Variable Cyanobacterial Toxin and Metabolite Profiles across Six Eutrophic Lakes of Differing Physiochemical Characteristics

Abstract
:1. Introduction
2. Results
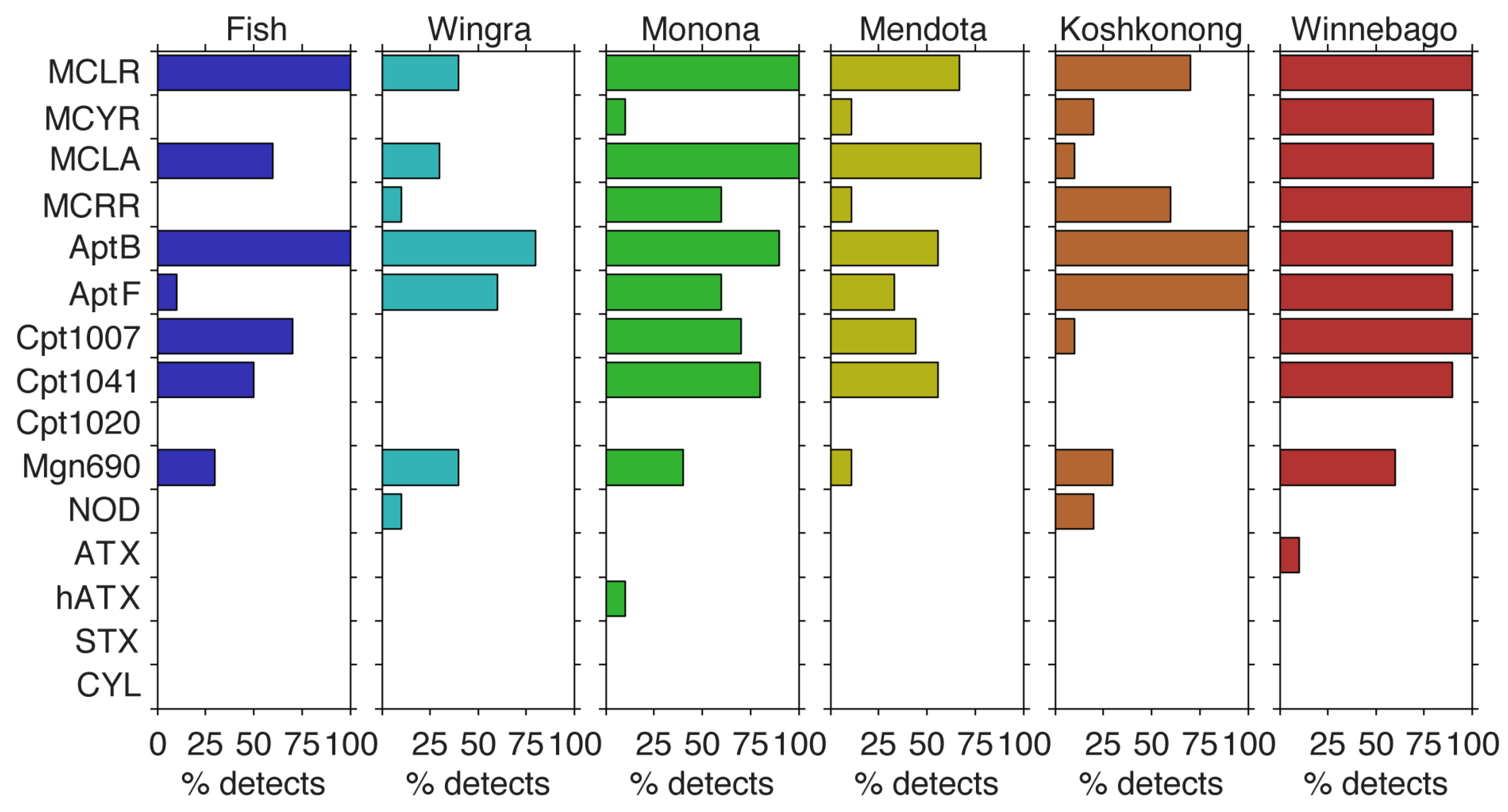
2.1. Metabolite Detects and Ranges Observed
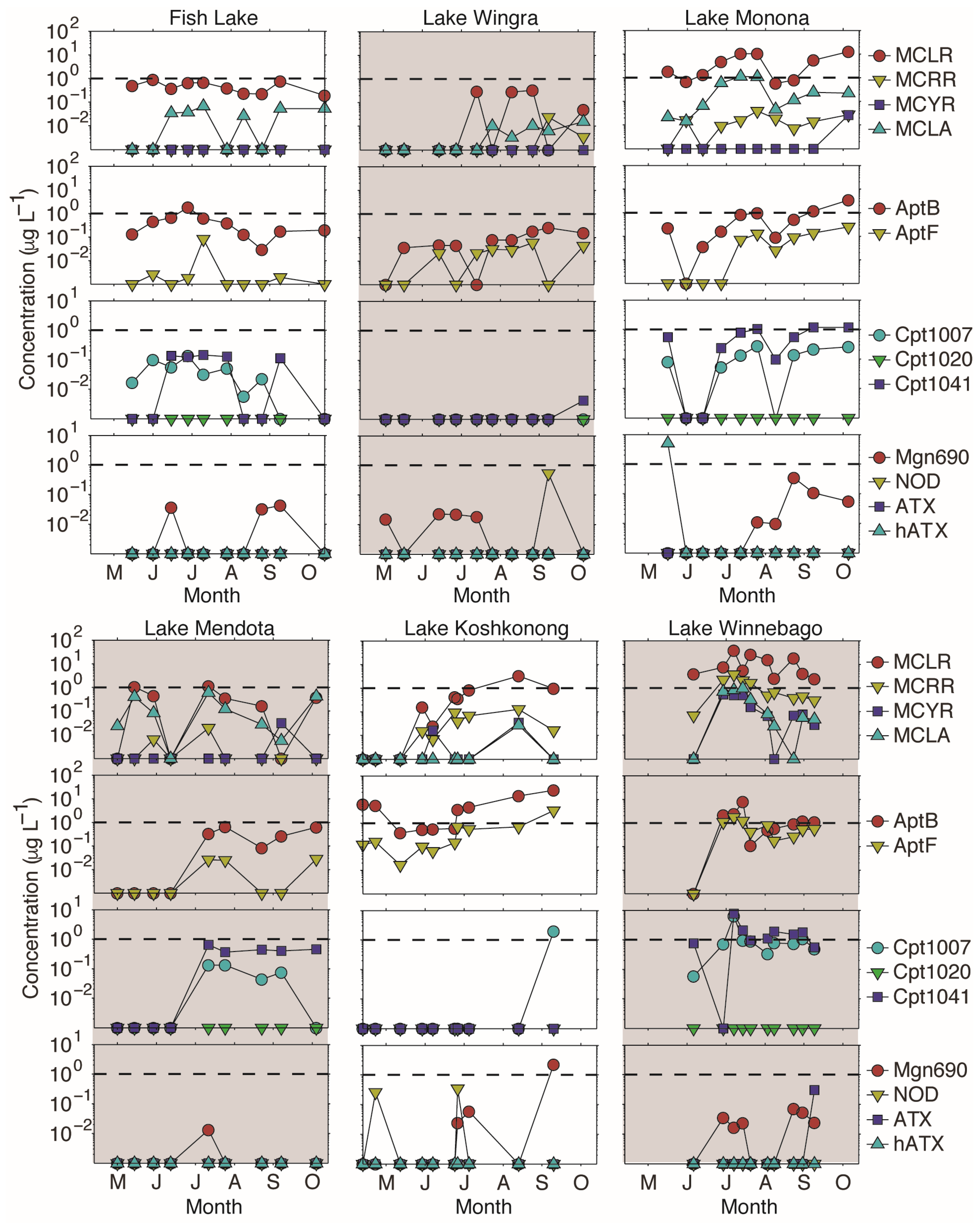
2.2. Intra-Lake Differences in Observed Metabolites
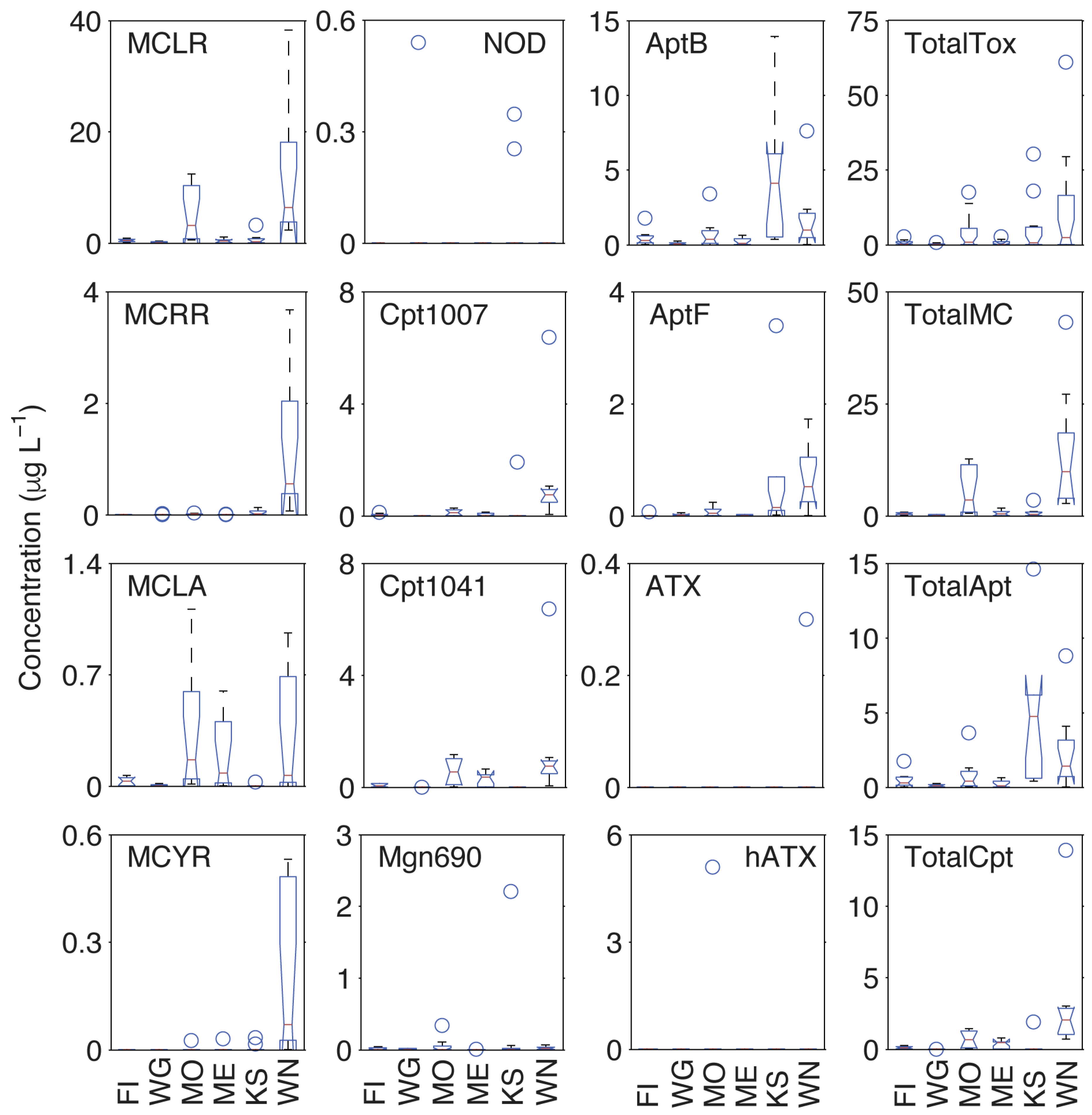
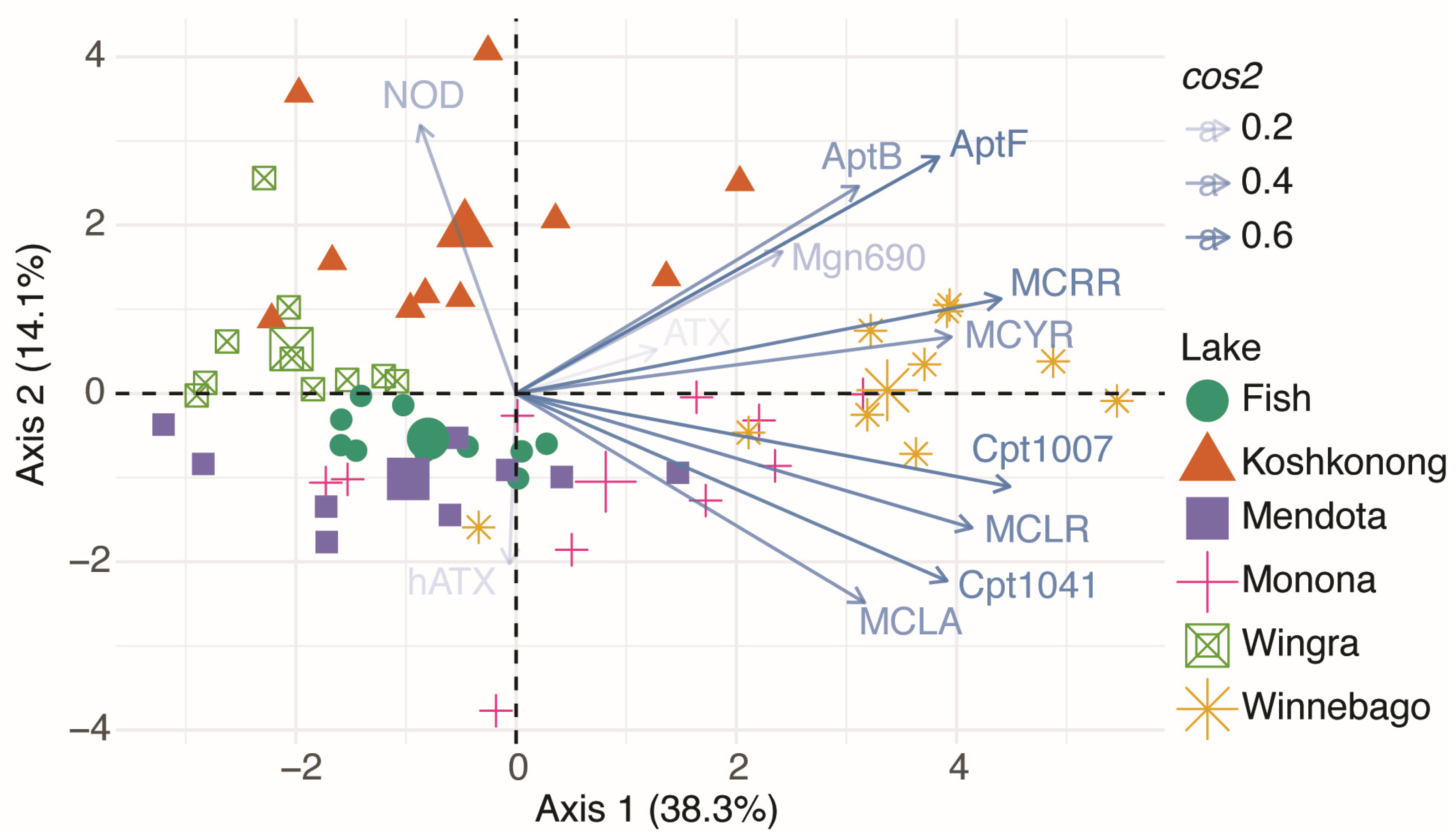
2.3. Inter-lake Differences in Observed Metabolites
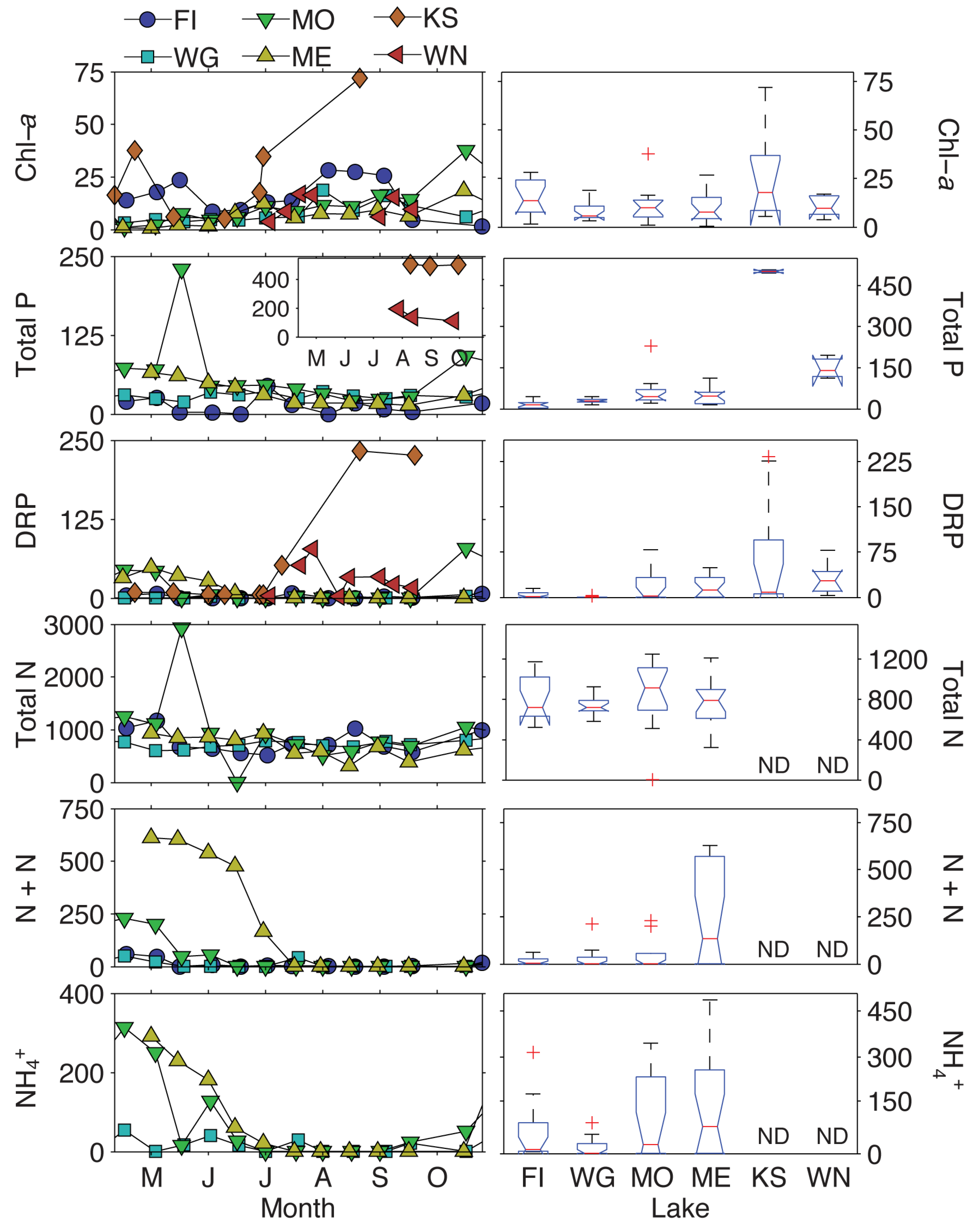
2.4. Physiochemical Characteristics
3. Discussion
4. Conclusions
- We measured 15 cyanobacterial compounds, nine of which are known human toxins, in six eutrophic lakes.
- Anabaenopeptins were the most frequently detected compounds, followed by microcystins and cyanopeptolins.
- In two highly populated lakes, one of which is used as a drinking water source, total microcystin concentrations were above 1 µg·L−1 in all samples collected.
- Nodularin, a more common marine toxin, was detected in the two shallowest lakes.
- Lake size and dissolved nitrogen and phosphorus concentrations (not TN and TP) were the most probable variables driving metabolite profiles.
- In order to determine the impact of these metabolites on ecosystem function and public health and manage them appropriately, similar studies are needed to elicit the eco-physiological role of these toxins.
5. Materials and Methods
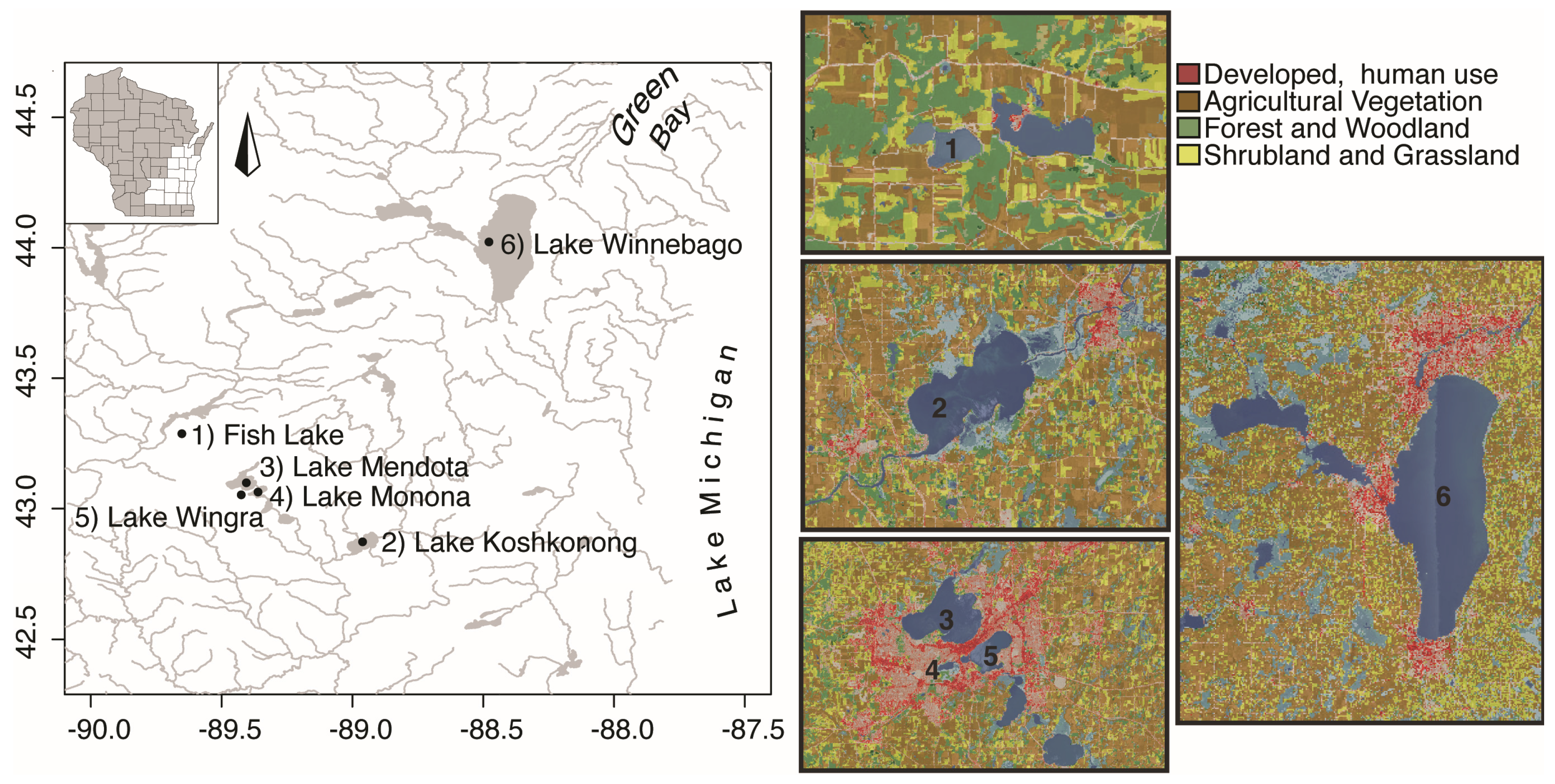
5.1. Study Sites
5.2. Sample Collection and Preservation
5.3. Chemicals
5.4. Chemical Extraction and Analysis
5.5. Ancillary Analytical Data and Measurements
5.6. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Welker, M.; Von Döhren, H. Cyanobacterial peptides—Nature’s own combinatorial biosynthesis. FEMS Microbiol. Rev. 2006, 30, 530–563. [Google Scholar] [CrossRef] [PubMed]
- Dittmann, E.; Fewer, D.P.; Neilan, B.A. Cyanobacterial toxins: Biosynthetic routes and evolutionary roots. FEMS Microbiol. Rev. 2013, 37, 23–43. [Google Scholar] [CrossRef] [PubMed]
- Balbus, J.M.; Boxall, A.B.A.; Fenske, R.A.; McKone, T.E.; Zeise, L. Implications of global climate change for the assessment and management of human health risks of chemicals in the natural environment. Environ. Toxicol. Chem. 2013, 32, 62–78. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Huisman, J. Climate change: A catalyst for global expansion of harmful cyanobacterial blooms. Environ. Microbiol. Rep. 2009, 1, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Chlipala, G.E.; Mo, S.; Orjala, J. Chemodiversity in freshwater and terrestrial cyanobacteria—A source for drug discovery. Curr. Drug Targets 2011, 12, 1654–1673. [Google Scholar] [CrossRef] [PubMed]
- Chorus, I.; Bartram, J. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; E & FN Spon: London, UK, 1999; p. 400. [Google Scholar]
- MacKintosh, C.; Beattie, K.A.; Klumpp, S.; Cohen, P.; Codd, G.A. Cyanobacterial microcystin-LR is a potent and specific inhibitor of protein phosphatases 1 and 2A from both mammals and higher plants. FEBS Lett. 1990, 264, 187–192. [Google Scholar] [CrossRef]
- Falconer, I.R. Health problems from exposure to cyanobacteria and proposed safety guidelines for drinking and recreational water. In Detection Methods for Cyanobacterial Toxins; Codd, G.A., Jefferies, T.M., Keevil, C.W., Potter, E., Eds.; Royal Society of Chemistry: Cambridge, UK, 1994; Volume 149, pp. 3–10. [Google Scholar]
- Fawell, J.; Mitchell, R.; Everett, D.; Hill, R. The toxicity of cyanobacterial toxins in the mouse: I microcystin-LR. Hum. Exp. Toxicol. 1999, 18, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-B.; Zhang, X.; Ju, J.; Li, Y.; Yin, L.; Pu, Y. Alterations in neurobehaviors and inflammation in hippocampus of rats induced by oral administration of microcystin-LR. Environ. Sci. Pollut. Res. 2014, 21, 12419–12425. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, J.A.; Zhao, Q.; Pu, C.; Qiu, Z.; Zhang, R.; Shu, W. A cross-sectional investigation of chronic exposure to microcystin in relationship to childhood liver damage in the three gorges reservoir region, China. Environ. Health Perspect. 2011, 119, 1483–1488. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Sheng, J.; Sha, J.; Han, X. The toxic effects of microcystin-LR on the reproductive system of male rats in vivo and in vitro. Reprod. Toxicol. 2008, 26, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Falconer, I.R.; Humpage, A.R. Tumour promotion by cyanobacterial toxins. Phycologia 1996, 35, 74–79. [Google Scholar] [CrossRef]
- Humpage, A.R.; Hardy, S.J.; Moore, E.J.; Froscio, S.M.; Falconer, I.R. Microcystins (cyanobacterial toxins) in drinking water enhance the growth of aberrant crypt foci in the mouse colon. J. Toxicol. Environ. Health Part A 2000, 61, 155–165. [Google Scholar] [PubMed]
- Nishiwaki-Matsushima, R.; Ohta, T.; Nishiwaki, S.; Suganuma, M.; Kohyama, K.; Ishikawa, T.; Carmichael, W.; Fujiki, H. Liver tumor promotion by the cyanobacterial cyclic peptide toxin microcystin-LR. J. Cancer Res. Clin. Oncol. 1992, 118, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Fleming, L.E.; Rivero, C.; Burns, J.; Williams, C.; Bean, J.A.; Shea, K.A.; Stinn, J. Blue green algal (cyanobacterial) toxins, surface drinking water, and liver cancer in Florida. Harmful Algae 2002, 1, 157–168. [Google Scholar] [CrossRef]
- Lun, Z.; Hai, Y.; Kun, C. Relationship between microcystin in drinking water and colorectal cancer. Biomed. Environ. Sci. 2002, 15, 166–171. [Google Scholar]
- Yu, S.; Zhao, N.; Zi, X. The relationship between cyanotoxin (microcystin, MC) in pond-ditch water and primary liver cancer in China. Zhonghua zhong liu za zhi [Chinese journal of oncology] 2001, 23, 96–99. [Google Scholar] [PubMed]
- Svirčev, Z.; Krstić, S.; Miladinov-Mikov, M.; Baltić, V.; Vidović, M. Freshwater cyanobacterial blooms and primary liver cancer epidemiological studies in Serbia. J. Environ. Sci. Health Part C 2009, 27, 36–55. [Google Scholar] [CrossRef] [PubMed]
- Faltermann, S.; Zucchi, S.; Kohler, E.; Blom, J.F.; Pernthaler, J.; Fent, K. Molecular effects of the cyanobacterial toxin cyanopeptolin (CP1020) occurring in algal blooms: Global transcriptome analysis in zebrafish embryos. Aquat. Toxicol. 2014, 149, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Gademann, K.; Portmann, C.; Blom, J.F.; Zeder, M.; Jüttner, F. Multiple toxin production in the cyanobacterium Microcystis: Isolation of the toxic protease inhibitor cyanopeptolin 1020. J. Nat. Prod. 2010, 73, 980–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okino, T.; Matsuda, H.; Murakami, M.; Yamaguchi, K. Microginin, an angiotensin-converting enzyme inhibitor from the blue-green alga Microcystis aeruginosa. Tetrahedron Lett. 1993, 34, 501–504. [Google Scholar] [CrossRef]
- Bagchi, S.N.; Das, P.K.; Ghosh, S.K. Screening and evaluation of protease inhibitory peptides in Microcystis spp.—Dominant water blooms. Microorg. Environ. Manag. 2012, 175–190. [Google Scholar]
- Namikoshi, M.; Rinehart, K.L. Bioactive compounds produced by cyanobacteria. J. Ind. Microbiol. Biotechnol. 1996, 17, 373–384. [Google Scholar] [CrossRef]
- Sano, T.; Usui, T.; Ueda, K.; Osada, H.; Kaya, K. Isolation of new protein phosphatase inhibitors from two cyanobacteria species, Planktothrix spp. J. Nat. Prod. 2001, 64, 1052–1055. [Google Scholar] [CrossRef] [PubMed]
- Sedmak, B.; Carmeli, S.; Eleršek, T. “Non-toxic” cyclic peptides induce lysis of cyanobacteria—An effective cell population density control mechanism in cyanobacterial blooms. Microb. Ecol. 2008, 56, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Yoshizawa, S.; Matsushima, R.; Watanabe, M.F.; Harada, K.; Ichihara, A.; Carmichael, W.W.; Fujiki, H. Inhibition of protein phosphatases by microcystins and nodularin associated with hepatotoxicity. J. Cancer Res. Clin. Oncol. 1990, 116, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Beattie, K.A.; Kaya, K.; Codd, G.A. The cyanobacterium Nodularia PCC 7804, of freshwater origin, produces [L-Har2]nodularin. Phytochemistry 2000, 54, 57–61. [Google Scholar] [CrossRef]
- Pearson, L.; Mihali, T.; Moffitt, M.; Kellmann, R.; Neilan, B. On the chemistry, toxicology and genetics of the cyanobacterial toxins, microcystin, nodularin, saxitoxin and cylindrospermopsin. Mar. Drugs 2010, 8, 1650. [Google Scholar] [CrossRef] [PubMed]
- Briand, J.-F.; Leboulanger, C.; Humbert, J.-F.; Bernard, C.; Dufour, P. Cylindrospermompsis raciborskii (cyanobacteria) invasion at mid-latitudes: Selection, wise physiological tolerance, or global warming? J. Phycol. 2004, 40, 231–238. [Google Scholar] [CrossRef]
- Runnegar, M.T.; Kong, S.M.; Zhong, Y.Z.; Ge, J.L.; Lu, S.C. The role of glutathione in the toxicity of a novel cyanobacterial alkaloid cylindrospermopsin in cultured rat hepatocytes. Biochem. Biophys. Res. Commun. 1994, 201, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Runnegar, M.T.; Xie, C.; Snider, B.B.; Wallace, G.A.; Weinreb, S.M.; Kuhlenkamp, J. In vitro hepatotoxicity of the cyanobacterial alkaloid cylindrospermopsin and related synthetic analogues. Toxicol. Sci. 2002, 67, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Fawell, J.K.; Mitchell, R.E.; Hill, R.E.; Everett, D.J. The toxicity of cyanobacterial toxins in the mouse: II anatoxin-a. Hum. Exp. Toxicol. 1999, 18, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Weirich, C.A.; Miller, T.R. Freshwater harmful algal blooms: Toxins and children’s health. Curr. Probl. Pediatr. Adolesc. Health Care 2014, 44, 2–24. [Google Scholar] [CrossRef] [PubMed]
- Fingerman, M.; Forester, R.H.; Stover, J.H., Jr. Action of shellfish poison on peripheral nerve and skeletal muscle. Proc. Soc. Exp. Biol. Med. 1953, 84, 643–646. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Sheets, M.; Ishida, H.; Li, F.; Barry, W.H. Saxitoxin blocks L-type ICa. J. Pharmacol. Exp. Ther. 2004, 308, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Blom, J.F.; Brütsch, T.; Barbaras, D.; Bethuel, Y.; Locher, H.H.; Hubschwerlen, C.; Gademann, K. Potent algicides based on the cyanobacterial alkaloid nostocarboline. Org. Lett. 2006, 8, 737–740. [Google Scholar] [CrossRef] [PubMed]
- Becher, P.G.; Baumann, H.I.; Gademann, K.; Jüttner, F. The cyanobacterial alkaloid nostocarboline: An inhibitor of acetylcholinesterase and trypsin. J. Appl. Phycol. 2008, 21, 103–110. [Google Scholar] [CrossRef]
- Dunn, O.J. On multiple tests and confidence intervals. Commun. Stat. Theory Methods 1974, 3, 101–103. [Google Scholar] [CrossRef]
- Heresztyn, T.; Nicholson, B.C. Nodularin concentrations in Lakes Alexandrina and Albert, South Australia, during a bloom of the cyanobacterium (blue-green alga) Nodularia spumigena and degradation of the toxin. Environ. Toxicol. Water Qual. 1997, 12, 273–282. [Google Scholar] [CrossRef]
- Wood, S.A.; Phillips, N.R.; de Winton, M.; Gibbs, M. Consumption of benthic cyanobacterial mats and nodularin-R accumulation in freshwater crayfish (Paranephrops planifrons) in Lake Tikitapu (Rotorua, New Zealand). Harmful Algae 2012, 20, 175–179. [Google Scholar] [CrossRef]
- Kemp, A.; John, J. Microcystins associated with Microcystis dominated blooms in the Southwest wetlands, Western Australia. Environ. Toxicol. 2006, 21, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Gehringer, M.M.; Adler, L.; Roberts, A.A.; Moffitt, M.C.; Mihali, T.K.; Mills, T.J.T.; Fieker, C.; Neilan, B.A. Nodularin, a cyanobacterial toxin, is synthesized in planta by symbiotic Nostoc sp. ISME J. 2012, 6, 1834–1847. [Google Scholar] [CrossRef] [PubMed]
- Wenta, R.; Sorsa, K. Road Salt Report; Dane County Public Health Department: Madison, WI, USA, 2011; pp. 1–26.
- Miller, T.R.; Beversdorf, L.; Chaston, S.D.; McMahon, K.D. Spatiotemporal molecular analysis of cyanobacteria blooms reveals Microcystis-Aphanizomenon interactions. PLoS ONE 2013, 8, e74933. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, R.P.; Madamwar, D.; Incharoensakdi, A. Bloom dynamics of cyanobacteria and their toxins: Environmental health impacts and mitigation strategies. Front. Microbiol. 2015, 6, 1254. [Google Scholar] [CrossRef] [PubMed]
- De Stasio, B.T. Harmful Algal Blooms in the Fox Valley: Recent Patterns of Cyanobacteria Abundance and Composition in the Fox-wolf Watershed and Green Bay; Wisconsin Harmful Algal Bloom Stakeholder Workshop; WI Sea Grant: Green Bay, WI, USA, 2007. [Google Scholar]
- Beversdorf, L.J.; Miller, T.R.; McMahon, K.D. Long-term monitoring reveals carbon-nitrogen metabolism key to microcystin production in eutrophic lakes. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, S.; Moore, R.E.; Niemczura, W.P.; Carmichael, W.W. Anatoxin-a(s), a potent anticholinesterase from Anabaena flos-aquae. J. Am. Chem. Soc. 1989, 111, 8021–8023. [Google Scholar] [CrossRef]
- Beversdorf, L.J.; Chaston, S.D.; Miller, T.R.; McMahon, K.D. Microcystin mcyA and mcyE gene abundances are not appropriate indicators of microcystin concentrations in lakes. PLoS ONE 2015, 10, e0125353. [Google Scholar] [CrossRef] [PubMed]
- Chorus, I. Toxic effects and substances in cyanobacteria other than microcystins, anatoxin-a and saxitoxins. In Cyanotoxins: Occurrence, Causes, Consequences; Chorus, I., Ed.; Springer: Berlin/Heidelberg, Germany, 2001; pp. 281–315. [Google Scholar]
- Fastner, J.; Erhard, M.; von Döhren, H. Determination of oligopeptide diversity within a natural population of Microcystis spp. (cyanobacteria) by typing single colonies by matrix-assisted laser desorption ionization–time of flight mass spectrometry. Appl. Environ. Microbiol. 2001, 67, 5069–5076. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.R.; McMahon, K.D. Genetic diversity of cyanobacteria in four eutrophic lakes. FEMS Microbiol. Ecol. 2011, 78, 336–348. [Google Scholar] [CrossRef] [PubMed]
- Furey, A.; Crowley, J.; Hamilton, B.; Lehane, M.; James, K.J. Strategies to avoid the mis-identification of anatoxin-a using mass spectrometry in the forensic investigation of acute neurotoxic poisoning. J. Chromatogr. A 2005, 1082, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, A.E.; Clesceri, L.S.; Eaton, A.D. Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 1992. [Google Scholar]
- Tett, P.; Kelly, M.G.; Hornberger, G.M. Method for spectrophotmetric measurement of chlorophyll-a and pheophytin-a in benthic microalgae. Limnol. Oceanogr. 1975, 20, 887–896. [Google Scholar] [CrossRef]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta Bioenerg. 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Matlab, 2012a; The MathWorks Inc.: Natick, MA, USA, 2012.
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lake | Trophic State * | Lake Type | Majority Land Use | Area (km2) | Max Depth (m) | Volume (106 m3) ** | Secchi (m) *** |
|---|---|---|---|---|---|---|---|
| Fish | Meso/eutrophic | Seepage | Ag | 0.87 | 18.9 | 8.2 | 2.8 |
| Wingra | Eutrophic | Drainage | Ur | 1.40 | 4.3 | 3.0 | 2.2 |
| Monona | Eutrophic | Drainage | Ur/Ag | 13.2 | 22.5 | 149 | 1.2 |
| Mendota | Eutrophic | Drainage | Ur/Ag | 39.4 | 25.3 | 498 | 1.7 |
| Koshkonong | Hyper/eutrophic | Drainage | Ag | 42.9 | 2.1 | 45.0 | 0.3 |
| Winnebago | Hyper/eutrophic | Drainage | Ur/Ag | 534 | 6.4 | 1709 | 0.6 |
| Lake | Function | MCLR | MCYR | MCLA | MCRR | Total MC | Apt B | AptF | Total Apt | NOD |
| Fish | Mean | 0.48 | BDL | 0.03 | BDL | 0.51 | 0.45 | 0.01 | 0.46 | BDL |
| Median | 0.44 | BDL | 0.03 | BDL | 0.47 | 0.28 | BDL | 0.28 | BDL | |
| Variance | 0.06 | 0.00 | 0.00 | 0.00 | 0.06 | 0.26 | 0.00 | 0.26 | 0.00 | |
| Max | 0.87 | BDL | 0.07 | BDL | 0.94 | 1.78 | 0.08 | 1.86 | BDL | |
| Koshkonong | Mean | 0.61 | 0.01 | BDL | 0.04 | 0.65 | 5.97 | 0.59 | 6.56 | 0.06 |
| Median | 0.26 | BDL | BDL | 0.02 | 0.28 | 4.13 | 0.15 | 4.28 | BDL | |
| Variance | 1.03 | 0.00 | 0.00 | 0.00 | 1.03 | 57.54 | 1.04 | 58.58 | 0.02 | |
| Max | 3.32 | 0.03 | 0.03 | 0.13 | 3.51 | 24.05 | 3.40 | 27.45 | 0.35 | |
| Monona | Mean | 4.83 | BDL | 0.35 | 0.01 | 5.20 | 0.73 | 0.07 | 0.80 | BDL |
| Median | 3.22 | BDL | 0.17 | 0.01 | 3.40 | 0.36 | 0.05 | 0.41 | BDL | |
| Variance | 21.35 | 0.00 | 0.18 | 0.00 | 21.54 | 1.05 | 0.01 | 1.06 | 0.00 | |
| Max | 12.40 | 0.03 | 1.11 | 0.04 | 13.58 | 3.41 | 0.24 | 3.65 | BDL | |
| Mendota | Mean | 0.38 | BDL | 0.19 | BDL | 0.58 | 0.21 | 0.01 | 0.22 | BDL |
| Median | 0.34 | BDL | 0.08 | BDL | 0.42 | 0.08 | BDL | 0.08 | BDL | |
| Variance | 0.18 | 0.00 | 0.05 | 0.00 | 0.23 | 0.07 | 0.00 | 0.07 | 0.00 | |
| Max | 1.11 | 0.03 | 0.60 | 0.02 | 1.76 | 0.63 | 0.03 | 0.66 | BDL | |
| Wingra | Mean | 0.10 | BDL | 0.01 | BDL | 0.11 | 0.09 | 0.02 | 0.11 | 0.06 |
| Median | BDL | BDL | BDL | BDL | 0.01 | 0.06 | 0.02 | 0.08 | BDL | |
| Variance | 0.02 | 0.00 | 0.00 | 0.00 | 0.02 | 0.01 | 0.00 | 0.01 | 0.03 | |
| Max | 0.33 | BDL | 0.02 | 0.02 | 0.37 | 0.25 | 0.06 | 0.31 | 0.54 | |
| Winnebago | Mean | 12.21 | 0.19 | 0.30 | 1.18 | 13.87 | 1.64 | 0.66 | 2.30 | BDL |
| Median | 6.37 | 0.07 | 0.07 | 0.56 | 7.07 | 0.97 | 0.52 | 1.49 | BDL | |
| Variance | 142.79 | 0.05 | 0.14 | 1.36 | 144.34 | 5.05 | 0.28 | 5.33 | 0.00 | |
| Max | 38.25 | 0.53 | 0.96 | 3.67 | 43.41 | 7.64 | 1.73 | 9.37 | BDL | |
| Lake | Function | Cpt1007 | Cpt1041 | Cpt1020 | Total Cpt | Mgn690 | ATX | hATX | STX | CYL |
| Fish | Mean | 0.04 | 0.06 | BDL | 0.11 | 0.01 | BDL | BDL | BDL | BDL |
| Median | 0.03 | 0.06 | BDL | 0.08 | BDL | BDL | BDL | BDL | BDL | |
| Variance | 0.00 | 0.00 | 0.00 | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Max | 0.14 | 0.14 | BDL | 0.28 | 0.04 | BDL | BDL | BDL | BDL | |
| Koshkonong | Mean | 0.19 | BDL | BDL | 0.19 | 0.23 | BDL | BDL | BDL | BDL |
| Median | BDL | BDL | BDL | BDL | BDL | BDL | BDL | BDL | BDL | |
| Variance | 0.37 | 0.00 | 0.00 | 0.37 | 0.48 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Max | 1.92 | BDL | BDL | 1.92 | 2.21 | BDL | BDL | BDL | BDL | |
| Monona | Mean | 0.11 | 0.55 | BDL | 0.67 | 0.05 | BDL | 0.51 | BDL | BDL |
| Median | 0.11 | 0.54 | BDL | 0.65 | 0.01 | BDL | BDL | BDL | BDL | |
| Variance | 0.01 | 0.22 | 0.00 | 0.23 | 0.01 | 0.00 | 2.60 | 0.00 | 0.00 | |
| Max | 0.28 | 1.17 | BDL | 1.45 | 0.35 | BDL | 5.10 | BDL | BDL | |
| Mendota | Mean | 0.04 | 0.26 | BDL | 0.30 | BDL | BDL | BDL | BDL | BDL |
| Median | BDL | 0.36 | BDL | 0.36 | BDL | BDL | BDL | BDL | BDL | |
| Variance | 0.00 | 0.06 | 0.00 | 0.07 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Max | 0.13 | 0.65 | BDL | 0.78 | 0.01 | BDL | BDL | BDL | BDL | |
| Wingra | Mean | BDL | BDL | BDL | BDL | 0.01 | BDL | BDL | BDL | BDL |
| Median | BDL | BDL | BDL | BDL | BDL | BDL | BDL | BDL | BDL | |
| Variance | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| Max | BDL | BDL | BDL | 0.01 | 0.02 | BDL | BDL | BDL | BDL | |
| Winnebago | Mean | 1.23 | 1.82 | BDL | 3.05 | 0.02 | 0.03 | BDL | BDL | BDL |
| Median | 0.75 | 1.32 | BDL | 2.07 | 0.02 | BDL | BDL | BDL | BDL | |
| Variance | 3.35 | 4.47 | 0.00 | 7.83 | 0.00 | 0.01 | 0.00 | 0.00 | 0.00 | |
| Max | 6.38 | 7.54 | BDL | 13.92 | 0.07 | 0.30 | BDL | BDL | BDL |
| Lake | Function | Chl-a | TP | DRP | TN | N + N | NH4+ | TN:TP | DIN:DRP | pH |
|---|---|---|---|---|---|---|---|---|---|---|
| Fish | Mean | 14.7 | 14.6 | 3.60 | 798 | 16.7 | 85.7 | 9900 | 970 | 8.5 |
| (n = 13) | Median | 13.8 | 15.0 | 1.00 | 716 | 5.00 | 12.0 | 57.3 | 174 | 8.7 |
| Max | 28.2 | 45.0 | 14.5 | 1170 | 62.0 | 504.0 | 72000 | 6900 | 9.1 | |
| Koshkonong | Mean | 27.2 | 501 | 61.4 | ND | ND | ND | ND | ND | ND |
| (n = 10 *) | Median | 17.8 | 36.0 | 9.00 | ND | ND | ND | ND | ND | ND |
| Max | 72.1 | 507 | 233 | ND | ND | ND | ND | ND | ND | |
| Monona | Mean | 10.8 | 62.4 | 17.0 | 970 | 54.6 | 89.8 | 17.9 | 820 | 8.6 |
| (n = 14) | Median | 9.85 | 45.5 | 2.50 | 911 | 0.01 | 25.5 | 17.6 | 9.00 | 8.8 |
| Max | 37.7 | 230 | 78.9 | 2937 | 229 | 314 | 32.1 | 630 | 8.9 | |
| Mendota | Mean | 9.49 | 44.9 | 16.2 | 750 | 267 | 126 | 20.6 | 1195 | 8.5 |
| (n = 17) | Median | 7.75 | 46.5 | 12.0 | 786 | 135 | 77.0 | 17.5 | 16.7 | 8.6 |
| Max | 26.7 | 112 | 49.0 | 1210 | 627 | 436 | 40.0 | 18800 | 8.9 | |
| Wingra | Mean | 8.15 | 28.9 | 0.53 | 728 | 27.9 | 17.9 | 26.4 | 3800 | 8.6 |
| (n = 15) | Median | 5.85 | 29.0 | 0.00 | 720 | 0.00 | 0.00 | 24.0 | 1600 | 8.4 |
| Max | 18.9 | 44.0 | 3.00 | 923 | 211 | 88.0 | 38.7 | 21000 | 9.2 | |
| Winnebago | Mean | 11.0 | 149 | 30.4 | ND | ND | ND | ND | ND | ND |
| (n = 10 *) | Median | 9.80 | 139 | 27.5 | ND | ND | ND | ND | ND | ND |
| Max | 16.9 | 195 | 78.0 | ND | ND | ND | ND | ND | ND |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beversdorf, L.J.; Weirich, C.A.; Bartlett, S.L.; Miller, T.R. Variable Cyanobacterial Toxin and Metabolite Profiles across Six Eutrophic Lakes of Differing Physiochemical Characteristics. Toxins 2017, 9, 62. https://doi.org/10.3390/toxins9020062
Beversdorf LJ, Weirich CA, Bartlett SL, Miller TR. Variable Cyanobacterial Toxin and Metabolite Profiles across Six Eutrophic Lakes of Differing Physiochemical Characteristics. Toxins. 2017; 9(2):62. https://doi.org/10.3390/toxins9020062
Chicago/Turabian StyleBeversdorf, Lucas J., Chelsea A. Weirich, Sarah L. Bartlett, and Todd. R. Miller. 2017. "Variable Cyanobacterial Toxin and Metabolite Profiles across Six Eutrophic Lakes of Differing Physiochemical Characteristics" Toxins 9, no. 2: 62. https://doi.org/10.3390/toxins9020062
APA StyleBeversdorf, L. J., Weirich, C. A., Bartlett, S. L., & Miller, T. R. (2017). Variable Cyanobacterial Toxin and Metabolite Profiles across Six Eutrophic Lakes of Differing Physiochemical Characteristics. Toxins, 9(2), 62. https://doi.org/10.3390/toxins9020062