
Nitrate Increased Cucumber Tolerance to Fusarium Wilt by Regulating Fungal Toxin Production and Distribution

Abstract
:1. Introduction
2. Results
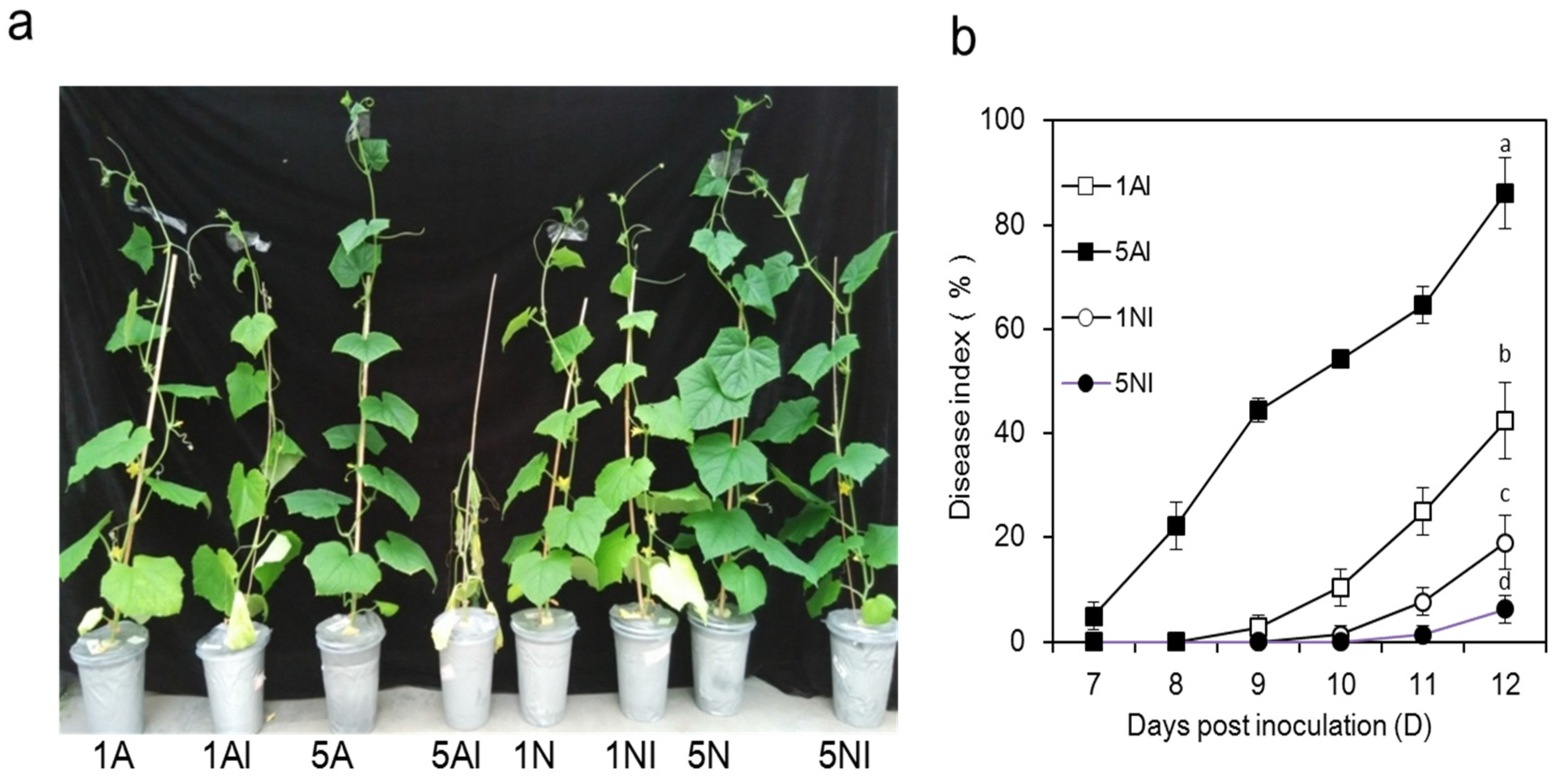
2.1. Effects of Different N Sources on the Growth and Disease Index of Cucumber Plants
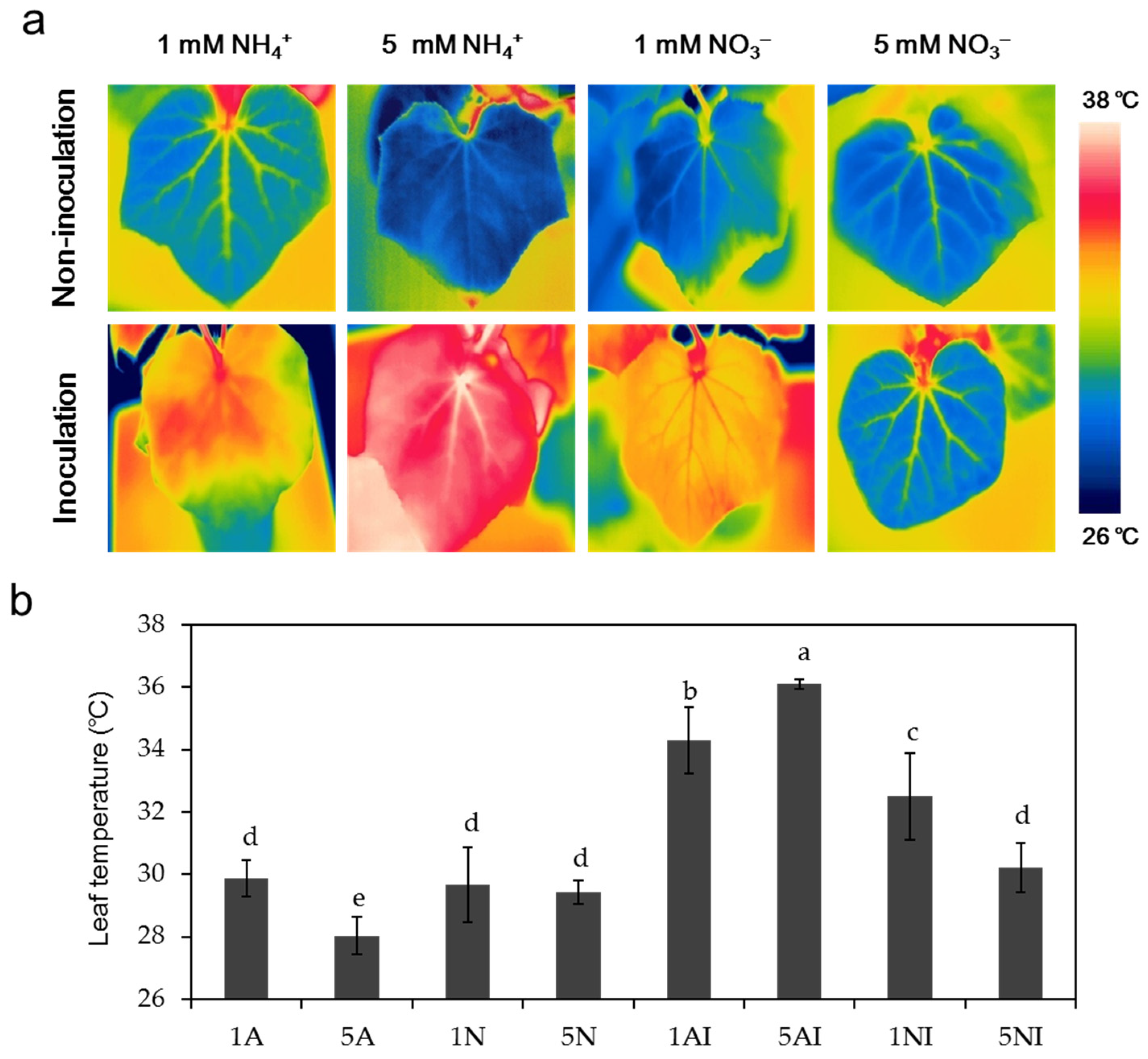
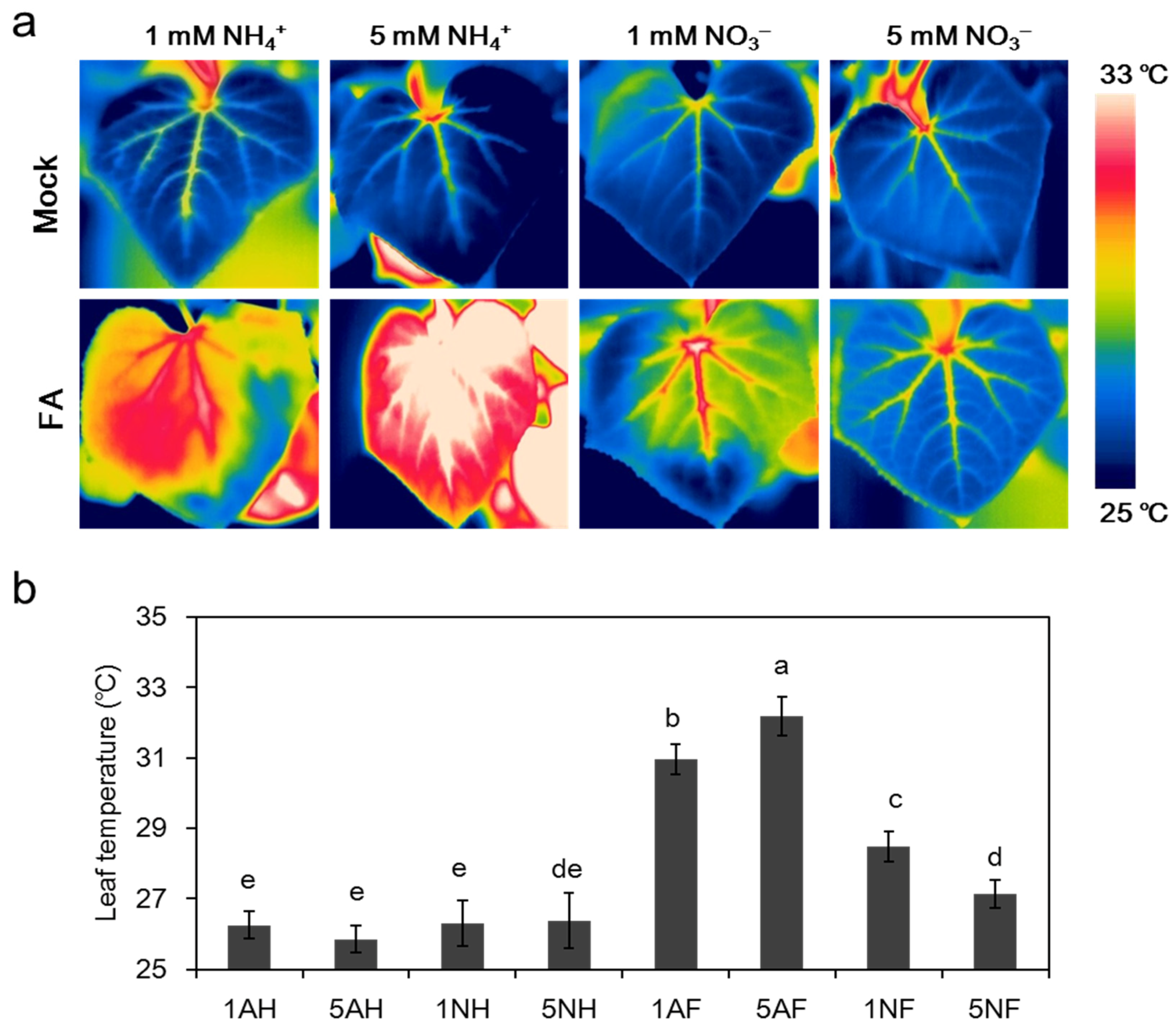
2.2. Effects of Different N Sources on the Leaf Temperature of Cucumber Plants
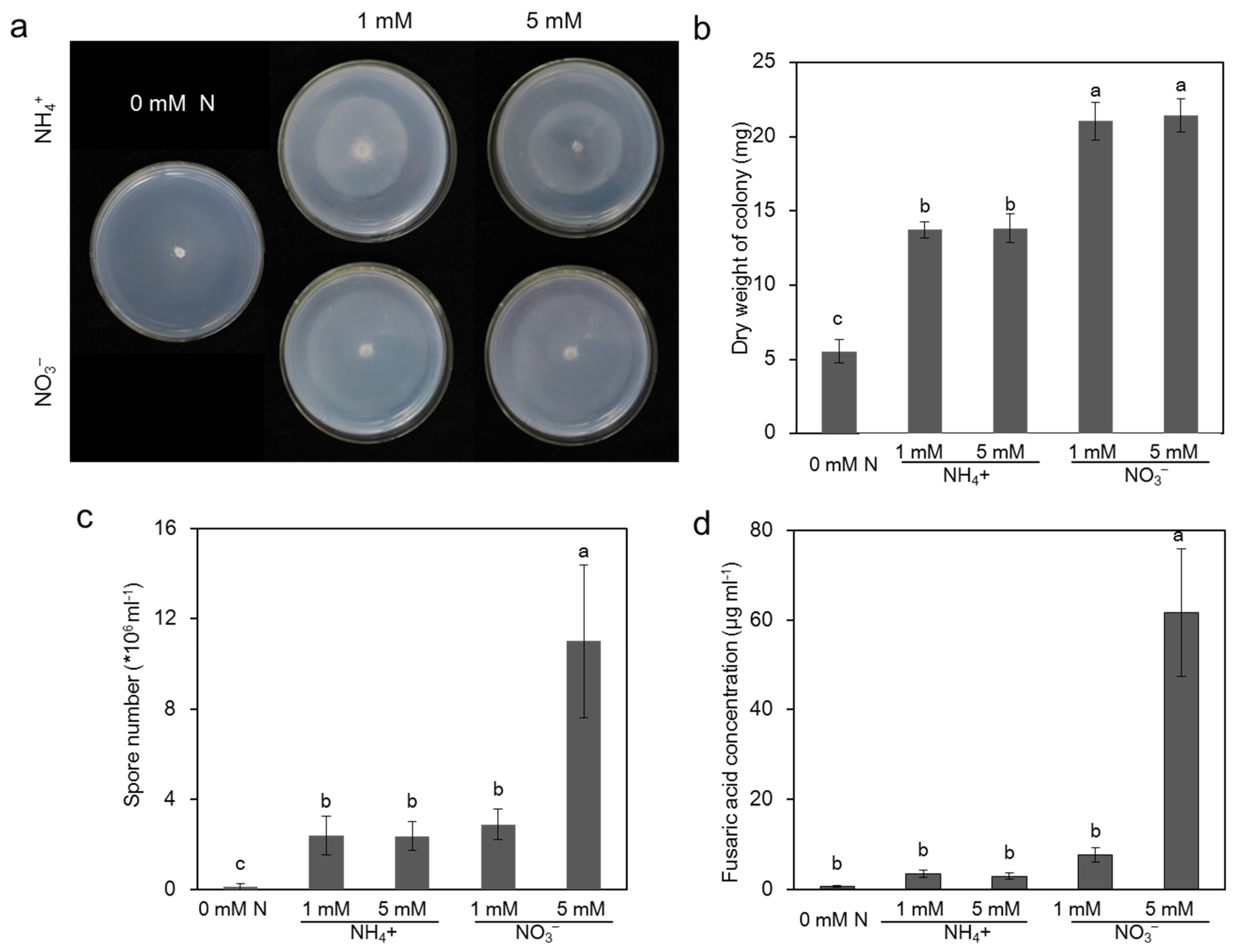
2.3. Effect of Different N Sources on the Pathogen Growth In Vitro Culture
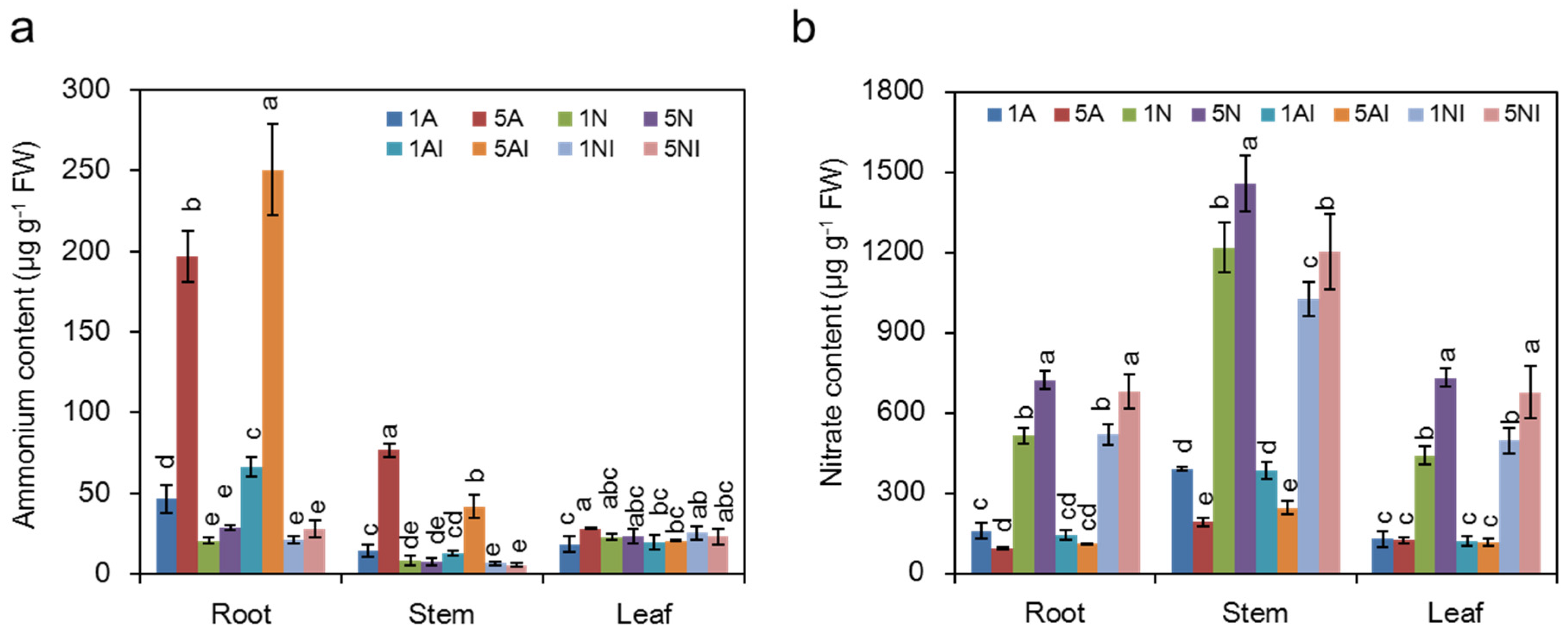
2.4. Effects of Different N Sources on the NH4+ and NO3− Content of Cucumber Plants
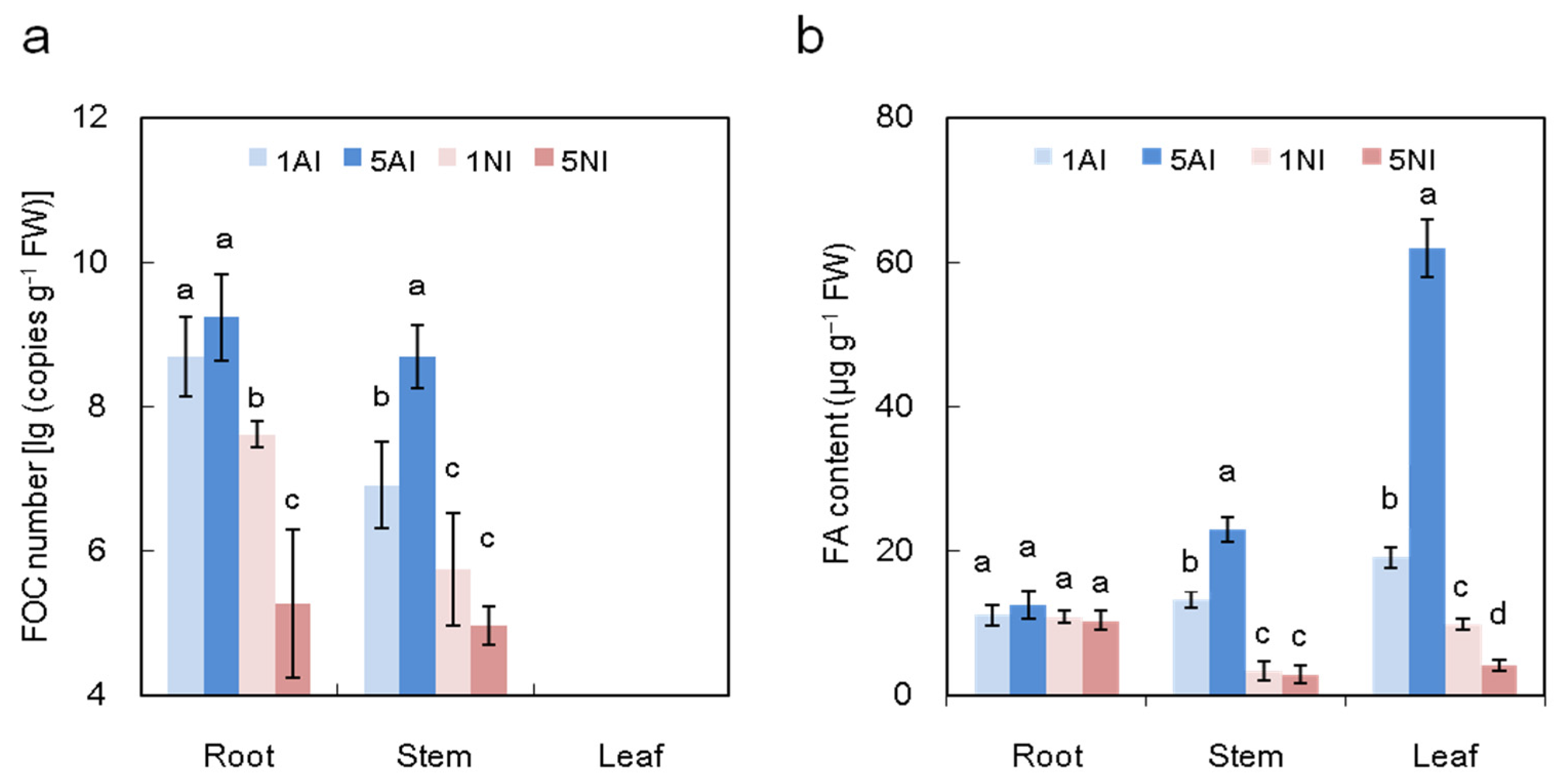
2.5. Effect of Different N Sources on the Pathogen Distributions and FA Content of Cucumber Plants
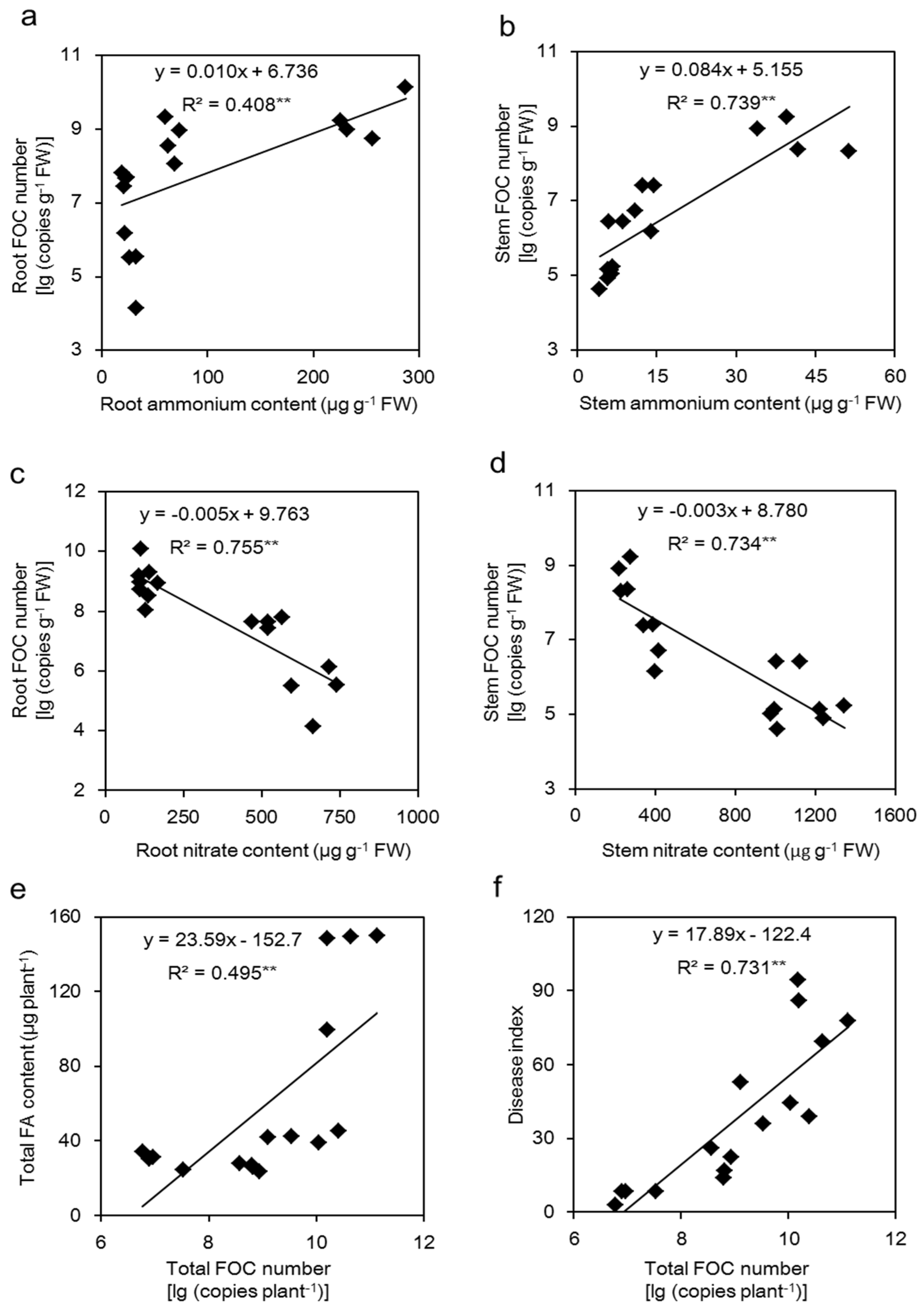
2.6. Correlations between the NH4+ Content or NO3− Content and the FOC Number and between the Total FOC Number and the Total FA Content or Disease Index
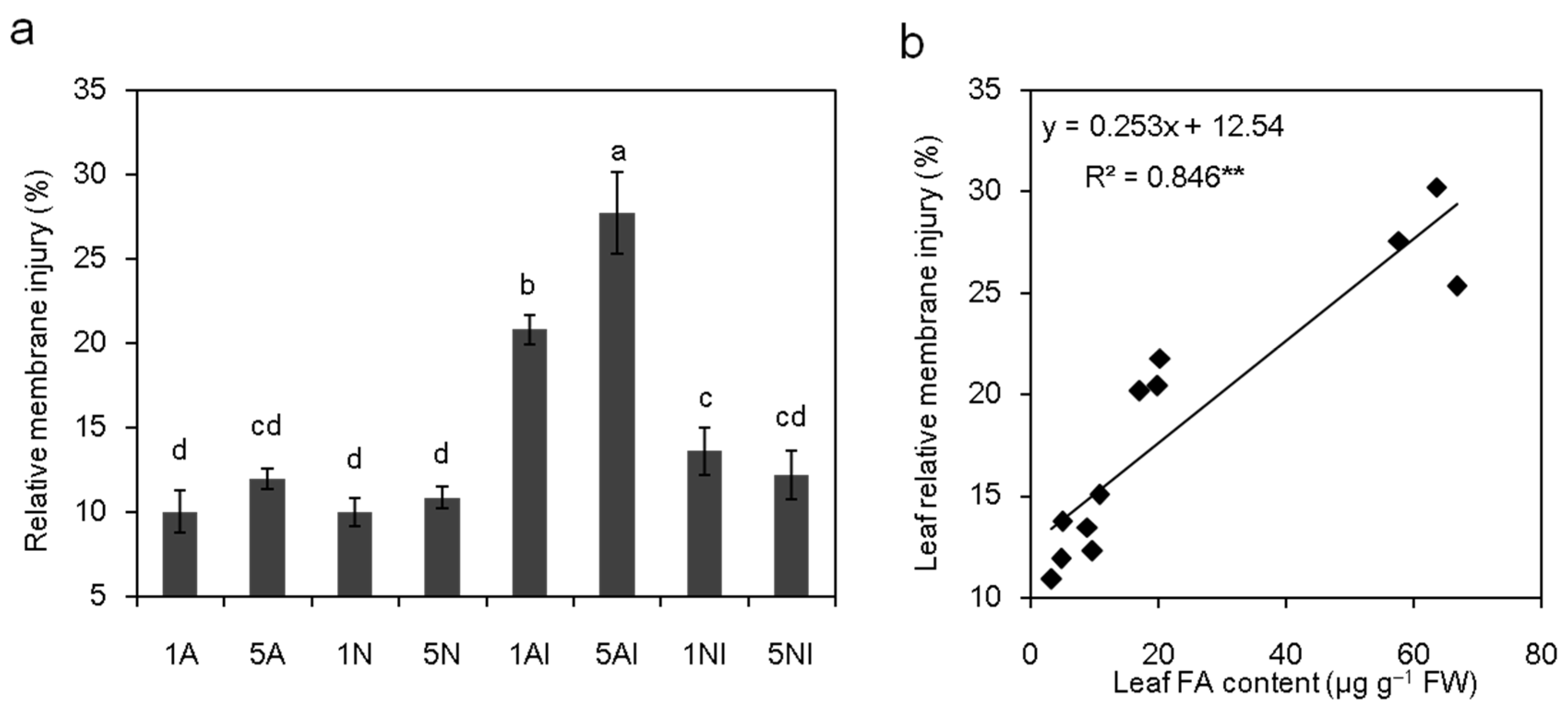
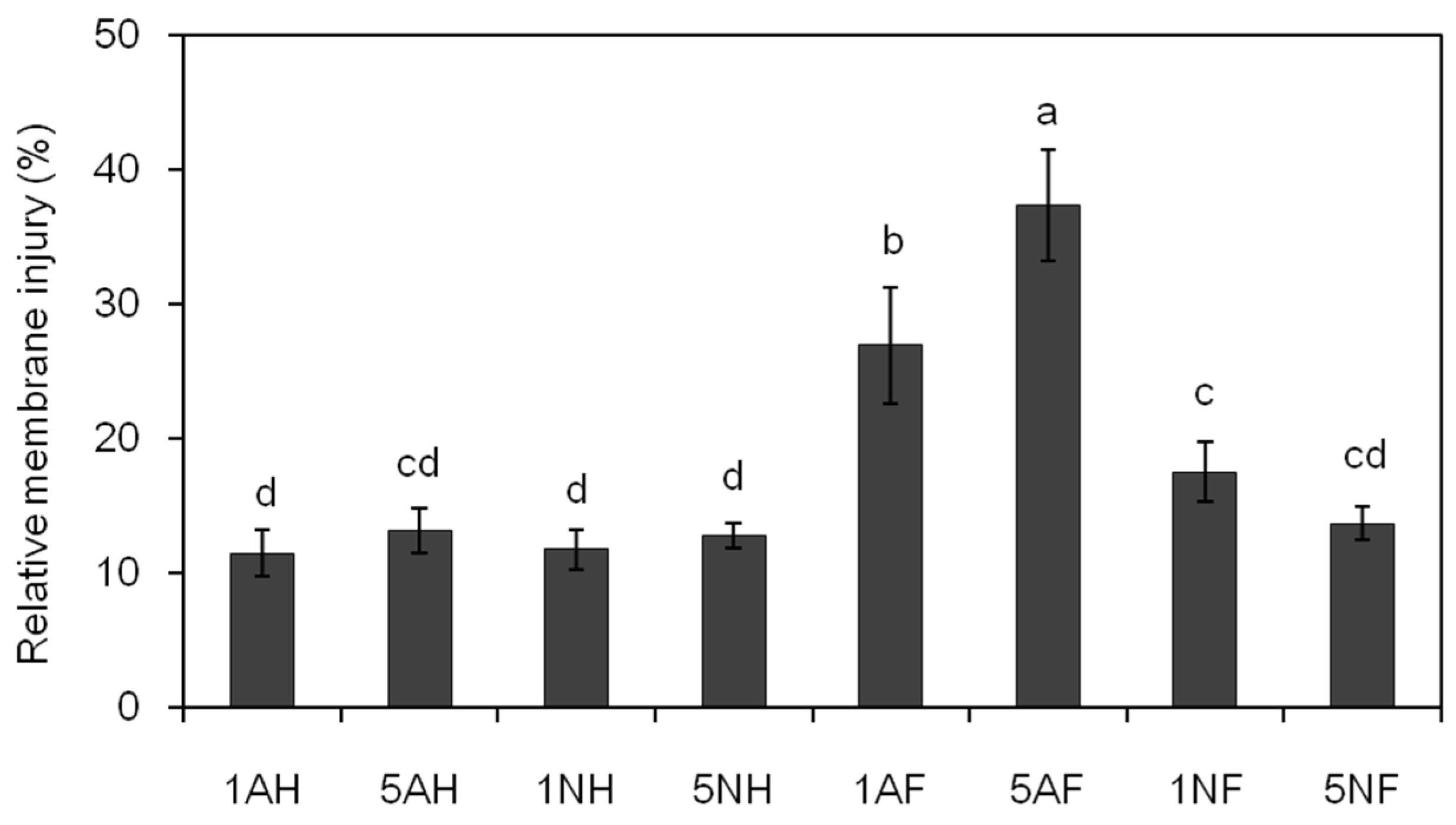
2.7. Effect of Different N Sources on the Leaf Relative Membrane Injury of Cucumber Plants after FOC Infection
2.8. Effect of FA on the Cucumber Plants Treated with Different N Sources
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Thermal Imaging
4.3. Plant Biomass Measurements
4.4. Fungal Colony Dry Weight, Conidial Number and Pathogen Crude Toxin Levels Grown under Different N Sources
4.5. Pathogen Incubation and Infection
4.6. Determination of the Disease Index
4.7. FA Extraction and Analysis
4.8. Specific Detection of FOC by Real-Time PCR
4.9. FA Response Experiment
4.10. Relative Leaf Membrane Injury
4.11. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ahn, P.; Chung, H.S.; Lee, Y.H. Vegetative compatibility groups and pathogenicity among isolates of Fusarium oxysporum f. sp. cucumerinum. Plant Dis. 1998, 82, 244–246. [Google Scholar] [CrossRef]
- Owen, J.H. Fusarium wilt of cucumber. Phytopathology 1955, 45, 435–439. [Google Scholar]
- Pietro, A.D.; Madrid, M.P.; Caracuel, Z.; Delgado-Jarana, J.; Roncero, M.I.G. Fusarium oxysporum: Exploring the molecular arsenal of a vascular wilt fungus. Mol. Plant Pathol. 2003, 4, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Knogge, W. Fungal infection of plants. Plant Cell 1996, 8, 1711–1772. [Google Scholar] [CrossRef] [PubMed]
- Bacon, C.; Porter, J.; Norred, W.; Leslie, J. Production of fusaric acid by Fusarium species. Appl. Environ. Microbiol. 1996, 62, 4039–4043. [Google Scholar] [PubMed]
- Dong, X.; Ling, N.; Wang, M.; Shen, Q.R.; Guo, S.W. Fusaric acid is a crucial factor in the disturbance of leaf water imbalance in Fusarium-infected banana plants. Plant Physiol. Biochem. 2012, 60, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Ling, N.; Dong, X.; Liu, X.K.; Shen, Q.R.; Guo, S.W. Effect of fusaric acid on the leaf physiology of cucumber seedlings. Eur. J. Plant Pathol. 2014, 138, 103–112. [Google Scholar] [CrossRef]
- Bouizgarne, B.; El-Maarouf-Bouteau, H.; Frankart, C.; Reboutier, D.; Madiona, K.; Pennarun, A.; Monestiez, M.; Trouverie, J.; Amiar, Z.; Briand, J. Early physiological responses of Arabidopsis thaliana cells to fusaric acid: Toxic and signalling effects. New Phytol. 2006, 169, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Pavlovkin, J.; Mistrik, I.; Prokop, M. Some aspects of the phytotoxic action of fusaric acid on primary Ricinus roots. Plant Soil Environ. 2004, 50, 397–401. [Google Scholar]
- Marrè, M.; Vergani, P.; Albergoni, F. Relationship between fusaric acid uptake and its binding to cell structures by leaves of Egeria densa and its toxic effects on membrane permeability and respiration. Physiol. Mol. Plant Pathol. 1993, 42, 141–157. [Google Scholar] [CrossRef]
- Gapillout, I.; Milat, M.L.; Blein, J.P. Effects of fusaric acid on cells from tomato cultivars resistant or susceptible to Fusarium oxysporum f. sp. lycopersici. Eur. J. Plant Pathol. 1996, 102, 127–132. [Google Scholar] [CrossRef]
- Jiao, J.; Sun, L.; Zhou, B.; Gao, Z.; Hao, Y.; Zhu, X.; Liang, Y. Hydrogen peroxide production and mitochondrial dysfunction contribute to the fusaric acid-induced programmed cell death in tobacco cells. J. Plant Physiol. 2014, 171, 1197–1203. [Google Scholar] [CrossRef] [PubMed]
- Curir, P.; Guglieri, L.; Dolci, M.; Capponi, A.; Aurino, G. Fusaric acid production by Fusarium oxysporum f. sp. lilii and its role in the lily basal rot disease. Eur. J. Plant Pathol. 2000, 106, 849–856. [Google Scholar] [CrossRef]
- Wu, H.S.; Bao, W.; Liu, D.Y.; Ling, N.; Ying, R.R.; Raza, W.; Shen, Q.R. Effect of fusaric acid on biomass and photosynthesis of watermelon seedlings leaves. Caryologia 2008, 61, 258–268. [Google Scholar] [CrossRef]
- D’Alton, A.; Etherton, B. Effects of fusaric acid on tomato root hair membrane potentials and ATP levels. Plant Physiol. 1984, 74, 39–42. [Google Scholar] [CrossRef] [PubMed]
- Kuźniak, E. Effects of fusaric acid on reactive oxygen species and antioxidants in tomato cell cultures. J. Phytopathol. 2001, 149, 575–582. [Google Scholar] [CrossRef]
- Samadi, L.; Behboodi, B.S. Fusaric acid induces apoptosis in saffron root-tip cells: Roles of caspase-like activity, cytochrome c, and H2O2. Planta 2006, 225, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Harborne, J.B. Plant toxins and their effects on animals. In Introduction to Ecological Biochemistry; Academic Press: London, UK, 1993; pp. 71–103. [Google Scholar]
- Fagard, M.; Launay, A.; Clément, G.; Courtial, J.; Dellagi, A.; Farjad, M.; Krapp, A.; Soulié, M.C.; Masclaux-Daubresse, C. Nitrogen metabolism meets phytopathology. J. Exp. Bot. 2014, 65, 5643–5656. [Google Scholar] [CrossRef] [PubMed]
- Huber, D.; Watson, R. Nitrogen form and plant disease. Annu. Rev. Phytopathol. 1974, 12, 139–165. [Google Scholar] [CrossRef] [PubMed]
- Huber, D.; Wilhelm, N. The role of manganese in resistance to plant diseases. In Manganese in Soils and Plants; Springer: Dordrecht, The Netherlands, 1988; pp. 155–173. [Google Scholar]
- Walters, D.; Bingham, I. Influence of nutrition on disease development caused by fungal pathogens: implications for plant disease control. Ann. Appl. Biol. 2007, 151, 307–324. [Google Scholar] [CrossRef]
- Engelsberger, W.R.; Schulze, W.X. Nitrate and ammonium lead to distinct global dynamic phosphorylation patterns when resupplied to nitrogen-starved Arabidopsis seedlings. Plant J. 2012, 69, 978–995. [Google Scholar] [CrossRef] [PubMed]
- Horchani, F.; Hajri, R.; Aschi-Smiti, S. Effect of ammonium or nitrate nutrition on photosynthesis, growth, and nitrogen assimilation in tomato plants. J. Plant Nutr. Soil Sci. 2010, 173, 610–617. [Google Scholar] [CrossRef]
- Guo, S.W.; Chen, G.; Zhou, Y.; Shen, Q.R. Ammonium nutrition increases photosynthesis rate under water stress at early development stage of rice (Oryza sativa L.). Plant Soil 2007, 296, 115–124. [Google Scholar] [CrossRef]
- Yang, X.X.; Li, Y.; Ren, B.B.; Ding, L.; Gao, C.M.; Shen, Q.R.; Guo, S.W. Drought-induced root aerenchyma formation restrains water uptake in nitrate-supplied rice seedlings. Plant Cell Environ. 2012, 53, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Patterson, K.; Cakmak, T.; Cooper, A.; Lager, I.; Rasmusson, A.G.; Escobar, M.A. Distinct signalling pathways and transcriptome response signatures differentiate ammonium-and nitrate-supplied plants. Plant Cell Environ. 2010, 33, 1486–1501. [Google Scholar] [CrossRef] [PubMed]
- Solomon, P.S.; Tan, K.C.; Oliver, R.P. The nutrient supply of pathogenic fungi; a fertile field for study. Mol. Plant Pathol. 2003, 4, 203–210. [Google Scholar] [CrossRef] [PubMed]
- López-Berges, M.S.; Rispail, N.; Prados-Rosales, R.C.; Di Pietro, A. A nitrogen response pathway regulates virulence functions in Fusarium oxysporum via the protein kinase TOR and the bZIP protein MeaB. Plant Cell 2010, 22, 2459–2475. [Google Scholar] [CrossRef] [PubMed]
- Stout, M.J.; Brovont, R.A.; Duffey, S.S. Effect of nitrogen availability on expression of constitutive and inducible chemical defenses in tomato, Lycopersicon esculentum. J. Chem. Ecol. 1998, 24, 945–963. [Google Scholar] [CrossRef]
- Neumann, S.; Paveley, N.; Beed, F.; Sylvester-Bradley, R. Nitrogen per unit leaf area affects the upper asymptote of Puccinia striiformis f. sp. tritici epidemics in winter wheat. Plant Pathol. 2004, 53, 725–732. [Google Scholar] [CrossRef]
- Jensen, B.; Munk, L. Nitrogen-induced changes in colony density and spore production of Erysiphe graminis f. sp. hordei on seedlings of six spring barley cultivars. Plant Pathol. 1997, 46, 191–202. [Google Scholar] [CrossRef]
- Hoffland, E.; van Beusichem, M.L.; Jeger, M.J. Nitrogen availability and susceptibility of tomato leaves to Botrytis cinerea. Plant Soil 1999, 210, 263–272. [Google Scholar] [CrossRef]
- Dietrich, R.; Ploss, K.; Heil, M. Constitutive and induced resistance to pathogens in Arabidopsis thaliana depends on nitrogen supply. Plan Cell Environ. 2004, 27, 896–906. [Google Scholar] [CrossRef]
- Snoeijers, S.S.; Pérez-García, A.; Joosten, M.H.; De Wit, P.J. The effect of nitrogen on disease development and gene expression in bacterial and fungal plant pathogens. Eur. J. Plant Pathol. 2000, 106, 493–506. [Google Scholar] [CrossRef]
- Hoffland, E.; Jeger, M.J.; van Beusichem, M.L. Effect of nitrogen supply rate on disease resistance in tomato depends on the pathogen. Plant Soil 2000, 218, 239–247. [Google Scholar] [CrossRef]
- Lecompte, F.; Abro, M.A.; Nicot, P. Contrasted responses of Botrytis cinerea isolates developing on tomato plants grown under different nitrogen nutrition regimes. Plant Pathol. 2010, 59, 891–899. [Google Scholar] [CrossRef]
- Bolton, M.D.; Thomma, B.P. The complexity of nitrogen metabolism and nitrogen-regulated gene expression in plant pathogenic fungi. Physiol. Mol. Plant Pathol. 2008, 72, 104–110. [Google Scholar] [CrossRef]
- Ben-Yephet, Y.; Reuven, M.; Zviebil, A.; Szmulewich, Y.; Lavkovits, I.; Markovits, T.; Soriano, S.; Bar-Yosef, B. Effect of nutrition on deformation disease in Gypsophila paniculata mother plants. Phytopathology 2006, 96, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Crespo, E.; Scalschi, L.; Llorens, E.; García-Agustín, P.; Camañes, G. NH4+ protects tomato plants against Pseudomonas syringae by activation of systemic acquired acclimation. J. Exp. Bot. 2015, 66, 6777–6790. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.J.; Brotman, Y.; Segu, S.; Zeier, T.; Zeier, J.; Persijn, S.T.; Cristescu, S.M.; Harren, F.J.; Bauwe, H.; Fernie, A.R. The form of nitrogen nutrition affects resistance against Pseudomonas syringae pv. phaseolicola in tobacco. J. Exp. Bot. 2013, 64, 553–568. [Google Scholar] [CrossRef] [PubMed]
- Lemmens, M.; Buerstmayr, H.; Krska, R.; Schuhmacher, R.; Grausgruber, H.; Ruckenbauer, P. The effect of inoculation treatment and long-term application of moisture on Fusarium head blight symptoms and deoxynivalenol contamination in wheat grains. Eur. J. Plant Pathol. 2004, 110, 299–308. [Google Scholar] [CrossRef]
- Oerke, E.; Steiner, U.; Dehne, H.; Lindenthal, M. Thermal imaging of cucumber leaves affected by downy mildew and environmental conditions. J. Exp. Bot. 2006, 57, 2121–2132. [Google Scholar] [CrossRef] [PubMed]
- Lindenthal, M.; Steiner, U.; Dehne, H.W.; Oerke, E.C. Effect of downy mildew development on transpiration of cucumber leaves visualized by digital infrared thermography. Phytopathology 2005, 95, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.W.; Brück, H.; Sattelmacher, B. Effects of supplied nitrogen form on growth and water uptake of french bean (phaseolus vulgaris L.) plants. Plant Soil 2002, 239, 267–275. [Google Scholar] [CrossRef]
- Iii, F.S.C.; Walter, C.H.S.; Clarkson, D.T. Growth response of barley and tomato to nitrogen stress and its control by abscisic acid, water relations and photosynthesis. Planta 1988, 173, 352–366. [Google Scholar]
- Chaerle, L.; Van, C.W.; Messens, E.; Lambers, H.; Van, M.M.; Van, D.S.D. Presymptomatic visualization of plant-virus interactions by thermography. Nat. Biotechnol. 1999, 17, 813–816. [Google Scholar] [PubMed]
- Mcdonald, K.L.; Cahill, D.M. Evidence for a transmissible factor that causes rapid stomatal closure in soybean at sites adjacent to and remote from hypersensitive cell death induced by phytophthora sojae. Physiol. Mol. Plant Pathol. 1999, 55, 197–203. [Google Scholar] [CrossRef]
- Wang, M.; Ling, N.; Dong, X.; Zhu, Y.Y.; Shen, Q.R.; Guo, S.W. Thermographic visualization of leaf response in cucumber plants infected with the soil-borne pathogen Fusarium oxysporum f. sp. cucumerinum. Plant Physiol. Biochem. 2012, 61, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Chaerle, L.; De Boever, F.; Montagu, M.V.; Straeten, D. Thermographic visualization of cell death in tobacco and Arabidopsis. Plant Cell Environ. 2001, 24, 15–25. [Google Scholar] [CrossRef]
- Lorenzini, G.; Guidi, L.; Nali, C.; Ciompi, S.; Soldatini, G.F. Photosynthetic response of tomato plants to vascular wilt diseases. Plant Sci. 1997, 124, 143–152. [Google Scholar] [CrossRef]
- Mur, L.A.; Simpson, C.; Kumari, A.; Gupta, A.K.; Gupta, K.J. Moving nitrogen to the centre of plant defence against pathogens. Ann. Bot. 2016, mcw 179. [Google Scholar] [CrossRef]
- Solomon, P.S.; Oliver, R.P. Evidence that gamma-aminobutyric acid is major nitrogen source during Cladosporium fulvum infection of tomato. Planta 2002, 214, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.W.; Zhou, Y.; Shen, Q.R.; Zhang, F.S. Effect of ammonium and nitrate nutrition on some physiological processes in higher plants-growth, photosynthesis, photorespiration, and water relations. Plant Biol. 2007, 9, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Aked, J.; Hall, J. Effect of powdery mildew infection on concentrations of apoplastic sugars in pea leaves. New Phytol. 1993, 123, 283–288. [Google Scholar] [CrossRef]
- Tetlow, I.; Farrar, J. Apoplastic sugar concentration and pH in barley leaves infected with brown rust. J. Exp. Bot. 1993, 44, 929–936. [Google Scholar] [CrossRef]
- Barna, B.; Sarhan, A.R.T.; Kiraly, Z. The influence of nitrogen nutrition on the sensitivity of tomato plants to culture filtrates of Fusarium and to fusaric acid. Physiol. Plant Pathol. 1983, 23, 257–263. [Google Scholar] [CrossRef]
- Fakhouri, W.; Walker, F.; Armbruster, W.; Buchenauer, H. Detoxification of fusaric acid by a nonpathogenic Colletotrichum sp. Physiol. Mol. Plant Pathol. 2003, 63, 263–269. [Google Scholar] [CrossRef]
- Delledonne, M.; Zeier, J.; Marocco, A.; Lamb, C. Signal interactions between nitric oxide and reactive oxygen intermediates in the plant hypersensitive disease resistance response. Proc. Natl. Acad. Sci. USA 2001, 98, 13454–13459. [Google Scholar] [CrossRef] [PubMed]
- Mur, L.A.J.; Prats, E.; Pierre, S.; Hall, M.A.; Hebelstrup, K.H. Integrating nitric oxide into salicylic acid and jasmonic acid/ethylene plant defense pathways. Front. Plant Sci. 2013, 4, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Xiong, Y.F.; Ling, N.; Feng, X.M.; Zhong, Z.T.; Shen, Q.R.; Guo, S.W. Detection of the dynamic response of cucumber leaves to fusaric acid using thermal imaging. Plant Physiol. Biochem. 2013, 66, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.C.; Flematti, G.R.; Ghisalberti, E.L.; Sivasithamparam, K.; Chakraborty, S.; Obanor, F.; Barbetti, M.J. Mycotoxins produced by Fusarium species associated with annual legume pastures and “sheep feed refusal disorders” in western Australia. Mycotoxin Res. 2011, 27, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Zhang, Z.H.; Ling, N.; Yuan, Y.J.; Zheng, X.Y.; Shen, B.; Shen, Q.R. Bacillus subtilis SQR 9 can control Fusarium wilt in cucumber by colonizing plant roots. Biol. Fertil. Soils 2011, 47, 495–506. [Google Scholar] [CrossRef]
- Wang, M.; Sun, Y.M.; Gu, Z.C.; Wang, R.R.; Sun, G.M.; Zhu, C.; Guo, S.W.; Shen, Q.R. Nitrate protects cucumber plants against Fusarium oxysporum by regulating citrate exudation. Plant Cell Physiol. 2016, 57, 2001–2012. [Google Scholar] [CrossRef] [PubMed]
- Tjamos, S.E.; Flemetakis, E.; Paplomatas, E.J.; Katinakis, P. Induction of resistance to Verticillium dahliae in Arabidopsis thaliana by the biocontrol agent k-165 and pathogenesis-related proteins gene expression. Mol. Plant Micro. Interact. 2005, 18, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Lievens, B.; Claes, L.; Vakalounakis, D.J.; Vanachter, A.C.; Thomma, B.P. A robust identification and detection assay to discriminate the cucumber pathogens Fusarium oxysporum f. sp. cucumerinum and f. sp. radicis-cucumerinum. Environ. Microbiol. 2007, 9, 2145–2161. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Root (g) | Stem (g) | Leaf (g) |
|---|---|---|---|
| 1A | 0.54 ± 0.03 d | 1.21 ± 0.03 cd | 1.68 ± 0.08 c |
| 5A | 0.73 ± 0.02 b | 1.96 ± 0.20 b | 2.58 ± 0.10 b |
| 1N | 0.61 ± 0.01 c | 1.40 ± 0.14 c | 1.97 ± 0.21 c |
| 5N | 0.85 ± 0.03 a | 2.66 ± 0.33 a | 3.81 ± 0.58 a |
| 1AI | 0.48 ± 0.02 e | 1.07 ± 0.05 d | 1.18 ± 0.11 d |
| 5AI | 0.36 ± 0.07 f | 1.38 ± 0.27 c | 1.62 ± 0.28 c |
| 1NI | 0.57 ± 0.04 cd | 1.18 ± 0.11 cd | 1.63 ± 0.09 c |
| 5NI | 0.80 ± 0.05 a | 2.39 ± 0.14 a | 3.64 ± 0.21 a |
| Treatments | Root (μg·g−1 FW) | Stem (μg·g−1 FW) | Leaf (μg·g−1 FW) |
|---|---|---|---|
| 1AF | 195 ± 9 c | 273 ± 12 a | 142 ± 6 b |
| 5AF | 206 ± 21 c | 238 ± 14 b | 174 ± 14 a |
| 1NF | 270 ± 13 b | 205 ± 7 b | 120 ± 10 c |
| 5NF | 306 ± 17 a | 168 ± 12 bc | 100 ±5 d |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, J.; Wang, M.; Sun, Y.; Gu, Z.; Wang, R.; Saydin, A.; Shen, Q.; Guo, S. Nitrate Increased Cucumber Tolerance to Fusarium Wilt by Regulating Fungal Toxin Production and Distribution. Toxins 2017, 9, 100. https://doi.org/10.3390/toxins9030100
Zhou J, Wang M, Sun Y, Gu Z, Wang R, Saydin A, Shen Q, Guo S. Nitrate Increased Cucumber Tolerance to Fusarium Wilt by Regulating Fungal Toxin Production and Distribution. Toxins. 2017; 9(3):100. https://doi.org/10.3390/toxins9030100
Chicago/Turabian StyleZhou, Jinyan, Min Wang, Yuming Sun, Zechen Gu, Ruirui Wang, Asanjan Saydin, Qirong Shen, and Shiwei Guo. 2017. "Nitrate Increased Cucumber Tolerance to Fusarium Wilt by Regulating Fungal Toxin Production and Distribution" Toxins 9, no. 3: 100. https://doi.org/10.3390/toxins9030100
APA StyleZhou, J., Wang, M., Sun, Y., Gu, Z., Wang, R., Saydin, A., Shen, Q., & Guo, S. (2017). Nitrate Increased Cucumber Tolerance to Fusarium Wilt by Regulating Fungal Toxin Production and Distribution. Toxins, 9(3), 100. https://doi.org/10.3390/toxins9030100