
Metabolism of Zearalenone and Its Major Modified Forms in Pigs


 , ,
, ,  and
and
Abstract
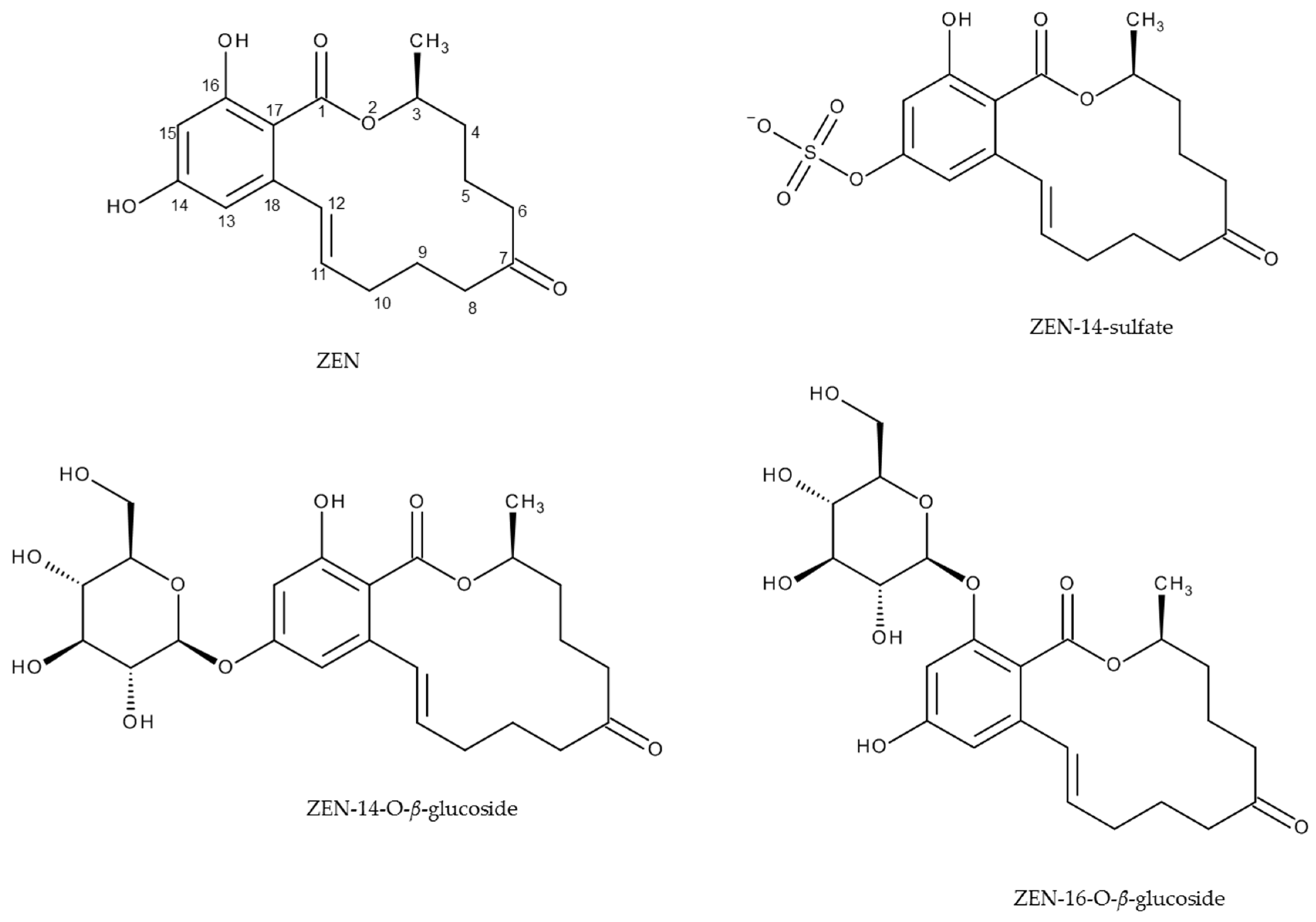
:1. Introduction
2. Results and Discussion
2.1. Clinical Symptoms
2.2. Sample Preparation and Method Performance
2.3. Amounts of Excreted Toxins and Biological Recoveries in Urine and Feces
2.3.1. Negative Control (Water Application)
2.3.2. Positive Control (ZEN Application)
2.3.3. Administration of ZEN-14-S
2.3.4. Administration of ZEN-14-Glc and ZEN-16-Glc
3. Conclusions
4. Materials and Methods
4.1. Chemicals and Standard Solutions
4.2. Animals and Study Design
4.3. Sample Preparation
4.4. HPLC-MS/MS Parameters
4.5. Method Validation
4.6. Analysis of Samples and Data Evaluation
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed]
- Vincelli, P.; Parker, G. Fumonisin, vomitoxin, and other mycotoxins in corn produced by Fusarium fungi. University of Kentucky, Cooperative Extension Service, 2002. ID 121. p. 8. Available online: http://www2.ca.uky.edu/agcomm/pubs/id/id121/id121.pdf (accessed on 22 December 2016).
- Dänicke, S.; Döll, S.; Goyarts, T.; Valenta, H.; Ueberschär, K.-H.; Flachowsky, G. Zur Beurteilung des Vorkommens der Fusarium-Toxine Deoxynivalenol (DON) und Zearalenon (ZON) sowie ihrer Metaboliten in physiologischen Substraten des Schweins. Tierärztliche Prax. 2008, 36, 35–47. [Google Scholar]
- Minervini, F.; Dell’Aquila, M.E. Zearalenone and reproductive function in farm animals. Int. J. Mol. Sci. 2008, 9, 2570–2584. [Google Scholar] [CrossRef] [PubMed]
- Dacasto, M.; Nachtmann, C.; Ceppa, L.; Nebbia, C. Zearalenone mycotoxicosis in piglets suckling sows fed contaminated grain. Vet. Hum. Toxicol. 1995, 37, 359–361. [Google Scholar] [PubMed]
- Richter, W.; Schuster, M.; Tischner, H.; Weigand, S.; Zimmermann, G.; Eder, J.; Doleschel, P.; Beck, R.; Lepschy, V.; Gleissenthall, J. Schimmelpilze und Mykotoxine in Futtermitteln (Futtergetreide, Grünfutter, Silage, Heu, Stroh). Vorkommen, Bewerten, Vermeiden. Länderübergreifende Zusammenarbeit der Landesanstalten für Landwirtschaft der Bundesländer Baden-Württemberg, Bayern, Sachsen und Thüringen. 2007, pp. 1–59. Available online: www.lfl.bayern.de/mam/cms07/ite/dateien/31386_schimmelpilze_und_mykotoxine_in_futtermitteln.pdf (accessed on 22 December 2016).
- EFSA Panel on Contaminants in the Food Chain. Scientific Opinion on the risks for human and animal health related to the presence of modified forms of certain mycotoxins in food and feed. EFSA J. 2014, 12, 3916. [Google Scholar]
- Berthiller, F.; Crews, C.; Dall’Asta, C.; de Saeger, S.; Haesaert, G.; Karlovsky, P.; Oswald, I.; Seefelder, W.; Speijers, G.; Stroka, J. Masked mycotoxins: A review. Mol. Nutr. Food Res. 2013, 57, 165–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rychlik, M.; Humpf, H.-U.; Marko, D.; Dänicke, S.; Mally, A.; Berthiller, F.; Klaffke, H.; Lorenz, N. Proposal of a comprehensive definition of modified and other forms of mycotoxins including “masked” mycotoxins. Mycotoxin Res. 2014, 30, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Kamimura, H. Conversion of zearalenone to zearalenone glycoside by Rhizopus. sp. Appl. Environ. Microbiol. 1986, 52, 515–519. [Google Scholar] [PubMed]
- Schneweis, I.; Meyer, K.; Engelhardt, G.; Bauer, J. Occurrence of zearalenone-4-β-D-glucopyranoside in wheat. J. Agric. Food Chem. 2002, 50, 1736–1738. [Google Scholar] [CrossRef] [PubMed]
- De Boevre, M.; Di Mavungu, J.D.; Landschoot, S.; Audenaert, K.; Eeckhout, M.; Maene, P.; Haesaert, G.; de Saeger, S. Natural occurrence of mycotoxins and their masked forms in food and feed products. World Mycotoxin J. 2012, 5, 207–219. [Google Scholar] [CrossRef]
- Poppenberger, B.; Berthiller, F.; Bachmann, H.; Lucyshyn, D.; Peterbauer, C.; Mitterbauer, R.; Schuhmacher, R.; Krska, R.; Glössl, J.; Adam, G. Heterologous expression of Arabidopsis UDP-glucosyltransferases in Saccharomyces cerevisiae for production of zearalenone-4-O-glucoside. Appl. Environ. Microbiol. 2006, 72, 4404–4410. [Google Scholar] [CrossRef] [PubMed]
- Kovalsky Paris, M.P.; Schweiger, W.; Hametner, C.; Stückler, R.; Muehlbauer, G.J.; Varga, E.; Krska, R.; Berthiller, F.; Adam, G. Zearalenone-16-O-glucoside: A new masked mycotoxin. J. Agric. Food Chem. 2014, 62, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Plasencia, J.; Mirocha, C.J. Isolation and characterization of zearalenone sulfate produced by Fusarium spp. Appl. Environ. Microbiol. 1991, 57, 146–150. [Google Scholar] [PubMed]
- El-Sharkawy, S.H.; Selim, M.I.; Afifi, M.S.; Halaweish, F.T. Microbial transformation of zearalenone to a zearalenone sulfate. Appl. Environ. Microbiol. 1991, 57, 549–552. [Google Scholar]
- Vendl, O.; Crews, C.; MacDonald, S.; Krska, R.; Berthiller, F. Occurrence of free and conjugated Fusarium mycotoxins in cereal-based food. Food Addit. Contam. 2010, 27, 1148–1152. [Google Scholar] [CrossRef] [PubMed]
- Drzymala, S.S.; Binder, J.; Brodehl, A.; Penkert, M.; Rosowski, M.; Garbe, L.-A.; Koch, M. Estrogenicity of novel phase I and phase II metabolites of zearalenone and cis-zearalenone. Toxicon 2015, 105, 10–12. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Commission recommendation (EC) of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding. Off. J. Eur. Union 2006, 229, 7–9. [Google Scholar]
- EFSA Panel on Contaminants in the Food Chain. Scientific Opinion on the risks for public health related to the presence of zearalenone in food. EFSA J. 2011, 9, 2197. [Google Scholar]
- EFSA Panel on Contaminants in the Food Chain. Appropriateness to set a group health-based guidance value for zearalenone and its modified forms. EFSA J. 2016, 14, 4425. [Google Scholar]
- Wang, J.P.; Chi, F.; Kim, I.H. Effects of montmorillonite clay on growth performance, nutrient digestibility, vulva size, faecal microflora, and oxidative stress in weaning gilts challenged with zearalenone. Anim. Feed Sci. Technol. 2012, 178, 158–166. [Google Scholar] [CrossRef]
- Thanner, S.; Czeglédi, L.; Schwartz-Zimmermann, H.E.; Berthiller, F.; Gutzwiller, A. Urinary deoxynivalenol (DON) and zearalenone (ZEA) as biomarkers of DON and ZEA exposure of pigs. Mycotoxin Res. 2016, 32, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Gambacorta, L.; Solfrizzo, M.; Visconti, A.; Powers, S.; Cossalter, A.M.; Pinton, P.; Oswald, I.P. Validation study on urinary biomarkers of exposure for aflatoxin B1, ochratoxin A, fumonisin B1, deoxynivalenol and zearalenone in piglets. World Mycotox. J. 2013, 6, 299–308. [Google Scholar] [CrossRef]
- Pfeiffer, E.; Hildebrand, A.; Mikula, H.; Metzler, M. Glucuronidation of zearalenone, zeranol and four metabolites in vitro: Formation of glucuronides by various microsomes and human UDP-glucuronosyltransferase isoforms. Mol. Nutr. Food Res. 2010, 54, 1468–1476. [Google Scholar] [CrossRef] [PubMed]
- Biehl, M.L.; Prelusky, D.B.; Koritz, G.D.; Hartin, K.E.; Buck, W.B.; Trenholm, H.L. Biliary excretion and enterohepatic cycling of zearalenone in immature pigs. Toxicol. Appl. Pharmcol. 1993, 121, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Olsen, M.; Pettersson, H.; Sandholm, K.; Visconti, A.; Kiessling, K.H. Metabolism of zearalenone by sow intestinal muscosa in vitro. Food Chem. Toxicol. 1987, 25, 681–683. [Google Scholar] [CrossRef]
- Malekinejad, H.; Maas-Bakker, R.; Fink-Gremmels, J. Species differences in the hepatic biotransformation of zearalenone. Vet. J. 2006, 172, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Döll, S.; Dänicke, S.; Ueberschär, K.-H.; Valenta, H.; Schnurrbusch, U.; Ganter, M.; Klobasa, F.; Flachowsky, G. Effects of graded levels of Fusarium toxin contaminated maize in diets for female weaned piglets. Arch. Anim. Nutr. 2003, 57, 311–334. [Google Scholar] [CrossRef]
- Zöllner, P.; Jodlbauer, J.; Kleinova, M.; Kahlbacher, H.; Kuhn, T.; Hochsteiner, W.; Lindner, W. Concentration levels of zearalenone and its metabolites in urine, muscle tissue, and liver samples of pigs fed with mycotoxin-contaminated oats. J. Agric. Food Chem. 2002, 50, 2494–2501. [Google Scholar] [CrossRef] [PubMed]
- Dänicke, S.; Swiech, E.; Buraczewska, L.; Ueberschär, K.-H. Kinetics and metabolism of zearalenone in young female pigs. J. Anim. Physiol. Anim. Nutr. 2005, 89, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Brezina, U.; Rempe, I.; Kersten, S.; Valenta, H.; Humpf, H.-U.; Dänicke, S. Diagnosis of intoxications of piglets fed with Fusarium toxin-contaminated maize by the analysis of mycotoxin residues in serum, liquor and urine with LC-MS/MS. Arch. Anim. Nutr. 2014, 68, 425–447. [Google Scholar] [CrossRef] [PubMed]
- Mirocha, C.J.; Pathre, S.V.; Robison, T.S. Comparative metabolism of zearalenone and transmission into bovine milk. Food Cosmet. Toxicol. 1981, 19, 25–30. [Google Scholar] [CrossRef]
- Mikula, H.; Hametner, C.; Berthiller, F.; Warth, B.; Krska, R.; Adam, G.; Fröhlich, J. Fast and reproducible chemical synthesis of zearalenone-14-β,D-glucuronide. World Mycotoxin J. 2012, 5, 289–296. [Google Scholar] [CrossRef]
- Olsen, M.E.; Pettersson, H.I.; Sandholm, K.A.; Kiessling, K.H. Quantitative liquid chromatographic method using fluorescence detection for determining zearalenone and its metabolites in blood plasma and urine. J. Assoc. Off. Anal. Chem. 1985, 68, 632–635. [Google Scholar] [PubMed]
- Dall’Erta, A.; Cirlini, M.; Dall’Asta, M.; Del Rio, D.; Galaverna, G.; Dall’Asta, C. Masked mycotoxins are efficiently hydrolyzed by human colonic microbiota releasing their aglycones. Chem. Res. Toxicol. 2013, 26, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Gratz, S.W.; Dinesh, R.; Yoshinari, T.; Holtrop, G.; Richardson, A.J.; Duncan, G.; MacDonald, S.; Lloyd, A.; Tarbin, J. Masked trichothecene and zearalenone mycotoxins withstand digestion and absorption in the upper GI tract but are efficiently hydrolyzed by human gut microbiota in vitro. Mol. Nutr. Food Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Dellafiora, L.; Perotti, A.; Galaverna, G.; Buschini, A.M.; Dall’Asta, C. On the masked mycotoxin zearalenone-14-glucoside. Does the mask truly hide? Toxicon 2016, 111, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Dellafiora, L.; Galaverna, G.; Righi, F.; Cozzini, P.; Dall’Asta, C. Assessing the hydrolytic fate of the masked mycotoxin zearalenone-14-glucoside—A warning light for the need to look at the “maskedome”. Food Chem. Toxicol. 2017, 99, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Gareis, M.; Bauer, J.; Thiem, J.; Plank, G.; Grabley, S.; Gedek, B. Cleavage of zearalenone-glycoside, a “masked” mycotoxin during digestion in swine. J. Vet. Med. B 1990, 37, 236–240. [Google Scholar] [CrossRef]
- Veršilovskis, A.; Geys, J.; Huybrechts, B.; Goossens, E.; de Saeger, S.; Callebaut, A. Simultaneous determination of masked forms of deoxynivalenol and zearalenone after oral dosing in rats by LC-MS/MS. World Mycotoxin J. 2012, 5, 303–318. [Google Scholar] [CrossRef]
- Michlmayr, H.; Varga, E.; Lupi, F.; Malachová, A.; Hametner, C.; Berthiller, F.; Adam, G. Synthesis of mono- and di-glucosides of zearalenone and α-/β-zearalenol by recombinant barley glucosyltransferase HvUGT14077. Toxins accepted for publication. 2017. [Google Scholar]
- Mikula, H.; Weber, J.; Lexmüller, S.; Bichl, G.; Schwartz, H.; Varga, E.; Berthiller, F.; Hametner, C.; Krska, R.; Fröhlich, J. Simultaneous preparation of α/β-zearalenol glucosides and glucuronides. Carbohydr. Res. 2013, 373, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Bundesgesetzblatt für die Republik Österreich. Teil I. 2012, 114, 1–22. Available online: https://www.ris.bka.gv.at/Dokumente/BgblAuth/BGBLA_2012_I_114/BGBLA_2012_I_114.pdf (accessed on 30 December 2016). [Google Scholar]
- Warth, B.; Sulyok, M.; Fruhmann, P.; Berthiller, F.; Schuhmacher, R.; Hametner, C.; Adam, G.; Fröhlich, J.; Krska, R. Assessment of human deoxynivalenol exposure using an LC-MS/MS based biomarker method. Toxicol. Lett. 2012, 211, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Sulyok, M.; Berthiller, F.; Krska, R.; Schuhmacher, R. Development and validation of a liquid chromatography/tandem mass spectrometric method for the determination of 39 mycotoxins in wheat and maize. Rapid Commun. Mass Spectrom. 2006, 20, 2649–2659. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Analyte | RA a ± RSD e (%) | SSE b ± RSD e (%) | RE c ± RSD e (%) | LOD d in Solution (ng/mL) | LOQ d in Solution (ng/mL) |
|---|---|---|---|---|---|
| α-ZEL f | 49 ± 14 | 47 ± 15 | 103 ± 12 | 0.08 | 0.28 |
| β-ZEL g | 104 ± 5 | 75 ± 16 | 139 ± 14 | 0.30 | 1.0 |
| ZEN h | 85 ± 9 | 77 ± 9 | 113 ± 11 | 0.10 | 0.33 |
| ZEN-14-Glc i | 75 ± 3 | 66 ± 4 | 114 ± 5 | 0.06 | 0.19 |
| ZEN-16-Glc j | 86 ± 5 | 75 ± 7 | 115 ± 6 | 0.73 | 2.4 |
| ZEN-14-S k | 124 ± 5 | 104 ± 5 | 119 ± 4 | 0.21 | 0.70 |
| Analyte | RA a ± RSD c (%) | LOD b in Solution (ng/mL) | LOQ b in Solution (ng/mL) |
|---|---|---|---|
| α-ZEL d | 89 ± 8 | 0.11 | 0.38 |
| β-ZEL e | 84 ± 9 | 0.16 | 0.54 |
| ZEN f | 95 ± 9 | 0.15 | 0.49 |
| ZEN-14-Glc g | 88 ± 9 | 0.07 | 0.23 |
| ZEN-16-Glc h | 96 ± 9 | 0.18 | 0.60 |
| ZEN-14-S i | 98 ± 10 | 0.02 | 0.08 |
| α-ZEL-GlcA j | 113 ± 6 | 0.73 | 2.4 |
| β-ZEL-GlcA k | 110 ± 7 | 0.42 | 1.4 |
| ZEN-14-GlcA l | 96 ± 9 | 0.30 | 1.0 |
| Average Amounts Excreted (nmol ± std. dev. a) b | ||||||
|---|---|---|---|---|---|---|
| Treatment | Matrix | Time Period | ZEN c | α-ZEL d | ZEN-14-GlcA e | Total f |
| ZEN | Feces | 0–24 h | 7 | 4 (n = 1) | n.d. g | 8 |
| 24–48 h | 24 | 13 | n.d. | 37 | ||
| 0–48 h | 31 ± 9 | 14 ± 6 | n.d. | 45 ± 14 | ||
| Urine | 0–24 h | 20 (n = 3) | n.d. | 62 (n = 3) | 62 | |
| 24–48 h | 13 (n = 3) | n.d. | 13 (n = 2) | 16 | ||
| 0–48 h | 33 ± 12 (n = 3) | n.d. | 53 ± 44 | 78 ± 31 | ||
| ZEN-14-S h | Urine | 0–24 h | 44 | 8 (n = 1) | 44 (n = 1) | 57 |
| 24–48 h | 11 | n.d. | n.d. | 11 | ||
| 0–48 h | 55 ± 19 | 8 (n = 1) | 44 (n = 1) | 68 ± 21 | ||
| ZEN-14-Glc i | Feces | 0–24 h | 40 | 21 (n = 2) | n.d. | 50 |
| 24–48 h | 48 | 29 | n.d. | 77 | ||
| 0–48 h | 88 ± 21 | 40 ± 19 | n.d. | 127 ± 32 | ||
| Urine | 0–24 h | 52 | 17 (n = 1) | 46 (n = 2) | 80 | |
| 24–48 h | n.d. | n.d. | 21 (n = 1) | 21 (n = 1) | ||
| 0–48 h | 52 ± 15 | 17 (n = 1) | 57 ± 45 (n = 2) | 85 ± 46 | ||
| ZEN–16–Glc j | Feces | 0–24 h | 25 | 12 (n = 2) | n.d. | 31 |
| 24–48 h | 36 | 16 | n.d. | 52 | ||
| 0–48 h | 62 ± 21 | 22 ± 7 | n.d. | 84 ± 23 | ||
| Urine | 0–24 h | 32 | 19 (n = 2) | 18 (n = 1) | 46 | |
| 24–48 h | 9 (n = 2) | n.d. | n.d. | 9 (n = 2) | ||
| 0–48 h | 36 ± 19 | 19 ± 3 (n = 2) | 18 (n = 1) | 51 ± 28 | ||
| Biological Recovery ± std. dev. (%) a,b | |||||
|---|---|---|---|---|---|
| Matrix | Time Period | ZEN c | ZEN-14-S d | ZEN-14-Glc e | ZEN-16-Glc f |
| Feces | 0–24 h | 3 ± 2 | n.d. g | 12 ± 13 | 9 ± 11 |
| 24–48 h | 12 ± 4 | n.d. | 17 ± 5 | 13 ± 7 | |
| 0–48 h | 14 ± 3 | n.d. | 29 ± 8 | 22 ± 7 | |
| Urine | 0–24 h | 20 ± 11 | 16 ± 5 | 18 ± 9 | 11 ± 6 |
| 24–48 h | 6 ± 2 | 3 ± <1 | 5 (n = 1) | 2 ± <1 (n = 2) | |
| 0–48 h | 26 ± 10 | 19 ± 5 | 19 ± 11 | 13 ± 7 | |
| Total | 0–24 h | 22 ± 10 | 16 ± 5 | 30 ± 9 | 20 ± 5 |
| 24–48 h | 18 ± 3 | 3 ± <1 | 18 ± 7 | 14 ± 7 | |
| 0–48 h | 40 ± 8 | 19 ± 5 | 48 ± 7 | 34 ± 3 | |
| Analyte | Retention Time (min) | [M − H]− (m/z) | DP c (V) | Product Ions (m/z) a | CE d (eV) | Relative Intensity b |
|---|---|---|---|---|---|---|
| ZEN e | 13.20 | 317.1 | −120 | 131.0/175.0 | −42/−34 | 0.70 |
| α-ZEL f | 11.22 | 319.1 | −125 | 160.0/275.1 | −30/−42 | 2.7 |
| β-ZEL g | 10.12 | 319.1 | −125 | 160.0/275.1 | −30/−42 | 2.6 |
| ZEN-14-Glc h | 8.80 | 479.1 | −125 | 317.0/175.0 | −22/−54 | 0.10 |
| ZEN-16-Glc i | 7.35 | 479.2 | −140 | 149.0/160.8 | −54/−54 | 0.49 |
| ZEN-14-S j | 8.89 | 397.0 | −115 | 317.0/131.0 | −34/−58 | 0.08 |
| ZEN-14-GlcA k | 8.50 | 493.1 | −115 | 317.0/175.0 | −36/−26 | 0.36 |
| α-ZEL-GlcA l | 7.23 | 495.1 | −110 | 319.0/112.8 | −38/−28 | 0.31 |
| β-ZEL-GlcA m | 6.13 | 495.1 | −110 | 319.0/112.8 | −38/−28 | 0.30 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Binder, S.B.; Schwartz-Zimmermann, H.E.; Varga, E.; Bichl, G.; Michlmayr, H.; Adam, G.; Berthiller, F. Metabolism of Zearalenone and Its Major Modified Forms in Pigs. Toxins 2017, 9, 56. https://doi.org/10.3390/toxins9020056
Binder SB, Schwartz-Zimmermann HE, Varga E, Bichl G, Michlmayr H, Adam G, Berthiller F. Metabolism of Zearalenone and Its Major Modified Forms in Pigs. Toxins. 2017; 9(2):56. https://doi.org/10.3390/toxins9020056
Chicago/Turabian StyleBinder, Sabina B., Heidi E. Schwartz-Zimmermann, Elisabeth Varga, Gerlinde Bichl, Herbert Michlmayr, Gerhard Adam, and Franz Berthiller. 2017. "Metabolism of Zearalenone and Its Major Modified Forms in Pigs" Toxins 9, no. 2: 56. https://doi.org/10.3390/toxins9020056
APA StyleBinder, S. B., Schwartz-Zimmermann, H. E., Varga, E., Bichl, G., Michlmayr, H., Adam, G., & Berthiller, F. (2017). Metabolism of Zearalenone and Its Major Modified Forms in Pigs. Toxins, 9(2), 56. https://doi.org/10.3390/toxins9020056