, H.-B. The individual and combined effects of deoxynivalenol and aflatoxin B1 on primary hepatocytes of Cyprinus carpio. Int. J. Mol. Sci. 2010, 11, 3760–3768. [Google Scholar] [CrossRef] [PubMed]
- Heussner, A.H.; Dietrich, D.R.; O’Brien, E. In vitro investigation of individual and combined cytotoxic effects of ochratoxin A and other selected mycotoxins on renal cells. Toxicol. In Vitro 2006, 20, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Heussner, A.H.; O’Brien, E.; Haehnlein, J.; Biester, M.A.; Dietrich, D.R. Comparison of interactive cytotoxic effects of selected mycotoxins on renal cells. Toxicol. Sci. 2004, 78, 89. [Google Scholar]
- Klarić, M.Š.; Medić, N.; Hulina, A.; Žanić Grubišić, T.; Rumora, L. Disturbed Hsp70 and Hsp27 expression and thiol redox status in porcine kidney PK15 cells provoked by individual and combined ochratoxin A and citrinin treatments. Food Chem. Toxicol. 2014, 71, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Klarić, M.Š.; Želježić, D.; Rumora, L.; Peraica, M.; Pepeljnjak, S.; Domijan, A.-M. A potential role of calcium in apoptosis and aberrant chromatin forms in porcine kidney PK15 cells induced by individual and combined ochratoxin A and citrinin. Arch. Toxicol. 2012, 86, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Klarić, M.Š.; Daraboš, D.; Rozgaj, R.; Kašuba, V.; Pepeljnjak, S. Beauvericin and ochratoxin A genotoxicity evaluated using the alkaline comet assay: single and combined genotoxic action. Arch. Toxicol. 2010, 84, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Klarić, M.Š.; Pepeljnjak, S.; Rozgaj, R. Genotoxicity of fumonisin B1, beauvericin and ochratoxin A in porcine kidney PK15 cells: Effects of individual and combined treatment. Croat. Chem. Acta 2008, 81, 139–146. [Google Scholar]
- Klarić, M.Š.; Rumora, L.; Ljubanović, D.; Pepeljnjak, S. Cytotoxicity and apoptosis induced by fumonisin B1, beauvericin and ochratoxin A in porcine kidney PK15 cells: effects of individual and combined treatment. Arch. Toxicol. 2008, 82, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Klarić, M.Š.; Pepeljnjak, S.; Domijan, A.-M.; Petrik, J. Lipid peroxidation and glutathione levels in porcine kidney PK15 cells after individual and combined treatment with fumonisin B1, beauvericin and ochratoxin A. Basic Clin. Pharmacol. Toxicol. 2006, 100, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Knecht, A.; Schwerdt, G.; Gekle, M.; Humpf, H.-U. Combinatory effects of citrinin and ochratoxin A in immortalized human proximal tubule cells. Mycotoxin Res. 2005, 21, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Koshinsky, H.A.; Hannan, P.J.; Khachatourians, G.G. HT-2 toxin, roridin A, T-2 toxin, and verrucarin A mycotoxins inhibit carbon dioxide production by Kluyveromyces marxianus. Can. J. Microbiol. 1991, 37, 933–938. [Google Scholar] [CrossRef]
- Koshinsky, H.A.; Khachatourians, G.G. Trichothecene synergism, additivity, and antagonism: The significance of the maximally quiescent ratio. Nat. Toxins 1992, 1, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Koshinsky, H.A.; Khachatourians, G.G. Bioassay for deoxynivalenol based on the interaction of T-2 toxin with trichothecene mycotoxins. Bull. Environ. Contam. Toxicol. 1992, 49, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Kouadio, J.H.; Brou, K.; Gnakri, D. Low level of ochratoxin A enhances aflatoxin B1 induced cytotoxicity and lipid peroxydation in both human intestinal (Caco-2) and hepatoma (HepG2) cells lines. Int. J. Nutr. Food Sci. 2013, 2, 294–300. [Google Scholar]
- Kouadio, J.H.; Moukha, S.; Brou, K.; Gnakri, D. Modulation of fumonisin B1 toxic action-induced by zeralenone in human intestinal cells Caco-2. Int. J. Sci. Technol. Res. 2013, 2, 315–320. [Google Scholar]
- Kouadio, J.H.; Dano, S.D.; Moukha, S.; Mobio, T.A.; Creppy, E.E. Effects of combinations of Fusarium mycotoxins on the inhibition of macromolecular synthesis, malondialdehyde levels, DNA methylation and fragmentation, and viability in Caco-2 cells. Toxicon 2007, 49, 306–317. [Google Scholar] [CrossRef] [PubMed]
- Lei, M.; Zhang, N.; Qi, D. In vitro investigation of individual and combined cytotoxic effects of aflatoxin B1 and other selected mycotoxins on the cell line porcine kidney 15. Exp. Toxicol. Pathol. 2013, 65, 1149–1157. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Fernández-Franzón, M.; Font, G.; Ruiz, M.J. Toxicity evaluation of individual and mixed enniatins using an in vitro method with CHO-K1 cells. Toxicol. In Vitro 2013, 27, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Luongo, D.; De Luna, R.; Russo, R.; Severino, L. Effects of four Fusarium toxins (fumonisin B1, α-zearalenol, nivalenol and deoxynivalenol) on porcine whole-blood cellular proliferation. Toxicon 2008, 52, 156–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luongo, D.; Severino, L.; Bergamo, P.; De Luna, R.; Lucisano, A.; Rossi, M. Interactive effects of fumonisin B1 and α-zearalenol on proliferation and cytokine expression in Jurkat T cells. Toxicol. In Vitro 2006, 20, 1403–1410. [Google Scholar] [CrossRef] [PubMed]
- Madhyastha, M.S.; Marquardt, R.R.; Abramson, D. Structure-activity relationships and interactions among trichothecene mycotoxins as assessed by yeast bioassay. Toxicon 1994, 32, 1147–1152. [Google Scholar] [CrossRef]
- Manderville, R.; Pfohl-Leszkowicz, A. Bioactivation and DNA adduction as a rationale for ochratoxin A carcinogenesis. World Mycotoxin J. 2008, 1, 357–367. [Google Scholar] [CrossRef]
- Marzocco, S.; Russo, R.; Bianco, G.; Autore, G.; Severino, L. Pro-apoptotic effects of nivalenol and deoxynivalenol trichothecenes in J774A.1 murine macrophages. Toxicol. Lett. 2009, 189, 21–26. [Google Scholar] [CrossRef] [PubMed]
- McKean, C.; Tang, L.; Tang, M.; Billam, M.; Wang, Z.; Theodorakis, C.W.; Kendall, R.J.; Wang, J.-S. Comparative acute and combinative toxicity of aflatoxin B1 and fumonisin B1 in animals and human cells. Food Chem. Toxicol. 2006, 44, 868–876. [Google Scholar] [CrossRef] [PubMed]
- Prosperini, A.; Font, G.; Ruiz, M.J. Interaction effects of Fusarium enniatins (A, A1, B and B1) combinations on in vitro cytotoxicity of Caco-2 cells. Toxicol. In Vitro 2014, 28, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, M.-J.; Franzova, P.; Juan-García, A.; Font, G. Toxicological interactions between the mycotoxins beauvericin, deoxynivalenol and T-2 toxin in CHO-K1 cells in vitro. Toxicon 2011, 58, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, M.-J.; Macáková, P.; Juan-García, A.; Font, G. Cytotoxic effects of mycotoxin combinations in mammalian kidney cells. Food Chem. Toxicol. 2011, 49, 2718–2724. [Google Scholar] [CrossRef] [PubMed]
- Rumora, L.; Domijan, A.-M.; Žanić Grubišić, T.; Klarić, M.Š. Differential activation of MAPKs by individual and combined ochratoxin A and citrinin treatments in porcine kidney PK15 cells. Toxicon 2014, 90, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Severino, L.; Luongo, D.; Bergamo, P.; Lucisano, A.; Rossi, M. Mycotoxins nivalenol and deoxynivalenol differentially modulate cytokine mRNA expression in Jurkat T cells. Cytokine 2006, 36, 75–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoev, S.; Denev, S.; Dutton, M.; Nkosi, B. Cytotoxic effect of some mycotoxins and their combinations on human peripheral blood mononuclear cells as measured by the MTT assay. Open Toxinology J. 2009, 2, 1–8. [Google Scholar] [CrossRef]
- Tajima, O.; Schoen, E.D.; Feron, V.J.; Groten, J.P. Statistically designed experiments in a tiered approach to screen mixtures of Fusarium mycotoxins for possible interactions. Food Chem. Toxicol. 2002, 40, 685–695. [Google Scholar] [CrossRef]
- Tammer, B.; Lehmann, I.; Nieber, K.; Altenburger, R. Combined effects of mycotoxin mixtures on human T cell function. Toxicol. Lett. 2007, 170, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Tatay, E.; Meca, G.; Font, G.; Ruiz, M.-J. Interactive effects of zearalenone and its metabolites on cytotoxicity and metabolization in ovarian CHO-K1 cells. Toxicol. In Vitro 2014, 28, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Theumer, M.G.; Cánepa, M.C.; López, A.G.; Mary, V.S.; Dambolena, J.S.; Rubinstein, H.R. Subchronic mycotoxicoses in Wistar rats: Assessment of the in vivo and in vitro genotoxicity induced by fumonisins and aflatoxin B1, and oxidative stress biomarkers status. Toxicology 2010, 268, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Thompson, W.L.; Wannemacher, R.W., Jr. Structure-function relationships of 12,13-epoxytrichothecene mycotoxins in cell culture: Comparison to whole animal lethality. Toxicon 1986, 24, 985–994. [Google Scholar] [CrossRef]
- Thuvander, A.; Wikman, C.; Gadhasson, I. In vitro exposure of human lymphocytes to trichothecenes: Individual variation in sensitivity and effects of combined exposure on lymphocyte function. Food Chem. Toxicol. 1999, 37, 639–648. [Google Scholar] [CrossRef]
- Wang, H.W.; Wang, J.Q.; Zheng, B.Q.; Li, S.L.; Zhang, Y.D.; Li, F.D.; Zheng, N. Cytotoxicity induced by ochratoxin A, zearalenone, and α-zearalenol: Effects of individual and combined treatment. Food Chem. Toxicol. 2014, 71, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.Y.M.; Turner, P.C.; El-Nezami, H. Individual and combined cytotoxic effects of Fusarium toxins (deoxynivalenol, nivalenol, zearalenone and fumonisins B1) on swine jejunal epithelial cells. Food Chem. Toxicol. 2013, 57, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Anninou, N.; Chatzaki, E.; Papachristou, F.; Pitiakoudis, Μ.; Simopoulos, C. Mycotoxins’ activity at toxic and sub-toxic concentrations: differential cytotoxic and genotoxic effects of single and combined administration of sterigmatocystin, ochratoxin A and citrinin on the hepatocellular cancer cell line Hep3B. Int. J. Environ. Res. Public. Health 2014, 11, 1855–1872. [Google Scholar] [CrossRef] [PubMed]
- Klarić, M.Š.; Rašić, D.; Peraica, M. Deleterious effects of mycotoxin combinations involving ochratoxin A. Toxins 2013, 5, 1965–1987. [Google Scholar] [CrossRef] [PubMed]
- Weber, F.; Freudinger, R.; Schwerdt, G.; Gekle, M. A rapid screening method to test apoptotic synergisms of ochratoxin A with other nephrotoxic substances. Toxicol. In Vitro 2005, 19, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.-C.; Talalay, P. Quantitative analysis of dose-effect relationships: the combined effects of multiple drugs or enzyme inhibitors. Adv. Enzyme Regul. 1984, 22, 27–55. [Google Scholar] [CrossRef]
- Chou, T.-C. Theoretical basis, experimental design, and computerized simulation of synergism and antagonism in drug combination studies. Pharmacol. Rev. 2006, 58, 621–681. [Google Scholar] [CrossRef] [PubMed]


 = mixtures no cited;
= mixtures no cited;  = mixtures cited between 1 and 3 times;
= mixtures cited between 1 and 3 times;  = mixtures cited between 3 and 5 times;
= mixtures cited between 3 and 5 times;  = mixtures cited between 5 and 7 times;
= mixtures cited between 5 and 7 times;  = mixtures cited between 7 and 9 times;
= mixtures cited between 7 and 9 times;  = mixtures cited more than 9 times). Reading from left to right on the x-axis:
= mixtures no cited; = mixtures cited between 1 and 3 times; = mixtures cited between 3 and 5 times; = mixtures cited between 5 and 7 times; = mixtures cited between 7 and 9 times; = mixtures cited more than 9 times). Reading from left to right on the x-axis:
= mixtures cited more than 9 times). Reading from left to right on the x-axis:
= mixtures no cited; = mixtures cited between 1 and 3 times; = mixtures cited between 3 and 5 times; = mixtures cited between 5 and 7 times; = mixtures cited between 7 and 9 times; = mixtures cited more than 9 times). Reading from left to right on the x-axis:




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mycotoxin | Fungi Source | Product of Primary Concern | Geographical Occurrence |
|---|---|---|---|
| AFs * (B1, B2, G1, G2) | Aspergillus (bombycis, flavus, nomius, ochraceoroseus, parasiticus, parvisclerotigenus, pseudotamarii, rambellii, toxicarius) | Cereals and cereal-based products (mainly corn), nuts, nut products and seeds, dried fruits, spices, milk and dairy products, meat, eggs | Temperate, tropical and subtropical regions (Southern Asia and Africa) |
| OTA * | Aspergillus (alliaceus, auricomus, carbonarius, cretensis, flocculosus, glaucus, lacticoffeatus, meleus, niger, ochraceus, pseudoelegans, roseoglobulosum, sclerotioniger, sclerotiorum, steynii, sulphureus, westerdijkiae); Penicillium (nordicum, verrucosum) | Cereals and cereal-based products (mainly rice and wheat), coffee and cocoa beans; wine, beer, dried fruits, spices, meat | From cool-temperate to tropical regions (Northern and Southern America, Northern and Western Europe, Africa and South Asia) |
| TCTs * (DON, NIV, T-2, HT-2, DAS) | Fusarium (acuminatum, armeniacum, culmorum, crookwellense, equisetii, graminearum, kyushuense, langsethiae, poae, pseudograminearum, sambucinum, scirpi, sporotrichioides, venamtum) | All cereals and cereal-based products | Northern temperate regions (Europe, America and Asia) |
| ZEA * | Fusarium (crookwellense, culmorum, equiseti, graminearum, incarnatum, pseudograminearum, semitectum, sporotrichioides, verticillioides) | All cereals and cereal-based products, and banana | Northern temperate regions (Europe, America and Asia) |
| FUM * (B1, B2, B3) | Fusarium (anthophilum, dlamini, fujikuroi, globosum, napiforme, nygamai, oxysporum, polyphialidicum, proliferatum, pseudonygamai, thapsinum, verticillioides) | Corn, millet, sorghum, rice and their derivatives | Hot-temperate regions (Europe, Africa) |
| BEA * | Fusarium (acuminatum, armeniacum, anthophilum, avenaceum, beomiforme, dlamini, equiseti, fujikuroi, globosum, langsethiae, longipes, nygamai, oxysporum, poae, proliferatum, pseudoanthophilum, sambucinum, semitectum, sporotrichioides, subglutinans) | All cereals and cereal-based products | Temperate regions (Europe) |
| ENs * (A, A1, B, B1) | Fusarium.(acuminatum, avenaceum, langsethiae, lateritium, poae, proliferatum, sambucinum, sporotrichioides, tricinctum) | All cereals and cereal-based products | Temperate regions (Europe) |
| MON * | Fusarium (acuminatum, avenaceum, culmorum, equiseti, fujikuroi, napiforme, nygamai, oxysporum, proliferatum, pseudonygamai, sporotrichioides, subglutinans, thapsinum, tricinctum, verticillioides) | All cereals and cereal-based products | Temperate regions (Europe) |
| Mycotoxin Couples/Cells | Doses (µM) | Exposure | Toxicological Effect | References | |
|---|---|---|---|---|---|
| Interaction between TCT | |||||
| DON+15-ADON | DON: 0.25–4 | 48 h | Synergistic | at low inhibitory concentration levels (IC10, 20, 30) | [155] |
| Human epithelial colorectal adenocarcinoma cells: Caco-2 | 15-ADON: 0.25–4 | Additive | at medium inhibit concentration levels (IC40, 50) | ||
| DON+15-ADON | DON: 0.2–15 | 24 h | Synergistic | from IC10 to IC80 | [154] |
| Intestinal porcine epithelial cells (ileum + jejunum): IPEC-1 | 15-ADON: 0.2–15 | ||||
| DON+3-ADON | DON: 0.25–4 | 48 h | Synergistic | at low and medium inhibitory concentration levels (IC10, 20, 30, 40) | [155] |
| Human epithelial colorectal adenocarcinoma cells: Caco-2 | 3-ADON: 0.42–6.67 | Additive | at the 50% growth inhibition level (IC50) | ||
| DON+3-ADON Intestinal porcine epithelial cells (ileum + jejunum): IPEC-1 | DON: 0.2–15 3-ADON: 2–150 | 24 h | Antagonistic | at low inhibitory concentration levels (IC10–IC30) | [154] |
| Additive | at medium inhibitory concentration levels (IC30–IC60) | ||||
| Synergistic | at high inhibitory concentration levels (IC60–IC80) | ||||
| 15-ADON+3-ADON | 15-ADON: 0.25–4 | 48 h | Synergistic | at low cytotoxicity levels (IC10, 20, 30) | [155] |
| Human epithelial colorectal adenocarcinoma cells: Caco-2 | 3-ADON: 0.42–6.67 | Additive | at medium inhibitory concentration levels (IC40, 50) | ||
| 15-ADON+3-ADON | 15-ADON: 0.2–15 | 24 h | Synergistic | at all cytotoxicity levels (IC10–IC80) | [154] |
| Intestinal porcine epithelial cells (ileum + jejunum): IPEC-1 | 3-ADON: 2–150 | ||||
| DON+15-ADON+3-ADON Human epithelial colorectal adenocarcinoma cells: Caco-2 | DON: 0.25–4 | 48 h | Synergistic | at low cytotoxicity levels (IC10, 20, 30) | [155] |
| 15-ADON: 0.25–4 | Additive | at the 40% growth inhibition level (IC40) | |||
| 3-ADON: 0.42–6.67 | Antagonistic | from the 50% growth inhibition level (IC50) | |||
| DON+NIV | DON: 10–100 | 24 h, 48 h and 72 h | Additive | at 50% growth inhibition level (IC50) | [194] |
| Murine monocyte macrophage cells: J774A.1 | NIV: 10–100 | ||||
| DON+NIV | DON: 0.5–2 | 48 h | Antagonistic | at the lowest dose | [209] |
| Intestinal porcine epithelial cells (jejunum): IPEC-J2 | NIV: 0.5–2 | Synergistic | at the highest dose | ||
| DON+NIV | DON: 0.25–4 | 48 h | Synergistic | at all cytotoxicity levels (from IC10 to IC50) | [155] |
| Human epithelial colorectal adenocarcinoma cells: Caco-2 | NIV: 0.2–3.2 | ||||
| DON+NIV | DON: 0.2–15 | 24 h | Synergistic | at all cytotoxicity levels (from IC10 to IC80) | [154] |
| Intestinal porcine epithelial cells (ileum + jejunum): IPEC-1 | NIV: 0.2–15 | ||||
| DON+FX | DON: 0.25–4 | 48 h | Synergistic | at all cytotoxicity levels (from IC10 to IC50) | [155] |
| Human epithelial colorectal adenocarcinoma cells: Caco-2 | FX: 7.5–120 | ||||
| DON+FX | DON: 0.2–15 | 24 h | Antagonistic | at all inhibitory concentration levels (IC10-IC80) | [154] |
| Intestinal porcine epithelial cells (ileum + jejunum): IPEC-1 | FX: 0.12–9 | ||||
| NIV+FX | NIV: 0.2–3.2 | 48 h | Synergistic | at low cytotoxicity levels (IC10, 20) | [155] |
| Human epithelial colorectal adenocarcinoma cells: Caco-2 | FX: 7.5–120 | Additive | at medium cytotoxicity levels (IC30, 40, 50) | ||
| NIV+FX | NIV: 0.2–15 | 24 h | Additive | at all cytotoxicity levels (IC10-IC80) | [154] |
| Intestinal porcine epithelial cells (ileum + jejunum): IPEC-1 | FX: 0.16–12 | ||||
| DON+NIV+FX Human epithelial colorectal adenocarcinoma cells: Caco-2 | DON: 0.25–4 | 48 h | Antagonistic Additive | at low cytotoxicity levels (IC10, 20) | [155] |
| NIV: 0.2–3.2 | at medium cytotoxicity levels (IC30, 40, 50) | ||||
| FX: 7.5–120 | |||||
| DON+T2 | DON: 0.25–4 | 24 h, 48 h and 72 h | Antagonistic | [197] | |
| Chinese hamster ovary cells: CHO-K1 | T2: 0.006–0.1 | ||||
| DON+T2 | DON: 0.25–8 | 24 h, 48 h and 72 h | Antagonistic | [198] | |
| Monkey kidney epithelial cells: Vero | T2: 0.001–0.05 | ||||
| DON+T2 | DON: 0.04–0.1 | 14 days | Additive | [166] | |
| Hematopoietic progenitors: CFU-GM | T2: 0.0005–0.0016 | ||||
| Interaction between TCT and other fusariotoxins | |||||
| DON+BEA | DON: 0.25–4 | 24 h, 48 h and 72 h | Antagonistic | [197] | |
| Chinese hamster ovary cells: CHO-K1 | BEA: 0.78–12.5 | ||||
| DON+BEA | DON: 0.25–8 | 24 h, 48 h and 72 h | Antagonistic | [198] | |
| Monkey kidney epithelial cells: Vero | BEA: 0.78–25 | ||||
| DON+BEA | DON: 0.04–0.1 | 14 days | Synergistic | [166] | |
| Hematopoietic progenitors: CFU-GM | BEA: 0.064–3.2 | ||||
| T2+BEA | T2: 0.006–0.1 | 24 h, 48 h and 72 h | Synergistic | [197] | |
| Chinese hamster ovary cells: CHO-K1 | BEA: 0.78–12.5 | ||||
| T2+BEA | T2: 0.001–0.05 | 24 h, 48 h and 72 h | Antagonistic | [198] | |
| Monkey kidney epithelial cells: Vero | BEA: 0.78–25 | ||||
| DON+FB1 | DON: 4–20 | 72 h | Additive | [187] | |
| Human epithelial colorectal adenocarcinoma cells: Caco-2 | FB1: 10 | ||||
| DON+FB1 | DON: 0.04–0.1 | 14 days | Antagonistic | [166] | |
| Hematopoietic progenitors: CFU-GM | FB1: 0.5–2 | ||||
| DON+FB1 | DON: 0.5–2 | 48 h | Antagonistic | at the lowest dose | [209] |
| Intestinal porcine epithelial cells (jejunum): IPEC-J2 | FB1: 20–40 | Synergistic | at the highest dose | ||
| NIV+FB1 | NIV: 0.5–2 | 48 h | Antagonistic | at the lowest dose | [209] |
| Intestinal porcine epithelial cells (jejunum): IPEC-J2 | FB1: 20–40 | Synergistic | at the highest dose | ||
| DON+ZEA | DON: 10–20 | 72 h | Additive | [187] | |
| Human epithelial colorectal adenocarcinoma cells: Caco-2 | ZEA: 10–20 | ||||
| DON+ZEA | DON: 0.04–0.1 | 14 days | Additive | [166] | |
| Hematopoietic progenitors: CFU-GM | ZEA: 0.2–10 | ||||
| DON+ZEA | DON: 0.5–2 | 48 h | Antagonistic | at the lowest dose | [209] |
| Intestinal porcine epithelial cells (jejunum): IPEC-J2 | ZEA: 10–40 | Synergistic | at the highest dose | ||
| DON+ZEA | DON: 100 | 24 h | Antagonistic | [156] | |
| Human colon carcinoma cells: HCT116 | ZEA: 40 | ||||
| NIV+ZEA | NIV: 0.5–2 | 48 h | Antagonistic | at the lowest dose | [209] |
| Intestinal porcine epithelial cells (jejunum): IPEC-J2 | ZEA: 10–40 | Synergistic | at the highest dose | ||
| T2+ZEA | T2: 0.0005–0.0016 | 14 days | Additive | [166] | |
| Hematopoietic progenitors: CFU-GM | ZEA: 0.2–10 | ||||
| T2+ZEA | T2: 0.025–0.1 | 24 h | Additive | [158] | |
| Monkey kidney epithelial cells: Vero | ZEA: 0.025–0.1 | ||||
| DON+T2+BEA Chinese hamster ovary cells: CHO-K1 | DON: 0.25–4 | 24 h, 48 h, and 72 h | Synergistic | [197] | |
| T2: 0.006–0.1 | |||||
| BEA: 0.78–12.5 | |||||
| DON+T2+BEA Monkey kidney epithelial cells: Vero | DON: 0.25–8 | 24 h, 48 h, and 72 h | Antagonistic | [198] | |
| T2: 0.001–0.05 | |||||
| BEA: 0.78–25 | |||||
| DON+NIV+ZEA Intestinal porcine epithelial cells (jejunum): IPEC-J2 | DON: 0.5–2 | 48 h | Antagonistic | at the lowest dose | [209] |
| NIV: 0.5–2 | |||||
| ZEA: 10–40 | Synergistic | at the highest dose | |||
| DON+NIV+FB1 | DON: 0.5–2 | 48 h | Antagonistic | at the lowest dose | [209] |
| NIV: 0.5–2 | |||||
| Intestinal porcine epithelial cells (jejunum): IPEC-J2 | FB1: 20–40 | Synergistic | at the highest dose | ||
| DON+ZEA+FB1 Human epithelial colorectal adenocarcinoma cells: Caco-2 | DON: 10–20 ZEA: 10–20 FB1: 10 | 72 h | Additive | [187] | |
| DON+ZEA+FB1 Intestinal porcine epithelial cells (jejunum): IPEC-J2 | DON: 0.5–2Z EA: 10–40 FB1: 20–40 | 48 h | Antagonistic Synergistic | at the lowest dose at the highest dose | [209] |
| NIV+ZEA+FB1 Intestinal porcine epithelial cells (jejunum): IPEC-J2 | NIV: 0.5–2 ZEA: 10–40 FB1: 20–40 | 48 h | Antagonistic Synergistic | at the lowest dose at the highest dose | [209] |
| DON+NIV+ZEA+FB1 Intestinal porcine epithelial cells (jejunum): IPEC-J2 | DON: 0.5–2 NIV: 0.5–2 ZEA: 10–40 FB1: 20–40 | 48 h | Antagonistic Synergistic | at the lowest dose at the highest dose | [209] |
| Interaction between other fusariotoxins | |||||
| ZEA+α-ZOL Human hepatocellular carcinoma cells: HepG2 | ZEA: 0.5–50 α-ZOL: 1–100 | 24 h and 72 h | Antagonistic | at all cytotoxicity levels (from IC10 to IC90) | [208] |
| 48 h | Antagonistic | at IC10, 20, 30, 40 | |||
| Additive | at IC50, 60, 70 | ||||
| Synergistic | at IC80, 90 | ||||
| ZEA+α-ZOL Chinese hamster ovary cells: CHO-K1 | ZEA: 12.5–50 α-ZOL: 6.25–25 | 24 h | Synergistic | at low cytotoxicity level (IC25) | [204] |
| Additive | at medium and high cytotoxicity levels (from IC50 to IC90) | ||||
| 48 h and 72 h | Additive | at all cytotoxicity levels (from IC25 to IC90) | |||
| ZEA+β-ZOL Chinese hamster ovary cells: CHO-K1 | ZEA: 12.5–50 β-ZOL: 6.25–25 | 24 h, 48 h and 72 h | Additive | at all cytotoxicity levels (from IC25 to IC90) | [204] |
| α-ZOL+β-ZOL Chinese hamster ovary cells: CHO-K1 | α-ZOL: 6.25–25 | 24 h | Additive | at all cytotoxicity levels (IC25, 50, 75, 90) | [204] |
| 48 h | Antagonistic | at low and medium cytotoxicity levels (IC25, 50) | |||
| β-ZOL: 6.25–25 | Additive | at high cytotoxicity levels (IC75, 90) | |||
| 72 h | Additive | at low and high cytotoxicity levels (IC25, 75, 90) | |||
| Antagonistic | at medium cytotoxicity level (IC50) | ||||
| ZEA+α-ZOL+β-ZOL | ZEA: 12.5–5 | 24 h and 48 h | Antagonistic | at low and medium cytotoxicity levels (IC25, 50) | [204] |
| Synergistic | at high cytotoxicity levels (IC75, 90) | ||||
| Chinese hamster ovary cells: CHO-K1 | α-ZOL: 6.25–25 β-ZOL: 6.25–25 | 72 h | Antagonistic | at low cytotoxicity level (IC25) | |
| Synergistic | at medium and high cytotoxicity levels (IC50, 75, 90) | ||||
| ZEA+FB1 Human epithelial colorectal adenocarcinoma cells: Caco-2 | ZEA: 5–20 FB1: 10 | 72 h | Antagonistic | [187] | |
| ZEA+FB1 Intestinal porcine epithelial cells (jejunum): IPEC-J2 | ZEA: 10–40 FB1: 20–40 | 48 h | Antagonistic | at the lowest dose | [209] |
| Synergistic | at the highest dose | ||||
| ZEA+FB1 Human epithelial colorectal adenocarcinomia cells: Caco-2 | ZEA: 10 FB1: 10 | 72 h | Antagonistic | [186] | |
| BEA+FB1 Porcine renal proximal tubular epithelial cells: PK15 | BEA: 0.064–6.4 µM FB1: 0.069–6.9 µM | 24 h | Additive | at low doses | [179] |
| Antagonistic | at the highest dose | ||||
| BEA+ENB Hematopoietic progenitors: CFU-GM | BEA: 0.064–3.2 ENB: 2–6 | 14 days | Additive | [166] | |
| ENA+ENA1 Chinese hamster ovary cells: CHO-K1 | ENA: 0.365–5 ENA1: 0.625–5 | 24 h | Synergistic | at low cytotoxicity levels (IC25) | [189] |
| Additive | at medium and high cytotoxicity levels (IC50, 75, 90) | ||||
| ENA+ENA1 Human epithelial colorectal adenocarcinomia cells: Caco-2 | ENA: 0.365–5 ENA1: 0.625–5 | 24 h | Antagonistic | at the lowest fraction affected (IC5) | [196] |
| Additive | at other fractions affected (IC25, 50, 75, 90) | ||||
| ENA+ENB Chinese hamster ovary cells: CHO-K1 | ENA: 0.365–5 ENB: 0.625–5 | 24 h | Synergistic | at low and medium cytotoxicity levels (IC25, 50) | [189] |
| Additive | at high cytotoxicity levels (IC75, 90) | ||||
| ENA+ENB Human epithelial colorectal adenocarcinomia cells: Caco-2 | ENA: 0.365–5 ENB: 0.625–5 | 24 h | Antagonistic | at the lowest fraction affected (IC5) | [196] |
| Additive | at other fractions affected (IC25, 50, 75, 90) | ||||
| ENA+ENB1 Chinese hamster ovary cells: CHO-K1 | ENA: 0.365–5 ENB1: 0.625–5 | 24 h | Additive | at all inhibitory concentration levels (IC25, 50, 75, 90) | [189] |
| ENA+ENB1 Human epithelial colorectal adenocarcinomia cells: Caco-2 | ENA: 0.365–5 ENB1: 0.625–5 | 24 h | Antagonistic | at the lowest fraction affected (IC5) | [196] |
| Additive | at other fractions affected (IC25, 50, 75, 90) | ||||
| ENA1+ENB | EN A1: 0.365–5 | 24 h | Additive | at all inhibitory concentration levels (IC25, 50, 75, 90) | [189] |
| Chinese hamster ovary cells: CHO-K1 | ENB: 0.625–5 | ||||
| ENA1+ENB Human epithelial colorectal adenocarcinoma cells: Caco-2 | EN A1: 0.365–5 ENB: 0.625–5 | 24 h | Antagonistic | at the lowest fraction affected (IC5) | [196] |
| Additive | at medium fractions affected (IC25, 50, 75) | ||||
| Synergistic | at the highest fraction affected (IC90) | ||||
| ENA1+ ENB1 | EN A1: 0.365–5 | 24 h | Synergistic | at low, medium and high cytotoxicity levels (IC25, 50, 75) | [189] |
| Chinese hamster ovary cells: CHO-K1 | ENB1: 0.625–5 | Additive | at very high cytotoxicity levels (IC 90) | ||
| ENA1+ ENB1 Human epithelial colorectal adenocarcinoma cells: Caco-2 | EN A1: 0.365–5 ENB1: 0.625–5 | 24 h | Additive Synergistic | at the lowest fraction affected (IC5) | [196] |
| at medium fractions affected (IC25, 50) | |||||
| at the two highest fractions affected (IC75, 90) | |||||
| ENB+ENB1 | ENB: 0.365–5 | 24 h | Additive | at all inhibitory concentration levels (IC25, 50, 75, 90) | [189] |
| Chinese hamster ovary cells: CHO-K1 | ENB1: 0.625–5 | ||||
| ENB+ENB1 Human epithelial colorectal adenocarcinoma cells: Caco-2 | ENB: 0.365–5ENB1: 0.625–5 | 24 h | Antagonistic | at the two lowest fractions affected (IC5, 25) | [196] |
| Additive | at other fractions affected (IC50, 75, 90) | ||||
| ENA+ENA1+ENB Chinese hamster ovary cells: CHO-K1 | ENA: 0.3125–2.5 | 24 h | Synergistic Additive | at low and medium cytotoxicity levels (IC25, 50) at high cytotoxicity levels (IC75, 90) | [189] |
| ENA1: 0.3125–2.5 | |||||
| ENB: 0.3125–2.5 | |||||
| ENA+ENA1+ENB Human epithelial colorectal adenocarcinoma cells: Caco-2 | ENA: 1.25–5 | 24 h | Antagonistic | at the lowest fraction affected (IC5) | [196] |
| ENA1: 1.25–5 | Additive | at medium fractions affected (IC25, 50) | |||
| ENB: 1.25–5 | Synergistic | at the two highest fractions affected (IC75, 90) | |||
| ENA+ENA1+ENB1 Chinese hamster ovary cells: CHO-K1 | ENA: 0.3125–2.5 | 24 h | Synergistic | at low and medium cytotoxicity levels (IC25, 50) | [189] |
| ENA1: 0.3125–2.5 | Additive | at high cytotoxicity level (IC75) | |||
| ENB1: 0.3125–2.5 | Antagonistic | at very high cytotoxicity level (IC90) | |||
| ENA+ENA1+ENB1 Human epithelial colorectal adenocarcinoma cells: Caco-2 | ENA: 1.25–5 | 24 h | Antagonistic Additive | at the lowest fraction affected (IC5) at other fractions affected (IC25, 50, 75, 90) | [196] |
| ENA1: 1.25–5 | |||||
| ENB1: 1.25–5 | |||||
| ENA+ENB+ENB1 Chinese hamster ovary cells: CHO-K1 | ENA: 0.3125–2.5 | 24 h | Synergistic Additive | at low and medium cytotoxicity levels (IC25, 50) at high cytotoxicity levels (IC75, 90) | [189] |
| ENB: 0.3125–2.5 | |||||
| ENB1: 0.3125–2.5 | |||||
| ENA+ENB+ENB1 Human epithelial colorectal adenocarcinoma cells: Caco-2 | ENA: 1.25–5 | 24 h | Antagonistic Additive | at the lowest fraction affected (IC5) at other fractions affected (IC25, 50, 75, 90) | [196] |
| ENB: 1.25–5 | |||||
| ENB1: 1.25–5 | |||||
| ENA1+ENB+ENB1 Chinese hamster ovary cells: CHO-K1 | ENA1: 0.3125–2.5 | 24 h | Synergistic | at low cytotoxicity level (IC25) | [189] |
| ENB: 0.3125–2.5 | Additive | at medium and high cytotoxicity levels (IC50,75) | |||
| ENB1: 0.3125–2.5 | Antagonistic | at very high cytotoxicity level (IC90) | |||
| EN A1+ENB+ENB1 Human epithelial colorectal adenocarcinoma cells: Caco-2 | ENA1: 1.25–5 | 24 h | Antagonistic Additive | at the lowest fraction affected (IC5) at other fractions affected (IC25, 50, 75, 90) | [196] |
| ENB: 1.25–5 | |||||
| ENB1: 1.25–5 | |||||
| ENA+ENA1+ENB+ENB1 Human epithelial colorectal adenocarcinoma cells: Caco-2 | ENA: 1.25–5 | 24 h | Antagonistic Additive | at the lowest fraction affected (IC5) at other fractions affected (IC25, 50, 75, 90) | [196] |
| ENA1: 1.25–5 | |||||
| ENB: 1.25–5 | |||||
| ENB1: 1.25–5 | |||||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smith, M.-C.; Madec, S.; Coton, E.; Hymery, N. Natural Co-Occurrence of Mycotoxins in Foods and Feeds and Their in vitro Combined Toxicological Effects. Toxins 2016, 8, 94. https://doi.org/10.3390/toxins8040094
Smith M-C, Madec S, Coton E, Hymery N. Natural Co-Occurrence of Mycotoxins in Foods and Feeds and Their in vitro Combined Toxicological Effects. Toxins. 2016; 8(4):94. https://doi.org/10.3390/toxins8040094
Chicago/Turabian StyleSmith, Marie-Caroline, Stéphanie Madec, Emmanuel Coton, and Nolwenn Hymery. 2016. "Natural Co-Occurrence of Mycotoxins in Foods and Feeds and Their in vitro Combined Toxicological Effects" Toxins 8, no. 4: 94. https://doi.org/10.3390/toxins8040094
APA StyleSmith, M. -C., Madec, S., Coton, E., & Hymery, N. (2016). Natural Co-Occurrence of Mycotoxins in Foods and Feeds and Their in vitro Combined Toxicological Effects. Toxins, 8(4), 94. https://doi.org/10.3390/toxins8040094