
Cardiac Cell Patterning on Customized Microelectrode Arrays for Electrophysiological Recordings



Abstract
:1. Introduction
2. Materials and Methods
2.1. MEA Device Fabrication
2.2. Surface Topographic Feature and Blocker Fabrication
2.3. Cell Patterning and Culture
2.4. Immunofluorescence Staining
2.5. Ca2+ Indicator Loading and Video Analysis
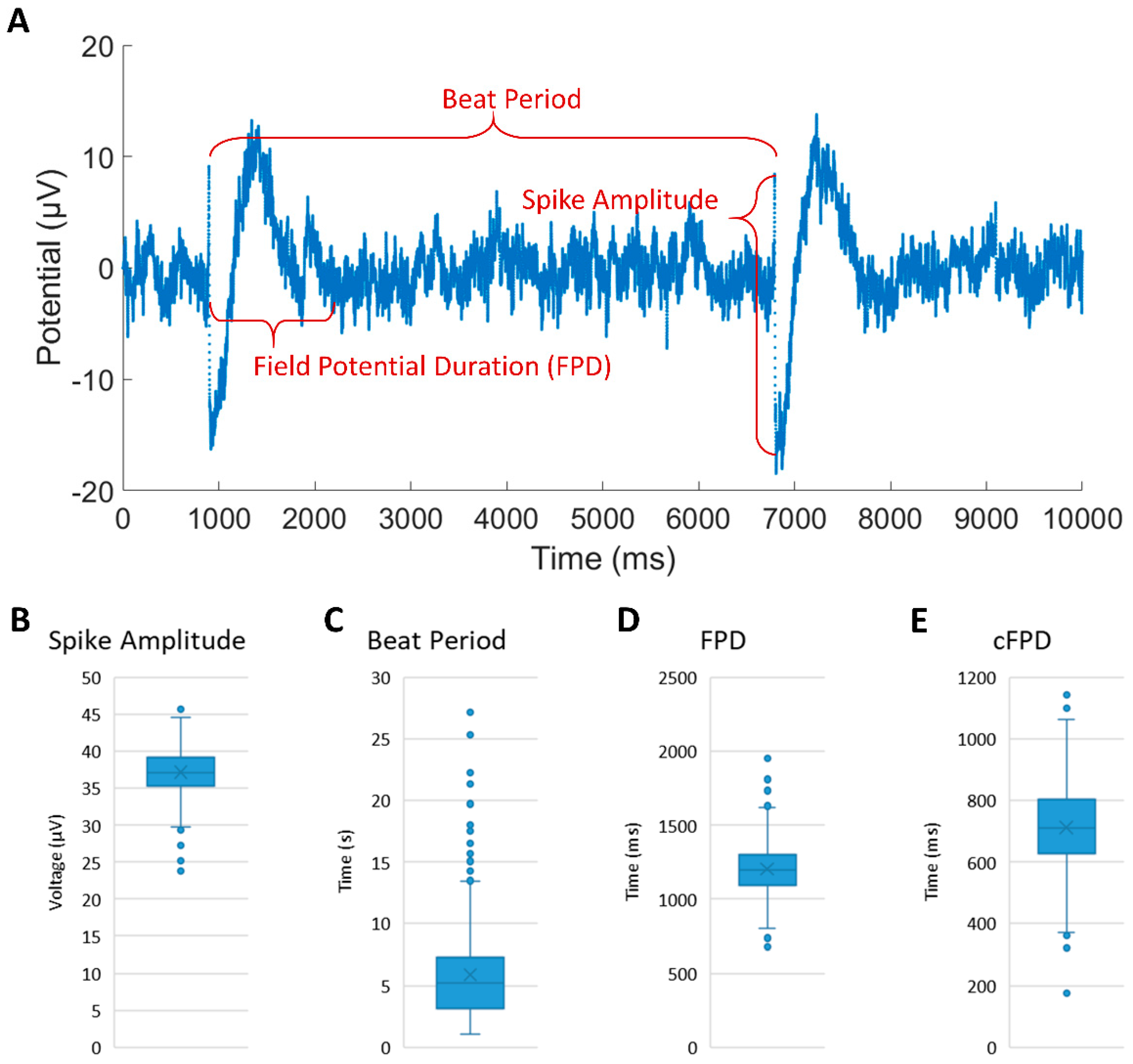
2.6. Electrical Data Analysis
3. Results and Discussion
3.1. MEA Sensor Device and Topographic Feature Fabrication
3.2. Cell Patterning with Surface Topographic Features
3.3. Extracellular Recording of the Synchronized rCM-Fibroblast Network
3.4. Extracellular Recording of iCMs
4. Summary and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Spach, M.S.; Boineau, J.P. Microfibrosis produces electrical load variations due to loss of side-to-side cell connections: A major mechanism of structural heart disease arrhythmias. Pacing Clin. Electrophysiol. 1997, 20, 397–413. [Google Scholar] [CrossRef] [PubMed]
- Vasquez, C.; Morley, G.E. The origin and arrhythmogenic potential of fibroblasts in cardiac disease. J. Cardiovasc. Trans. Res. 2012, 5, 760–767. [Google Scholar] [CrossRef] [Green Version]
- Camelliti, P.; Green, C.R.; Kohl, P. Structural and functional coupling of cardiac myocytes and fibroblasts. Adv. Cardiol. 2006, 42, 132–149. [Google Scholar] [CrossRef]
- Casini, S.; Verkerk, A.O.; Remme, C.A. Human iPSC-Derived Cardiomyocytes for Investigation of Disease Mechanisms and Therapeutic Strategies in Inherited Arrhythmia Syndromes: Strengths and Limitations. Cardiovasc. Drugs Ther. 2017, 31, 325–344. [Google Scholar] [CrossRef] [Green Version]
- Kussauer, S.; David, R.; Lemcke, H. hiPSCs Derived Cardiac Cells for Drug and Toxicity Screening and Disease Modeling: What micro-electrode-array analyses can tell us. Cells 2019, 8, 1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, V.; Chong, N.; Ellis, B.; Ren, X.; Senapati, S.; Chang, H.-C.; Zorlutuna, P. Constant-potential environment for activating and synchronizing cardiomyocyte colonies with on-chip ion-depleting perm-selective membranes. Lab Chip 2020, 20, 4273–4284. [Google Scholar] [CrossRef]
- Connolly, P.; Clark, P.; Curtis, A.; Dow, J.; Wilkinson, C. An Extracellular microelectrode Array for monitoring electrogenic cells in culture. Biosens. Bioelectron. 1990, 5, 223–234. [Google Scholar] [CrossRef]
- Thomas, C.A.; Springer, P.A.; Loeb, G.E.; Berwald-Netter, Y.; Okun, L.M. A miniature microelectrode array to monitor the bioelectric activity of cultured cells. Exp. Cell Res. 1972, 74, 61–66. [Google Scholar] [CrossRef]
- Caluori, G.; Pribyl, J.; Pesl, M.; Jelinkova, S.; Rotrekl, V.; Skladal, P.; Raiteri, R. Non-invasive electromechanical cell-based biosensors for improved investigation of 3D cardiac models. Biosens. Bioelectron. 2019, 124–125, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Simeonov, S.; Schäffer, T.E. Ultrafast Imaging of Cardiomyocyte Contractions by Combining Scanning Ion Conductance Microscopy with a Microelectrode Array. Anal. Chem. 2019, 91, 9648–9655. [Google Scholar] [CrossRef]
- Agladze, N.N.; Halaidych, O.V.; Tsvelaya, V.A.; Bruegmann, T.; Kilgus, C.; Sasse, P.; Agladze, K.I. Synchronization of excitable cardiac cultures of different origin. Biomater. Sci. 2017, 5, 1777–1785. [Google Scholar] [CrossRef]
- Kaneko, T.; Toriumi, H.; Shimada, J.; Nomura, F. Extracellular field potential recording of single cardiomyocytes in agarose microchambers using microelectrode array. Jpn. J. Appl. Phys. 2018, 57, 03EB03. [Google Scholar] [CrossRef]
- Alassaf, A.; Tansik, G.; Mayo, V.; Wubker, L.; Carbonero, D.; Agarwal, A. Engineering anisotropic cardiac monolayers on microelectrode arrays for non-invasive analyses of electrophysiological properties. Analyst 2019, 145, 139–149. [Google Scholar] [CrossRef]
- Tijore, A.; Irvine, S.A.; Sarig, U.; Mhaisalkar, P.; Baisane, V.; Venkatraman, S. Contact guidance for cardiac tissue engineering using 3D bioprinted gelatin patterned hydrogel. Biofabrication 2018, 10, 25003. [Google Scholar] [CrossRef]
- Kim, M.-C.; Neal, D.M.; Kamm, R.D.; Asada, H.H. Dynamic modeling of cell migration and spreading behaviors on fibronectin coated planar substrates and micropatterned geometries. PLoS Comput. Biol. 2013, 9, e1002926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneko, T.; Nomura, F.; Yasuda, K. On-chip constructive cell-network study (I): Contribution of cardiac fibroblasts to cardiomyocyte beating synchronization and community effect. J. Nanobiotechnol. 2011, 9, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gherghiceanu, M.; Barad, L.; Novak, A.; Reiter, I.; Itskovitz-Eldor, J.; Binah, O.; Popescu, L.M. Cardiomyocytes derived from human embryonic and induced pluripotent stem cells: Comparative ultrastructure. J. Cell. Mol. Med. 2011, 15, 2539–2551. [Google Scholar] [CrossRef] [Green Version]
- Davis, R.P.; Casini, S.; van den Berg, C.W.; Hoekstra, M.; Remme, C.A.; Dambrot, C.; Salvatori, D.; Oostwaard, D.W.; Wilde, A.A.M.; Bezzina, C.R.; et al. Cardiomyocytes derived from pluripotent stem cells recapitulate electrophysiological characteristics of an overlap syndrome of cardiac sodium channel disease. Circulation 2012, 125, 3079–3091. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Gao, J.; Pi, R.; Liu, P. An Optimized Protocol for Culture of Cardiomyocyte from Neonatal Rat. Cytotechnology 2005, 49, 109–116. [Google Scholar] [CrossRef] [Green Version]
- Ellis, B.W.; Acun, A.; Can, U.I.; Zorlutuna, P. Human iPSC-derived myocardium-on-chip with capillary-like flow for personalized medicine. Biomicrofluidics 2017, 11, 24105. [Google Scholar] [CrossRef] [Green Version]
- Yeung, C.-K.; Sommerhage, F.; Wrobel, G.; Law, J.K.-Y.; Offenhäusser, A.; Rudd, J.A.; Ingebrandt, S.; Chan, M. To establish a pharmacological experimental platform for the study of cardiac hypoxia using the microelectrode array. J. Pharmacol. Toxicol. Methods 2009, 59, 146–152. [Google Scholar] [CrossRef]
- Ren, X.; Gomez, J.; Bashar, M.K.; Ji, J.; Can, U.I.; Chang, H.-C.; Shukla, N.; Datta, S.; Zorlutuna, P. Cardiac Muscle Cell-Based Coupled Oscillator Network for Collective Computing. Adv. Intell. Syst. 2021, 2000253. [Google Scholar] [CrossRef]
- Yamamoto, W.; Asakura, K.; Ando, H.; Taniguchi, T.; Ojima, A.; Uda, T.; Osada, T.; Hayashi, S.; Kasai, C.; Miyamoto, N.; et al. Electrophysiological Characteristics of Human iPSC-Derived Cardiomyocytes for the Assessment of Drug-Induced Proarrhythmic Potential. PLoS ONE 2016, 11, e0167348. [Google Scholar] [CrossRef] [Green Version]
- Zwartsen, A.; de Korte, T.; Nacken, P.; de Lange, D.W.; Westerink, R.H.S.; Hondebrink, L. Cardiotoxicity screening of illicit drugs and new psychoactive substances (NPS) in human iPSC-derived cardiomyocytes using microelectrode array (MEA) recordings. J. Mol. Cell. Cardiol. 2019, 136, 102–112. [Google Scholar] [CrossRef]
- Natarajan, A.; Stancescu, M.; Dhir, V.; Armstrong, C.; Sommerhage, F.; Hickman, J.J.; Molnar, P. Patterned cardiomyocytes on microelectrode arrays as a functional, high information content drug screening platform. Biomaterials 2011, 32, 4267–4274. [Google Scholar] [CrossRef] [Green Version]
- Werdich, A.A.; Lima, E.A.; Ivanov, B.; Ges, I.; Anderson, M.E.; Wikswo, J.P.; Baudenbacher, F.J. A microfluidic device to confine a single cardiac myocyte in a sub-nanoliter volume on planar microelectrodes for extracellular potential recordings. Lab Chip 2004, 4, 357–362. [Google Scholar] [CrossRef]
- Klauke, N.; Smith, G.L.; Cooper, J. Stimulation of Single Isolated Adult Ventricular Myocytes within a Low Volume Using a Planar Microelectrode Array. Biophys. J. 2003, 85, 1766–1774. [Google Scholar] [CrossRef] [Green Version]
- Joo, S.; Lim, J.; Nam, Y. Design and Fabrication of Miniaturized Neuronal Circuits on Microelectrode Arrays Using Agarose Hydrogel Micro-molding Technique. BioChip J. 2018, 12, 193–201. [Google Scholar] [CrossRef]
- Smith, A.S.T.; Choi, E.; Gray, K.; Macadangdang, J.; Ahn, E.H.; Clark, E.C.; Laflamme, M.A.; Wu, J.C.; Murry, C.E.; Tung, L.; et al. NanoMEA: A Tool for High-Throughput, Electrophysiological Phenotyping of Patterned Excitable Cells. Nano Lett. 2020, 20, 1561–1570. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Minami, I.; Yu, L.; Tsuji, K.; Nakajima, M.; Qiao, J.; Suzuki, M.; Shimono, K.; Nakatsuji, N.; Kotera, H.; et al. Extracellular Recordings of Patterned Human Pluripotent Stem Cell-Derived Cardiomyocytes on Aligned Fibers. Stem Cells Int. 2016, 2016, 2634013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moutaux, E.; Charlot, B.; Genoux, A.; Saudou, F.; Cazorla, M. An integrated microfluidic/microelectrode array for the study of activity-dependent intracellular dynamics in neuronal networks. Lab Chip 2018, 18, 3425–3435. [Google Scholar] [CrossRef] [PubMed]
- Gladkov, A.; Pigareva, Y.; Kutyina, D.; Kolpakov, V.; Bukatin, A.; Mukhina, I.; Kazantsev, V.; Pimashkin, A. Design of Cultured Neuron Networks in vitro with Predefined Connectivity Using Asymmetric Microfluidic Channels. Sci. Rep. 2017, 7, 15625. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Cell Source | Patterns Description | MEA Platform and Electrode Materials | Electrode Arrangement | Ref. |
|---|---|---|---|---|---|
| Study the activity-dependent intracellular dynamics in reconstituted neuron networks | Neuron | Single directed axon isolated by microfluidic device | Pt electrode on custom MEA | In the target chambers | Moutaux et al. [31] |
| Propose a design of organized neuron network with directed connectivity | Neuron | Unidirectional axon growth in microchambers with various shapes | Ti electrode on commercial MEA | In segments of microchambers | Gladkov et al. [32] |
| Design neuronal circuits with various topologies | Neuron | Neuron bodies in microwell and neurites in microgroove | TiN electrode on commercial MEA | In microwell | Joo et al. [28] |
| Describe a microfluidic device to measure FP from a single CM | CM | Single CM trapped in microfluidic device | Pt electrode on custom MEA | In microchambers | Werdich et al. [26] |
| Describe a microchamber to isolate single CM with electrical stimulation in small volumes | CM | Single isolated CM cell in each microchannel | Au electrode on custom MEA | Stimulation and reference electrodes in each microchamber | Klauke et al. [27] |
| Enable measurement of extracellular FP of a single CM | CM | Single CM individually seeded in hollow microchambers | ITO electrode on custom MEA | In each microchamber | Kaneko et al. [12] |
| Enable measurement of conductivity, viscosity and refractory period of CMs | CM | A monolayer CM with long path on the surface modified by PEG and fibronectin | Ti electrode on commercial MEA | In the start and end positions of path | Natarajan et al. [25] |
| Develop multi-well MEA plates with nanotopographic patterns | CM /Neuron | Aligned cells along the topographic nanogrooves | Custom MEA | Electrode matrix in each well | Smith et al. [29] |
| Study the electrical activity of iCMs on aligned fibers | CM | Anisotropic iCMs on the surface modified by electrospun aligned fiber | Ti electrode on commercial MEA | / | Li et al. [30] |
| Investigate the physical and protein cues to engineer aligned iCMs | CM | Anisotropic monolayer of iCMs along the direction of printed proteins and microgrooves | Ti electrode on commercial MEA | / | Alassaf et al. [13] |
| Study the coupling dynamics of a controlled network of rCM and fibroblast connections | CM /Fibroblast | Three individual rCM clusters and connected fibroblast bridges patterned by topographic features and mobile blockers | Au electrode on custom MEA | In CM clusters and fibroblast connection pathway | This paper |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, J.; Ren, X.; Zorlutuna, P. Cardiac Cell Patterning on Customized Microelectrode Arrays for Electrophysiological Recordings. Micromachines 2021, 12, 1351. https://doi.org/10.3390/mi12111351
Ji J, Ren X, Zorlutuna P. Cardiac Cell Patterning on Customized Microelectrode Arrays for Electrophysiological Recordings. Micromachines. 2021; 12(11):1351. https://doi.org/10.3390/mi12111351
Chicago/Turabian StyleJi, Jiaying, Xiang Ren, and Pinar Zorlutuna. 2021. "Cardiac Cell Patterning on Customized Microelectrode Arrays for Electrophysiological Recordings" Micromachines 12, no. 11: 1351. https://doi.org/10.3390/mi12111351
APA StyleJi, J., Ren, X., & Zorlutuna, P. (2021). Cardiac Cell Patterning on Customized Microelectrode Arrays for Electrophysiological Recordings. Micromachines, 12(11), 1351. https://doi.org/10.3390/mi12111351