
Ameliorative Effects of Grape Seed Proanthocyanidin Extract on Growth Performance, Immune Function, Antioxidant Capacity, Biochemical Constituents, Liver Histopathology and Aflatoxin Residues in Broilers Exposed to Aflatoxin B1

 ,
, 
 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Growth Performance
2.2. Serum Biochemistry
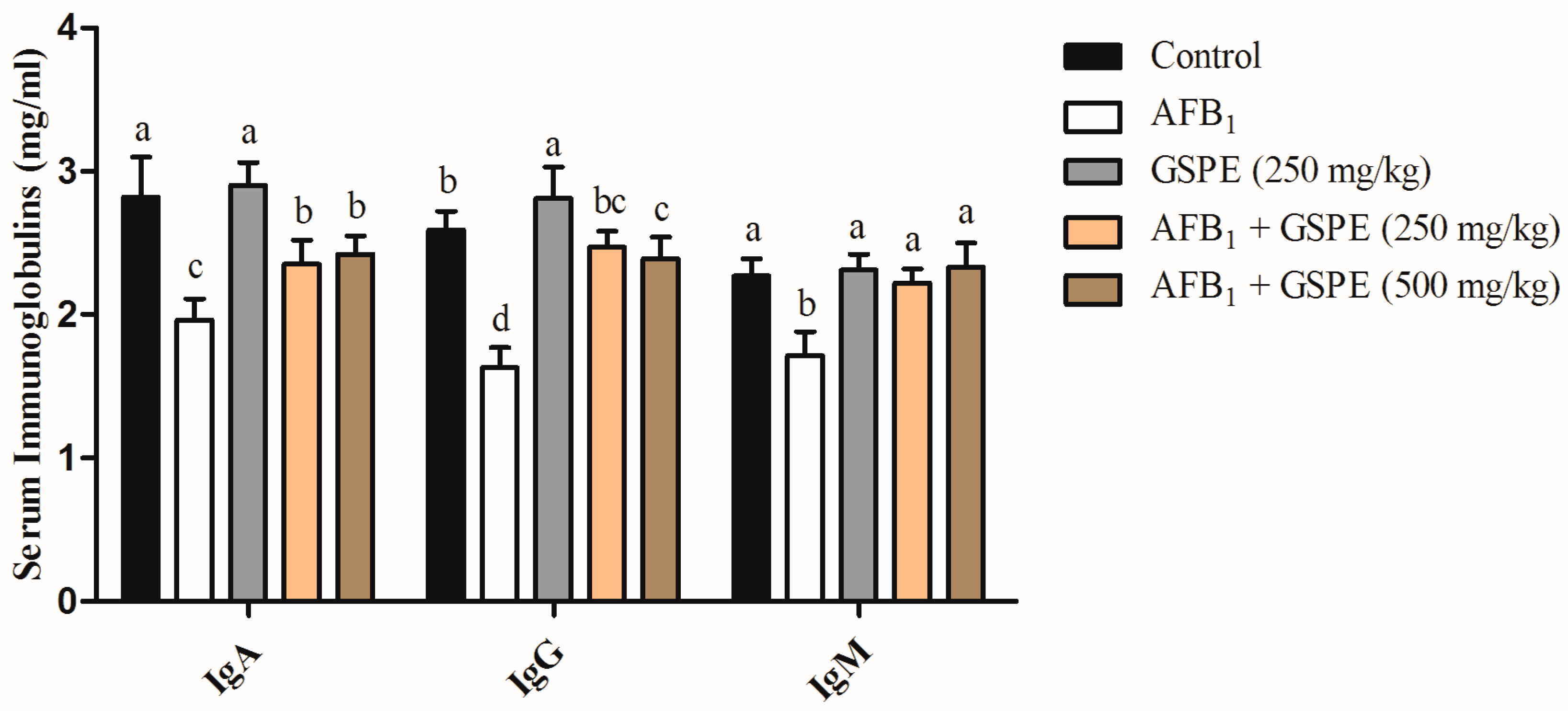
2.3. Serum Immunoglobulins
2.4. Serum Antioxidant Parameters
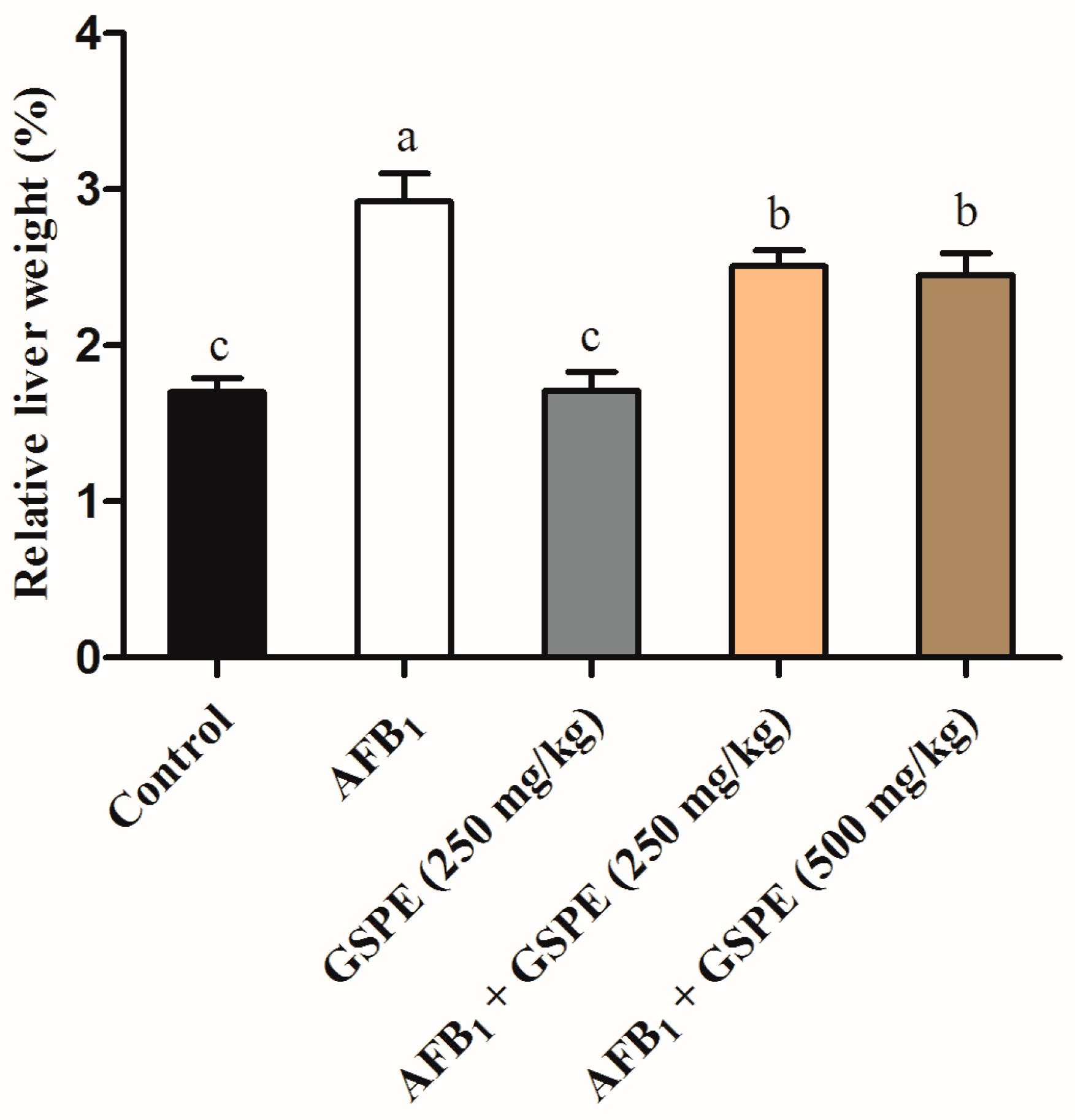
2.5. Histopathological Variations and Relative Weight of Liver
2.6. Hepatic Antioxidant Parameters
2.7. Aflatoxin B1 Residues in Liver
3. Discussion
3.1. Growth Performance
3.2. Serum Biochemistry
3.3. Serum Immunoglobulins
3.4. Serum and Liver Antioxidant Parameters
3.5. Histopathological Variations and Relative Weight of Liver
3.6. AFB1 Residues
4. Conclusions
5. Material and Methods
5.1. Birds, Diets, and Management
5.2. Aflatoxin B1 Production and Analysis
5.3. Collection of Samples and Measurements
5.4. Serum Biochemical and Histopathological Analysis
5.5. Serum and Liver Antioxidant Enzymes Assays
5.6. Serum Concentrations of Immunoglobulins Analysis
5.7. Analysis of Aflatoxin Residues in Liver
5.8. Statistical Analysis
Acknowledgments
Authors Contributions
Conflicts of Interest
References
- Diaz, D.E.; Hagler, W.M.; Hopkins, B.A.; Whitlow, L.W. Aflatoxin binders I: In vitro binding assay for aflatoxin B1 by several potential sequestering agents. Mycopathologia 2003, 156, 223–226. [Google Scholar] [CrossRef]
- Alpsoy, L.; Yalvac, M.E. Key roles of vitamins A, C, and E in aflatoxin B1-induced oxidative stress. Vitam. Horm. 2010, 86, 287–305. [Google Scholar]
- Dalvi, R. An overview of aflatoxicosis of poultry: Its characteristics, prevention and reduction. Vet. Res. Commun. 1986, 10, 429–443. [Google Scholar] [CrossRef] [PubMed]
- Kermanshahi, H.; Hazegh, A.; Afzali, N. Effect of sodium bentonite in broiler chickens fed diets contaminated with aflatoxin B1. J. Anim. Vet. Adv. 2009, 8, 1631–1636. [Google Scholar]
- Magnoli, A.; Monge, M.; Miazzo, R.; Cavaglieri, L.; Magnoli, C.; Merkis, C.; Cristofolini, A.; Dalcero, A.; Chiacchiera, S. Effect of low levels of aflatoxin B1 on performance, biochemical parameters, and aflatoxin B1 in broiler liver tissues in the presence of monensin and sodium bentonite. Poult. Sci. 2011, 90, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Surai, P.F. Natural Antioxidants in Avian Nutrition and Reproduction; Nottingham University Press: Nottingham, UK, 2002. [Google Scholar]
- Shen, H.-M.; Shi, C.-Y.; Lee, H.-P.; Ong, C.-N. Aflatoxin B1-induced lipid peroxidation in rat liver. Toxicol. Appl. Pharmacol. 1994, 127, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Aflatoxins. IARC; International Agency for the Research on Cancer: Lyon, France, 2012; Volume 100F. [Google Scholar]
- Hussain, Z.; Khan, M.Z.; Khan, A.; Javed, I.; Saleemi, M.K.; Mahmood, S.; Asi, M.R. Residues of aflatoxin B1 in broiler meat: Effect of age and dietary aflatoxin B1 levels. Food Chem. Toxicol. 2010, 48, 3304–3307. [Google Scholar] [CrossRef] [PubMed]
- Bintvihok, A.; Kositcharoenkul, S. Effect of dietary calcium propionate on performance, hepatic enzyme activities and aflatoxin residues in broilers fed a diet containing low levels of aflatoxin B1. Toxicon 2006, 47, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.-Y.; Qi, M.; Zhao, L.; Zhu, M.-K.; Guo, J.; Liu, J.; Gu, C.-Q.; Rajput, S.A.; Krumm, C.S.; Qi, D.-S. Curcumin prevents aflatoxin B1 hepatoxicity by inhibition of cytochrome p450 isozymes in chick liver. Toxins (Basel) 2016, 8, 327. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ma, Q.; Zhao, L.; Guo, Y.; Duan, G.; Zhang, J.; Ji, C. Protective efficacy of alpha-lipoic acid against aflatoxinB1-induced oxidative damage in the liver. Asian-Australas. J. Anim. Sci. 2014, 27, 907. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Hamid, A.A.; Firgany, A.E.-D.L. Vitamin e supplementation ameliorates aflatoxin B1-induced nephrotoxicity in rats. Acta Histochem. 2015, 117, 767–779. [Google Scholar] [CrossRef] [PubMed]
- Zhen, J.; Qu, Z.; Fang, H.; Fu, L.; Wu, Y.; Wang, H.; Zang, H.; Wang, W. Effects of grape seed proanthocyanidin extract on pentylenetetrazole-induced kindling and associated cognitive impairment in rats. Int. J. Mol. Med. 2014, 34, 391–398. [Google Scholar] [CrossRef] [PubMed]
- El-Ashmawy, I.M.; Saleh, A.; Salama, O.M. Effects of marjoram volatile oil and grape seed extract on ethanol toxicity in male rats. Basic Clin. Pharmacol. Toxicol. 2007, 101, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.D.; Katiyar, S.K. Dietary grape seed proanthocyanidins inhibit uvb-induced cyclooxygenase-2 expression and other inflammatory mediators in uvb-exposed skin and skin tumors of SKH-1 hairless mice. Pharm. Res. 2010, 27, 1092–1102. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, D.; Sen, C.K.; Ray, S.D.; Das, D.K.; Bagchi, M.; Preuss, H.G.; Vinson, J.A. Molecular mechanisms of cardioprotection by a novel grape seed proanthocyanidin extract. Mutat. Res. 2003, 523, 87–97. [Google Scholar] [CrossRef]
- Demirkaya, E.; Avci, A.; Kesik, V.; Karslioglu, Y.; Oztas, E.; Kismet, E.; Gokcay, E.; Durak, I.; Koseoglu, V. Cardioprotective roles of aged garlic extract, grape seed proanthocyanidin, and hazelnut on doxorubicin-induced cardiotoxicity. Can. J. Physiol. Pharmacol. 2009, 87, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.-H.; Kim, H.J.; Jeong, I.; Hong, Y.J.; Kim, M.-J.; Rhie, D.-J.; Jo, Y.-H.; Hahn, S.J.; Yoon, S.H. Grape seed proanthocyanidin extract inhibits glutamate-induced cell death through inhibition of calcium signals and nitric oxide formation in cultured rat hippocampal neurons. BMC Neurosci. 2011, 12, 78. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Kemper, L.J.; Wang, J.; Zahs, K.R.; Ashe, K.H.; Pasinetti, G.M. Grape seed polyphenolic extract specifically decreases aβ* 56 in the brains of tg2576 mice. J. Alzheimers Dis. 2011, 26, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Sayin, V.I.; Ibrahim, M.X.; Larsson, E.; Nilsson, J.A.; Lindahl, P.; Bergo, M.O. Antioxidants accelerate lung cancer progression in mice. Sci. Transl. Med. 2014, 6, 221ra15. [Google Scholar] [CrossRef] [PubMed]
- Ouédraogo, M.; Charles, C.; Ouédraogo, M.; Guissou, I.P.; Stévigny, C.; Duez, P. An overview of cancer chemopreventive potential and safety of proanthocyanidins. Nutr. Cancer 2011, 63, 1163–1173. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, D.; Bagchi, M.; Stohs, S.J.; Das, D.K.; Ray, S.D.; Kuszynski, C.A.; Joshi, S.S.; Pruess, H.G. Free radicals and grape seed proanthocyanidin extract: Importance in human health and disease prevention. Toxicology 2000, 148, 187–197. [Google Scholar] [CrossRef]
- Xu, Z.-C.; Yin, J.; Zhou, B.; Liu, Y.-T.; Yu, Y.; Li, G.-Q. Grape seed proanthocyanidin protects liver against ischemia/reperfusion injury by attenuating endoplasmic reticulum stress. World J. Gastroenterol. 2015, 21, 7468–7477. [Google Scholar] [CrossRef] [PubMed]
- Long, M.; Yang, S.-H.; Han, J.-X.; Li, P.; Zhang, Y.; Dong, S.; Chen, X.; Guo, J.; Wang, J.; He, J.-B. The protective effect of grape-seed proanthocyanidin extract on oxidative damage induced by zearalenone in kunming mice liver. Int. J. Mol. Sci. 2016, 17, 808. [Google Scholar] [CrossRef] [PubMed]
- Long, M.; Zhang, Y.; Li, P.; Yang, S.-H.; Zhang, W.-K.; Han, J.-X.; Wang, Y.; He, J.-B. Intervention of grape seed proanthocyanidin extract on the subchronic immune injury in mice induced by aflatoxin B1. Int. J. Mol. Sci. 2016, 17, 516. [Google Scholar] [CrossRef] [PubMed]
- Dai, N.; Zou, Y.; Zhu, L.; Wang, H.-F.; Dai, M.-G. Antioxidant properties of proanthocyanidins attenuate carbon tetrachloride (CCL4)–induced steatosis and liver injury in rats via CYP2E1 regulation. J. Med. Food 2014, 17, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Sano, T.; Oda, E.; Yamashita, T.; Naemura, A.; Ijiri, Y.; Yamakoshi, J.; Yamamoto, J. Anti-thrombotic effect of proanthocyanidin, a purified ingredient of grape seed. Thromb. Res. 2005, 115, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Engelbrecht, A.-M.; Mattheyse, M.; Ellis, B.; Loos, B.; Thomas, M.; Smith, R.; Peters, S.; Smith, C.; Myburgh, K. Proanthocyanidin from grape seeds inactivates the PI3-kinase/PKB pathway and induces apoptosis in a colon cancer cell line. Cancer Lett. 2007, 258, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Qiu, J.; Zhao, S.; You, B.; Ji, X.; Wang, Y.; Cui, X.; Wang, Q.; Gao, H. Grape seed proanthocyanidin extract alleviates ouabain-induced vascular remodeling through regulation of endothelial function. Mol. Med. Rep. 2012, 6, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Liu, G.; Hu, Z.; Li, X.; Yang, X.; Jiang, B.; Li, X. Grape seed proanthocyanidin extract protects from cisplatin-induced nephrotoxicity by inhibiting endoplasmic reticulum stress-induced apoptosis. Mol. Med. Rep. 2014, 9, 801–807. [Google Scholar] [CrossRef] [PubMed]
- Al-Sowayan, N.S.; Kishore, U. Prophylactic efficacy of a combination of proanthocyanidin and vitamin e on hepatotoxicity induced by doxorubicin in rats. Int. Res. J. Pharm. 2012, 2, 161–169. [Google Scholar]
- Mansouri, E.; Khorsandi, L.; Moaiedi, M.Z. Grape seed proanthocyanidin extract improved some of biochemical parameters and antioxidant disturbances of red blood cells in diabetic rats. Iran. J. Pharm. Res. 2015, 14, 329–334. [Google Scholar] [PubMed]
- Ortatatli, M.; Oğuz, H.; Hatipoğlu, F.; Karaman, M. Evaluation of pathological changes in broilers during chronic aflatoxin (50 and 100 ppb) and clinoptilolite exposure. Res. Vet. Sci. 2005, 78, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Gowda, N.K.; Ledoux, D.R.; Rottinghaus, G.E.; Bermudez, A.J.; Chen, Y.C. Antioxidant efficacy of curcuminoids from turmeric (Curcuma longa L.) powder in broiler chickens fed diets containing aflatoxin B1. Br. J. Nutr. 2009, 102, 1629–1634. [Google Scholar] [CrossRef] [PubMed]
- Gowda, N.; Ledoux, D.; Rottinghaus, G.; Bermudez, A.; Chen, Y. Efficacy of turmeric (curcuma longa), containing a known level of curcumin, and a hydrated sodium calcium aluminosilicate to ameliorate the adverse effects of aflatoxin in broiler chicks. Poult. Sci. 2008, 87, 1125–1130. [Google Scholar] [CrossRef] [PubMed]
- Bagherzadeh Kasmani, F.; Karimi Torshizi, M.; Allameh, A.; Shariatmadari, F. A novel aflatoxin-binding bacillus probiotic: Performance, serum biochemistry, and immunological parameters in japanese quail. Poult. Sci. 2012, 91, 1846–1853. [Google Scholar] [CrossRef] [PubMed]
- Osborne, D.; Huff, W.; Hamilton, P.; Burmeister, H. Comparison of ochratoxin, aflatoxin, and t-2 toxin for their effects on selected parameters related to digestion and evidence for specific metabolism of carotenoids in chickens 1, 2. Poult. Sci. 1982, 61, 1646–1652. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Wahhab, M.; Nada, S.; Amra, H. Effect of aluminosilicates and bentonite on aflatoxin-induced developmental toxicity in rat. J. Appl. Toxicol. 1999, 19, 199–204. [Google Scholar] [CrossRef]
- Oguz, H.; Kececi, T.; Birdane, Y.; Önder, F.; Kurtoglu, V. Effect of clinoptilolite on serum biochemical and haematological characters of broiler chickens during aflatoxicosis. Res. Vet. Sci. 2000, 69, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Xu, Z.; Feng, J.; Wang, C. Efficacy of modified montmorillonite nanocomposite to reduce the toxicity of aflatoxin in broiler chicks. Anim. Feed Sci. Technol. 2006, 129, 138–148. [Google Scholar] [CrossRef]
- Wu, J.-W.; Lin, L.-C.; Hung, S.-C.; Lin, C.-H.; Chi, C.-W.; Tsai, T.-H. Hepatobiliary excretion of silibinin in normal and liver cirrhotic rats. Drug Metab. Dispos. 2007, 36, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Yunus, A.W.; Razzazi-Fazeli, E.; Bohm, J. Aflatoxin B1 in affecting broiler’s performance, immunity, and gastrointestinal tract: A review of history and contemporary issues. Toxins (Basel) 2011, 3, 566–590. [Google Scholar] [CrossRef] [PubMed]
- Miazzo, R.; Peralta, M.; Magnoli, C.; Salvano, M.; Ferrero, S.; Chiacchiera, S.; Carvalho, E.; Rosa, C.; Dalcero, A. Efficacy of sodium bentonite as a detoxifier of broiler feed contaminated with aflatoxin and fumonisin. Poult. Sci. 2005, 84, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ozer, J.; Ratner, M.; Shaw, M.; Bailey, W.; Schomaker, S. The current state of serum biomarkers of hepatotoxicity. Toxicology 2008, 245, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Fink-Gremmels, J. The role of mycotoxins in the health and performance of dairy cows. Vet. J. 2008, 176, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Meissonnier, G.M.; Pinton, P.; Laffitte, J.; Cossalter, A.-M.; Gong, Y.Y.; Wild, C.P.; Bertin, G.; Galtier, P.; Oswald, I.P. Immunotoxicity of aflatoxin B1: Impairment of the cell-mediated response to vaccine antigen and modulation of cytokine expression. Toxicol. Appl. Pharmacol. 2008, 231, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Fang, J.; Peng, X.; Cui, H.; Chen, J.; Wang, F.; Chen, Z.; Zuo, Z.; Deng, J.; Lai, W. Effect of selenium supplementation on aflatoxin B1-induced histopathological lesions and apoptosis in bursa of fabricius in broilers. Food Chem. Toxicol. 2014, 74, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Ma, Q.; Zhao, L.; Jia, R.; Zhang, J.; Ji, C.; Wang, X. Protective effects of sporoderm-broken spores of ganderma lucidum on growth performance, antioxidant capacity and immune function of broiler chickens exposed to low level of aflatoxin B1. Toxins (Basel) 2016, 8, 278. [Google Scholar] [CrossRef] [PubMed]
- Thaxton, J.; Tung, H.; Hamilton, P. Immunosuppression in chickens by aflatoxin 1, 2. Poult. Sci. 1974, 53, 721–725. [Google Scholar] [CrossRef] [PubMed]
- Ul-Hassan, Z.; Zargham Khan, M.; Khan, A.; Javed, I. Immunological status of the progeny of breeder hens kept on ochratoxin a (OTA)-and aflatoxin B1 (AFB1)-contaminated feeds. J. Immunotoxicol. 2012, 9, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-F.; Liu, J.; Wasser, S.; Shen, H.-M.; Tan, C.E.-L.; Ong, C.-N. Inhibition of ebselen on aflatoxin B1-induced hepatocarcinogenesis in fischer 344 rats. Carcinogenesis 2000, 21, 2237–2243. [Google Scholar] [CrossRef] [PubMed]
- Kodama, M.; Inoue, F.; Akao, M. Enzymatic and non-enzymatic formation of free radicals from aflatoxin B1. Free Radic. Res. Commun. 1990, 10, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.-C.; Lee, B.-S.; Chung, W.-T.; Choi, M.-S.; Lee, J.-C. Protective effects of apigenin and quercetin on aflatoxin B1-induced immunotoxicity in mice. Food Sci. Biotechnol. 2010, 19, 987–992. [Google Scholar] [CrossRef]
- Naaz, F.; Abdin, M.; Javed, S. Protective effect of esculin against prooxidant aflatoxin B1-induced nephrotoxicity in mice. Mycotoxin Res. 2014, 30, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Yener, Z.; Celik, I.; Ilhan, F.; Bal, R. Effects of urtica dioica l. Seed on lipid peroxidation, antioxidants and liver pathology in aflatoxin-induced tissue injury in rats. Food Chem. Toxicol. 2009, 47, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Zhao, L.; Ji, C.; Li, X.; Jia, R.; Xi, L.; Zhang, J.; Ma, Q. Protective effects of bacillus subtilis ansb060 on serum biochemistry, histopathological changes and antioxidant enzyme activities of broilers fed moldy peanut meal naturally contaminated with aflatoxins. Toxins (Basel) 2015, 7, 3330–3343. [Google Scholar] [CrossRef] [PubMed]
- Zuo, R.-Y.; Chang, J.; Yin, Q.-Q.; Wang, P.; Yang, Y.-R.; Wang, X.; Wang, G.-Q.; Zheng, Q.-H. Effect of the combined probiotics with aflatoxin B1-degrading enzyme on aflatoxin detoxification, broiler production performance and hepatic enzyme gene expression. Food Chem. Toxicol. 2013, 59, 470–475. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Horn, N.; Applegate, T. Efficiency of hydrated sodium calcium aluminosilicate to ameliorate the adverse effects of graded levels of aflatoxin B1 in broiler chicks. Poult. Sci. 2014, 93, 2037–2047. [Google Scholar] [CrossRef] [PubMed]
- Taub, R. Liver regeneration: From myth to mechanism. Nat. Rev. Mol. Cell Biol. 2004, 5, 836. [Google Scholar] [CrossRef] [PubMed]
- Berry, C.L. The pathology of mycotoxins. J. Pathol. 1988, 154, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Denli, M.; Blandon, J.; Guynot, M.; Salado, S.; Perez, J. Effects of dietary afladetox on performance, serum biochemistry, histopathological changes, and aflatoxin residues in broilers exposed to aflatoxin B1. Poult. Sci. 2009, 88, 1444–1451. [Google Scholar] [CrossRef] [PubMed]
- Zaghini, A.; Martelli, G.; Roncada, P.; Simioli, M.; Rizzi, L. Mannanoligosaccharides and aflatoxin B1 in feed for laying hens: Effects on egg quality, aflatoxins B1 and M1 residues in eggs, and aflatoxin B1 levels in liver. Poult. Sci. 2005, 84, 825–832. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Heydt, M.; Abdel-Hadi, A.; Magan, N.; Geisen, R. Complex regulation of the aflatoxin biosynthesis gene cluster of aspergillus flavus in relation to various combinations of water activity and temperature. Int. J. Food Microbiol. 2009, 135, 231–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Sun, L.; Zhang, N.; Zhang, J.; Guo, J.; Li, C.; Rajput, S.A.; Qi, D. Effects of nutrients in substrates of different grains on aflatoxin B1 production by aspergillus flavus. Biomed. Res. Int. 2016, 2016, 7232858. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Zhang, N.; Sun, L.; Qi, D. Effects of different substrates and oils on aflatoxin B1 production by aspergillus parasiticus. Eur. Food Res. Technol. 2015, 240, 627–634. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Dietary Treatments | ||||
|---|---|---|---|---|---|
| Control | AFB1 (1 mg/kg) | GSPE (250 mg/kg) | AFB1 + GSPE (250 mg/kg) | AFB1 + GSPE (500 mg/kg) | |
| 1–2 weeks | |||||
| ADFI (g/day) | 52.56 ± 0.97 a | 40.31 ± 1.54 c | 51.7 ± 1.19 a | 45.35 ± 1.21 b | 45.91 ± 2.70 b |
| ADG (g/day) | 39.09 ± 1.43 a | 25.77 ± 1.31 c | 38.92 ± 1.44 a | 30.85 ± 0.82 b | 30.61 ± 1.99 b |
| FCR (feed:gain) | 1.35 ± 0.05 c | 1.57 ± 0.13 a | 1.33 ± 0.04 c | 1.47 ± 0.04 b | 1.50 ± 0.05 a,b |
| 3–4 weeks | |||||
| ADFI (g/day) | 104.95 ± 9.16 a | 71.64 ± 4.42 c | 106.24 ± 2.44 a | 80.72 ± 2.62 b | 83.74 ± 1.62 b |
| ADG (g/day) | 69.69 ± 2.90 b | 45.14 ± 3.98 d | 73.78 ± 1.27 a | 53.13 ± 1.57 c | 54.08 ± 1.74 c |
| FCR (feed:gain) | 1.51 ± 0.12 a,b | 1.59 ± 0.07 a | 1.44 ± 0.02 b | 1.52 ± 0.08 a,b | 1.55 ± 0.03 a |
| 1–4 weeks | |||||
| ADFI (g/day) | 78.75 ± 4.78 a | 55.98 ± 2.79 c | 78.80 ± 1.02 a | 63.04 ± 1.17 b | 64.82 ± 2.07 b |
| ADG (g/day) | 54.40 ± 1.75 b | 35.46 ± 1.71 d | 56.43 ± 1.25 a | 41.99 ± 1.10 c | 42.35 ± 1.66 c |
| FCR (feed:gain) | 1.45 ± 0.07 c | 1.58 ± 0.03 a | 1.40 ± 0.02 c | 1.50 ± 0.06 b | 1.53 ± 0.02 a,b |
| Parameters | Dietary Treatments | ||||
|---|---|---|---|---|---|
| Control | AFB1 | GSPE | AFB1 + GSPE | AFB1 + GSPE | |
| (1 mg/kg) | (250 mg/kg) | (250 mg/kg) | (500 mg/kg) | ||
| ALT (U/L) | 1.76 ± 0.13 b | 2.43 ± 0.18 a | 1.68 ± 0.10 b | 1.87 ± 0.14 b | 1.85 ± 0.17 b |
| AST (U/L) | 272.57 ± 24.34 c | 352.63 ± 30.23 a | 257.53 ± 14.37 c | 312.25 ± 19.67 b | 298.67 ± 16.50 b |
| GGT (U/L) | 20.45 ± 1.37 c | 29.57 ± 1.69 a | 20.95 ± 1.43 c | 26.38 ± 1.85 b | 24.62 ± 1.65 b |
| ALP (U/L) | 1499.75 ± 73.22 c | 1891.43 ± 144.70 a | 1481.93 ± 88.92 c | 1730.88 ± 136.48 b | 1608.38 ± 124.54 b,c |
| TP (g/L) | 29.57 ± 2.07 a | 18.32 ± 1.43 c | 29.45 ± 1.01 a | 24.57 ± 2.10 b | 23.08 ± 1.86 b |
| Albumin (g/L) | 15.50 ± 0.96 a | 10.33 ± 0.65 c | 15.92 ± 0.88 a | 13.05 ± 0.94 b | 13.13 ± 0.82 b |
| Globulin (g/L) | 12.57 ± 0.43 a | 9.01 ± 0.65 c | 12.61 ± 0.63 a | 11.82 ± 0.61 b | 12.02 ± 0.52 a,b |
| Parameters | Dietary Treatments | ||||
|---|---|---|---|---|---|
| Control | AFB1 | GSPE | AFB1 + GSPE | AFB1 + GSPE | |
| (1 mg/kg) | (250 mg/kg) | (250 mg/kg) | (500 mg/kg) | ||
| MDA, nmol/mL | 2.46 ± 0.19 c | 4.05 ± 0.44 a | 2.32 ± 0.18 c | 3.24 ± 0.32 b | 3.37 ± 0.29 b |
| T-SOD, U/mL | 157.52 ± 13.20 b | 108.91 ± 10.55 c | 182.57 ± 9.47 a | 150.53 ± 11.02 b | 154.29 ± 9.35 b |
| GSH-Px, U/mL | 1544.56 ± 62.50 a | 913.67 ± 82.97 c | 1629.54 ± 133.29 a | 1293.92 ± 100.44 b | 1345.37 ± 88.06 b |
| CAT, U/mL | 2.80 ± 0.24 a,b | 1.83 ± 0.17 d | 2.91 ± 0.25 a | 2.59 ± 0.16 b,c | 2.36 ± 0.21 c |
| GSH, mg/L | 5.76 ± 0.23 b | 3.02 ± 0.18 c | 7.26 ± 0.63 a | 6.24 ± 0.47 b | 5.93 ± 0.40 b |
| GR, U/L | 27.39 ± 2.75 b | 16.67 ± 1.43 d | 31.28 ± 2.64 a | 24.17 ± 1.55 c | 25.08 ± 2.26 b,c |
| GST, U/mL | 57.25 ± 3.00 a | 39.76 ± 3.50 c | 54.81 ± 4.48 a,b | 50.31 ± 4.03 b | 51.95 ± 3.69 b |
| Parameters | Dietary Treatments | ||||
|---|---|---|---|---|---|
| Control | AFB1 | GSPE | AFB1 + GSPE | AFB1 + GSPE | |
| (1 mg/kg) | (250 mg/kg) | (250 mg/kg) | (500 mg/kg) | ||
| MDA, nmol/mgprot | 0.96 ± 0.05 c,d | 1.59 ± 0.17 a | 0.82 ± 0.10 d | 1.09 ± 0.08 b,c | 1.14 ± 0.13 b |
| T-SOD, U/mgprot | 71.87 ± 4.59 a | 52.70 ± 7.04 b | 79.19 ± 2.75 a | 76.14 ± 6.67 a | 76.52 ± 8.50 a |
| GSH-Px, U/mgprot | 51.65 ± 4.66 b | 29.77 ± 3.56 d | 62.46 ± 5.94 a | 40.29 ± 3.54 c | 43.81 ± 4.98 c |
| CAT, U/mgprot | 86.79 ± 9.14 a | 57.47 ± 9.12 c | 95.46 ± 8.85 a | 75.22 ± 5.16 b | 73.34 ± 6.42 b |
| GSH, mg/gprot | 2.83 ± 0.18 b | 2.09 ± 0.20 c | 3.10 ± 0.22 a | 2.78 ± 0.18 b | 2.71 ± 0.23 b |
| GR, U/gprot | 6.65 ± 0.65 a | 3.18 ± 0.30 c | 7.03 ± 0.48 a | 4.04 ± 0.42 b | 3.68 ± 0.34 b,c |
| GST, U/mgprot | 25.28 ± 1.44 a | 19.03 ± 1.48 c | 26.49 ± 2.25 a | 21.96 ± 1.46 b | 22.82 ± 1.77 b |
| Ingredients | Percentage % |
| Corn | 58.3 |
| Soybean meal | 30.2 |
| Fish meal | 5.6 |
| Soybean oil | 2.3 |
| Dicalcium phosphate | 1.2 |
| Lime stone | 1.00 |
| Salt | 0.2 |
| Methionine | 0.2 |
| Premix 1 | 1.00 |
| Total | 100.00 |
| Calculated chemical composition | |
| Crude protein | 21.87 |
| Metabolisable energy (MJ/kg) | 13.45 |
| Lysine | 1.14 |
| Methionine | 0.40 |
| Methionine + Cystine | 0.94 |
| Calcium | 0.95 |
| Available phosphorus | 0.49 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali Rajput, S.; Sun, L.; Zhang, N.; Mohamed Khalil, M.; Gao, X.; Ling, Z.; Zhu, L.; Khan, F.A.; Zhang, J.; Qi, D. Ameliorative Effects of Grape Seed Proanthocyanidin Extract on Growth Performance, Immune Function, Antioxidant Capacity, Biochemical Constituents, Liver Histopathology and Aflatoxin Residues in Broilers Exposed to Aflatoxin B1. Toxins 2017, 9, 371. https://doi.org/10.3390/toxins9110371
Ali Rajput S, Sun L, Zhang N, Mohamed Khalil M, Gao X, Ling Z, Zhu L, Khan FA, Zhang J, Qi D. Ameliorative Effects of Grape Seed Proanthocyanidin Extract on Growth Performance, Immune Function, Antioxidant Capacity, Biochemical Constituents, Liver Histopathology and Aflatoxin Residues in Broilers Exposed to Aflatoxin B1. Toxins. 2017; 9(11):371. https://doi.org/10.3390/toxins9110371
Chicago/Turabian StyleAli Rajput, Shahid, Lvhui Sun, Niya Zhang, Mahmoud Mohamed Khalil, Xin Gao, Zhao Ling, Luoyi Zhu, Farhan Anwar Khan, Jiacai Zhang, and Desheng Qi. 2017. "Ameliorative Effects of Grape Seed Proanthocyanidin Extract on Growth Performance, Immune Function, Antioxidant Capacity, Biochemical Constituents, Liver Histopathology and Aflatoxin Residues in Broilers Exposed to Aflatoxin B1" Toxins 9, no. 11: 371. https://doi.org/10.3390/toxins9110371
APA StyleAli Rajput, S., Sun, L., Zhang, N., Mohamed Khalil, M., Gao, X., Ling, Z., Zhu, L., Khan, F. A., Zhang, J., & Qi, D. (2017). Ameliorative Effects of Grape Seed Proanthocyanidin Extract on Growth Performance, Immune Function, Antioxidant Capacity, Biochemical Constituents, Liver Histopathology and Aflatoxin Residues in Broilers Exposed to Aflatoxin B1. Toxins, 9(11), 371. https://doi.org/10.3390/toxins9110371