
TGF-β1/Smad3 Signaling Pathway Mediates T-2 Toxin-Induced Decrease of Type II Collagen in Cultured Rat Chondrocytes

Abstract
:1. Introduction
2. Results
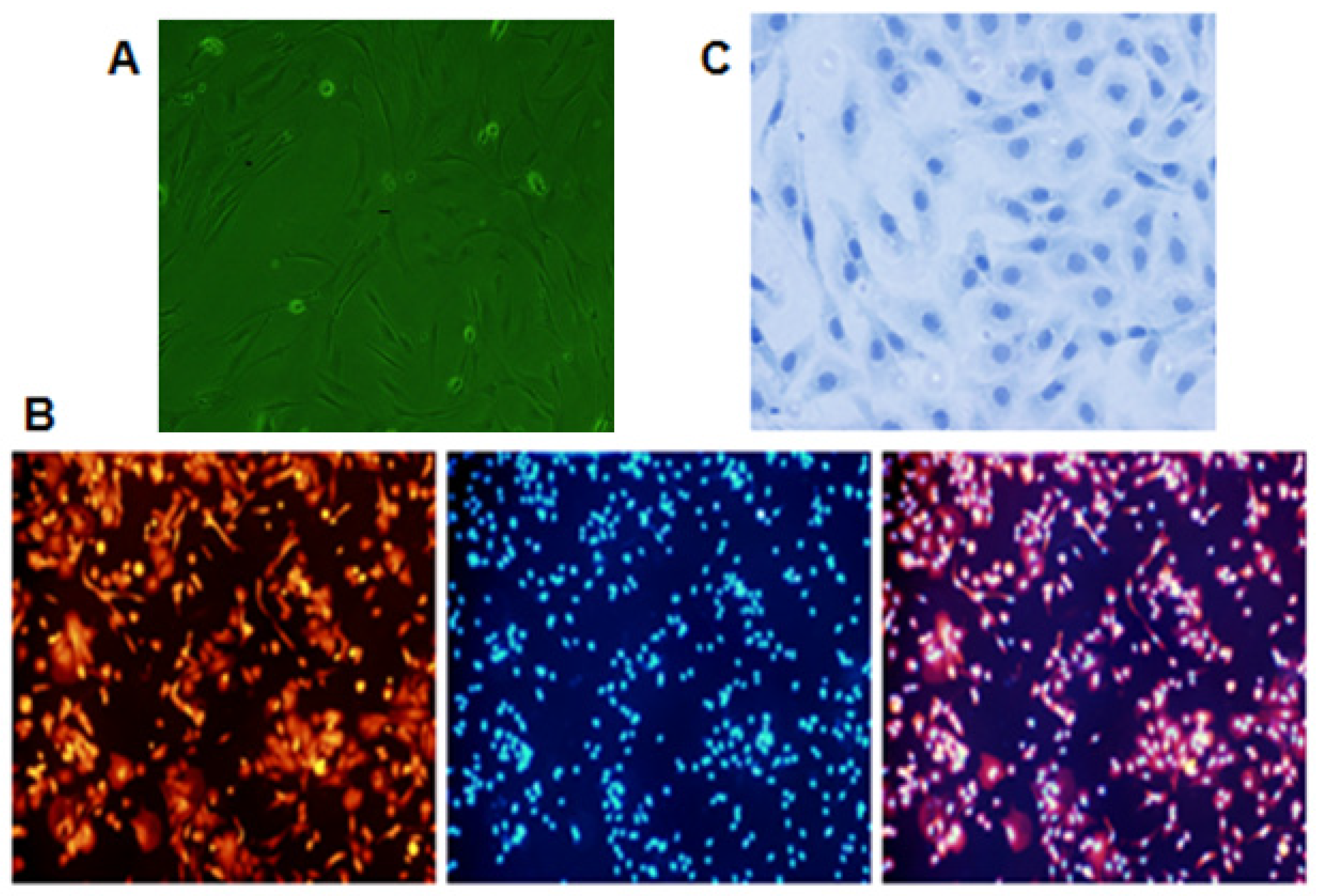
2.1. Effects of T-2 Toxin on Cell Survival Rate of Chondrocytes
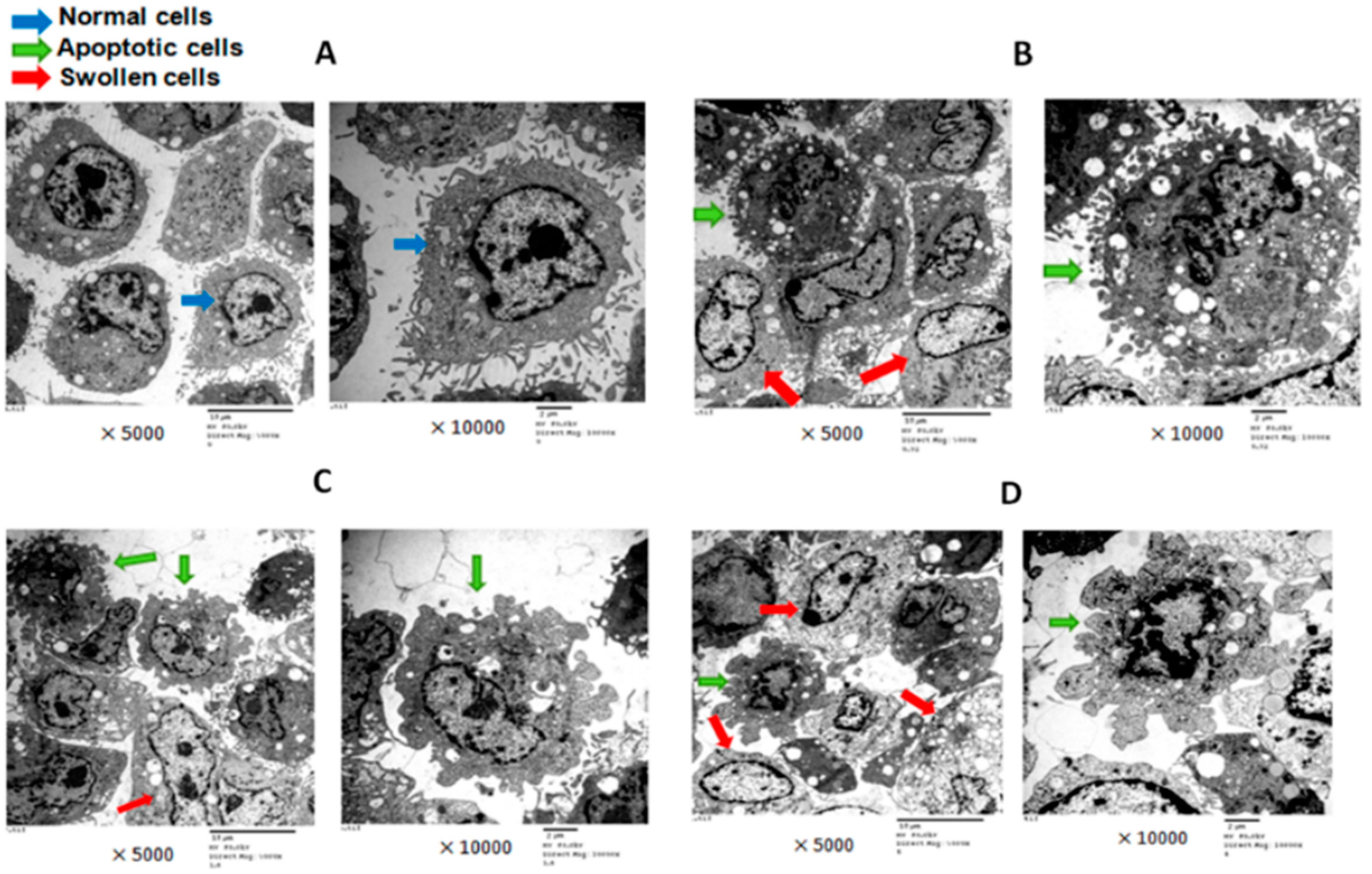
2.2. T-2 Toxin and Ultra-Structure of Chondrocytes
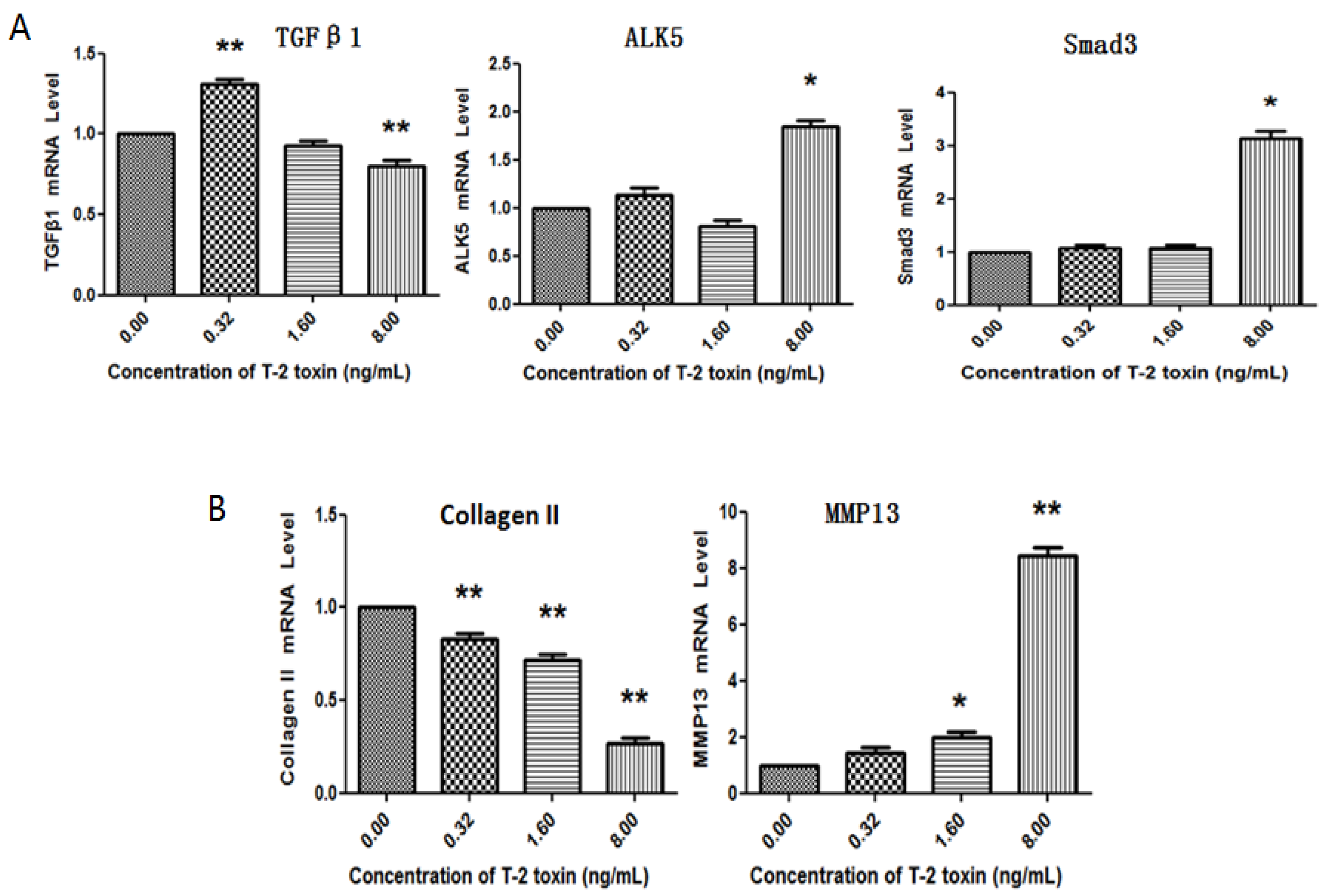
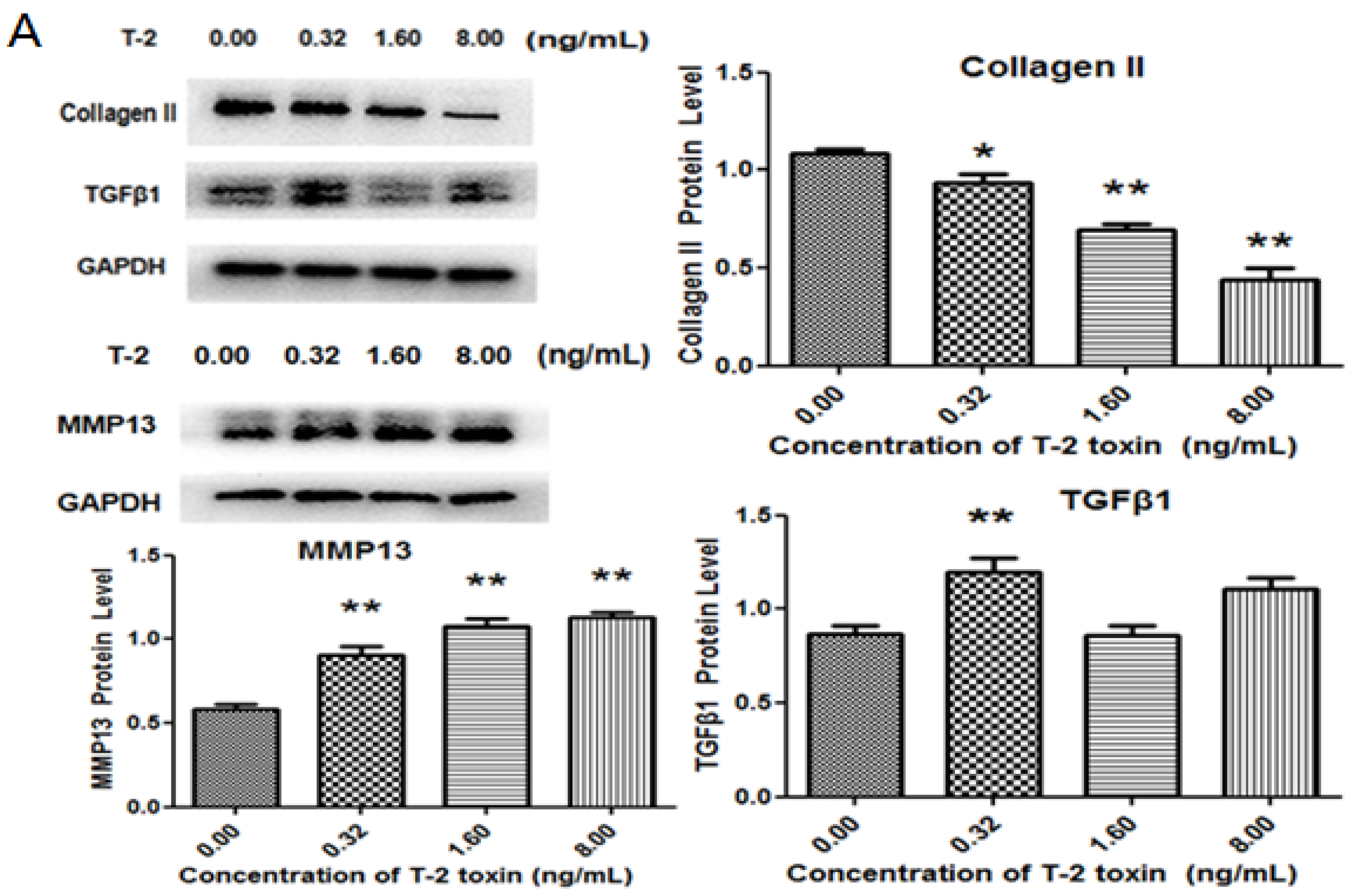
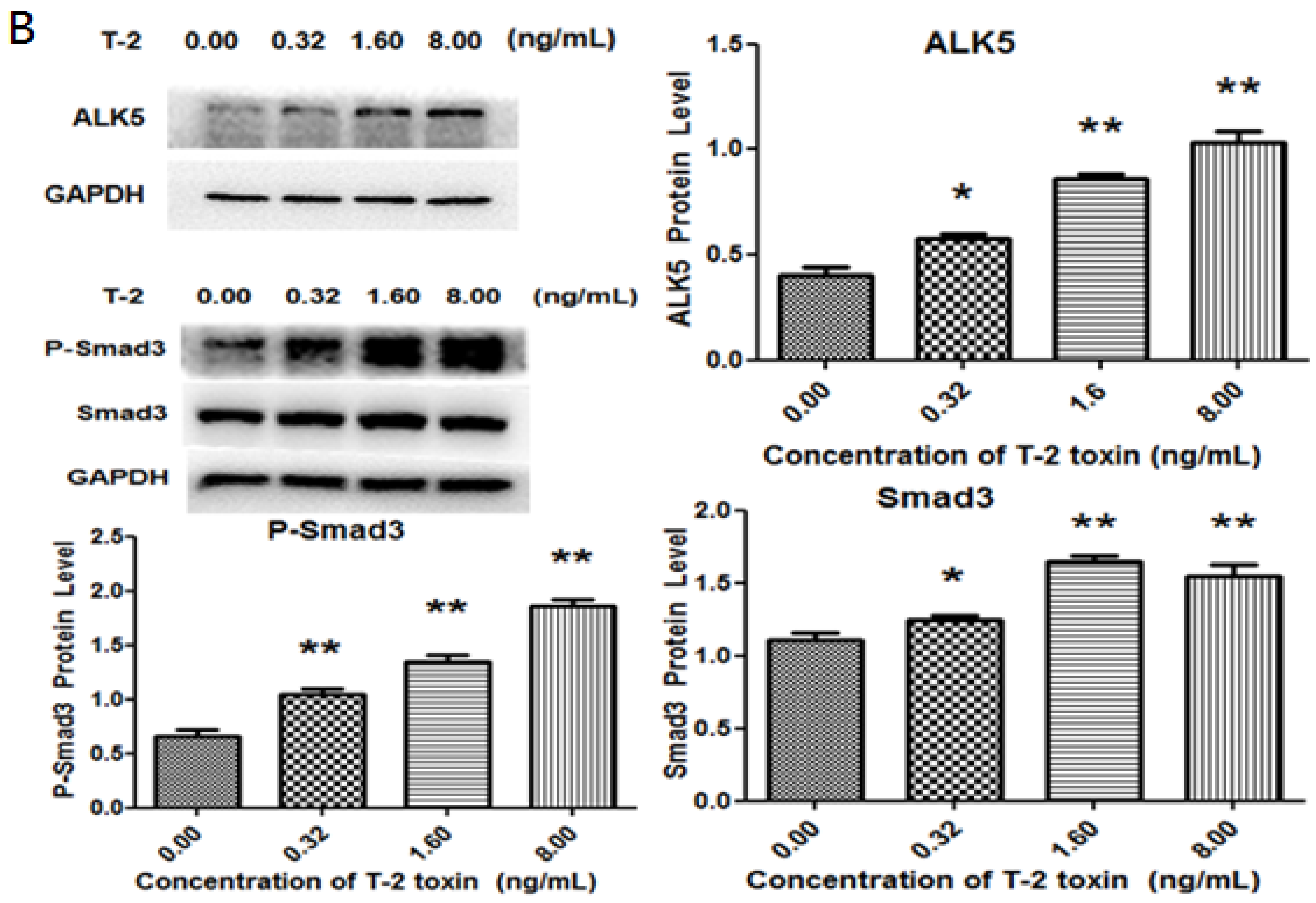
2.3. Effects of T-2 Toxin on Collagen Degradation-Related Proteins
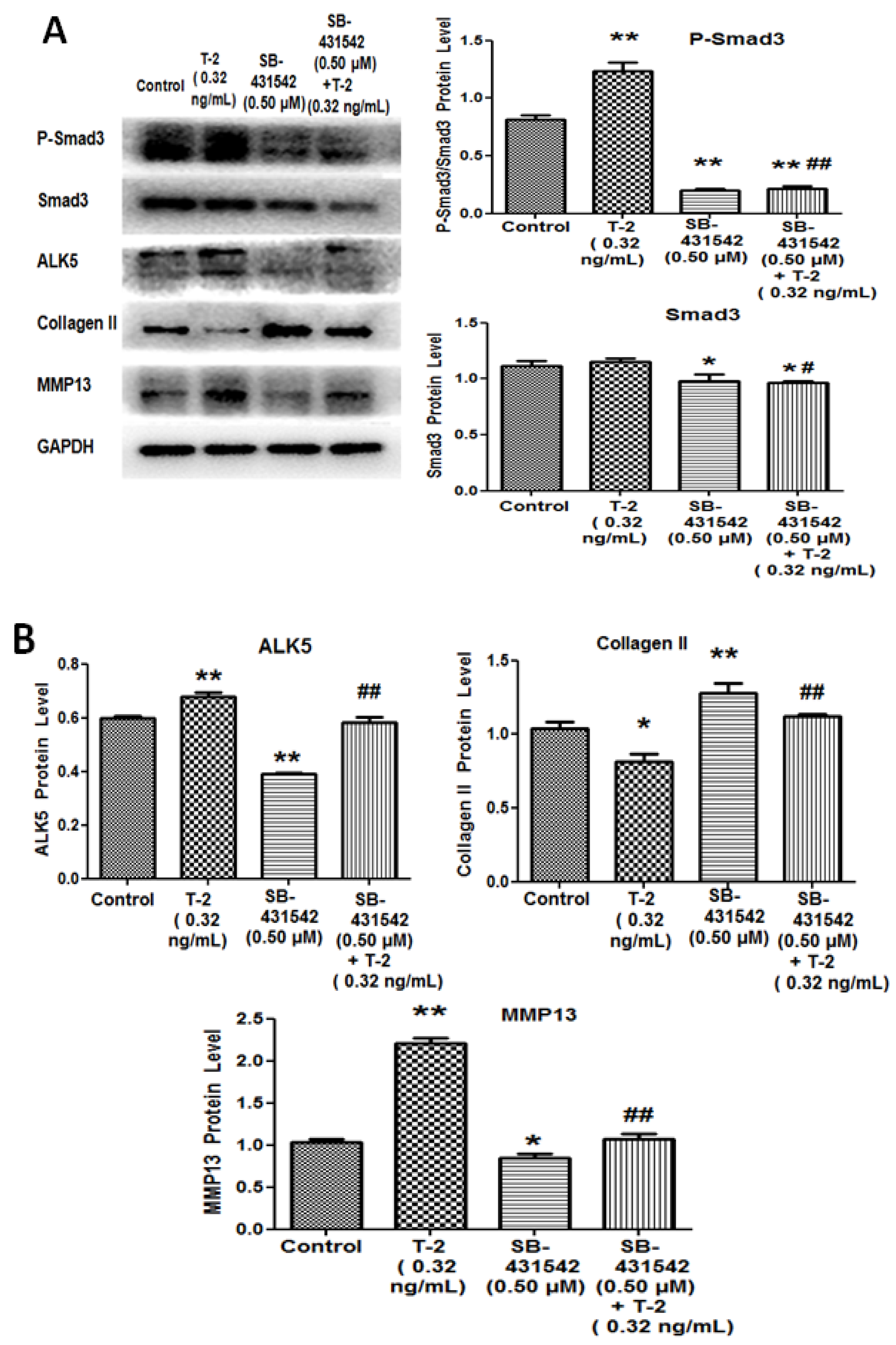
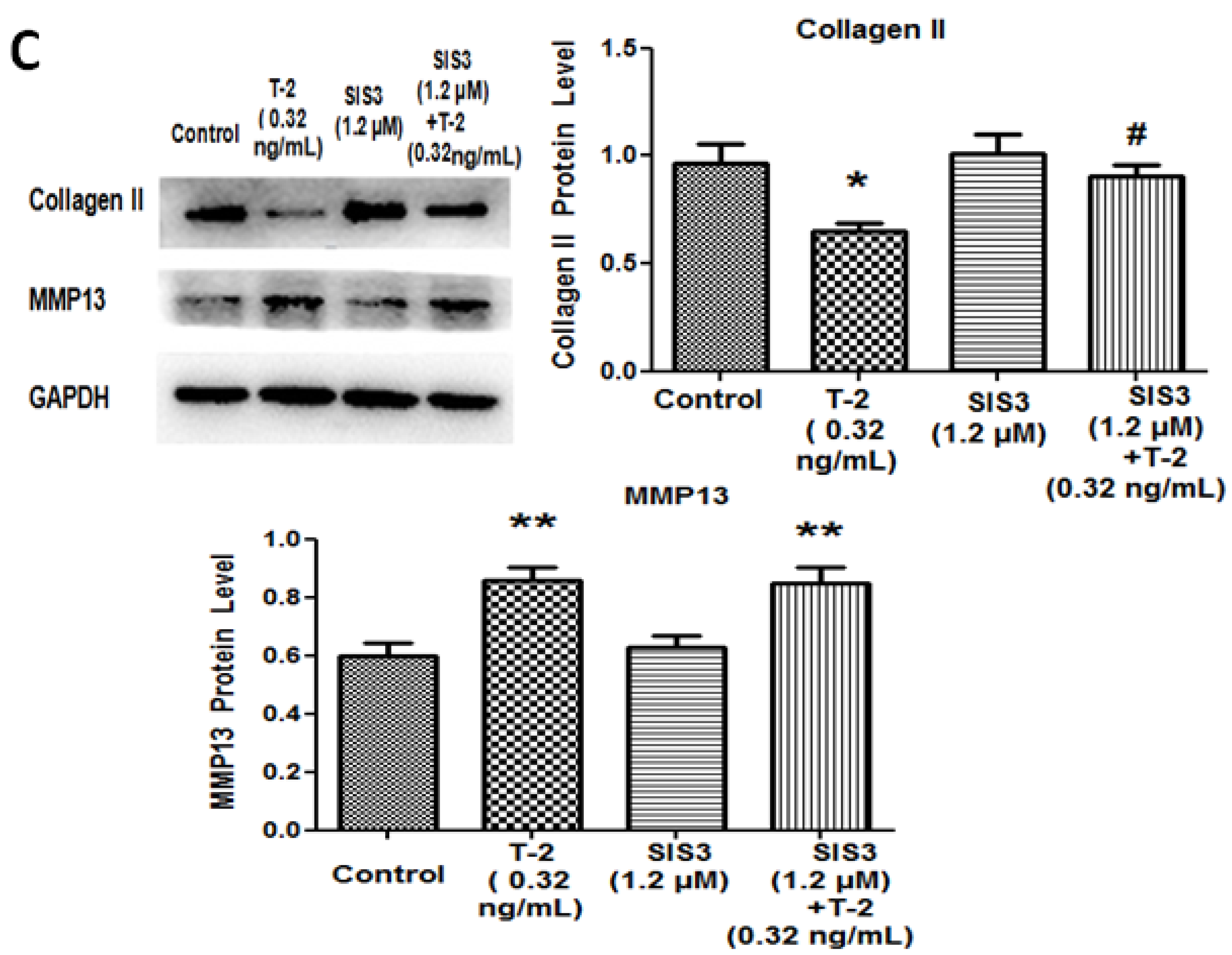
2.4. TGF-β1 Inhibitor and Smad3 Inhibitor Blocked the Effect of T-2 Toxin on Chondrocytes
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Immunofluorescence
4.3. Toluidine Blue Staining
4.4. Cell Viability Assay
4.5. Observation of Chondrocytes by Transmission Electron Microscopy
4.6. Real Time PCR
4.7. Western Blot
4.8. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Manning, B.B.; Li, M.H. Response of channel catfish to diets containing T-2 toxin. J. Aquat. Anim. Health 2003, 15, 229–238. [Google Scholar] [CrossRef]
- Guan, F.; Li, S.; Wang, Z.L.; Yang, H.; Xue, S.; Wang, W.; Song, D.; Zhou, X.; Zhou, W.; Chen, J.H.; et al. Histopathology of chondronecrosis development in knee articular cartilage in a rat model of Kashin-Beck disease using T-2 toxin and selenium deficiency conditions. Rheumatol. Int. 2013, 33, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.H.; Cao, J.L.; Chu, Y.L.; Yang, Z.T.; Shi, Z.L.; Wang, H.L.; Guo, X.; Wang, Z.L. Protective effect of selenium against T-2 toxin-induced inhibition of chondrocyte aggrecan and collagen II synthesis. J. South. Med. Univ. 2006, 26, 381–385. [Google Scholar]
- Chen, J.; Chu, Y.; Cao, J.; Wang, W.; Liu, J.; Wang, J. Effects of T-2 toxin and selenium on chondrocyte expression of matrix metalloproteinases (MMP-1, MMP-13), α2-macroglobulin (α2M) and TIMPs. Toxicol. In Vitro 2011, 25, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Cao, J.; Liu, F.; Li, S.; Chen, J.; Fu, Q.; Zhang, Z.; Liu, J.; Luo, M.; Wang, J.; et al. The effects of mycotoxins and selenium deficiency on tissue-engineered cartilage. Cells Tissues Organs 2012, 196, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Luo, M.; Wang, W.; Zhang, Z.; He, Y.; Duance, V.C.; Hughes, C.E.; Caterson, B.; Cao, J. Altered proteolytic activity and expression of MMPs and aggrecanases and their inhibitors in Kashin-Beck disease. J. Orthop. Res. 2015, 33, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Bruna, A.; Darken, R.S.; Rojo, F.; Ocaña, A.; Peñuelas, S.; Arias, A.; Paris, R.; Tortosa, A.; Mora, J.; Baselga, J.; et al. High TGFbeta-Smad activity confers poor prognosis in glioma patients and promotes cell proliferation depending on the methylation of the PDGF-B gene. Cancer Cell 2007, 11, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Hu, B.; Gu, W.; Xu, L.; Wang, D.; Huang, H.J.; Cavenee, W.K.; Cheng, S.Y. Platelet-derived growth factor-B enhances glioma angiogenesis by stimulating vascular endothelial growth factor expression in tumor endothelia and by promoting pericyte recruitment. Am. J. Pathol. 2003, 162, 1083–1093. [Google Scholar] [CrossRef]
- Guo, X.; Zuo, H.; Cao, C.X.; Zhang, Y.; Geng, D.; Zhang, Z.T.; Zhang, Y.G.; von der Mark, K.; von der Mark, H. Abnormal expression of Col X, PTHrP, TGF-β, bFGF, and VEGF in cartilage with Kashin-Beck disease. J. Bone Miner. Metab. 2006, 24, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Massagué, J. Mechanisms of TGF-beta signaling from cell membrane to the nucleus. Cell 2003, 113, 685–700. [Google Scholar] [CrossRef]
- Macias, M.J.; Martin-Malpartida, P.; Massagué, J. Structural determinants of Smad function in TGF-β signaling. Trends Biochem. Sci. 2015, 40, 296–308. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Zhou, P.Y.; Ni, W.; Wei, W.; Ben, D.F.; Lu, W.; Xia, Z.F. Inhibition of activin receptor-like kinase 5 induces matrix metallopeptidase 9 expression and aggravates lipopolysaccharide-induced pulmonary injury in mice. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 1051–1059. [Google Scholar] [PubMed]
- Aref-Eshghi, E.; Liu, M.; Harper, P.E.; Doré, J.; Martin, G.; Furey, A.; Green, R.; Rahman, P.; Zhai, G. Overexpression of MMP13 in human osteoarthritic cartilage is associated with the SMAD-independent TGF-β signalling pathway. Arthritis Res. Ther. 2015, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albarenque, S.M.; Shinozuka, J.; Suzuki, K.; Nakayama, H.; Doi, K. Kinetics and distribution of transforming growth factor (TGF)-beta 1 mRNA in the dorsal skin of hypotrichotic WBN/ILA-Ht rats following topical application of T-2 toxin. Exp. Toxicol. Pathol. 2000, 52, 297–301. [Google Scholar] [PubMed]
- Wang, J.; Ma, J.; Gu, J.H.; Wang, F.Y.; Shang, X.S.; Tao, H.R.; Wang, X. Regulation of type II collagen, matrix metalloproteinase-13 and cell proliferation by interleukin-1β is mediated by curcumin via inhibition of NF-κB signaling in rat chondrocytes. Mol. Med. Rep. 2017, 16, 1837–1845. [Google Scholar] [CrossRef] [PubMed]
- Pullig, O.; Kladny, B.; Weseloh, G.; Swoboda, B. Metabolic activation of chondrocytes in human osteoarthritis. Expression of type II collagen. Z. Orthop. Ihre Grenzgeb. 1999, 137, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Shen, J.; Jin, H.; Im, H.J.; Sandy, J.; Chen, D. Recent progress in understanding molecular mechanisms of cartilage degeneration during osteoarthritis. Ann. N. Y. Acad. Sci. 2011, 1240, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chu, Y.; Cao, J.; Yang, Z.; Guo, X.; Wang, Z. T-2 toxin induces apoptosis, and selenium partly blocks, T-2 toxin induced apoptosis in chondrocytes through modulation of the Bax/Bcl-2 ratio. Food Chem. Toxicol. 2006, 44, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.J.; Guo, X.; Zuo, H.; Zhang, Y.G.; Xu, P.; Ping, Z.G.; Zhang, Z.T.; Geng, D. Chondrocytc apoptosis and expression of Bcl-2, Bax, Fas, and iNOS in articular cartilage in patients with Kaschin-Beck disease. J. Rheumatol. 2006, 33, 615–619. [Google Scholar] [PubMed]
- Wang, L.H.; Fu, Y.; Shi, Y.X.; Wang, W.G. T-2 toxin induces degenerative articular changes in rodents: Link to Kaschin-Beck disease. Toxicol. Pathol. 2011, 39, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Serra, R.; Johnson, M.; Filvaroff, E.H.; LaBorde, J.; Sheehan, D.M.; Derynck, R.; Moses, H.L. Expression of a truncated, kinase-defective TGF-beta type II receptor in mouse skeletal tissue promotes terminal chondrocyte differentiation and osteoarthritis. J. Cell Biol. 1997, 139, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Letterio, J.J.; Lechleider, R.J.; Chen, L.; Hayman, R.; Gu, H.; Roberts, A.B.; Deng, C. Targeted disruption of SMAD3 results in impaired mucosal immunity and diminished T cell responsiveness to TGF-β. EMBO J. 1999, 18, 1280–1291. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, Y.; Chang, Y.; Duan, D.; Sun, Z.; Guo, X. Elevation of IGFBP2 contributes to mycotoxin T-2-induced chondrocyte injury and metabolism. Biochem. Biophys. Res. Commun. 2016, 478, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Kang, P.; Yang, J.; Shen, B.; Zhou, Z.; Duan, L.; Deng, J.; Huang, H.; Pei, F.X. The effect of Kashin-Beck disease-affected feed and T-2 toxin on the bone development of Wistar rats. Int. J. Rheum. Dis. 2010, 13, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.-Y.; Wang, Y.; An, J.; Mao, C.-M.; Hou, N.; Lv, Y.-X.; Wang, Y.-L.; Cui, F.; Huang, M.; Yang, X. Mutation analysis of the Smad3 gene in human osteoarthritis. Eur. J. Hum. Genet. 2003, 11, 714–717. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, J.; Serra, R. Unique and redundant roles of Smad3 in TGF-beta-mediated regulation of long bone development in organ culture. Dev. Dyn. 2004, 230, 685–699. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | mRNA | Primer Sequence (5′-3′) |
|---|---|---|
| 1 | TGFβ1 | F:GGAAGGACCTGGGTTGGAAG |
| R:GTAGTAGACGATGGGCAGTGG | ||
| 2 | ALK5 | F:AGGATGACTTACAGAGGCTTAGAC |
| R:TGATACCCAGTGACTCAAGGAAG | ||
| 3 | Smad3 | F:AGTTCTCCAGAGTTAAAAGCGA |
| R:CTTGACCGCCTTCTCGCA | ||
| 4 | Type II collagen | F:GCCAGGATGCCCGAAAATTAG |
| R:GTCACCTCTGGGTCCTTGTTC | ||
| 5 | MMP13 | F:TGCATACGAGCATCCATCCC |
| R:CTCAAAGTGAACCGCAGCAC | ||
| 6 | GAPDH | F:AACTCCCATTCTTCCACCTTTG |
| R:CTCTTGCTCTCAGTATCCTTGC |
| No. | Antibody | MW (kDa) | Dilution Ratio | Sources (Company) |
|---|---|---|---|---|
| 1 | TGF-β1 | 44 | 1:500 | Proteintech |
| 2 | ALK5 | 56 | 1:1000 | abcam |
| 3 | P-Smad3 | 52 | 1:1000 | Cell Signaling TECHNOLOGY |
| 4 | Smad3 | 48 | 1:3000 | abcam |
| 5 | Type II collagen | 142 | 1:10,000 | abcam |
| 6 | MMP13 | 54 | 1:3000 | abcam |
| 7 | GAPDH | 37 | 1:1000 | GOOD HERE |
| 8 | Peroxidase-conjugated Afinipure goat anti-rabbit IgG | 0.8 mg/mL | 1:25,000 | ZSGB-BIO |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Zou, N.; Wang, J.; Wang, K.-W.; Li, F.-Y.; Chen, F.-X.; Sun, B.-Y.; Sun, D.-J. TGF-β1/Smad3 Signaling Pathway Mediates T-2 Toxin-Induced Decrease of Type II Collagen in Cultured Rat Chondrocytes. Toxins 2017, 9, 359. https://doi.org/10.3390/toxins9110359
Li Y, Zou N, Wang J, Wang K-W, Li F-Y, Chen F-X, Sun B-Y, Sun D-J. TGF-β1/Smad3 Signaling Pathway Mediates T-2 Toxin-Induced Decrease of Type II Collagen in Cultured Rat Chondrocytes. Toxins. 2017; 9(11):359. https://doi.org/10.3390/toxins9110359
Chicago/Turabian StyleLi, Yang, Ning Zou, Jing Wang, Ke-Wei Wang, Fu-Yuan Li, Fu-Xun Chen, Bing-Yu Sun, and Dian-Jun Sun. 2017. "TGF-β1/Smad3 Signaling Pathway Mediates T-2 Toxin-Induced Decrease of Type II Collagen in Cultured Rat Chondrocytes" Toxins 9, no. 11: 359. https://doi.org/10.3390/toxins9110359
APA StyleLi, Y., Zou, N., Wang, J., Wang, K. -W., Li, F. -Y., Chen, F. -X., Sun, B. -Y., & Sun, D. -J. (2017). TGF-β1/Smad3 Signaling Pathway Mediates T-2 Toxin-Induced Decrease of Type II Collagen in Cultured Rat Chondrocytes. Toxins, 9(11), 359. https://doi.org/10.3390/toxins9110359