
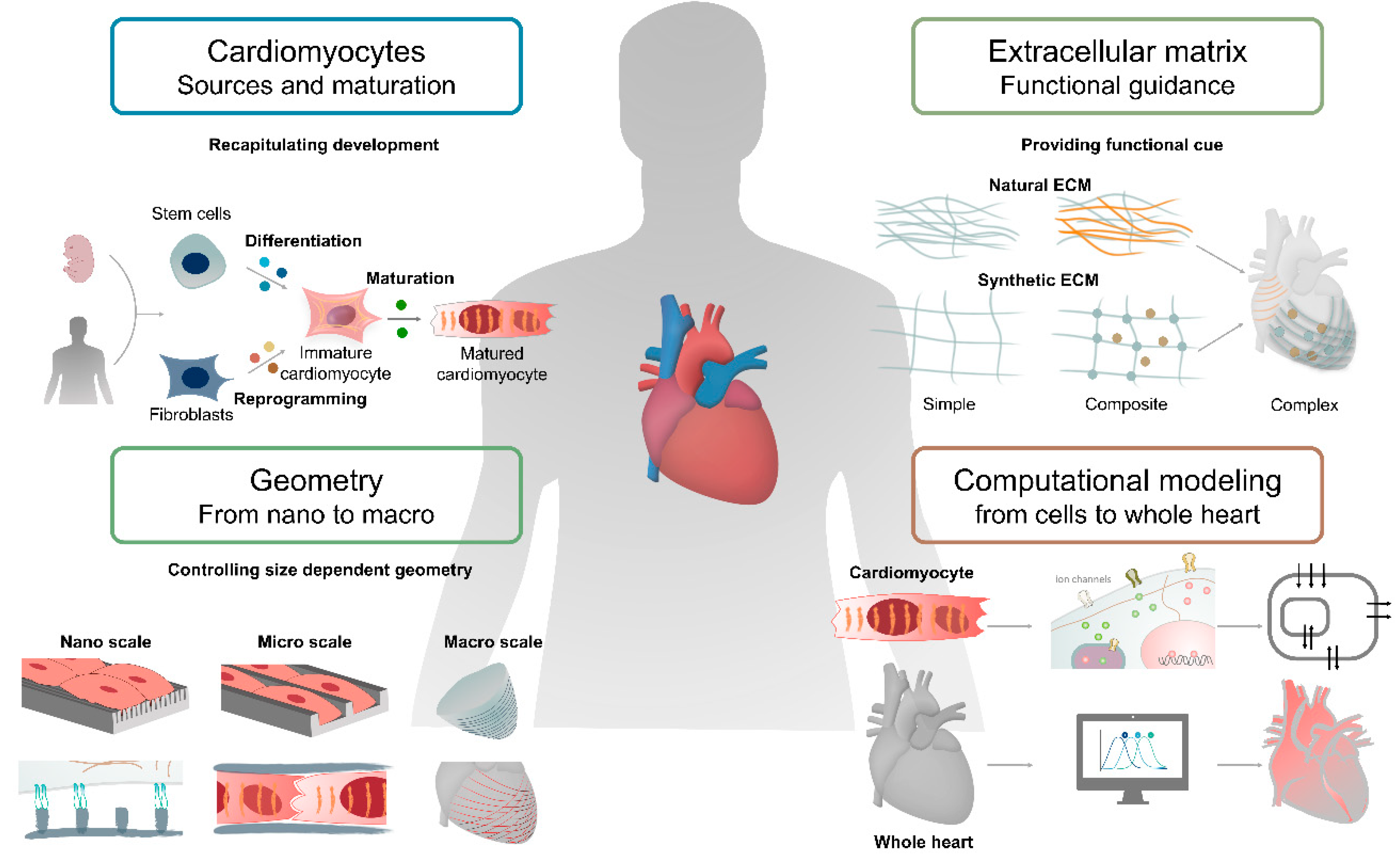
Recapitulating Cardiac Structure and Function In Vitro from Simple to Complex Engineering





Abstract
:
1. Introduction
2. Cardiomyocytes
2.1. Establishment of Cardiomyocyte Differentiation Using Rodent Cells
2.1.1. Primary Rodent Cardiomyocytes
2.1.2. Mouse Embryonic Stem Cell-Derived Cardiomyocytes
2.2. Generation of Cardiomyocytes from Human PSCs
2.2.1. Establishment of Human PSC-Derived Cardiomyocyte
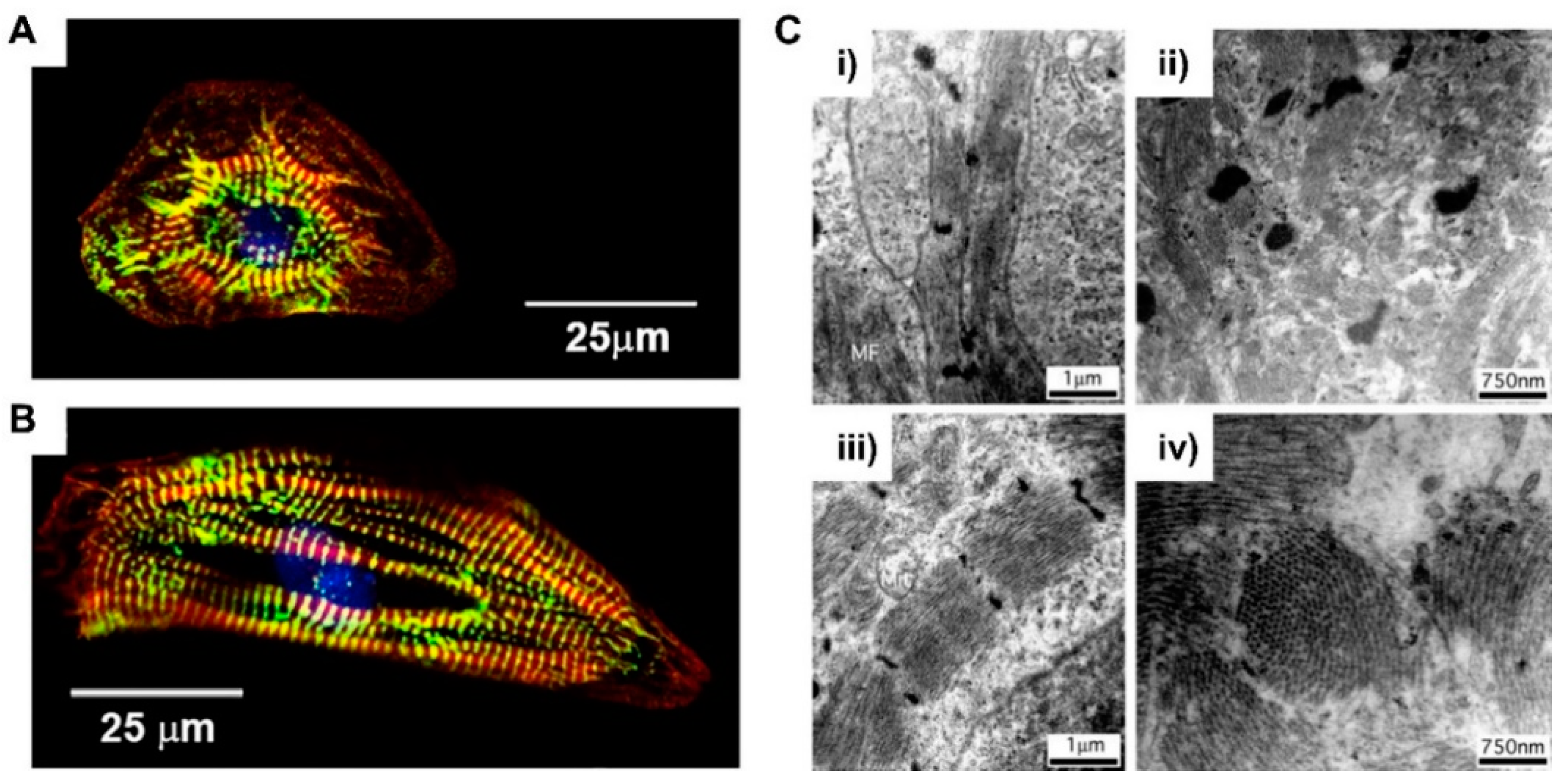
2.2.2. The Major Properties of Matured Cardiomyocyte
2.3. Cell-Based Application for Cardiac Tissue Engineering
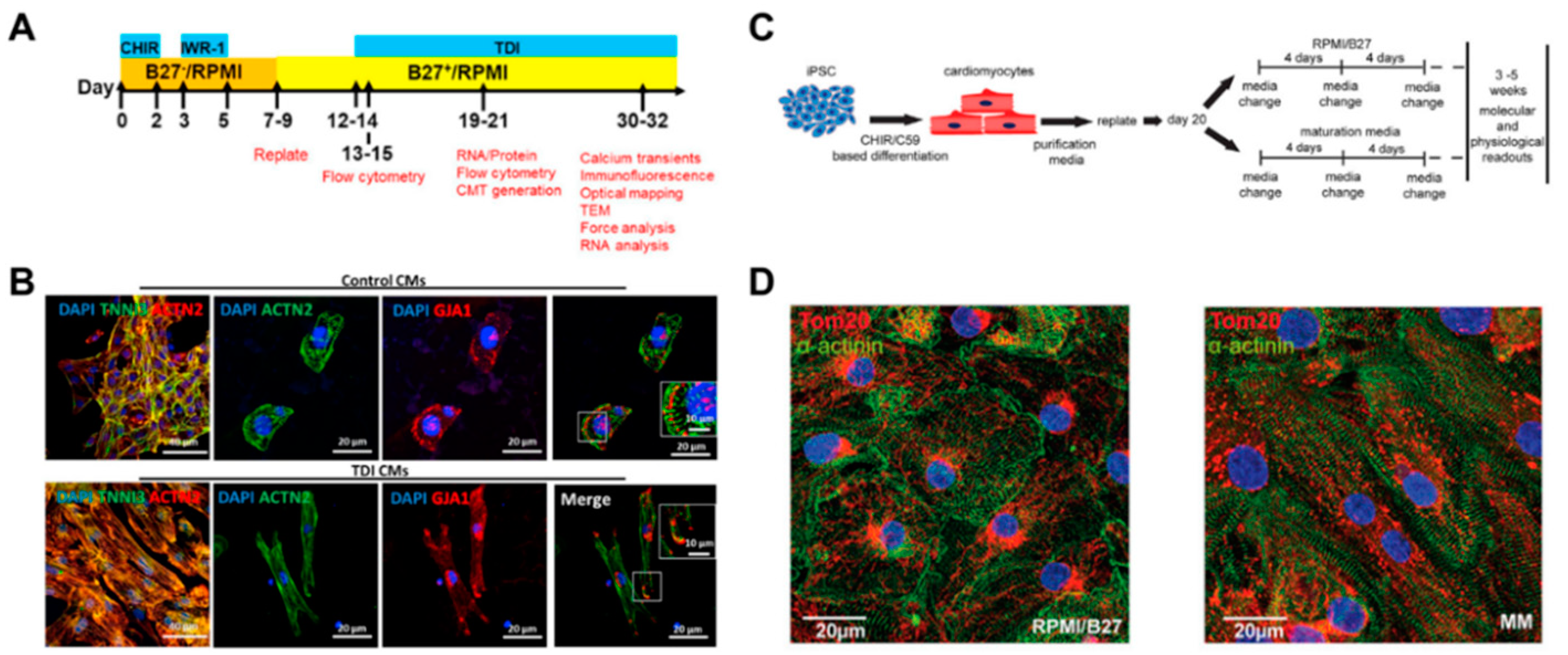
2.3.1. Cardiomyocyte Maturation by Biochemical Cues
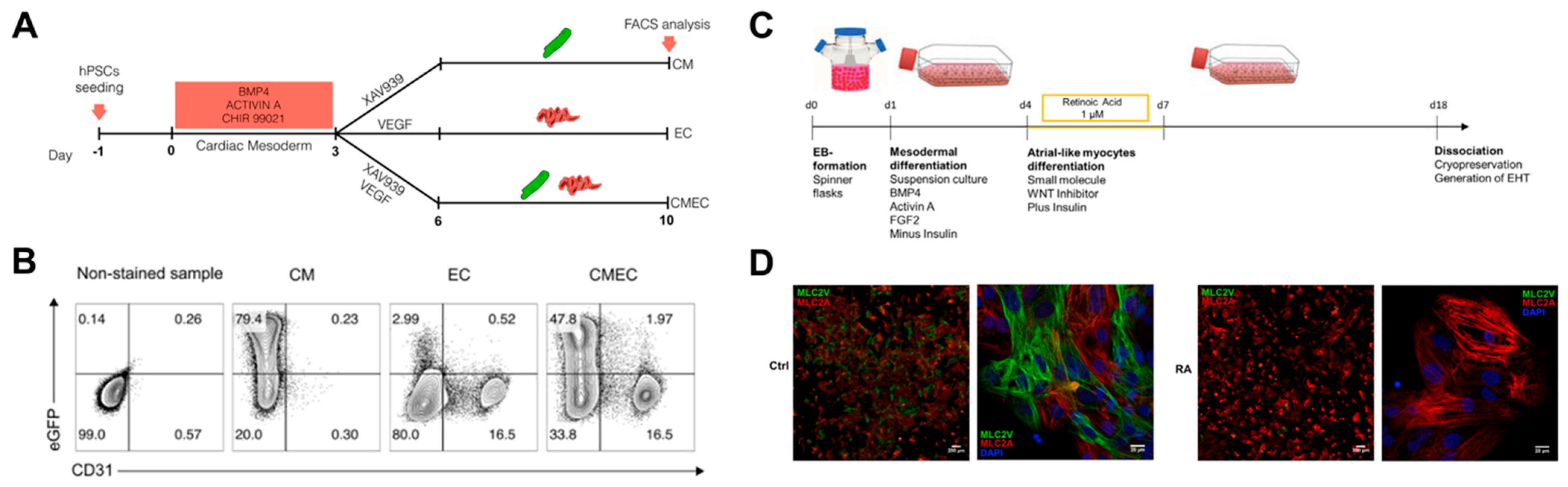
2.3.2. Chamber-Specific Differentiation by Biochemical Cues
2.3.3. Gene Editing Technologies for Disease Modeling In Vitro
3. The Cardiac ECM
3.1. Heart ECM Components
3.1.1. Basic Components of the Heart

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Structural | Semi-Structural | Non-Structural | |
|---|---|---|---|
| Glycoproteins | Fiber: Collagens Elastins (not glycosylated) [91] | Fibronectin Laminin [92] | Prototypical matricellular proteins [93] |
| Glycosaminoglycans | Chondroitin sulfate [94] | Hyaluronic acid(Hyaluronan) [95] | |
| Proteoglycans | Basement membrane proteoglycans Cell Surface Proteoglycans Small Leucine Rich Proteoglycans [96,97] |
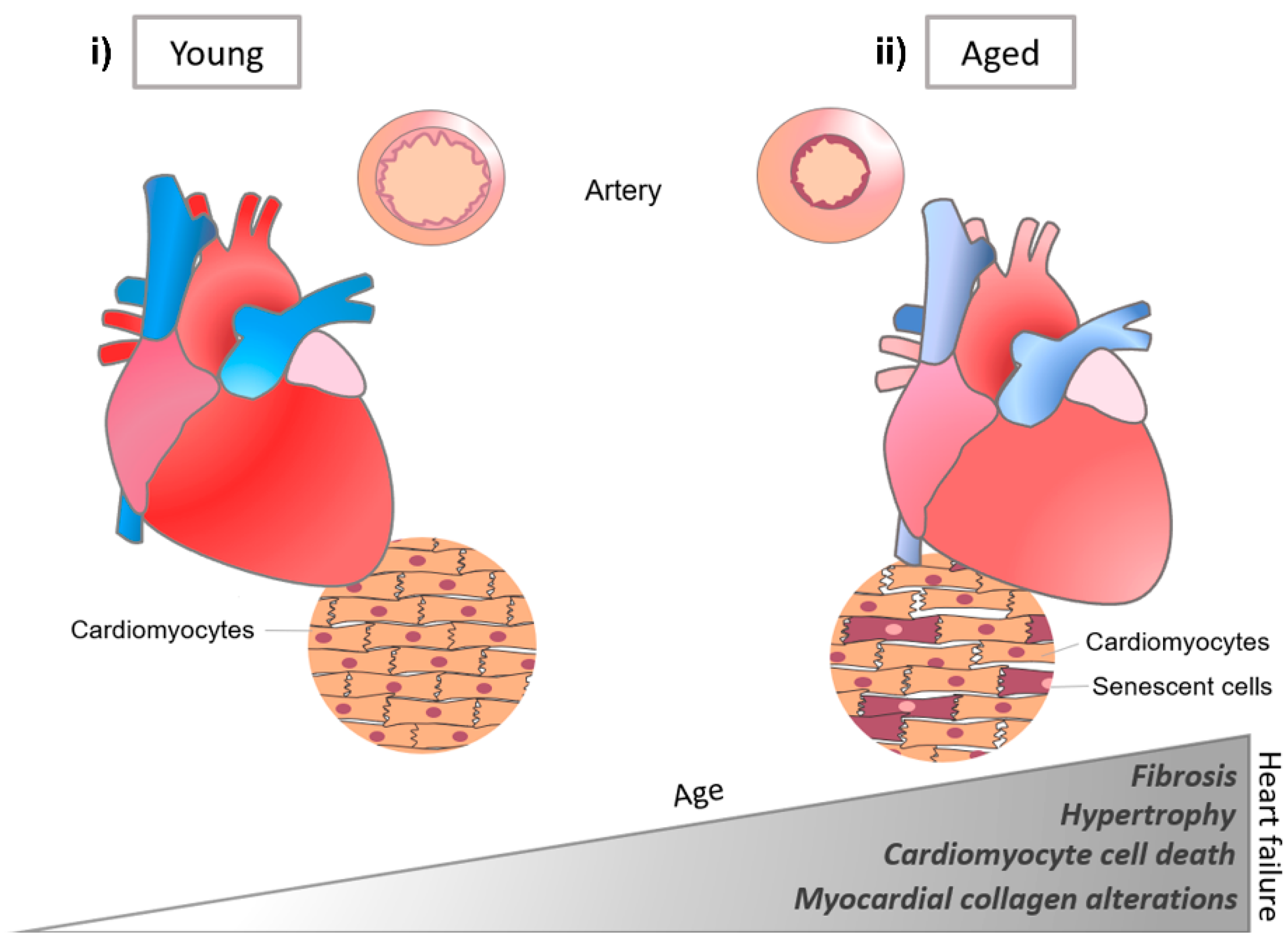
3.1.2. Cardiac ECM Components in Development and Disease
3.2. Naturally Derived ECMs in Cardiac Tissue Engineering
3.2.1. Collagen
3.2.2. Fibrin
3.2.3. Matrigel
3.2.4. Chitosan
3.2.5. Alginate
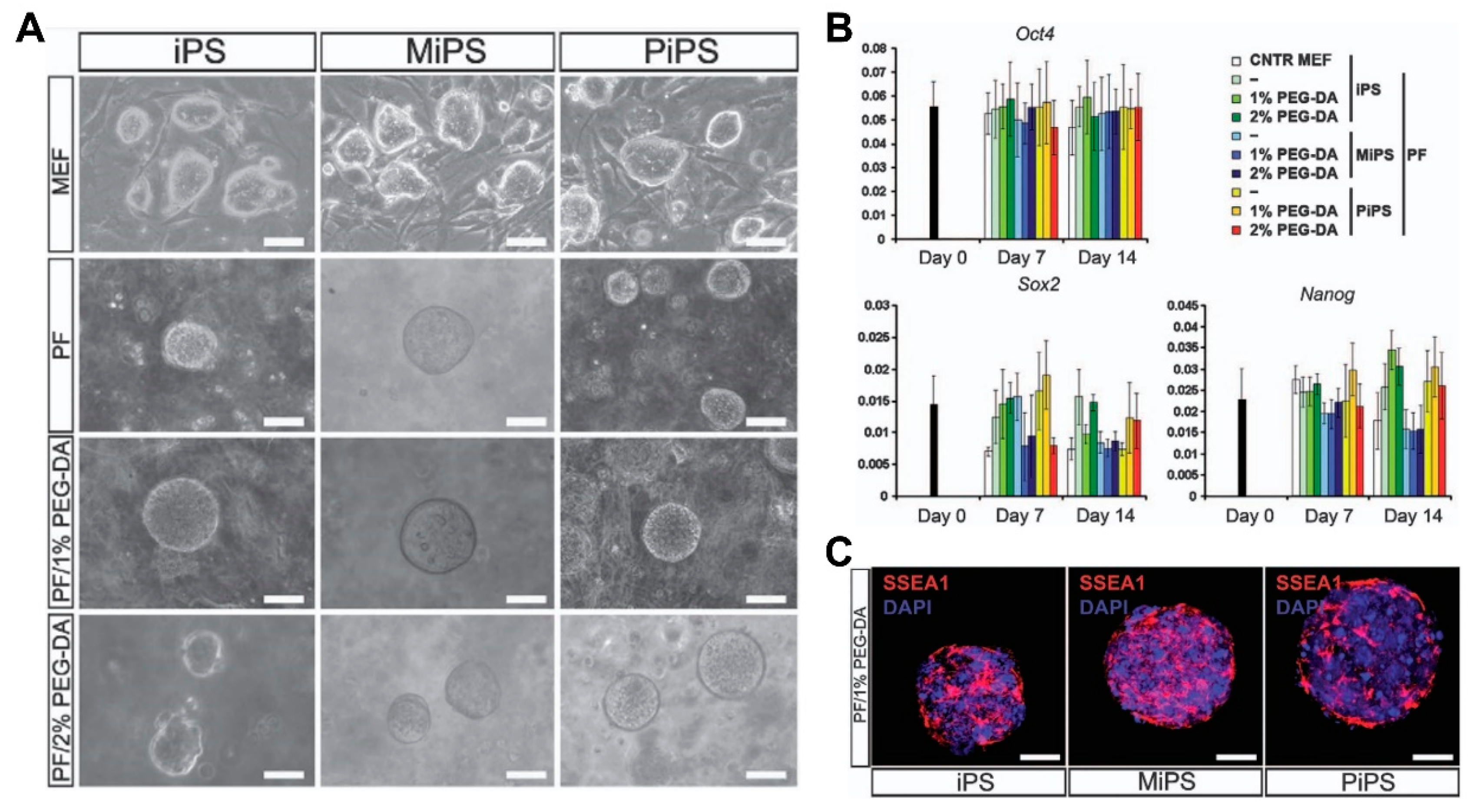
3.3. Decellularized ECM in Cardiac Tissue Engineering
3.3.1. Fabrication of dECM
3.3.2. dECM and Induced Cardiac Progenitor Cells (iCPCs)
3.4. Synthetic Materials for ECM in Cardiac Tissue Engineering
3.4.1. PEG (Poly(ethylene glycol))
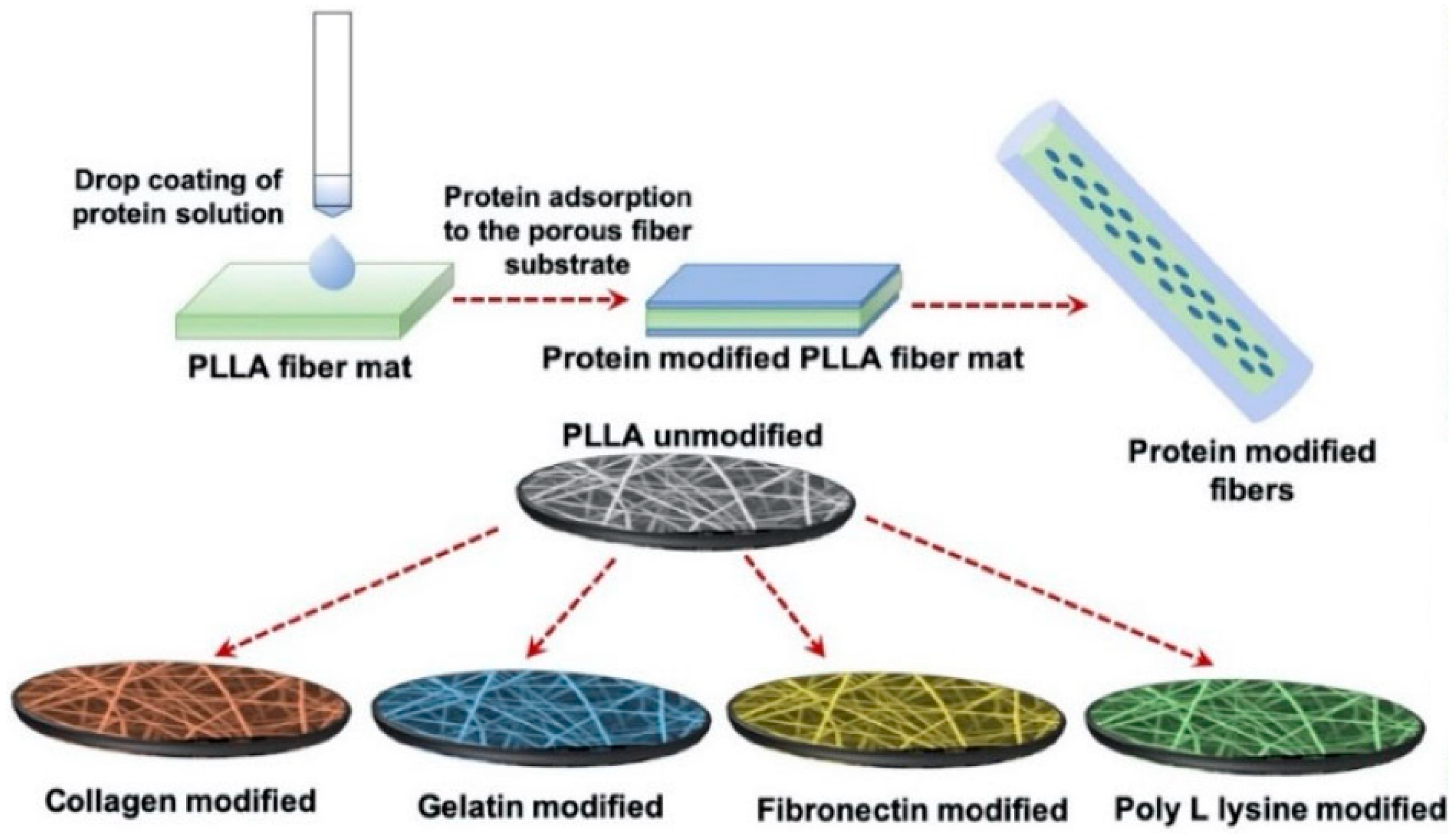
3.4.2. Poly(Lactic Acid)—PLLA Scaffolds
3.4.3. Poly(2-hydroxyethyl methacrylate)—PHEMA
4. Geometry
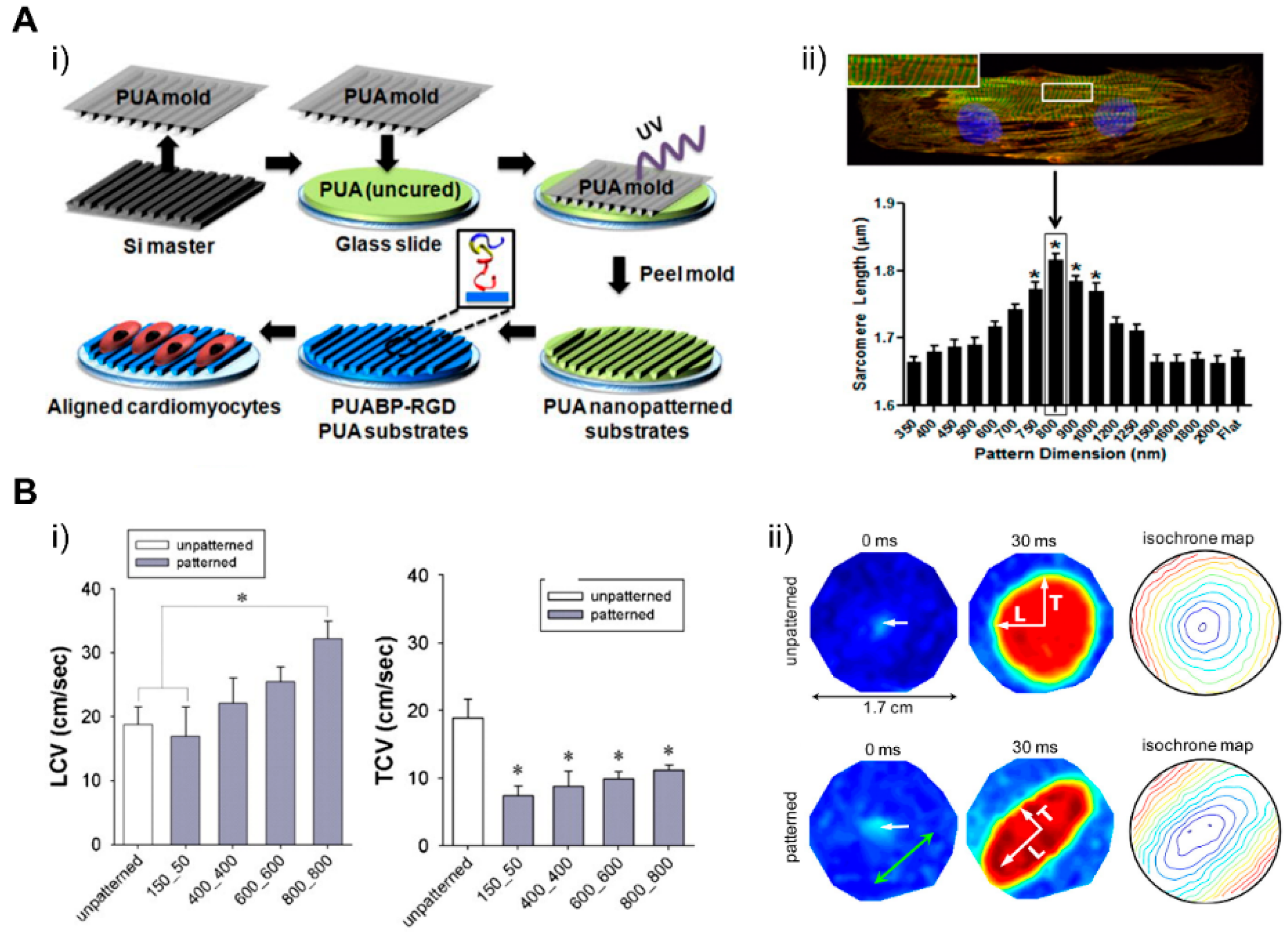
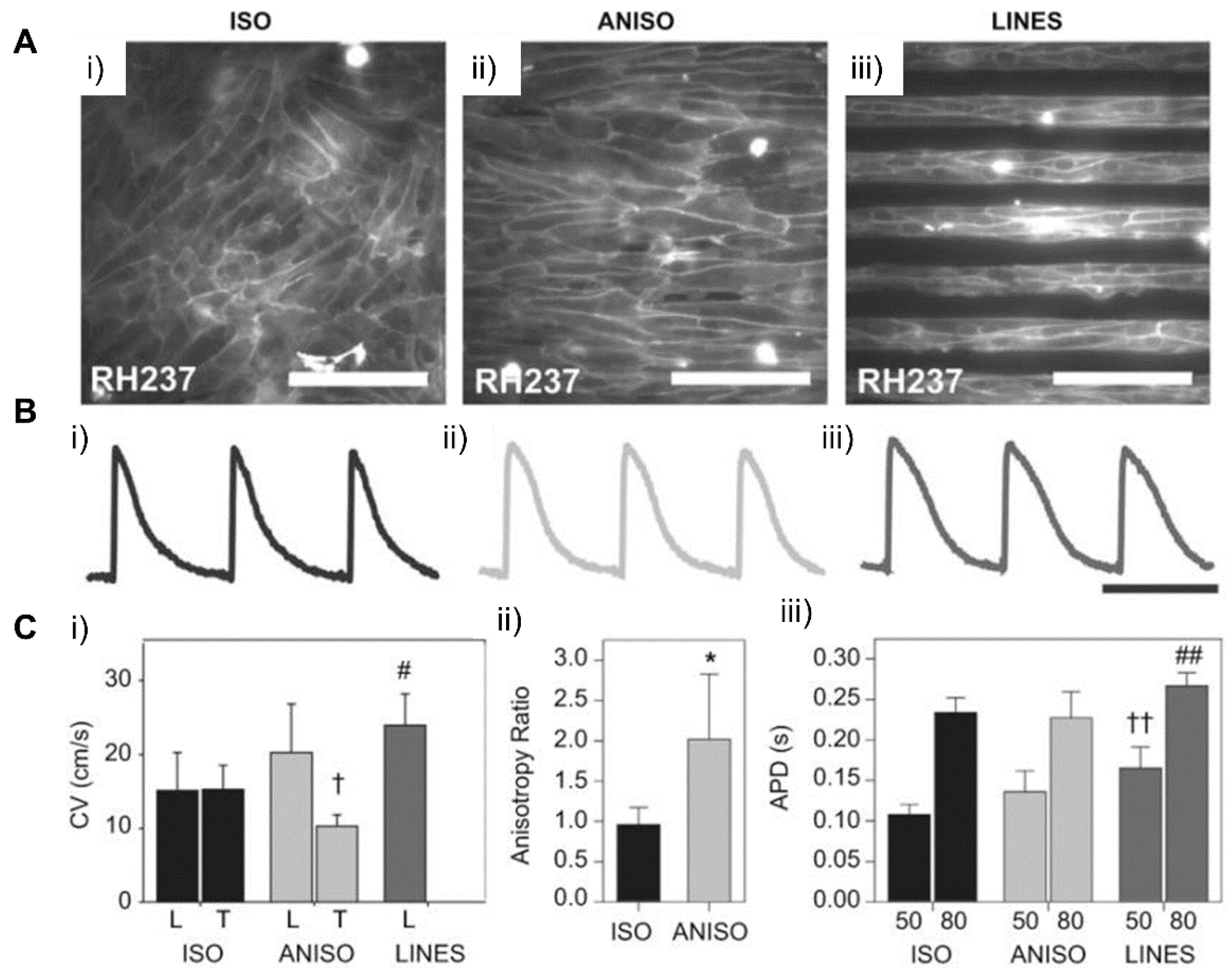
4.1. Patterning Geometry in Nanoscale
4.2. Patterning Geometry in Microscale
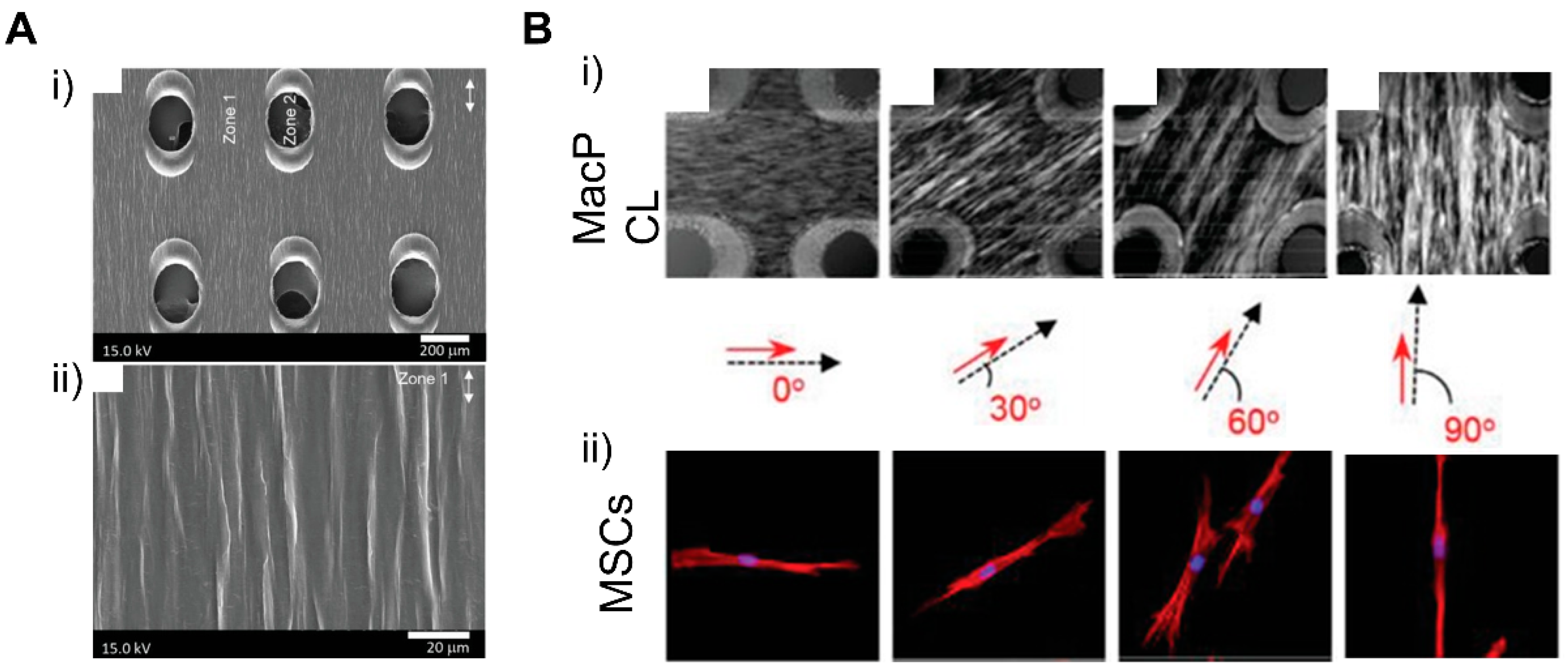
4.3. Patterning Geometry in Multiscale
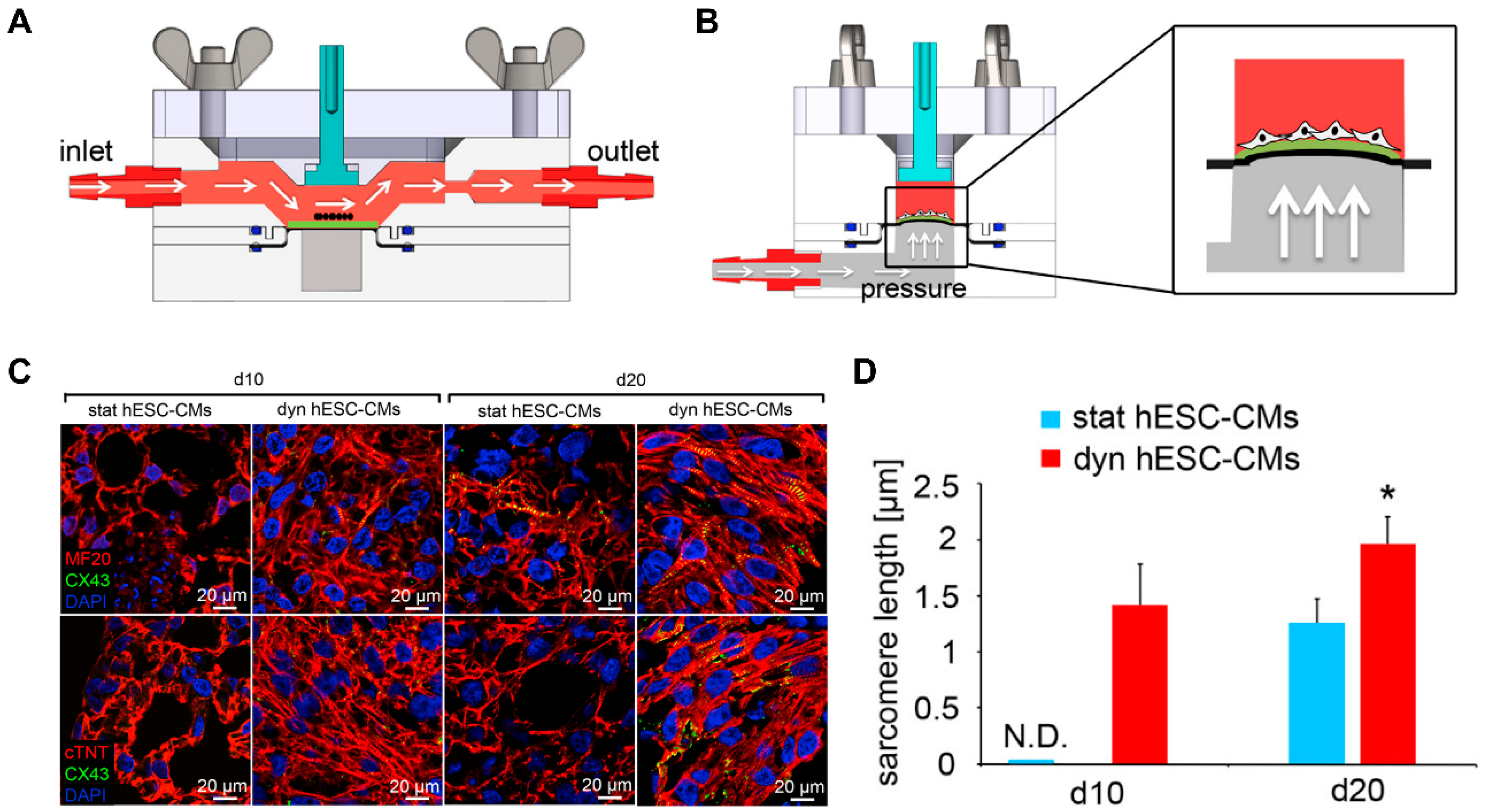
4.4. Dynamic Patterning
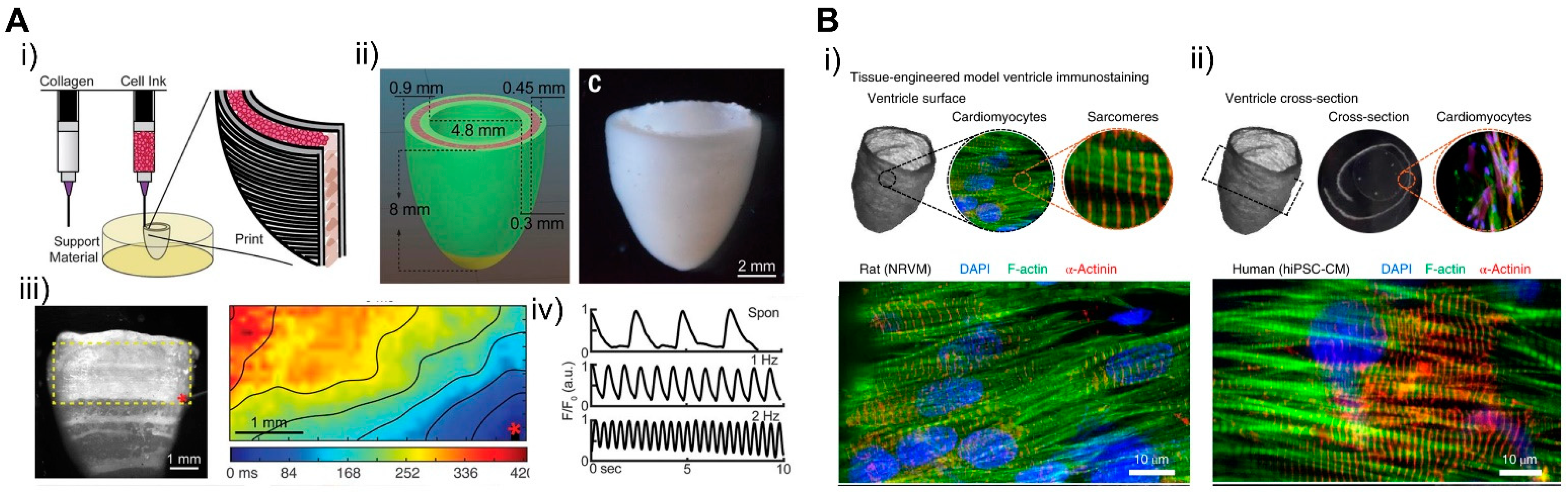
4.5. Patterning 3D in Macroscale
5. Computational Cardiology
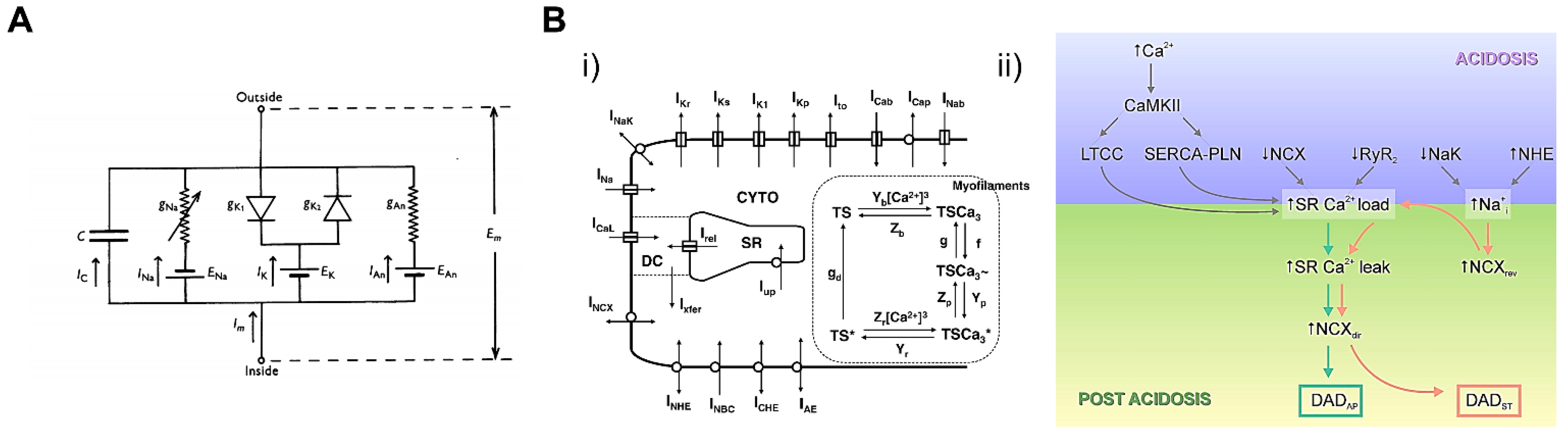
5.1. Cardiac Cellular Model
5.2. Whole-Heart Model
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mandrycky, C.J.; Williams, N.P.; Batalov, I.; El-Nachef, D.; De Bakker, B.S.; Davis, J.; Kim, D.-H.; Deforest, C.A.; Zheng, Y.; Stevens, K.R.; et al. Engineering Heart Morphogenesis. Trends Biotechnol. 2020, 38, 835–845. [Google Scholar] [CrossRef]
- Schwach, V.; Passier, R. Native cardiac environment and its impact on engineering cardiac tissue. Biomater. Sci. 2019, 7, 3566–3580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pomeroy, J.E.; Helfer, A.; Bursac, N. Biomaterializing the promise of cardiac tissue engineering. Biotechnol. Adv. 2020, 42, 107353. [Google Scholar] [CrossRef]
- Leitolis, A.; Robert, A.W.; Pereira, I.; Correa, A.; Stimamiglio, M.A. Cardiomyogenesis Modeling Using Pluripotent Stem Cells: The Role of Microenvironmental Signaling. Front. Cell Dev. Biol. 2019, 7, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.; Li, P.-H.; Chen, H.-Z. Cardiomyocyte Senescence and Cellular Communications Within Myocardial Microenvironments. Front. Endocrinol. 2020, 11, 280. [Google Scholar] [CrossRef]
- Vagnozzi, R.J.; Molkentin, J.D.; Houser, S.R. New Myocyte Formation in the Adult Heart. Circ. Res. 2018, 123, 159–176. [Google Scholar] [CrossRef]
- Porter, G.A.; Hom, J.R.; Hoffman, D.L.; Quintanilla, R.A.; Bentley, K.L.D.M.; Sheu, S.-S. Bioenergetics, mitochondria, and cardiac myocyte differentiation. Prog. Pediatr. Cardiol. 2011, 31, 75–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puente, B.N.; Kimura, W.; Muralidhar, S.A.; Moon, J.; Amatruda, J.F.; Phelps, K.L.; Grinsfelder, D.; Rothermel, B.A.; Chen, R.; Garcia, J.A.; et al. The Oxygen-Rich Postnatal Environment Induces Cardiomyocyte Cell-Cycle Arrest through DNA Damage Response. Cell 2014, 157, 565–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kempf, H.; Andree, B.; Zweigerdt, R. Large-scale production of human pluripotent stem cell derived cardiomyocytes. Adv. Drug Deliv. Rev. 2016, 96, 18–30. [Google Scholar] [CrossRef]
- Scuderi, G.J.; Butcher, J. Naturally Engineered Maturation of Cardiomyocytes. Front. Cell Dev. Biol. 2017, 5, 50. [Google Scholar] [CrossRef]
- Guimarães, C.F.; Gasperini, L.; Marques, A.P.; Reis, R.L. The stiffness of living tissues and its implications for tissue engineering. Nat. Rev. Mater. 2020, 5, 351–370. [Google Scholar] [CrossRef]
- Hussey, G.S.; Dziki, J.L.; Badylak, S.F. Extracellular matrix-based materials for regenerative medicine. Nat. Rev. Mater. 2018, 3, 159–173. [Google Scholar] [CrossRef]
- Jang, Y.; Park, Y.; Kim, J. Engineering Biomaterials to Guide Heart Cells for Matured Cardiac Tissue. Coatings 2020, 10, 925. [Google Scholar] [CrossRef]
- Danik, S.B.; Liu, F.; Zhang, J.; Suk, H.J.; Morley, G.E.; Fishman, G.I.; Gutstein, D.E. Modulation of cardiac gap junction expression and arrhythmic susceptibility. Circ. Res. 2004, 95, 1035–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagaraju, C.K.; Dries, E.; Gilbert, G.; Abdesselem, M.; Wang, N.; Amoni, M.; Driesen, R.B.; Sipido, K.R. Myofibroblast modulation of cardiac myocyte structure and function. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Seldin, M.M.; Fu, K.; Li, S.; Lam, L.; Wang, P.; Wang, Y.; Huang, D.; Nguyen, T.L.; Wei, B.; et al. Topological Arrangement of Cardiac Fibroblasts Regulates Cellular Plasticity. Circ. Res. 2018, 123, 73–85. [Google Scholar] [CrossRef]
- Carson, D.; Hnilova, M.; Yang, X.; Nemeth, C.L.; Tsui, J.H.; Smith, A.S.; Jiao, A.; Regnier, M.; Murry, C.E.; Tamerler, C.; et al. Nanotopography-Induced Structural Anisotropy and Sarcomere Development in Human Cardiomyocytes Derived from Induced Pluripotent Stem Cells. ACS Appl. Mater. Interfaces 2016, 8, 21923–21932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.-H.; Lipke, E.A.; Kim, P.; Cheong, R.; Thompson, S.; Delannoy, M.; Suh, K.-Y.; Tung, L.; Levchenko, A. Nanoscale cues regulate the structure and function of macroscopic cardiac tissue constructs. Proc. Natl. Acad. Sci. USA 2009, 107, 565–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, J.-L.; Tulloch, N.L.; Razumova, M.V.; Saiget, M.; Muskheli, V.; Pabon, L.; Reinecke, H.; Regnier, M.; Murry, C.E. Mechanical Stress Conditioning and Electrical Stimulation Promote Contractility and Force Maturation of Induced Pluripotent Stem Cell-Derived Human Cardiac Tissue. Circulation 2016, 134, 1557–1567. [Google Scholar] [CrossRef]
- Niederer, S.A.; Lumens, J.; Trayanova, N.A. Computational models in cardiology. Nat. Rev. Cardiol. 2019, 16, 100–111. [Google Scholar] [CrossRef]
- Mirams, G.R.; Pathmanathan, P.; Gray, R.A.; Challenor, P.; Clayton, R.H. Uncertainty and variability in computational and mathematical models of cardiac physiology. J. Physiol. 2016, 594, 6833–6847. [Google Scholar] [CrossRef]
- Usyk, T.P.; LeGrice, I.J.; McCulloch, A.D. Computational model of three-dimensional cardiac electromechanics. Comput. Vis. Sci. 2002, 4, 249–257. [Google Scholar] [CrossRef]
- Louch, W.E.; Sheehan, K.A.; Wolska, B.M. Methods in cardiomyocyte isolation, culture, and gene transfer. J. Mol. Cell. Cardiol. 2011, 51, 288–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaugg, M.; Xu, W.; Lucchinetti, E.; Shafiq, S.A.; Jamali, N.Z.; Siddiqui, M.A.Q. β-Adrenergic Receptor Subtypes Differentially Affect Apoptosis in Adult Rat Ventricular Myocytes. Circulation 2000, 102, 344–350. [Google Scholar] [CrossRef] [Green Version]
- Shizukuda, Y.; Buttrick, P.M. Subtype Specific Roles of β-Adrenergic Receptors in Apoptosis of Adult Rat Ventricular Myocytes. J. Mol. Cell. Cardiol. 2002, 34, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Rakhit, R.D.; Edwards, R.J.; Mockridge, J.W.; Baydoun, A.R.; Wyatt, A.W.; Mann, G.E.; Marber, M.S. Nitric oxide-induced cardioprotection in cultured rat ventricular myocytes. Am. J. Physiol. Circ. Physiol. 2000, 278, H1211–H1217. [Google Scholar] [CrossRef]
- Amin, J.K.; Xiao, L.; Pimental, D.R.; Pagano, P.J.; Singh, K.; Sawyer, D.B.; Colucci, W.S. Reactive Oxygen Species Mediate Alpha-adrenergic Receptor-stimulated Hypertrophy in Adult Rat Ventricular Myocytes. J. Mol. Cell. Cardiol. 2001, 33, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Ashley, E.A.; Sears, C.E.; Bryant, S.M.; Watkins, H.C.; Casadei, B. Cardiac Nitric Oxide Synthase 1 Regulates Basal and β-Adrenergic Contractility in Murine Ventricular Myocytes. Circulation 2002, 105, 3011–3016. [Google Scholar] [CrossRef] [Green Version]
- Snabaitis, A.K.; Hearse, D.J.; Avkiran, M. Regulation of sarcolemmal Na(+)/H(+) exchange by hydrogen peroxide in adult rat ventricular myocytes. Cardiovasc. Res. 2002, 53, 470–480. [Google Scholar] [CrossRef] [Green Version]
- Calaghan, S.; White, E. Caveolae modulate excitation–contraction coupling and β2-adrenergic signalling in adult rat ventricular myocytes. Cardiovasc. Res. 2006, 69, 816–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacot, J.G.; McCulloch, A.D.; Omens, J.H. Substrate Stiffness Affects the Functional Maturation of Neonatal Rat Ventricular Myocytes. Biophys. J. 2008, 95, 3479–3487. [Google Scholar] [CrossRef] [Green Version]
- Bian, W.; Badie, N.; Himel, H.D.; Bursac, N. Robust T-tubulation and maturation of cardiomyocytes using tissue-engineered epicardial mimetics. Biomaterials 2014, 35, 3819–3828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peter, A.K.; Bjerke, M.A.; Leinwand, L.A. Biology of the cardiac myocyte in heart disease. Mol. Biol. Cell 2016, 27, 2149–2160. [Google Scholar] [CrossRef] [PubMed]
- Cerbai, E.; Pino, R.; Sartiani, L.; Mugelli, A. Influence of postnatal-development on If occurrence and properties in neonatal rat ventricular myocytes. Cardiovasc. Res. 1999, 42, 416–423. [Google Scholar] [CrossRef] [Green Version]
- Müller-Ehmsen, J.; Whittaker, P.; Kloner, R.A.; Dow, J.S.; Sakoda, T.; Long, T.I.; Laird, P.W.; Kedes, L. Survival and Development of Neonatal Rat Cardiomyocytes Transplanted into Adult Myocardium. J. Mol. Cell. Cardiol. 2002, 34, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Jiang, C.; Hong, H.; Liu, J.; Qiu, L.; Huang, Y.; Ye, L. Effects of hypoxia on cardiomyocyte proliferation and association with stage of development. Biomed. Pharmacother. 2019, 118, 109391. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, Y.; Ding, G.; May, H.I.; Xu, J.; Gillette, T.G.; Wang, H.; Wang, Z.V. Temporal dynamics of cardiac hypertrophic growth in response to pressure overload. Am. J. Physiol. Circ. Physiol. 2017, 313, H1119–H1129. [Google Scholar] [CrossRef] [PubMed]
- Watkins, S.J.; Borthwick, G.M.; Arthur, H.M. The H9C2 cell line and primary neonatal cardiomyocyte cells show similar hypertrophic responses in vitro. Vitr. Cell. Dev. Biol. Anim. 2010, 47, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Hansen, A.; Eder, A.; Bönstrup, M.; Flato, M.; Mewe, M.; Schaaf, S.; Aksehirlioglu, B.; Schwörer, A.; Uebeler, J.; Eschenhagen, T. Development of a Drug Screening Platform Based on Engineered Heart Tissue. Circ. Res. 2010, 107, 35–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzatzalos, E.; Abilez, O.J.; Shukla, P.; Wu, J.C. Engineered heart tissues and induced pluripotent stem cells: Macro- and microstructures for disease modeling, drug screening, and translational studies. Adv. Drug Deliv. Rev. 2016, 96, 234–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wobus, A.M.; Wallukat, G.; Hescheler, J. Pluripotent mouse embryonic stem cells are able to differentiate into cardiomyocytes expressing chronotropic responses to adrenergic and cholinergic agents and Ca2+ channel blockers. Differentiation 1991, 48, 173–182. [Google Scholar] [CrossRef] [PubMed]
- A Maltsev, V.; Rohwedel, J.; Hescheler, J.; Wobus, A.M. Embryonic stem cells differentiate in vitro into cardiomyocytes representing sinusnodal, atrial and ventricular cell types. Mech. Dev. 1993, 44, 41–50. [Google Scholar] [CrossRef]
- Wobus, A.M.; Kaomei, G.; Shan, J.; Wellner, M.-C.; Rohwedel, J.; Guanju, J.; Fleischmann, B.; Katus, H.A.; Hescheler, J.; Franz, W.-M. Retinoic Acid Accelerates Embryonic Stem Cell-Derived Cardiac Differentiation and Enhances Development of Ventricular Cardiomyocytes. J. Mol. Cell. Cardiol. 1997, 29, 1525–1539. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Bradley, A. Mice deficient for BMP2 are nonviable and have defects in amnion/chorion and cardiac development. Development 1996, 122, 2977–2986. [Google Scholar]
- Mima, T.; Ueno, H.; Fischman, D.A.; Williams, L.T.; Mikawa, T. Fibroblast growth factor receptor is required for in vivo cardiac myocyte proliferation at early embryonic stages of heart development. Proc. Natl. Acad. Sci. USA 1995, 92, 467–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
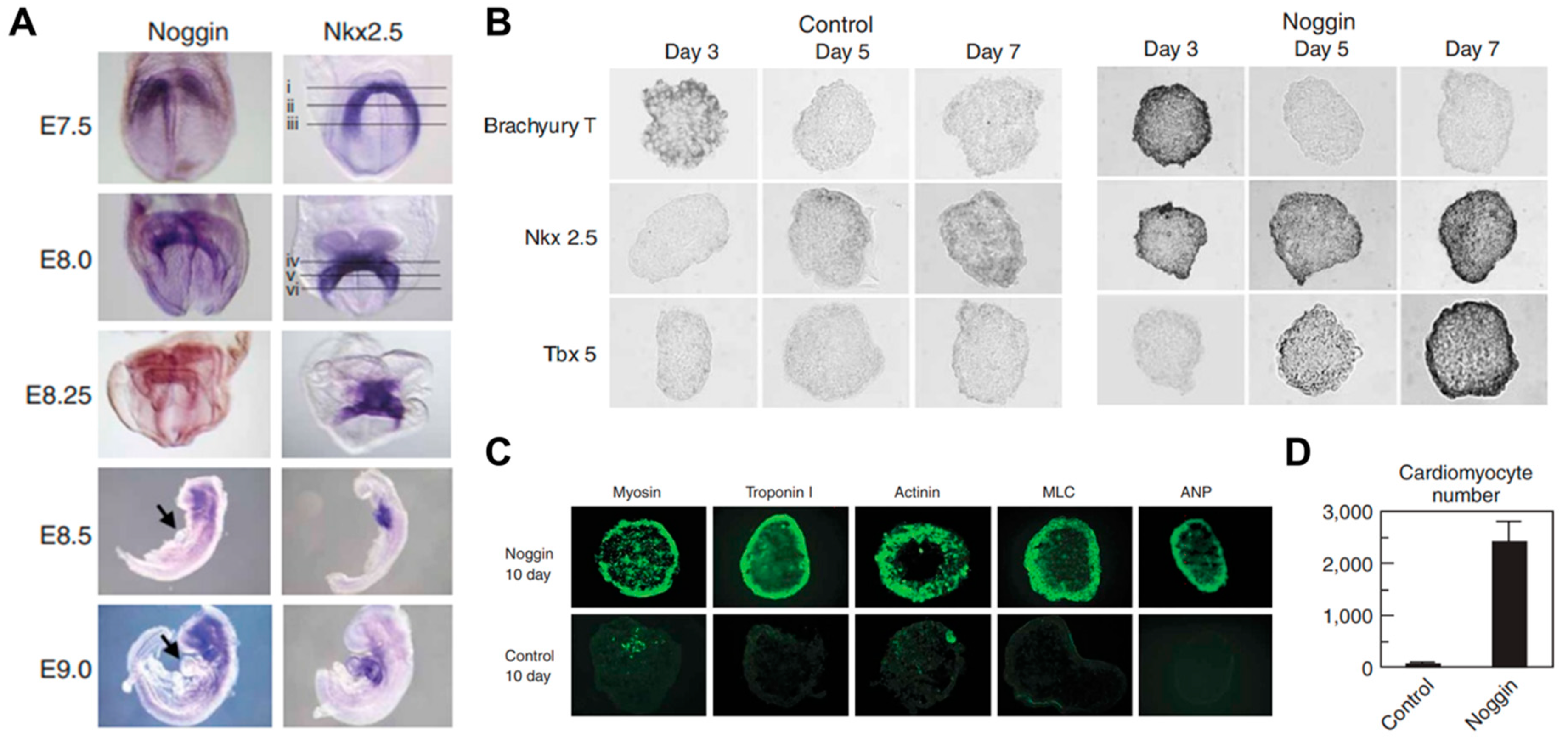
- Marvin, M.J.; Di Rocco, G.; Gardiner, A.; Bush, S.M.; Lassar, A.B. Inhibition of Wnt activity induces heart formation from posterior mesoderm. Genes Dev. 2001, 15, 316–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuasa, S.; Itabashi, Y.; Koshimizu, U.; Tanaka, T.; Sugimura, K.; Kinoshita, M.; Hattori, F.; Fukami, S.-I.; Shimazaki, T.; Okano, H.; et al. Transient inhibition of BMP signaling by Noggin induces cardiomyocyte differentiation of mouse embryonic stem cells. Nat. Biotechnol. 2005, 23, 607–611. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, H.; Olson, E.N.; Bassel-Duby, R. Therapeutic approaches for cardiac regeneration and repair. Nat. Rev. Cardiol. 2018, 15, 585–600. [Google Scholar] [CrossRef]
- Zwi, L.; Caspi, O.; Arbel, G.; Huber, I.; Gepstein, A.; Park, I.-H.; Gepstein, L. Cardiomyocyte Differentiation of Human Induced Pluripotent Stem Cells. Circulation 2009, 120, 1513–1523. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, Y.; Yamanaka, S. iPS cells: A source of cardiac regeneration. J. Mol. Cell. Cardiol. 2011, 50, 327–332. [Google Scholar] [CrossRef] [Green Version]
- Burridge, P.W.; Keller, G.; Gold, J.D.; Wu, J.C. Production of De Novo Cardiomyocytes: Human Pluripotent Stem Cell Differentiation and Direct Reprogramming. Cell Stem Cell 2012, 10, 16–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laflamme, M.A.; Chen, K.Y.; Naumova, A.V.; Muskheli, V.; Fugate, J.A.; Dupras, S.K.; Reinecke, H.; Xu, C.; Hassanipour, M.; Police, S.; et al. Cardiomyocytes derived from human embryonic stem cells in pro-survival factors enhance function of infarcted rat hearts. Nat. Biotechnol. 2007, 25, 1015–1024. [Google Scholar] [CrossRef]
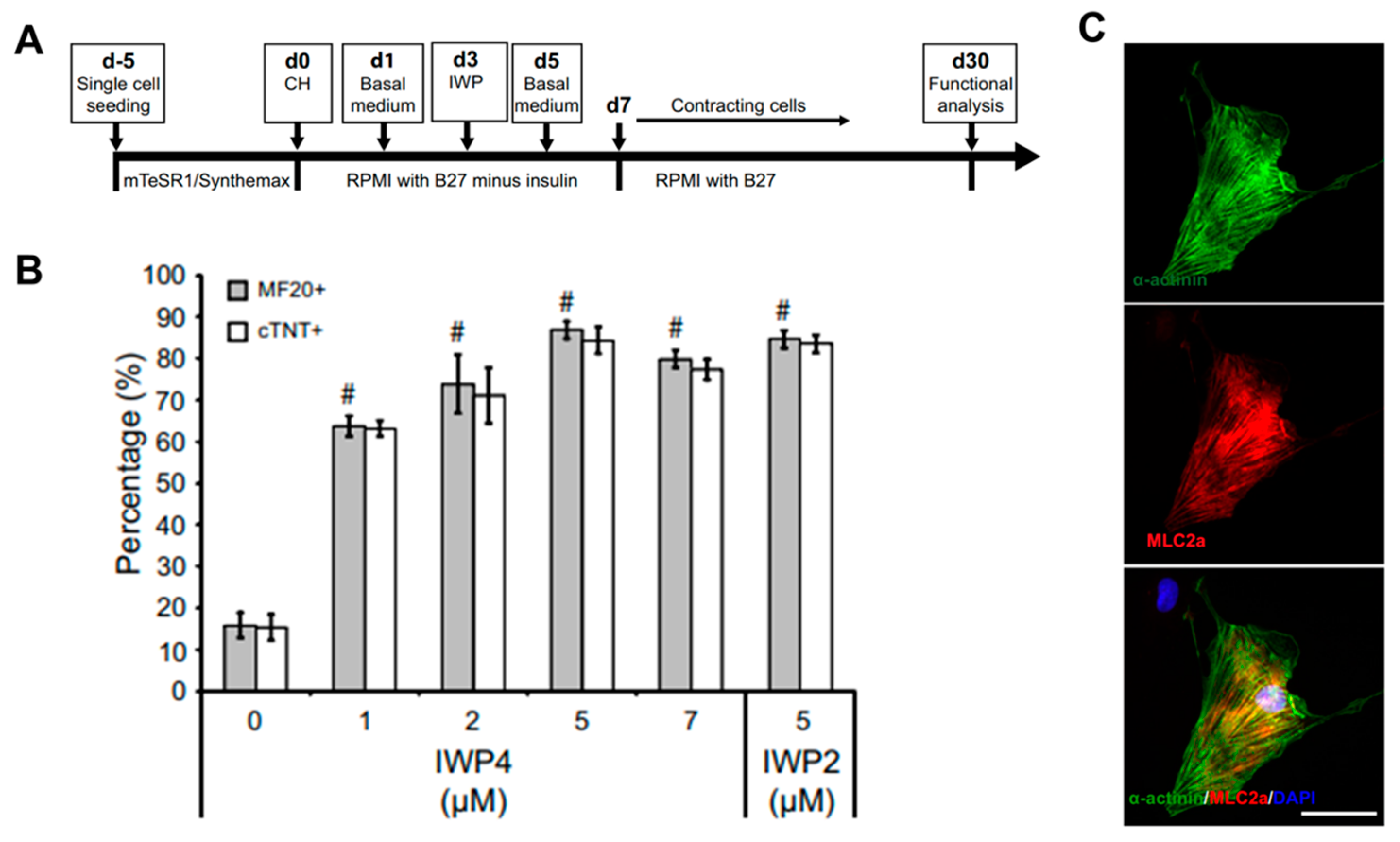
- Lian, X.; Hsiao, C.; Wilson, G.; Zhu, K.; Hazeltine, L.B.; Azarin, S.M.; Raval, K.K.; Zhang, J.; Kamp, T.J.; Palecek, S.P. Cozzarelli Prize Winner: Robust cardiomyocyte differentiation from human pluripotent stem cells via temporal modulation of canonical Wnt signaling. Proc. Natl. Acad. Sci. USA 2012, 109, E1848–E1857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burridge, P.W.; Matsa, E.; Shukla, P.; Lin, Z.C.; Churko, J.M.; Ebert, A.D.; Lan, F.; Diecke, S.; Huber, B.; Mordwinkin, N.M.; et al. Chemically defined generation of human cardiomyocytes. Nat. Methods 2014, 11, 855–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Pabon, L.; Murry, C.E. Engineering Adolescence. Circ. Res. 2014, 114, 511–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Rodriguez, M.L.; Leonard, A.; Sun, L.; Fischer, K.A.; Wang, Y.; Ritterhoff, J.; Zhao, L.; Kolwicz, S.C.; Pabon, L.; et al. Fatty Acids Enhance the Maturation of Cardiomyocytes Derived from Human Pluripotent Stem Cells. Stem Cell Rep. 2019, 13, 657–668. [Google Scholar] [CrossRef] [Green Version]
- Laflamme, M.A.; Murry, C.E. Heart regeneration. Nat. Cell Biol. 2011, 473, 326–335. [Google Scholar] [CrossRef] [Green Version]
- Gerdes, A.M.; Kellerman, S.E.; Moore, J.A.; Muffly, K.E.; Clarck, C.E.; Reaves, P.Y.; Malec, K.B.; McKeown, P.P.; Schocken, D.D. Structural Remodeling of Cardiac Myocytes in Patients With Ischemic Cardiomyopathy. Circulation 1992, 86, 426–430. [Google Scholar] [CrossRef] [Green Version]
- Snir, M.; Kehat, I.; Gepstein, A.; Coleman, R.; Itskovitz-Eldor, J.; Livne, E.; Gepstein, L. Assessment of the ultrastructural and proliferative properties of human embryonic stem cell-derived cardiomyocytes. Am. J. Physiol. Circ. Physiol. 2003, 285, H2355–H2363. [Google Scholar] [CrossRef] [Green Version]
- Gutstein, D.E.; Liu, F.-Y.; Meyers, M.B.; Choo, A.; Fishman, G.I. The organization of adherens junctions and desmosomes at the cardiac intercalated disc is independent of gap junctions. J. Cell Sci. 2003, 116, 875–885. [Google Scholar] [CrossRef] [Green Version]
- Sottas, V.; Wahl, C.-M.; Trache, M.C.; Bartolf-Kopp, M.; Cambridge, S.; Hecker, M.; Ullrich, N.D. Improving electrical properties of iPSC-cardiomyocytes by enhancing Cx43 expression. J. Mol. Cell. Cardiol. 2018, 120, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.; Choi, S.-C.; Lim, D.-S.; Kim, J.-H.; Kim, J.; Park, Y. Modulating cardiomyocyte and fibroblast interaction using layer-by-layer deposition facilitates synchronisation of cardiac macro tissues. Soft Matter 2020, 16, 428–434. [Google Scholar] [CrossRef] [PubMed]
- Lundy, S.D.; Zhu, W.-Z.; Regnier, M.; Laflamme, M.A. Structural and Functional Maturation of Cardiomyocytes Derived from Human Pluripotent Stem Cells. Stem Cells Dev. 2013, 22, 1991–2002. [Google Scholar] [CrossRef] [Green Version]
- Bird, S.; Doevendans, P.; Van Rooijen, M.; De La Riviere, A.B.; Hassink, R.; Passier, R.; Mummery, C. The human adult cardiomyocyte phenotype. Cardiovasc. Res. 2003, 58, 423–434. [Google Scholar] [CrossRef]
- Agarkova, I.; Perriard, J.-C. The M-band: An elastic web that crosslinks thick filaments in the center of the sarcomere. Trends Cell Biol. 2005, 15, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Laksman, Z.; Backx, P.H. The electrophysiological development of cardiomyocytes. Adv. Drug Deliv. Rev. 2016, 96, 253–273. [Google Scholar] [CrossRef]
- Liu, A.; Tang, M.; Xi, J.; Gao, L.; Zheng, Y.; Luo, H.; Hu, X.; Zhao, F.; Reppel, M.; Hescheler, J.; et al. Functional Characterization of Inward Rectifier Potassium Ion Channel in Murine Fetal Ventricular Cardiomyocytes. Cell. Physiol. Biochem. 2010, 26, 413–420. [Google Scholar] [CrossRef]
- Balse, E.; Steele, D.F.; Abriel, H.; Coulombe, A.; Fedida, D.; Hatem, S.N. Dynamic of Ion Channel Expression at the Plasma Membrane of Cardiomyocytes. Physiol. Rev. 2012, 92, 1317–1358. [Google Scholar] [CrossRef]
- Lopaschuk, G.D.; Jaswal, J.S. Energy Metabolic Phenotype of the Cardiomyocyte During Development, Differentiation, and Postnatal Maturation. J. Cardiovasc. Pharmacol. 2010, 56, 130–140. [Google Scholar] [CrossRef]
- Werner, J.C.; Sicard, R.E.; Schuller, H.G. Palmitate oxidation by isolated working fetal and newborn pig hearts. Endocrinol. Metab. 1989, 256, 315–321. [Google Scholar] [CrossRef]
- Schaper, J.; Meiser, E.; Stämmler, G. Ultrastructural morphometric analysis of myocardium from dogs, rats, hamsters, mice, and from human hearts. Circ. Res. 1985, 56, 377–391. [Google Scholar] [CrossRef] [Green Version]
- Parikh, S.S.; Blackwell, D.J.; Gomez-Hurtado, N.; Frisk, M.; Wang, L.; Kim, K.; Dahl, C.P.; Fiane, A.; Tønnessen, T.; Kryshtal, D.O.; et al. Thyroid and Glucocorticoid Hormones Promote Functional T-Tubule Development in Human-Induced Pluripotent Stem Cell–Derived Cardiomyocytes. Circ. Res. 2017, 121, 1323–1330. [Google Scholar] [CrossRef]
- Huang, C.Y.; Maia-Joca, R.P.M.; Ong, C.S.; Wilson, I.; DiSilvestre, D.; Tomaselli, G.F.; Reich, D.H. Enhancement of human iPSC-derived cardiomyocyte maturation by chemical conditioning in a 3D environment. J. Mol. Cell. Cardiol. 2020, 138, 1–11. [Google Scholar] [CrossRef]
- Makinde, A.-O.; Kantor, P.F.; Lopaschuk, G.D. Maturation of fatty acid and carbohydrate metabolism in the newborn heart. Mol. Cell. Biochem. 1998, 188, 49–56. [Google Scholar] [CrossRef]
- Feyen, D.A.; McKeithan, W.L.; Bruyneel, A.A.; Spiering, S.; Hörmann, L.; Ulmer, B.; Zhang, H.; Briganti, F.; Schweizer, M.; Hegyi, B.; et al. Metabolic Maturation Media Improve Physiological Function of Human iPSC-Derived Cardiomyocytes. Cell Rep. 2020, 32, 107925. [Google Scholar] [CrossRef]
- Giacomelli, E.; Bellin, M.; Sala, L.; Van Meer, B.J.; Tertoolen, L.G.J.; Orlova, V.V.; Mummery, C.L. Three-dimensional cardiac microtissues composed of cardiomyocytes and endothelial cells co-differentiated from human pluripotent stem cells. Development 2017, 144, 1008–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Protze, S.I.; Laksman, Z.; Backx, P.H.; Keller, G.M. Human Pluripotent Stem Cell-Derived Atrial and Ventricular Cardiomyocytes Develop from Distinct Mesoderm Populations. Cell Stem Cell 2017, 21, 179–194.e4. [Google Scholar] [CrossRef] [PubMed]
- Lemme, M.; Ulmer, B.M.; Lemoine, M.D.; Zech, A.T.; Flenner, F.; Ravens, U.; Reichenspurner, H.; Rol-Garcia, M.; Smith, G.; Hansen, A.; et al. Atrial-like Engineered Heart Tissue: An In Vitro Model of the Human Atrium. Stem Cell Rep. 2018, 11, 1378–1390. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Rafatian, N.; Feric, N.T.; Cox, B.J.; Aschar-Sobbi, R.; Wang, E.Y.; Aggarwal, P.; Zhang, B.; Conant, G.; Ronaldson-Bouchard, K.; et al. A Platform for Generation of Chamber-Specific Cardiac Tissues and Disease Modeling. Cell 2019, 176, 913–927.e18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaunbrecher, R.J.; Abel, A.N.; Kim, D.-H.; Linke, W.A.; Sniadecki, N.J.; Regnier, M.; Murry, C.E.; Beussman, K.; Leonard, A.; Von Frieling-Salewsky, M.; et al. Cronos Titin Is Expressed in Human Cardiomyocytes and Necessary for Normal Sarcomere Function. Circulation 2019, 140, 1647–1660. [Google Scholar] [CrossRef]
- Brandão, K.O.; Brink, L.V.D.; Miller, D.C.; Grandela, C.; Van Meer, B.J.; Mol, M.P.; De Korte, T.; Tertoolen, L.G.; Mummery, C.L.; Sala, L.; et al. Isogenic Sets of hiPSC-CMs Harboring Distinct KCNH2 Mutations Differ Functionally and in Susceptibility to Drug-Induced Arrhythmias. Stem Cell Rep. 2020, 15, 1127–1139. [Google Scholar] [CrossRef] [PubMed]
- Rienks, M.; Papageorgiou, A.-P.; Frangogiannis, N.G.; Heymans, S. Myocardial Extracellular Matrix. Circ. Res. 2014, 114, 872–888. [Google Scholar] [CrossRef] [Green Version]
- Baudino, T.A.; Carver, W.; Giles, W.; Borg, T.K. Cardiac fibroblasts: Friend or foe? Am. J. Physiol. Circ. Physiol. 2006, 291, H1015–H1026. [Google Scholar] [CrossRef]
- Frangogiannis, N.G. The Extracellular Matrix in Ischemic and Nonischemic Heart Failure. Circ. Res. 2019, 125, 117–146. [Google Scholar] [CrossRef]
- Horn, M.A.; Trafford, A.W. Aging and the cardiac collagen matrix: Novel mediators of fibrotic remodelling. J. Mol. Cell. Cardiol. 2016, 93, 175–185. [Google Scholar] [CrossRef] [Green Version]
- Kwak, H.-B. Aging, exercise, and extracellular matrix in the heart. J. Exerc. Rehabilitation 2013, 9, 338–347. [Google Scholar] [CrossRef]
- Brower, G.L.; Gardner, J.D.; Forman, M.F.; Murray, D.B.; Voloshenyuk, T.; Levick, S.P.; Janicki, J.S. The relationship between myocardial extracellular matrix remodeling and ventricular function? Eur. J. Cardio-Thoracic Surg. 2006, 30, 604–610. [Google Scholar] [CrossRef]
- Yokota, T.; McCourt, J.; Ma, F.; Ren, S.; Li, S.; Kim, T.-H.; Kurmangaliyev, Y.Z.; Nasiri, R.; Ahadian, S.; Nguyen, T.; et al. Type V Collagen in Scar Tissue Regulates the Size of Scar after Heart Injury. Cell 2020, 182, 545–562.e23. [Google Scholar] [CrossRef]
- Frangogiannis, N.G. The extracellular matrix in myocardial injury, repair, and remodeling. J. Clin. Investig. 2017, 127, 1600–1612. [Google Scholar] [CrossRef] [Green Version]
- Behrens, D.T.; Villone, D.; Koch, M.; Brunner, G.; Sorokin, L.; Robenek, H.; Bruckner-Tuderman, L.; Bruckner, P.; Hansen, U. The Epidermal Basement Membrane Is a Composite of Separate Laminin- or Collagen IV-containing Networks Connected by Aggregated Perlecan, but Not by Nidogens. J. Biol. Chem. 2012, 287, 18700–18709. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Mao, C.; Jia, Y.; Fu, Y.; Kong, W. Extracellular matrix dynamics in vascular remodeling. Am. J. Physiol. Physiol. 2020, 319, C481–C499. [Google Scholar] [CrossRef]
- Jallerat, Q.; Feinberg, A.W. Extracellular Matrix Structure and Composition in the Early Four-Chambered Embryonic Heart. Cells 2020, 9, 285. [Google Scholar] [CrossRef] [Green Version]
- McCurdy, S.M.; Dai, Q.; Zhang, J.; Zamilpa, R.; Ramirez, T.A.; Dayah, T.; Nguyen, N.; Jin, Y.-F.; Bradshaw, A.D.; Lindsey, M.L. SPARC mediates early extracellular matrix remodeling following myocardial infarction. Am. J. Physiol. Circ. Physiol. 2011, 301, H497–H505. [Google Scholar] [CrossRef] [Green Version]
- Murata, K.; Oshima, Y. Chondroitin sulfates in atherosclerotic human aorta. Atherosclerosis 1971, 14, 121–129. [Google Scholar] [CrossRef]
- Raman, R.; Sasisekharan, V.; Sasisekharan, R. Structural Insights into Biological Roles of Protein-Glycosaminoglycan Interactions. Chem. Biol. 2005, 12, 267–277. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Fan, C.; Zhuang, Y.; Sun, J.; Hou, X.; Chen, B.; Xiao, Z.; Chen, Y.; Zhan, Z.; Zhao, Y.; et al. Heparan sulfate proteoglycan promotes fibroblast growth factor-2 function for ischemic heart repair. Biomater. Sci. 2019, 7, 5438–5450. [Google Scholar] [CrossRef]
- Andenæs, K.; Lunde, I.G.; Mohammadzadeh, N.; Dahl, C.P.; Aronsen, J.M.; Strand, M.E.; Palmero, S.; Sjaastad, I.; Christensen, G.; Engebretsen, K.V.T.; et al. The extracellular matrix proteoglycan fibromodulin is upregulated in clinical and experimental heart failure and affects cardiac remodeling. PLoS ONE 2018, 13, e0201422. [Google Scholar] [CrossRef]
- Marijianowski, M.M.; Van Der Loos, C.M.; Mohrschladt, M.F.; Becker, A.E. The neonatal heart has a relatively high content of total collagen and type I collagen, a condition that may explain the less compliant state. J. Am. Coll. Cardiol. 1994, 23, 1204–1208. [Google Scholar] [CrossRef] [Green Version]
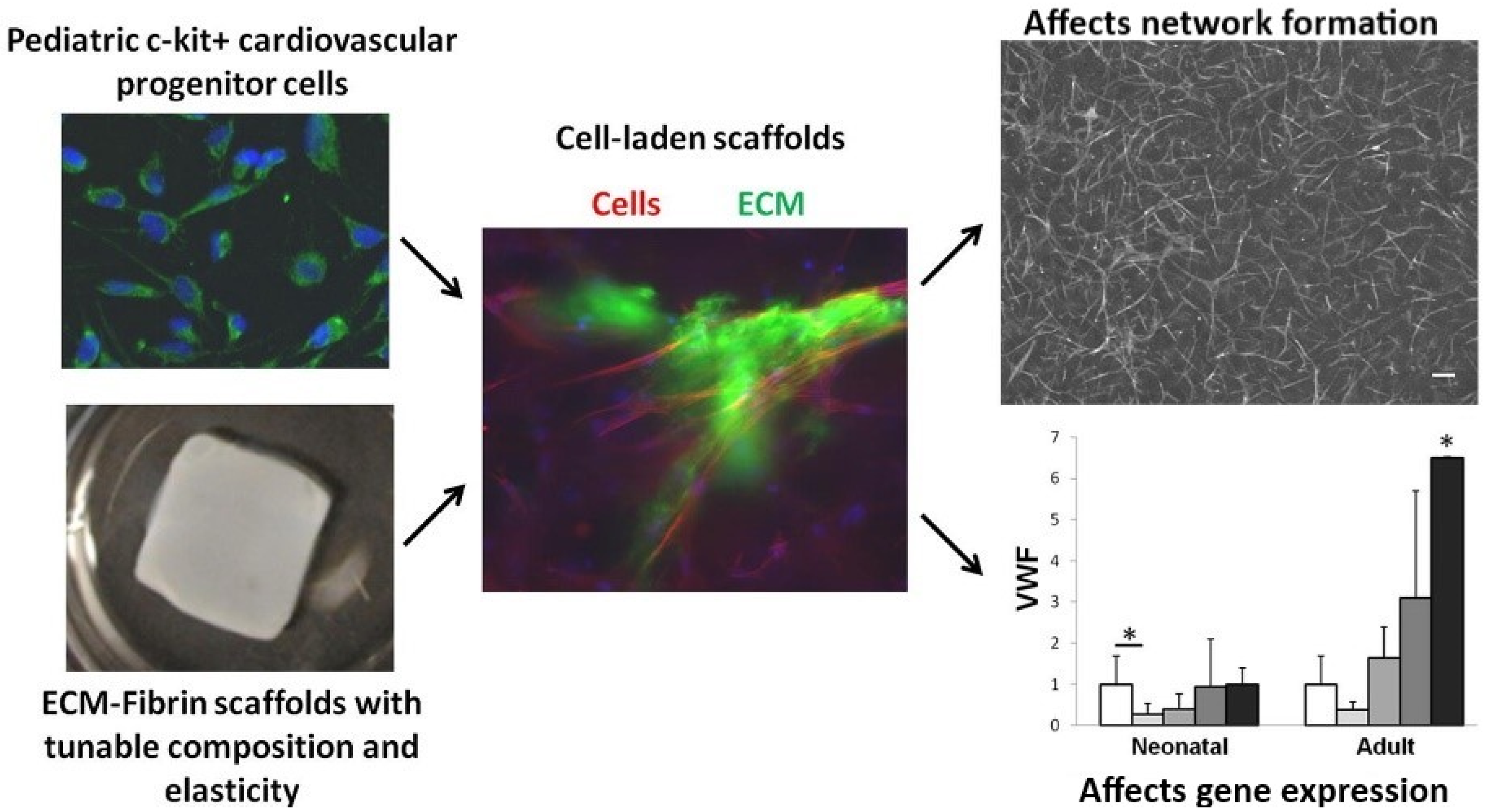
- Williams, C.; Quinn, K.; Georgakoudi, I.; Black, L. Young developmental age cardiac extracellular matrix promotes the expansion of neonatal cardiomyocytes in vitro. Acta Biomater. 2014, 10, 194–204. [Google Scholar] [CrossRef] [Green Version]
- I Jugdutt, B. Aging-related Changes in Cardiac Extracellular Matrix: Implications for Heart Failure in Older Patients. J. Cardiol. Curr. Res. 2015, 3, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Iop, L.; Sasso, E.D.; Menabò, R.; Di Lisa, F.; Gerosa, G. The Rapidly Evolving Concept of Whole Heart Engineering. Stem Cells Int. 2017, 2017, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Goldsmith, E.C.; Borg, T.K. The dynamic interaction of the extracellular matrix in cardiac remodeling. J. Card. Fail. 2002, 8, S314–S318. [Google Scholar] [CrossRef]
- Cleutjens, J.P.; Creemers, E.E. Integration of concepts: Cardiac extracellular matrix remodeling after myocardial infarction. J. Card. Fail. 2002, 8, S344–S348. [Google Scholar] [CrossRef]
- Pauschinger, M.; Knopf, D.; Petschauer, S.; Doerner, A.; Poller, W.; Schwimmbeck, P.L.; Kühl, U.; Schultheiss, H.-P. Dilated Cardiomyopathy Is Associated With Significant Changes in Collagen Type I/III ratio. Circulation 1999, 99, 2750–2756. [Google Scholar] [CrossRef] [Green Version]
- De Souza, R.R. Aging of myocardial collagen. Biogerontology 2002, 3, 325–335. [Google Scholar] [CrossRef]
- Hinderer, S.; Schenke-Layland, K. Cardiac fibrosis–A short review of causes and therapeutic strategies. Adv. Drug Deliv. Rev. 2019, 146, 77–82. [Google Scholar] [CrossRef]
- Copes, F.; Pien, N.; Van Vlierberghe, S.; Boccafoschi, F.; Mantovani, D. Collagen-Based Tissue Engineering Strategies for Vascular Medicine. Front. Bioeng. Biotechnol. 2019, 7, 166. [Google Scholar] [CrossRef] [Green Version]
- Perea-Gil, I.; Prat-Vidal, C.; Bayes-Genis, A. In vivo experience with natural scaffolds for myocardial infarction: The times they are a-changin’. Stem Cell Res. Ther. 2015, 6, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.-Q.; Peng, S.; Song, Z.-Y.; Lin, S. Collagen biomaterial for the treatment of myocardial infarction: An update on cardiac tissue engineering and myocardial regeneration. Drug Deliv. Transl. Res. 2019, 9, 1–15. [Google Scholar] [CrossRef]
- Chan, E.C.; Kuo, S.-M.; Kong, A.M.; Morrison, W.A.; Dusting, G.J.; Mitchell, G.M.; Lim, S.Y.; Liu, G.-S. Three Dimensional Collagen Scaffold Promotes Intrinsic Vascularisation for Tissue Engineering Applications. PLoS ONE 2016, 11, e0149799. [Google Scholar] [CrossRef] [Green Version]
- Rashedi, I.; Talele, N.; Wang, X.-H.; Hinz, B.; Radisic, M.; Keating, A. Collagen scaffold enhances the regenerative properties of mesenchymal stromal cells. PLoS ONE 2017, 12, e0187348. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; Radisic, M.; Lim, J.O.; Chang, B.H.; Vunjak-Novakovic, G. A novel composite scaffold for cardiac tissue engineering. Vitro Cell. Dev. Biol. Animal 2005, 41, 188–196. [Google Scholar] [CrossRef]
- Law, J.X.; Liau, L.L.; Saim, A.; Yang, Y.; Idrus, R. Electrospun Collagen Nanofibers and Their Applications in Skin Tissue Engineering. Tissue Eng. Regen. Med. 2017, 14, 699–718. [Google Scholar] [CrossRef]
- Liu, S.; Wu, J.; Liu, X.; Chen, D.; Bowlin, G.L.; Cao, L.; Lu, J.; Li, F.; Mo, X.; Fan, C. Osteochondral regeneration using an oriented nanofiber yarn-collagen type I/hyaluronate hybrid/TCP biphasic scaffold. J. Biomed. Mater. Res. Part A 2014, 103, 581–592. [Google Scholar] [CrossRef]
- Ishii, O.; Shin, M.; Sueda, T.; Vacanti, J.P. In vitro tissue engineering of a cardiac graft using a degradable scaffold with an extracellular matrix–like topography. J. Thorac. Cardiovasc. Surg. 2005, 130, 1358–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hitscherich, P.; Aphale, A.; Gordan, R.; Whitaker, R.; Singh, P.; Xie, L.-H.; Patra, P.; Lee, E.J. Electroactive graphene composite scaffolds for cardiac tissue engineering. J. Biomed. Mater. Res. Part A 2018, 106, 2923–2933. [Google Scholar] [CrossRef]
- O’Neill, H.S.; O’Sullivan, J.; Porteous, N.; Ruiz-Hernandez, E.; Kelly, H.M.; O’Brien, F.J.; Duffy, G.P. A collagen cardiac patch incorporating alginate microparticles permits the controlled release of hepatocyte growth factor and insulin-like growth factor-1 to enhance cardiac stem cell migration and proliferation. J. Tissue Eng. Regen. Med. 2018, 12, e384–e394. [Google Scholar] [CrossRef] [PubMed]
- Yost, M.J.; Baicu, C.F.; Stonerock, C.E.; Goodwin, R.L.; Price, R.L.; Davis, J.M.; Evans, H.; Watson, P.D.; Gore, C.M.; Sweet, J.; et al. A Novel Tubular Scaffold for Cardiovascular Tissue Engineering. Tissue Eng. 2004, 10, 273–284. [Google Scholar] [CrossRef]
- Tedder, M.E.; Liao, J.; Weed, B.; Stabler, C.; Zhang, H.; Simionescu, A.; Simionescu, D.T. Stabilized Collagen Scaffolds for Heart Valve Tissue Engineering. Tissue Eng. 2009, 15, 1257–1268. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.H.; Shoichet, M.S.; Radisic, M. Vascular endothelial growth factor immobilized in collagen scaffold promotes penetration and proliferation of endothelial cells. Acta Biomater. 2008, 4, 477–489. [Google Scholar] [CrossRef]
- McLaughlin, S.; McNeill, B.; Podrebarac, J.; Hosoyama, K.; Sedlakova, V.; Cron, G.; Smyth, D.; Seymour, R.; Goel, K.; Liang, W.; et al. Injectable human recombinant collagen matrices limit adverse remodeling and improve cardiac function after myocardial infarction. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Li, Q.; Zhao, Y.; Chen, W.; Chen, B.; Xiao, Z.; Lin, H.; Nie, L.; Wang, D.; Dai, J. Stem-cell-capturing collagen scaffold promotes cardiac tissue regeneration. Biomaterials 2011, 32, 2508–2515. [Google Scholar] [CrossRef]
- Neuta, P.A.; Rojas, D.M.; Agredo, W.; Gutierrez, J.O. Evaluation of the repairing effect of collagen type I and MaxGel on the infarcted myocardium in an animal model. In Proceedings of the 37th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Milan, Italy, 25–29 August 2015. [Google Scholar] [CrossRef]
- Kaiser, N.J.; Kant, R.J.; Minor, A.J.; Coulombe, K.L. Optimizing Blended Collagen-Fibrin Hydrogels for Cardiac Tissue Engineering with Human iPSC-derived Cardiomyocytes. ACS Biomater. Sci. Eng. 2019, 5, 887–899. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.; Budina, E.; Stoppel, W.L.; Sullivan, K.E.; Emani, S.; Emani, S.M.; Black, L.D. Cardiac extracellular matrix–fibrin hybrid scaffolds with tunable properties for cardiovascular tissue engineering. Acta Biomater. 2015, 14, 84–95. [Google Scholar] [CrossRef] [Green Version]
- Funamoto, S.; Hashimoto, Y.; Kishida, A.; Negishi, J. A fibrin-coated pericardial extracellular matrix prevented heart adhesion in a rat model. J. Biomed. Mater. Res. Part B Appl. Biomater. 2019, 107, 1088–1094. [Google Scholar] [CrossRef]
- Birla, R.K.; Borschel, G.H.; Dennis, R.G.; Brown, D.L. Myocardial Engineering in Vivo: Formation and Characterization of Contractile, Vascularized Three-Dimensional Cardiac Tissue. Tissue Eng. 2005, 11, 803–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.-C.; Khait, L.; Birla, R.K. Contractile three-dimensional bioengineered heart muscle for myocardial regeneration. J. Biomed. Mater. Res. Part A 2007, 80, 719–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, L.; Gregorich, Z.R.; Fast, V.G.; Hu, X.; Lloyd, S.G.; Ge, Y.; Zhang, J.; Zhu, W.; Mattapally, S.; Oduk, Y.; et al. Large Cardiac Muscle Patches Engineered From Human Induced-Pluripotent Stem Cell–Derived Cardiac Cells Improve Recovery From Myocardial Infarction in Swine. Circulation 2018, 137, 1712–1730. [Google Scholar] [CrossRef] [PubMed]
- Wendel, J.S.; Ye, L.; Tao, R.; Zhang, J.; Zhang, J.; Kamp, T.J.; Tranquillo, R.T. Functional Effects of a Tissue-Engineered Cardiac Patch From Human Induced Pluripotent Stem Cell-Derived Cardiomyocytes in a Rat Infarct Model. Stem Cells Transl. Med. 2015, 4, 1324–1332. [Google Scholar] [CrossRef]
- Arnaoutova, I.; George, J.; Kleinman, H.K.; Benton, G. The endothelial cell tube formation assay on basement membrane turns 20: State of the science and the art. Angiogenesis 2009, 12, 267–274. [Google Scholar] [CrossRef]
- Hughes, C.S.; Postovit, L.M.; Lajoie, G.A. Matrigel: A complex protein mixture required for optimal growth of cell culture. Proteomics 2010, 10, 1886–1890. [Google Scholar] [CrossRef]
- Li, Z.; Guan, J. Hydrogels for Cardiac Tissue Engineering. Polymers 2011, 3, 740–761. [Google Scholar] [CrossRef] [Green Version]
- Bakunts, K.; Gillum, N.; Karabekian, Z.; Sarvazyan, N. Formation of cardiac fibers in Matrigel matrix. Biotechniques 2008, 44, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Pan, J.; Qiu, F.; Wei, T.; Li, Z.; Jiang, X.; Zhuang, W.; Liu, Z.; Zhang, Z.; Dong, H. A novel modified physiologically relevant model for cardiac angiogenesis. Microvasc. Res. 2017, 114, 84–91. [Google Scholar] [CrossRef]
- Zhang, L.; Li, X.; Yu, X.; Li, Y.; Sun, A.; Huang, C.; Xu, F.; Guo, J.; Sun, Y.; Zhang, X.; et al. Construction of vascularized pacemaker tissues by seeding cardiac progenitor cells and endothelial progenitor cells into Matrigel. Life Sci. 2017, 179, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Tormos, C.; Madihally, S. Chitosan for cardiac tissue engineering and regeneration. In Chitosan Based Biomaterials Volume 2; Elsevier BV: Cambridge, UK, 2017; Volume 2, pp. 115–143. [Google Scholar] [CrossRef]
- Shigemasa, Y.; Saito, K.; Shashiwa, H.; Saimoto, H. Enzymatic degradation of chitins and partially deacetylated chitins. Int. J. Biol. Macromol. 1993, 16, 43–49. [Google Scholar] [CrossRef]
- Khor, E.; Lim, L.Y. Implantable applications of chitin and chitosan. Biomaterials 2003, 24, 2339–2349. [Google Scholar] [CrossRef]
- Madihally, S.V.; Matthew, H.W. Porous chitosan scaffolds for tissue engineering. Biomaterials 1999, 20, 1133–1142. [Google Scholar] [CrossRef]
- Shu, Y.; Hao, T.; Yao, F.; Qian, Y.; Wang, Y.; Yang, B.; Fanglian, Y.; Wang, C. RoY Peptide-Modified Chitosan-Based Hydrogel to Improve Angiogenesis and Cardiac Repair under Hypoxia. ACS Appl. Mater. Interfaces 2015, 7, 6505–6517. [Google Scholar] [CrossRef]
- Pourfarhangi, K.E.; Mashayekhan, S.; Asl, S.G.; Hajebrahimi, Z. Construction of scaffolds composed of acellular cardiac extracellular matrix for myocardial tissue engineering. Biologicals 2018, 53, 10–18. [Google Scholar] [CrossRef]
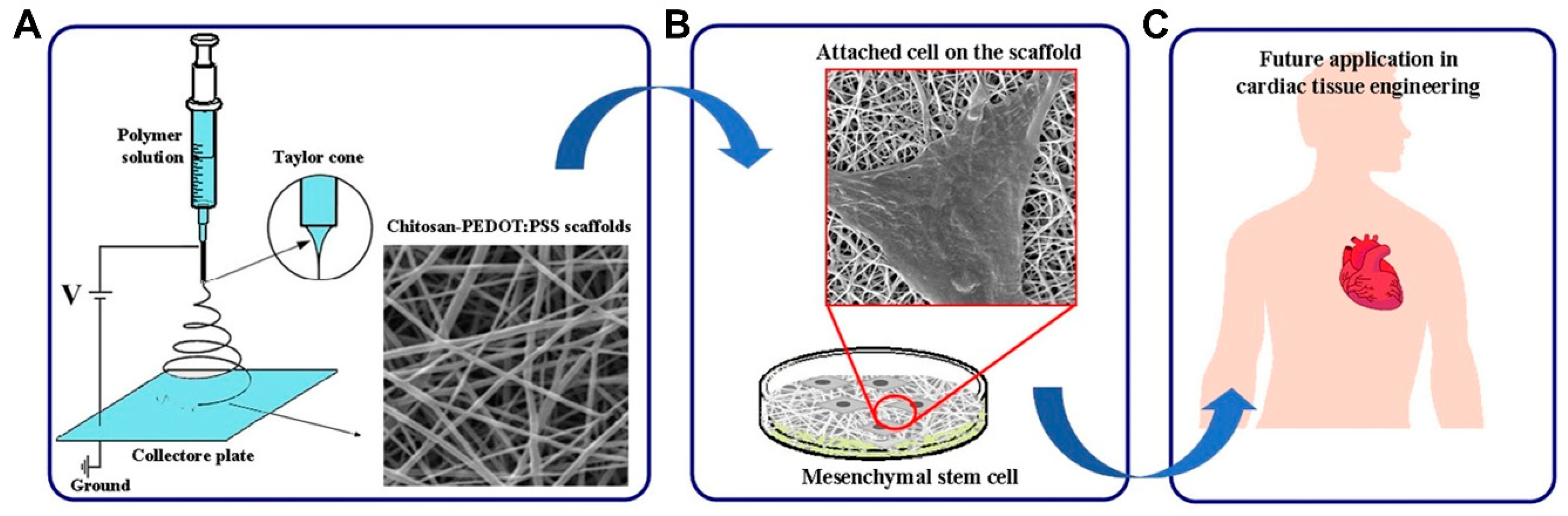
- Abedi, A.; Hasanzadeh, M.; Tayebi, L. Conductive nanofibrous Chitosan/PEDOT:PSS tissue engineering scaffolds. Mater. Chem. Phys. 2019, 237, 121882. [Google Scholar] [CrossRef]
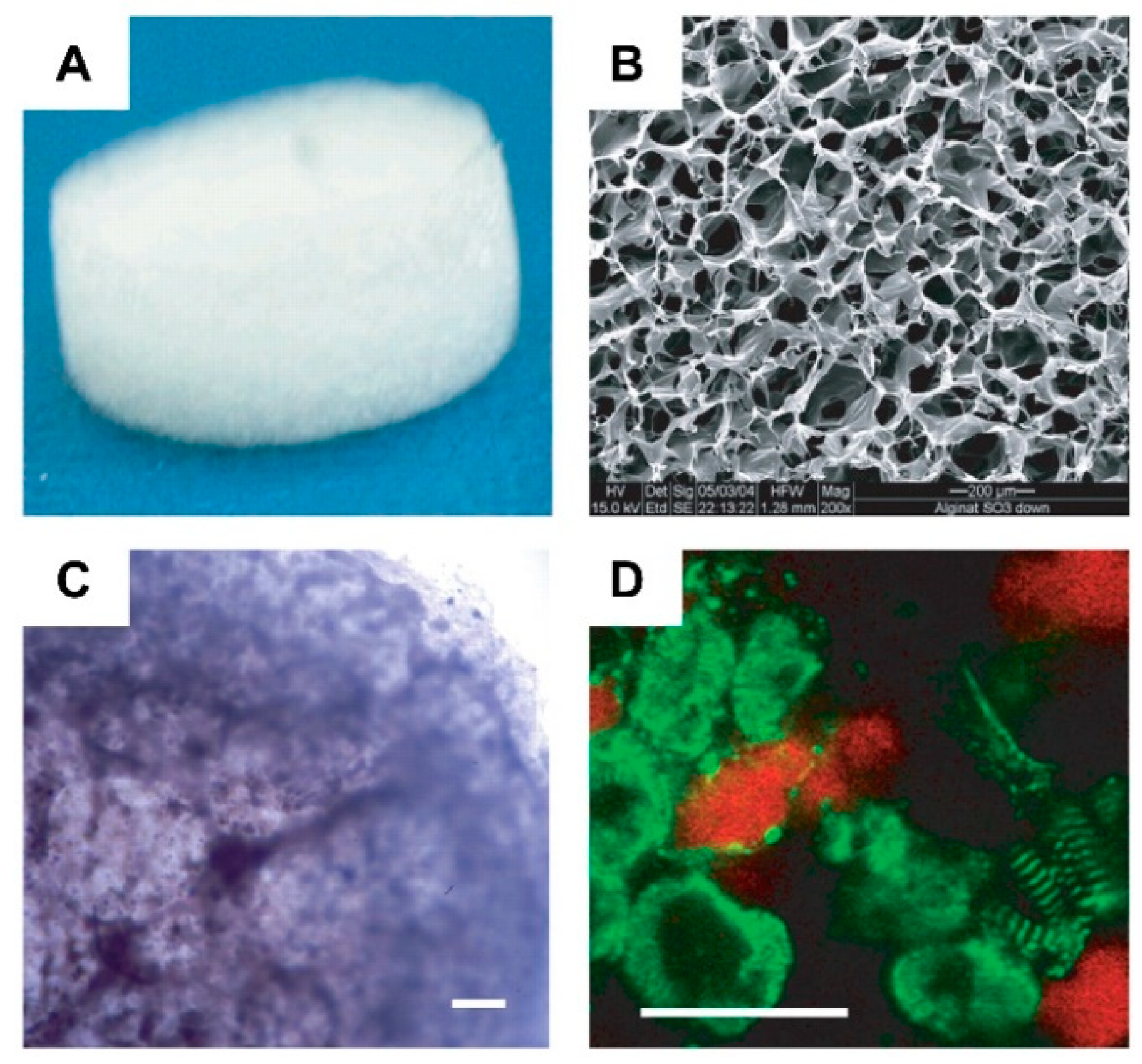
- Liberski, A.; Latif, N.; Raynaud, C.; Bollensdorff, C.; Yacoub, M. Alginate for cardiac regeneration: From seaweed to clinical trials. Glob. Cardiol. Sci. Pr. 2016, 2016, e201604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, T.; Li, J.; Yao, F.; Dong, D.; Wang, Y.; Yang, B.; Wang, C. Injectable Fullerenol/Alginate Hydrogel for Suppression of Oxidative Stress Damage in Brown Adipose-Derived Stem Cells and Cardiac Repair. ACS Nano 2017, 11, 5474–5488. [Google Scholar] [CrossRef]
- Curley, C.J.; Dolan, E.B.; Otten, M.; Hinderer, S.; Duffy, G.P.; Murphy, B.P. An injectable alginate/extra cellular matrix (ECM) hydrogel towards acellular treatment of heart failure. Drug Deliv. Transl. Res. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Xu, Z.; Lam, M.T. Alginate Application for Heart and Cardiovascular Diseases. In Alginates and Their Biomedical Applications; Springer: Singapore, 2017; Volume 11, pp. 185–212. [Google Scholar] [CrossRef]
- Zhang, P.; Zhang, H.; Wang, H.; Wei, Y.; Hu, S. Artificial Matrix Helps Neonatal Cardiomyocytes Restore Injured Myocardium in Rats. Artif. Organs 2006, 30, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.F. On the nature of biomaterials. Biomaterials 2009, 30, 5897–5909. [Google Scholar] [CrossRef] [PubMed]
- Dvir, T.; Kedem, A.; Leor, J.; Cohen, S.; Ruvinov, E.; Levy, O.; Freeman, I.; Landa, N.; Holbova, R.; Feinberg, M.S.; et al. Prevascularization of cardiac patch on the omentum improves its therapeutic outcome. Proc. Natl. Acad. Sci. USA 2009, 106, 14990–14995. [Google Scholar] [CrossRef] [Green Version]
- Christman, K.L.; Lee, R.J. Biomaterials for the Treatment of Myocardial Infarction. J. Am. Coll. Cardiol. 2006, 48, 907–913. [Google Scholar] [CrossRef] [Green Version]
- Rane, A.A.; Christman, K.L. Biomaterials for the Treatment of Myocardial Infarction: A 5-Year Update. J. Am. Coll. Cardiol. 2011, 58, 2615–2629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
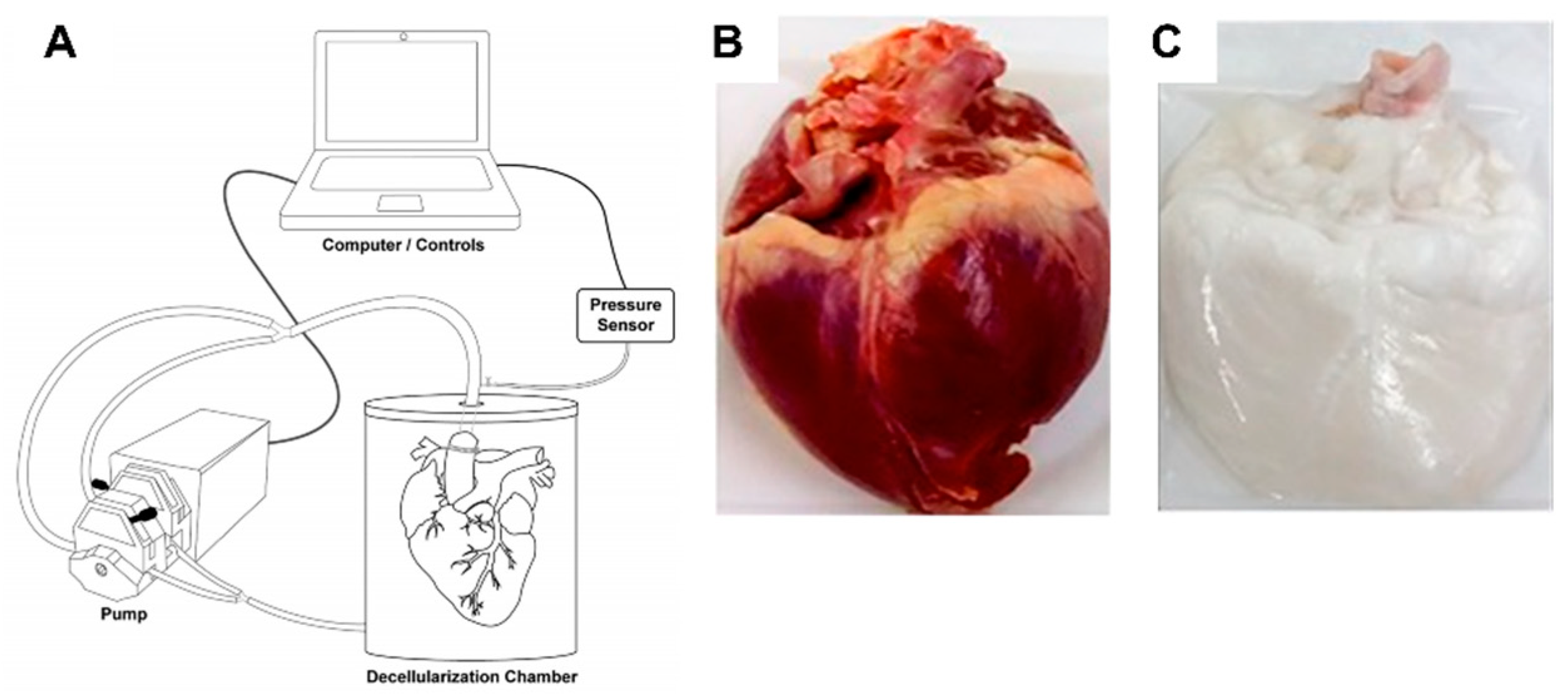
- Taylor, D.A.; Sampaio, L.C.; Ferdous, Z.; Gobin, A.S.; Taite, L.J. Decellularized matrices in regenerative medicine. Acta Biomater. 2018, 74, 74–89. [Google Scholar] [CrossRef]
- Garreta, E.; Oria, R.; Tarantino, C.; Pla-Roca, M.; Prado, P.; Fernández-Avilés, F.; Campistol, J.M.; Samitier, J.; Montserrat, N. Tissue engineering by decellularization and 3D bioprinting. Mater. Today 2017, 20, 166–178. [Google Scholar] [CrossRef] [Green Version]
- Bejleri, D.; Davis, M.E. Decellularized Extracellular Matrix Materials for Cardiac Repair and Regeneration. Adv. Heal. Mater. 2019, 8, e1801217. [Google Scholar] [CrossRef] [PubMed]
- Gaffney, L.; Wrona, E.A.; Freytes, D.O. Potential Synergistic Effects of Stem Cells and Extracellular Matrix Scaffolds. ACS Biomater. Sci. Eng. 2017, 4, 1208–1222. [Google Scholar] [CrossRef]
- Kc, P.; Hong, Y.; Zhang, G. Cardiac tissue-derived extracellular matrix scaffolds for myocardial repair: Advantages and challenges. Regen. Biomater. 2019, 6, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Khalpey, Z. Acellular porcine heart matrices: Whole organ decellularization with 3D-Bioscaffold & vascular preservation. J. Clin. Transl. Res. 2017. [Google Scholar] [CrossRef] [Green Version]
- Eitan, Y.; Sarig, U.; Dahan, N.; Machluf, M. Acellular Cardiac Extracellular Matrix as a Scaffold for Tissue Engineering: In Vitro Cell Support, Remodeling, and Biocompatibility. Tissue Eng. Part C Methods 2010, 16, 671–683. [Google Scholar] [CrossRef]
- Schoen, B.; Avrahami, R.; Baruch, L.; Efraim, Y.; Goldfracht, I.; Elul, O.; Davidov, T.; Gepstein, L.; Zussman, E.; Machluf, M. Electrospun Extracellular Matrix: Paving the Way to Tailor-Made Natural Scaffolds for Cardiac Tissue Regeneration. Adv. Funct. Mater. 2017, 27, 1700427. [Google Scholar] [CrossRef]
- Efraim, Y.; Sarig, H.; Anavy, N.C.; Sarig, U.; de Berardinis, E.; Chaw, S.-Y.; Krishnamoorthi, M.; Kalifa, J.; Bogireddi, H.; Duc, T.V.; et al. Biohybrid cardiac ECM-based hydrogels improve long term cardiac function post myocardial infarction. Acta Biomater. 2017, 50, 220–233. [Google Scholar] [CrossRef] [PubMed]
- Muniyandi, P.; Palaninathan, V.; Veeranarayanan, S.; Ukai, T.; Maekawa, T.; Hanajiri, T.; Mohamed, M.S. ECM Mimetic Electrospun Porous Poly (L-lactic acid) (PLLA) Scaffolds as Potential Substrates for Cardiac Tissue Engineering. Polymers 2020, 12, 451. [Google Scholar] [CrossRef] [Green Version]
- Seif-Naraghi, S.B.; Singelyn, J.M.; Salvatore, M.A.; Osborn, K.G.; Wang, J.J.; Sampat, U.; Kwan, O.L.; Strachan, G.M.; Wong, J.; Schup-Magoffin, P.J.; et al. Safety and Efficacy of an Injectable Extracellular Matrix Hydrogel for Treating Myocardial Infarction. Sci. Transl. Med. 2013, 5, 173ra25. [Google Scholar] [CrossRef] [Green Version]
- Alexanian, R.A.; Mahapatra, K.; Schmuck, E.G.; Raval, A.N.; Eckhardt, L.L.; Glukhov, A.V.; Lalit, P.A.; Kamp, T.J.; Lang, D.; Vaidyanathan, R.; et al. Induced cardiac progenitor cells repopulate decellularized mouse heart scaffolds and differentiate to generate cardiac tissue. Biochim. Biophys. Acta Bioenerg. 2020, 1867, 118559. [Google Scholar] [CrossRef]
- Reid, J.A.; Callanan, A. Hybrid cardiovascular sourced extracellular matrix scaffolds as possible platforms for vascular tissue engineering. J. Biomed. Mater. Res. Part B Appl. Biomater. 2019, 108, 910–924. [Google Scholar] [CrossRef] [Green Version]
- Jongpaiboonkit, L.; King, W.J.; Lyons, G.E.; Paguirigan, A.L.; Warrick, J.W.; Beebe, D.J.; Murphy, W.L. An adaptable hydrogel array format for 3-dimensional cell culture and analysis. Biomaterials 2008, 29, 3346–3356. [Google Scholar] [CrossRef] [Green Version]
- Kraehenbuehl, T.P.; Zammaretti, P.; Van Der Vlies, A.J.; Schoenmakers, R.G.; Lutolf, M.P.; Jaconi, M.E.; Hubbell, J.A. Three-dimensional extracellular matrix-directed cardioprogenitor differentiation: Systematic modulation of a synthetic cell-responsive PEG-hydrogel. Biomaterials 2008, 29, 2757–2766. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.S.T.; Yoo, H.; Yi, H.; Ahn, E.H.; Lee, J.H.; Shao, G.; Nagornyak, E.; Laflamme, M.A.; Murry, C.E.; Kim, D.-H. Micro- and nano-patterned conductive graphene–PEG hybrid scaffolds for cardiac tissue engineering. Chem. Commun. 2017, 53, 7412–7415. [Google Scholar] [CrossRef] [PubMed]
- Karahaliloğlu, Z. Electrospun PU-PEG and PU-PC hybrid scaffolds for vascular tissue engineering. Fibers Polym. 2017, 18, 2135–2145. [Google Scholar] [CrossRef]
- Cristallini, C.; Gagliardi, M.; Barbani, N.; Giannessi, D.; Guerra, G.D. Novel biodegradable, biomimetic and functionalised polymer scaffolds to prevent expansion of post-infarct left ventricular remodelling. J. Mater. Sci. Mater. Electron. 2012, 23, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Maiullari, F.; Costantini, M.; Milan, M.; Pace, V.; Chirivì, M.; Maiullari, S.; Rainer, A.; Baci, D.; Marei, H.E.S.; Seliktar, D.; et al. A multi-cellular 3D bioprinting approach for vascularized heart tissue engineering based on HUVECs and iPSC-derived cardiomyocytes. Sci. Rep. 2018, 8, 13532. [Google Scholar] [CrossRef]
- Bearzi, C.; Gargioli, C.; Baci, D.; Fortunato, O.; Shapira-Schweitzer, K.; Kossover, O.; Latronico, M.V.G.; Seliktar, D.; Condorelli, G.; Rizzi, R. PlGF–MMP9-engineered iPS cells supported on a PEG–fibrinogen hydrogel scaffold possess an enhanced capacity to repair damaged myocardium. Cell Death Dis. 2014, 5, e1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertuoli, P.T.; Ordoño, J.; Armelin, E.; Pérez-Amodio, S.; Baldissera, A.F.; Ferreira, C.A.; Puiggalí, J.; Engel, E.; Del Valle, L.J.; Alemán, C. Electrospun Conducting and Biocompatible Uniaxial and Core–Shell Fibers Having Poly(lactic acid), Poly(ethylene glycol), and Polyaniline for Cardiac Tissue Engineering. ACS Omega 2019, 4, 3660–3672. [Google Scholar] [CrossRef] [PubMed]
- Lopes, M.S.; Jardini, A.; Filho, R.M. Poly (Lactic Acid) Production for Tissue Engineering Applications. Procedia Eng. 2012, 42, 1402–1413. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Valtchev, P.; Dehghani, F. Synthesis and purification of poly(l-lactic acid) using a one step benign process. Green Chem. 2012, 14, 1357–1366. [Google Scholar] [CrossRef]
- Liu, Q.; Tian, S.; Zhao, C.; Chen, X.; Lei, I.; Wang, Z.; Ma, P.X. Porous nanofibrous poly( l -lactic acid) scaffolds supporting cardiovascular progenitor cells for cardiac tissue engineering. Acta Biomater. 2015, 26, 105–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, A.; Soliman, S.; El Hajj, F.; Tseng, Y.-T.; Yalcin, H.C.; Marei, H.E. Fabrication and In Vitro Characterization of a Tissue Engineered PCL-PLLA Heart Valve. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.S.; Blue, M.A.; Brandon, T.A.; Emmanual, J.; Guilbeau, E.J. Performance of a hy-drogel composite pericardial substitute after long-term implant studies. Asaio. J. 1992, 38, M550–M554. [Google Scholar] [CrossRef] [PubMed]
- Vijayasekaran, S.; Chirila, T.V.; Robertson, T.A.; Lou, X.; Fitton, J.H.; Hicks, C.R.; Constable, I.J. Calcification of poly(2-hydroxyethyl methacrylate) hydrogel sponges implanted in the rabbit cornea: A 3-month study. J. Biomater. Sci. Polym. Ed. 2000, 11, 599–615. [Google Scholar] [CrossRef] [PubMed]
- Madden, L.R.; Mortisen, D.J.; Sussman, E.M.; Dupras, S.K.; Fugate, J.A.; Cuy, J.L.; Hauch, K.D.; Laflamme, M.A.; Murry, C.E.; Ratner, B.D. Proangiogenic scaffolds as functional templates for cardiac tissue engineering. Proc. Natl. Acad. Sci. USA 2010, 107, 15211–15216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lao, S.; Xu, J.; Liu, Y.; Cai, S.; Lin, L.; Zhang, J.; Cai, N.; Yin, S. A comparative study of the influence of two types of PHEMA stents on the differentiation of ASCs to myocardial cells. Mol. Med. Rep. 2017, 16, 507–514. [Google Scholar] [CrossRef] [Green Version]
- Atzet, S.; Curtin, S.; Trinh, P.; Bryant, S.; Ratner, B. Degradable Poly(2-hydroxyethyl methacrylate)-co-polycaprolactone Hydrogels for Tissue Engineering Scaffolds. Biomacromolecules 2008, 9, 3370–3377. [Google Scholar] [CrossRef] [Green Version]
- Filipović, V.V.; Nedeljković, B.Đ.B.; Vukomanović, M.; Tomić, S.L. Biocompatible and degradable scaffolds based on 2-hydroxyethyl methacrylate, gelatin and poly(beta amino ester) crosslinkers. Polym. Test. 2018, 68, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Zong, X.; Bien, H.; Chung, C.-Y.; Yin, L.; Fang, D.; Hsiao, B.S.; Chu, B.; Entcheva, E. Electrospun fine-textured scaffolds for heart tissue constructs. Biomaterials 2005, 26, 5330–5338. [Google Scholar] [CrossRef] [PubMed]
- Abadi, P.P.S.S.; Garbern, J.C.; Behzadi, S.; Hill, M.J.; Tresback, J.S.; Heydari, T.; Ejtehadi, M.R.; Ahmed, N.; Copley, E.; Aghaverdi, H.; et al. Engineering of Mature Human Induced Pluripotent Stem Cell-Derived Cardiomyocytes Using Substrates with Multiscale Topography. Adv. Funct. Mater. 2018, 28. [Google Scholar] [CrossRef]
- Xu, C.; Inai, R.; Kotaki, M.; Ramakrishna, S. Aligned biodegradable nanofibrous structure: A potential scaffold for blood vessel engineering. Biomaterials 2004, 25, 877–886. [Google Scholar] [CrossRef]
- Jr, G.C.E.; Cheng, M.; Bettinger, C.J.; Borenstein, J.T.; Langer, R.; Freed, L.E. Accordion-like honeycombs for tissue engineering of cardiac anisotropy. Nat. Mater. 2008, 7, 1003–1010. [Google Scholar] [CrossRef]
- Nichol, J.W.; Engelmayr, G.C.; Cheng, M.; Freed, L.E. Co-culture induces alignment in engineered cardiac constructs via MMP-2 expression. Biochem. Biophys. Res. Commun. 2008, 373, 360–365. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Wang, J.; Loskill, P.; Huebsch, N.; Koo, S.; Svedlund, F.L.; Marks, N.C.; Hua, E.W.; Grigoropoulos, C.P.; Conklin, B.R.; et al. Self-organizing human cardiac microchambers mediated by geometric confinement. Nat. Commun. 2015, 6, 7413. [Google Scholar] [CrossRef] [Green Version]
- Hawkes, W.; Huang, D.; Reynolds, P.; Hammond, L.; Ward, M.; Gadegaard, N.; Marshall, J.F.; Iskratsch, T.; Palma, M. Probing the nanoscale organisation and multivalency of cell surface receptors: DNA origami nanoarrays for cellular studies with single-molecule control. Faraday Discuss. 2019, 219, 203–219. [Google Scholar] [CrossRef] [Green Version]
- Kankala, R.K.; Zhu, K.; Li, J.; Wang, C.-S.; Wang, S.-B.; Chen, A.-Z. Fabrication of arbitrary 3D components in cardiac surgery: From macro-, micro- to nanoscale. Biofabrication 2017, 9, 032002. [Google Scholar] [CrossRef] [Green Version]
- Martinelli, V.; Cellot, G.; Fabbro, A.; Bosi, S.; Mestroni, L.; Ballerini, L. Improving cardiac myocytes performance by carbon nanotubes platforms. Front. Physiol. 2013, 4, 239. [Google Scholar] [CrossRef] [Green Version]
- Feinberg, A.W.; Alford, P.W.; Jin, H.; Ripplinger, C.M.; Werdich, A.A.; Sheehy, S.P.; Grosberg, A.; Parker, K.K. Controlling the contractile strength of engineered cardiac muscle by hierarchal tissue architecture. Biomaterials 2012, 33, 5732–5741. [Google Scholar] [CrossRef] [Green Version]
- Pong, T.; Adams, W.J.; Bray, M.-A.; Feinberg, A.W.; Sheehy, S.P.; A Werdich, A.; Parker, K.K. Hierarchical architecture influences calcium dynamics in engineered cardiac muscle. Exp. Biol. Med. 2011, 236, 366–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaetani, R.; Doevendans, P.A.; Metz, C.H.; Alblas, J.; Messina, E.; Giacomello, A.; Sluijter, J.P. Cardiac tissue engineering using tissue printing technology and human cardiac progenitor cells. Biomaterials 2012, 33, 1782–1790. [Google Scholar] [CrossRef] [PubMed]
- Chan, V.; Raman, R.; Cvetkovic, C.; Bashir, R. Enabling Microscale and Nanoscale Approaches for Bioengineered Cardiac Tissue. ACS Nano 2013, 7, 1830–1837. [Google Scholar] [CrossRef] [PubMed]
- Yeo, Y.; Geng, W.; Ito, T.; Kohane, D.S.; Burdick, J.A.; Radisic, M. Photocrosslinkable hydrogel for myocyte cell culture and injection. J. Biomed. Mater. Res. Part B Appl. Biomater. 2006, 81, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Forte, G.; Carotenuto, F.; Pagliari, F.; Pagliari, S.; Cossa, P.; Fiaccavento, R.; Ahluwalia, A.; Vozzi, G.; Vinci, B.; Serafino, A.; et al. Criticality of the Biological and Physical Stimuli Array Inducing Resident Cardiac Stem Cell Determination. Stem Cells 2008, 26, 2093–2103. [Google Scholar] [CrossRef] [PubMed]
- Rehfeldt, F.; Engler, A.J.; Eckhardt, A.; Ahmed, F.; Discher, D.E. Cell responses to the mechanochemical microenvironment—Implications for regenerative medicine and drug delivery. Adv. Drug Deliv. Rev. 2007, 59, 1329–1339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix Elasticity Directs Stem Cell Lineage Specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef] [Green Version]
- Luna, J.I.; Ciriza, J.; García-Ojeda, M.E.; Kong, M.; Herren, A.; Lieu, D.K.; Li, R.A.; Fowlkes, C.C.; Khine, M.; McCloskey, K.E. Multiscale Biomimetic Topography for the Alignment of Neonatal and Embryonic Stem Cell-Derived Heart Cells. Tissue Eng. Part C Methods 2011, 17, 579–588. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Wang, Z.; Hu, X.; Chan, J.K.Y.; Wen, F.; Wang, Y.; Xie, C.; Wang, X.; Luo, F.; Hong, M.; et al. Scaffold with Micro/Macro-Architecture for Myocardial Alignment Engineering into Complex 3D Cell Patterns. Adv. Health Mater. 2019, 8, e1901015. [Google Scholar] [CrossRef]
- Hinman, S.S.; Kim, R.; Wang, Y.; Phillips, K.S.; Attayek, P.J.; Allbritton, N.L. Microphysiological system design: Simplicity is elegance. Curr. Opin. Biomed. Eng. 2020, 13, 94–102. [Google Scholar] [CrossRef]
- Kolanowski, T.J.; Busek, M.; Schubert, M.; Dmitrieva, A.; Binnewerg, B.; Pöche, J.; Fisher, K.; Schmieder, F.; Grünzner, S.; Hansen, S.; et al. Enhanced structural maturation of human induced pluripotent stem cell-derived cardiomyocytes under a controlled microenvironment in a microfluidic system. Acta Biomater. 2020, 102, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Granados-Riveron, J.T.; Brook, J.D. The Impact of Mechanical Forces in Heart Morphogenesis. Circ. Cardiovasc. Genet. 2012, 5, 132–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lux, M.; Andrée, B.; Horvath, T.; Nosko, A.; Manikowski, D.; Hilfiker-Kleiner, D.; Haverich, A.; Hilfiker, A. In vitro maturation of large-scale cardiac patches based on a perfusable starter matrix by cyclic mechanical stimulation. Acta Biomater. 2016, 30, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Shen, N.; Knopf, A.; Westendorf, C.; Kraushaar, U.; Riedl, J.; Bauer, H.; Pöschel, S.; Layland, S.L.; Holeiter, M.; Knolle, S.; et al. Steps toward Maturation of Embryonic Stem Cell-Derived Cardiomyocytes by Defined Physical Signals. Stem Cell Rep. 2017, 9, 122–135. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.; Ma, X.; Gou, M.; Mei, D.; Zhang, K.; Chen, S. 3D printing of functional biomaterials for tissue engineering. Curr. Opin. Biotechnol. 2016, 40, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Duan, B. State-of-the-Art Review of 3D Bioprinting for Cardiovascular Tissue Engineering. Ann. Biomed. Eng. 2017, 45, 195–209. [Google Scholar] [CrossRef]
- Shin, Y.J.; Shafranek, R.T.; Tsui, J.H.; Walcott, J.; Nelson, A.; Kim, D.-H. 3D bioprinting of mechanically tuned bioinks derived from cardiac decellularized extracellular matrix. Acta Biomater. 2021, 119, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Stoppel, W.L.; Hu, D.; Domian, I.J.; Kaplan, D.L.; Black, L.D. Anisotropic silk biomaterials containing cardiac extracellular matrix for cardiac tissue engineering. Biomed. Mater. 2015, 10, 034105. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.; Hudson, A.R.; Shiwarski, D.J.; Tashman, J.W.; Hinton, T.J.; Yerneni, S.; Bliley, J.M.; Campbell, P.G.; Feinberg, A.W. 3D bioprinting of collagen to rebuild components of the human heart. Science 2019, 365, 482–487. [Google Scholar] [CrossRef] [PubMed]
- MacQueen, L.A.; Sheehy, S.P.; Chantre, C.O.; Zimmerman, J.F.; Pasqualini, F.S.; Liu, X.; Goss, J.A.; Campbell, P.H.; Gonzalez, G.M.; Park, S.-J.; et al. A tissue-engineered scale model of the heart ventricle. Nat. Biomed. Eng. 2018, 2, 930–941. [Google Scholar] [CrossRef]
- Kupfer, M.E.; Lin, W.H.; Ravikumar, V.; Qiu, K.; Wang, L.; Gao, L.; Bhuiyan, D.; Lenz, M.; Ai, J.; Mahutga, R.R.; et al. In Situ Expansion, Differentiation and Electromechanical Coupling of Human Cardiac Muscle in a 3D Bioprinted, Chambered Organoid. Circ. Res. 2020, 127, 207–224. [Google Scholar] [CrossRef]
- Noble, D. A modification of the Hodgkin-Huxley equations applicable to Purkinje fibre action and pacemaker potentials. J. Physiol. 1962, 160, 317–352. [Google Scholar] [CrossRef]
- Puglisi, J.L.; Wang, F.; Bers, D.M. Modeling the isolated cardiac myocyte. Prog. Biophys. Mol. Biol. 2004, 85, 163–178. [Google Scholar] [CrossRef] [PubMed]
- Wolska, B.M.; Solaro, R.J. Method for isolation of adult mouse cardiac myocytes for studies of contraction and microfluorimetry. Am. J. Physiol. Circ. Physiol. 1996, 271, H1250–H1255. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.H.; Rudy, Y. A model of the ventricular cardiac action potential. Depolarization, repolarization, and their interaction. Circ. Res. 1991, 68, 1501–1526. [Google Scholar] [CrossRef] [Green Version]
- O’Hara, T.; Virág, L.; Varró, A.; Rudy, Y. Simulation of the Undiseased Human Cardiac Ventricular Action Potential: Model Formulation and Experimental Validation. PLoS Comput. Biol. 2011, 7, e1002061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heijman, J.; Volders, P.G.; Westra, R.L.; Rudy, Y. Local control of β-adrenergic stimulation: Effects on ventricular myocyte electrophysiology and Ca2+-transient. J. Mol. Cell. Cardiol. 2011, 50, 863–871. [Google Scholar] [CrossRef] [Green Version]
- Lascano, E.C.; Said, M.; Vittone, L.; Mattiazzi, A.; Mundiña-Weilenmann, C.; Negroni, J.A. Role of CaMKII in post acidosis arrhythmias: A simulation study using a human myocyte model. J. Mol. Cell. Cardiol. 2013, 60, 172–183. [Google Scholar] [CrossRef]
- Bai, J.; Wang, K.; Liu, Y.; Li, Y.; Liang, C.; Luo, G.; Dong, S.; Yuan, Y.; Zhang, H. Computational Cardiac Modeling Reveals Mechanisms of Ventricular Arrhythmogenesis in Long QT Syndrome Type 8: CACNA1C R858H Mutation Linked to Ventricular Fibrillation. Front. Physiol. 2017, 8, 771. [Google Scholar] [CrossRef]
- Fernandez-Chas, M.; Curtis, M.J.; A Niederer, S. Mechanism of doxorubicin cardiotoxicity evaluated by integrating multiple molecular effects into a biophysical model. Br. J. Pharmacol. 2018, 175, 763–781. [Google Scholar] [CrossRef] [Green Version]
- Moreno, J.D.; Zhu, Z.I.; Yang, P.-C.; Bankston, J.R.; Jeng, M.-T.; Kang, C.; Wang, L.; Bayer, J.D.; Christini, D.J.; Trayanova, N.A.; et al. A Computational Model to Predict the Effects of Class I Anti-Arrhythmic Drugs on Ventricular Rhythms. Sci. Transl. Med. 2011, 3, 98ra83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Perez, A.; Sebastian, R.; Ferrero, J.M. Three-dimensional cardiac computational modelling: Methods, features and applications. Biomed. Eng. Online 2015, 14, 1–31. [Google Scholar] [CrossRef] [Green Version]
- Baillargeon, B.; Rebelo, N.; Fox, D.D.; Taylor, R.L.; Kuhl, E. The Living Heart Project: A robust and integrative simulator for human heart function. Eur. J. Mech. A/Solids 2014, 48, 38–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plank, G.; Burton, R.A.B.; Mahmood, F.; Rodriguez, B.; Grau, V.; Schneider, J.E.; Gavaghan, D.; Kohl, P.; Hales, P.; Bishop, M.; et al. Generation of histo-anatomically representative models of the individual heart: Tools and application. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2009, 367, 2257–2292. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Constantino, J.; Vedula, V.; Trayanova, N.; Mittal, R. A New MRI-Based Model of Heart Function with Coupled Hemodynamics and Application to Normal and Diseased Canine Left Ventricles. Front. Bioeng. Biotechnol. 2015, 3, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mincholé, A.; Zacur, E.; Ariga, R.; Grau, V.; Rodriguez, B. MRI-based computational tor-so/biventricular multiscale models to investigate the impact of anatomical variability on the ECG QRS complex. Front. Physiol. 2019, 10, 1103. [Google Scholar] [CrossRef] [Green Version]
- Trayanova, N.A. Whole-Heart Modeling. Circ. Res. 2011, 108, 113–128. [Google Scholar] [CrossRef] [Green Version]
- Mittal, R.; Seo, J.H.; Vedula, V.; Choi, Y.J.; Liu, H.; Huang, H.H.; Jain, S.; Younes, L.; Abraham, T.; George, R.T. Computational modeling of cardiac hemodynamics: Current status and future outlook. J. Comput. Phys. 2016, 305, 1065–1082. [Google Scholar] [CrossRef]
- McCormick, M.; Nordsletten, D.A.; Kay, D.; Smith, N.P. Simulating left ventricular fluid–solid me-chanics through the cardiac cycle under LVAD support. J. Comput. Phys. 2013, 244, 80–96. [Google Scholar] [CrossRef]
- Taylor, C.A.; Fonte, T.A.; Min, J.K. Computational Fluid Dynamics Applied to Cardiac Computed Tomography for Noninvasive Quantification of Fractional Flow Reserve. J. Am. Coll. Cardiol. 2013, 61, 2233–2241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz, D.R.; Markl, M.; Mur, J.L.M.; Barker, A.; Fernández-Golfín, C.; Lancellotti, P.; Gómez, J.L.Z. Intracardiac flow visualization: Current status and future directions. Eur. Hear. J. Cardiovasc. Imaging 2013, 14, 1029–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augustin, C.M.; Crozier, A.; Neic, A.; Prassl, A.J.; Karabelas, E.; Da Silva, T.F.; Fernandes, J.F.; Campos, F.; Kuehne, T.; Plank, G. Patient-specific modeling of left ventricular electromechanics as a driver for haemodynamic analysis. Europace 2016, 18, iv121–iv129. [Google Scholar] [CrossRef] [PubMed]






















| Cell Source (Type) | Advantages | Disadvantages |
|---|---|---|
| Adult rat ventricular myocyte (ARVM) | Easy to obtain Mature | Xenogenic |
| Neonatal rat ventricular myocyte (NRVM) | Easy to obtain | Xenogenic Immature |
| Adult porcine ventricular myocyte | Genetically closer to human cells than rodent | Xenogenic |
| Human embryonic stem cell-derived cardiomyocyte (hESC-CM) | Same species Unlimited cell source | Ethical issue Allogenic Immature |
| Human-induced pluripotent stem cell-derived cardiomyocyte (hiPSC-CM) | Same species Unlimited cell source Autologous | Immature |
| Name | Advantages | Disadvantages | |
|---|---|---|---|
| Natural materials | Collagen; decellularized ECM | Biodegradable and biocompatible | Slow gelation, weak strength |
| Fibrin gel | Biodegradable and biocompatible | Fast degradation in vivo and slow gelation | |
| Matrigel | Similar to native ECM structure | Potentially carcinogenic | |
| Chitosan | Structurally similar to heart tissue components | Insufficient support for cardiac tissue growth | |
| Alginate | Biodegradable, biocompatible, non-toxic, and cost effective | Weak cell adhesion and proliferation | |
| Synthetic materials | PHEMA | Biocompatible | Non-degradable and mismatch of modulus |
| PEG | Biocompatible, bioinert, and FDA approved | Non-degradable, low cell adhesion, and toxicity | |
| Polyamides | Versatility in chemical modification, fast gelation time, and injectability | Non elastic and non-degradable |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, A.R.M.P.; Jang, Y.; Son, I.; Kim, J.; Park, Y. Recapitulating Cardiac Structure and Function In Vitro from Simple to Complex Engineering. Micromachines 2021, 12, 386. https://doi.org/10.3390/mi12040386
Santos ARMP, Jang Y, Son I, Kim J, Park Y. Recapitulating Cardiac Structure and Function In Vitro from Simple to Complex Engineering. Micromachines. 2021; 12(4):386. https://doi.org/10.3390/mi12040386
Chicago/Turabian StyleSantos, Ana Rita M. P., Yongjun Jang, Inwoo Son, Jongseong Kim, and Yongdoo Park. 2021. "Recapitulating Cardiac Structure and Function In Vitro from Simple to Complex Engineering" Micromachines 12, no. 4: 386. https://doi.org/10.3390/mi12040386
APA StyleSantos, A. R. M. P., Jang, Y., Son, I., Kim, J., & Park, Y. (2021). Recapitulating Cardiac Structure and Function In Vitro from Simple to Complex Engineering. Micromachines, 12(4), 386. https://doi.org/10.3390/mi12040386