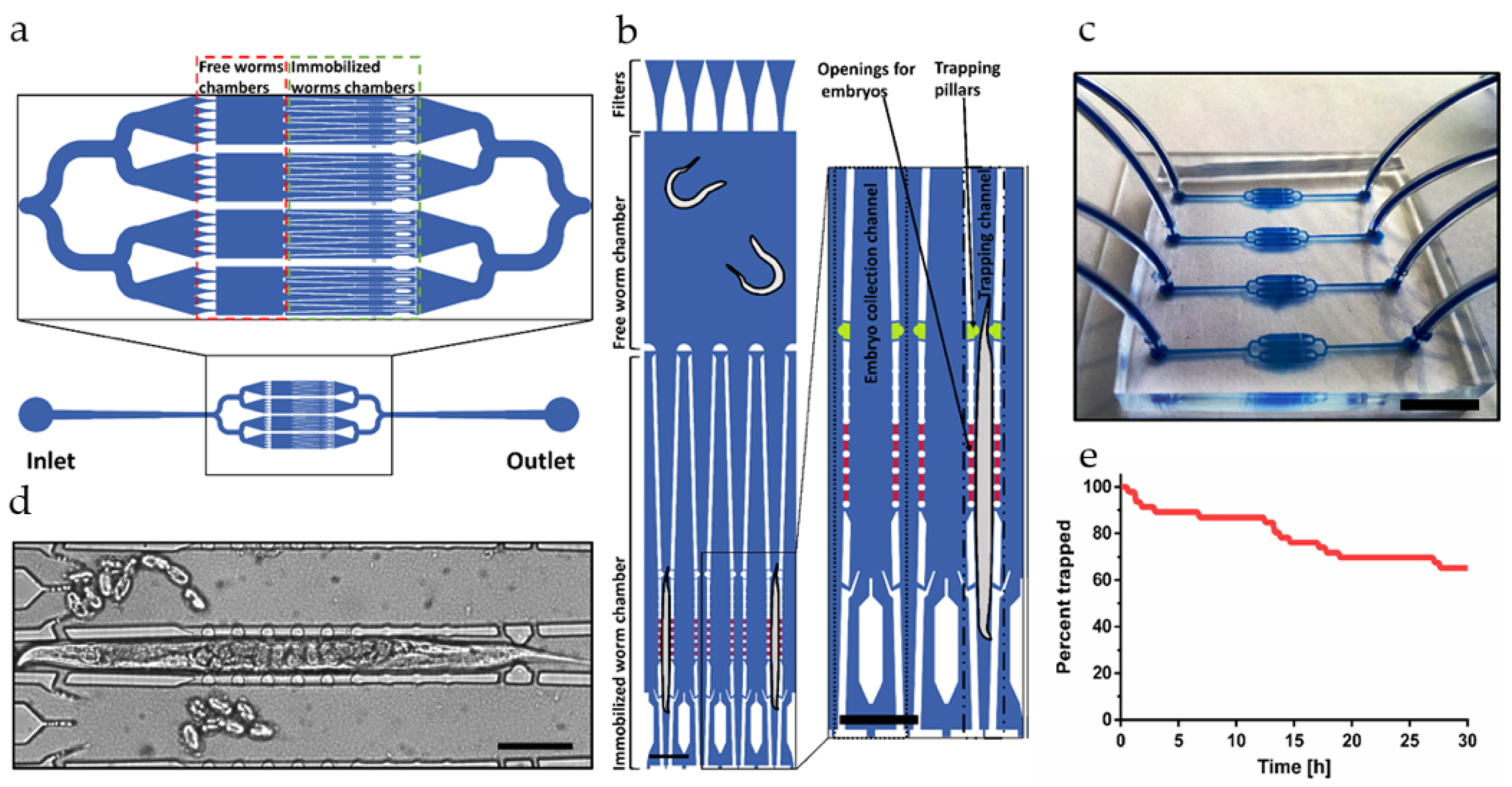
4.1. Monitoring of the Bacterial Load in the C. elegans Intestine
Worms mechanically disrupt ingested bacterial food in the pharyngeal grinder. Subsequently, peristaltic action transports bacterial fragments and nutrients into the mid and posterior regions of the gastrointestinal tract for chemical metabolization during the digestive process and finally residues are expelled by activation of a defecation motor program [
47]. Expulsion of undigested material takes place approximately every 45–50 s for N2 worms [
48]. We have used low-magnification (20×) imaging for monitoring the bacterial load of the entire gut of immobilized adult N2 worms. A representative example illustrating our custom-developed image analysis protocol is shown in
Figure 2a [
49]. In this specific case of a worm fed
E. coli OP50 RFP (adult N2 worm, trapped for 30 h, see
Figure 3a), the fluorescent image revealed enhanced bacterial load in the terminal bulb (grinder) of the pharynx, which is separated from the intestine by the pharyngeal-intestine valve, and in the posterior region of the intestine (hindgut) (
Figure 2a, upper image) [
50].
For quantification of bacterial accumulation, the fluorescent signal was measured along a manually defined line following the intestinal profile (
Figure 2a, lower image). Mean values were calculated over a width of 25 µm and represented graphically by plotting the fluorescence intensity as a function of the location in the worm gut (
Figure 2b).
The graph was then transposed into a linear color code scheme for more convenient visualization of the fluorescence distribution (panel in the upper part of
Figure 2b). For monitoring the dynamics of bacterial/microbead transit or accumulation, an adequate number of such color plots was recorded successively over the time interval of interest and grouped into larger panels.
Figure 2c shows a high-resolution brightfield/fluorescent image (50×) of the hindgut of an adult worm, allowing to locate precisely the bacterial load (
E. coli OP50 RFP). As shown in this example, we generally observed a superposition of diffuse fluorescence distributed in the lumen and brighter spots of small and varying size. As bacteria are mechanically disrupted in situ in the worm’s grinder, we assume that the diffuse signal may be attributed mainly to the presence of active fluorophores originating from membrane fragments and compounds from the cytoplasm [
51], i.e., nutritive compounds that can be biochemically transformed in the worm intestine. In the present context, quantification of the diffuse fluorescent signal is therefore important for evaluating the digestive process. On the other hand, depending on the efficacy of the grinding process, also a certain amount of intact single bacteria and bacterial clusters (identified as the brighter spots in
Figure 2c) can be found in locations posterior to the grinder.
In order to study the specific features and the fate of the bacterial load in more detail, we therefore designed an imaging protocol for discriminating between diffuse and discrete fluorescent signals in the intestinal track. We discriminated in vitro intact and disrupted fluorescently labeled bacteria by a simple brightness level thresholding protocol (
Figure S4). For bacteria detection in worms’ gut, two subsequent thresholding steps were applied for extracting the total fluorescent signal intensity
It (
Figure 2d) and the brighter fluorescent spots
Ii corresponding to intact bacteria that persist in the intestine (
Figure 2e). The signal
Id associated with diffuse fluorescence of disrupted bacteria was obtained by subtraction
Id =
It −
Ii (
Figure 2f). As will be reported later (see Figure 6e,f), this image processing protocol was particularly useful for evaluating the time constant
τ of biochemical digestion based on the decay of
Id.
The acidic environment in the
C. elegans intestine, in particular in the mid and posterior sections, may alter signals generated by the fluorescently labeled bacterial load [
15]. For this reason, and in particular for a correct and detailed interpretation of the presence, specific features and time dependence of the bacterial load, the pH sensitivity of the fluorophore expressed by the bacteria has to be considered. In a previous work, we characterized in vitro the pH sensitivity of two different
E. coli strains, OP50 RFP and HT115 GFP, expressing RFP or GFP fluorescent proteins, respectively. We showed that the RFP marker was not affected in the pH range between 3.7 and 6, while the GFP intensity rapidly decreased even in mildly acidic solutions (pH 6) [
49]. Here we studied the fate of these two
E. coli strains in vivo. YA N2 nematodes were loaded in the worm microfluidic traps and identical feeding protocols were applied, either using
E. coli OP50 RFP or HT115 GFP in different assay units of the chip. An example for each condition is shown in
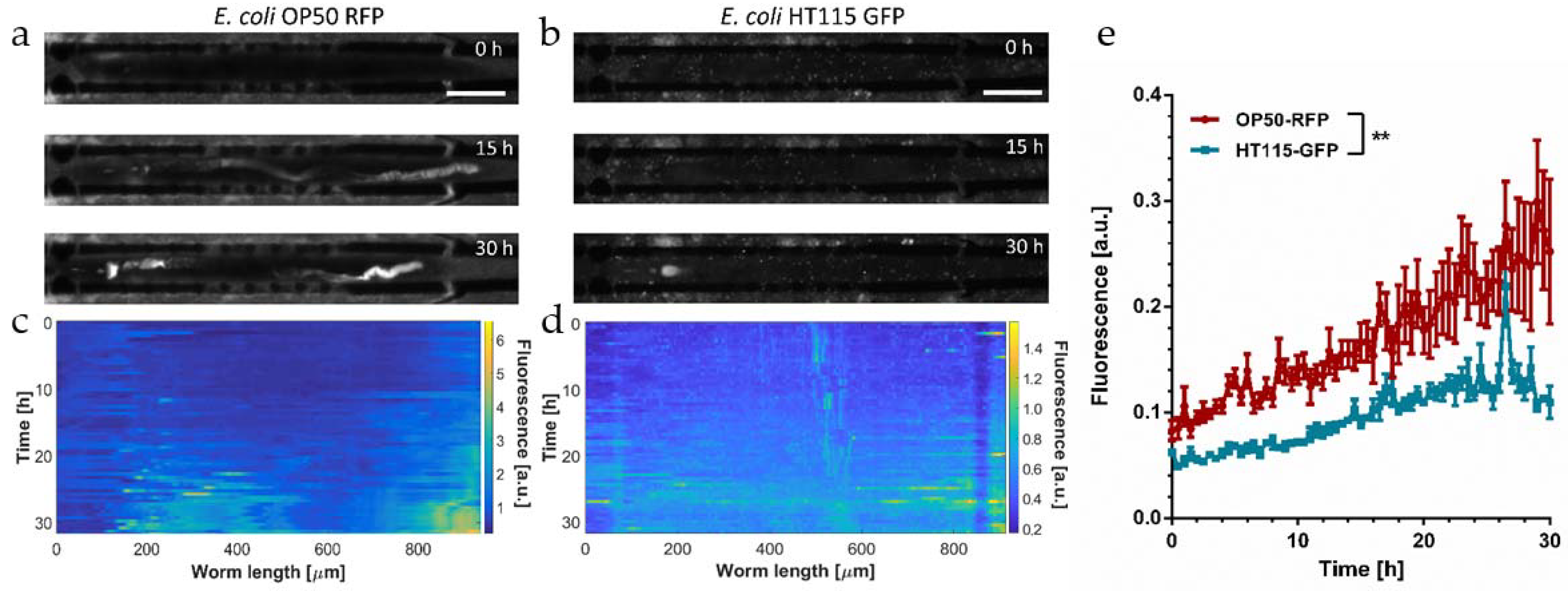
Figure 3a,b, respectively.
Representative fluorescence images (20×) of an immobilized N2 worm recorded at 0 h, 15 h, and 30 h characterized the long-term evolution of the bacterial load and/or the evolution of the fluorescence signal due to the pH sensitivity of the fluorophore. At the beginning of the observation period (YA worm at
t = 0 in
Figure 3a,b), signals were very low along the whole intestinal lumen in both cases.
During the YA stage, worms are capable to disrupt and digest ingested bacteria efficiently without significant transient or permanent accumulation. Initial exposure conditions for RFP and GFP recording have been adjusted to the limit of detection of the bacterial load in the pharynx in either cases; thus, signal intensities of RFP and GFP raw images may be qualitatively compared. The pharyngeal pumping rate of adult
C. elegans does not significantly differ for worms fed OP50 or HT115, respectively [
22]. Thus, we assume that the amount of bacteria in a completely filled pharyngeal terminal bulb is comparable for both cases. During aging (15 h to 30 h in
Figure 3a,b) a progressive increase in fluorescence was observed in the grinder and the hindgut regions for both cases, whereas the signal remained low in the mid-section of the intestine. The increase in fluorescence in the pharyngeal region may possibly be explained by the onset of a decreasing efficacy of the grinder function after 30 h. In fact, it was shown by analysis of incubated worm lysates that intestinal colonization levels, even for non-pathogenic
E. coli, already increase at the early adulthood stage [
21]. Moreover, pharyngeal swelling by bacterial infection has been identified as a reason for early death in
C. elegans [
52]. Furthermore, the hindgut region is particularly sensitive to bacterial proliferation fostered by senescent pathologies or genetic causes [
21,
28]. As will be discussed below, the strong fluorescent signal visible in the posterior region of the intestine of aging nematodes fed the OP50 RFP strain is generated by a mixture of fluorescent reporters originating from disrupted bacterial food and intact bacteria. Signals from both sources may persist in the hindgut due to the pH insensitivity of RFP. Color plot panels of the gut fluorescence corresponding to
Figure 3a,b are shown in
Figure 3c,d, respectively. These panels monitor the time evolution of the bacterial load more accurately with a time-lapse resolution of 30 min over 30 h. Overall, the signal intensity in nematodes fed OP50 RFP is considerably higher with respect to the HT115 GFP counterpart. Nevertheless, in both cases an onset of bacterial accumulation can be identified for
t ≥ 20 h (
Figure 3c,d). As the worm feeding behavior is not visibly impacted by the two
E. coli strains, we conclude that the low GFP fluorescence in the hindgut is due to the pH sensitivity of this marker. In fact, only the diffuse GFP signal component related to membrane disruption is affected by the acidic environment (see discussion of
Figure 4a); thus, we assume that the weak signal observed in this case is generated by a small amount of intact
E. coli HT115 GFP persisting in the worm gut. The graphs shown in
Figure 3e compare the evolution of the apparent average bacterial load over 30 h in worms fed
E. coli OP50 RFP and HT115 GFP, respectively. The bacterial load increased over time in both cases, in particular for worms fed the OP50 RFP strain, reflecting mainly the strong increase in the hindgut signal. Overall, GFP values are below RFP values due to the pH sensitivity of the former. In
Figure 3e, values have been normalized with respect to the maximum transient RFP or GFP fluorescent signal recorded in the pharynx, whereby we assume that these signals correspond to a comparable maximum amount of bacterial intake; therefore, both graphs in
Figure 3e may be compared in a quasi-quantitative way.
We also recorded the bacterial load evolution in free-moving worms in the dedicated chambers on the chip.
Figure S1b,c compares both conditions for the two
E. coli strains and shows that worm immobilization did not affect food uptake behavior and bacterial load accumulation in the gut, indicating physiological conditions during immobilization. In order to verify whether the observed fluorescent signal in the gut reflects accurately the amount of ingested, disrupted, and digested bacteria, we investigated the impact of pH in more detail. In particular, as discussed above, for the pH-sensitive GFP reporter, the fluorescent signal may fade in the acidic worm gut, possibly falsely indicating a signal loss caused by chemical digestion. Here, we focus on single representative worms. Results obtained on larger populations showing bacterial distribution patterns in wild-type worms and
eat-2 mutants were presented in our previous paper [
49].
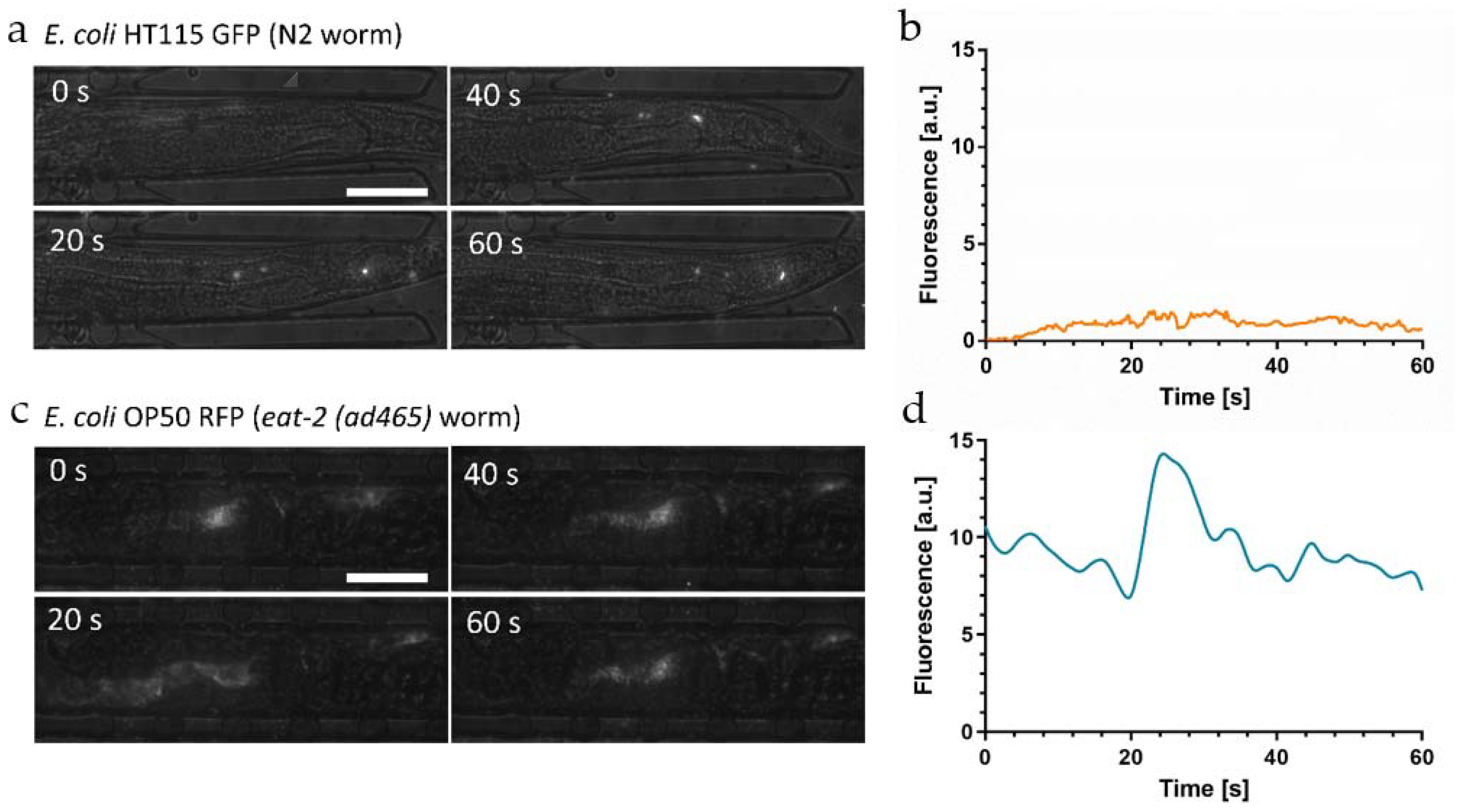
Figure 4a shows a time-lapse sequence of high-resolution images (50×) of the hindgut of an immobilized YA N2 worm fed
E. coli HT115 GFP.
In this image sequence, we could clearly resolve fluorescent spots generated by a few individual intact bacteria passing through the hindgut region, despite the high intestinal acidity in this region (pH~3.6). GFP fluorescence of intact bacteria thus appears not to be affected by the acidic environment. This observation indicated that the integrity of the bacterial membrane plays a significant role with respect to fluorescence emitted by the bacterial load during the ingestion and digestion process. On the other hand, no diffuse fluorescence associated to the presence of membrane fragments or other compounds originating from disrupted bacteria was observed in this case. GFP labeled bacteria thus turn out to be not suitable for monitoring digestion processes in the worm gut, as biochemical decomposition/nutrient absorption and fading of GFP signals due to low pH cannot be discriminated. The average fluorescent signal of the whole worm as a function of time is shown in
Figure 4b, reflecting a low but relatively stable signal originating from intact bacteria persisting in the gut.
According to the findings presented above, digestion assays require worm feeding with the pH-insensitive OP50 RFP strain. In this case, we are able to observe strong fluorescent signals in the hindgut, caused by the presence of a mixture of disrupted bacteria and intact bacteria (
Figure 3a). In order to confirm this hypothesis, we analyzed the bacterial load in
eat-2 mutants that are characterized by defective pharyngeal pumping, resulting in a conspicuous accumulation of live bacteria in the posterior region of the intestine [
53].
Figure 4c shows a time-lapse sequence of high-resolution images (50×) of the hindgut of an immobilized
eat-2 mutant fed
E. coli OP50 RFP.
We detected a mixed load of bacterial fragments and a high amount of intact bacteria generating superposed diffuse and clustered spot-like signals. The corresponding time evolution of the average fluorescent signal in the whole worm is shown in
Figure 4d.
Overall fluorescence remains on a constant high level on a time scale relevant for food processing cycles due to the stable presence of a considerable number of intact bacteria in the gut of the
eat-2 mutant. The peak in
Figure 4d is generated by a new food processing event, i.e., ingestion and subsequent digestion of a new bacterial aliquot.
4.2. Intestinal Load Dynamics and Biochemical Digestion of Bacterial Food
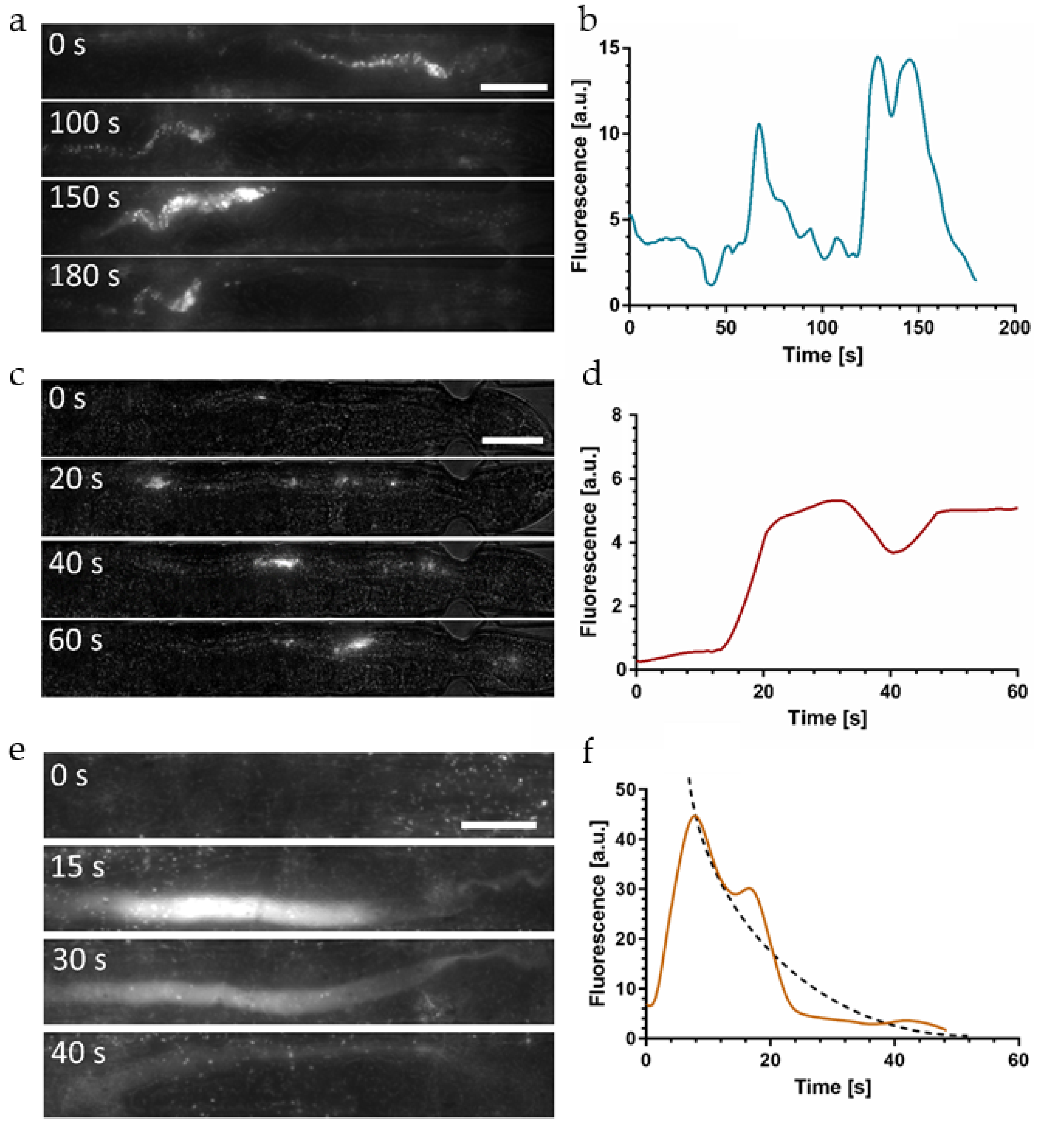
Based on the imaging protocols and preliminary studies presented above, we were able to evaluate the time constant of nutrient absorption in the worm intestine. We designed an assay for observing simultaneously the fate of
E. coli OP50 RFP fluorescence due to enzymatic digestion and the intestinal peristaltic action. For the latter, indigestible 1 µm-sized rhodamine B-marked melamine microbeads were used for tracking the dynamics and cyclic motion of the intestinal load. In order to obtain real-time recordings of the digestive processes, we performed short time-lapse imaging sequences (60 s to 180 s, frame interval of 250 ms) of confined YA N2
C. elegans worms, either at low resolution (20×) for a global view of the entire worm intestine (
Figure 5), or at high resolution (50×) for local and more precise observations, and in particular for the measurement of the decay of the diffuse fluorescent signal in the hindgut (
Figure 6). After worm loading and trapping, on-chip microfluidic units were filled either with (i) a mixture of
E. coli OP50 RFP and red fluorescent microparticles (~4 × 10
9 beads/mL in S-medium) in equal proportions, (ii) fluorescent microparticles only, or (iii)
E. coli OP50 RFP only, respectively. The chip was refilled with fresh suspension every 10 min in order to maintain the amount of bacteria or particles in the microfluidic chambers.
Representative low-resolution fluorescence images for all three conditions are shown in
Figure 5a–c, respectively.
Figure 5d–f are the corresponding color plots. When mixed, intact bacteria and microbeads cannot be distinguished in the worm intestine. In this case, the recorded signal sequences also show extended diffuse regions originating from bacterial membrane fragments. Fluorescent spots and diffuse signals are superposed, resulting in relatively broad features in the intestine (
Figure 5a,d). One the other hand, in the case of mere microbeads ingestion, spatially constrained features with stable fluorescence intensity were observed and bright spots generated by single beads and bead clusters can be more clearly identified in the absence of a diffuse signal (
Figure 5b,e). In both cases, resin microbeads cannot be digested and are periodically displaced by peristaltic contractions from the pharyngeal region to the hindgut and vice versa (
Figure 5d,e, see also
Figure 6b). Color panels similar to
Figure 5d were used to evaluate the periodicity of the intestinal load motion, and for that, the time evolution of the signal average over the hindgut region was plotted. Four representative graphs are reported in
Figure S5, from which we determined an average period of
Tp = 71 ± 5 s. In
Supplementary Video S1 multiple peristaltic contractions in the posterior intestinal lumen of a single N2 YA
C. elegans worm were recorded. Such periodical muscular contractions generate the observed cyclic motion of the intestinal load through the entire worm gut. On the contrary, when worms are fed only
E. coli OP50 RFP bacteria, the transient pattern of the observed fluorescent signals is very different and determined by the rapid biochemical digestion of bacterial food. This dynamic and kinetic behavior is exemplified in the image sequence shown in
Figure 5c and with higher time resolution in the color plot panel of
Figure 5f. Ingested bacteria, initially intact and concentrated in the worm pharynx, are physically disrupted by mechanical action of the grinder and subsequently, after passing the pharyngeal-intestinal valve, pushed towards the posterior region of the gastrointestinal tract until the pharyngeal lumen is empty. Strongest bacterial transfer from the pharynx to the adjacent intestinal lumen is visible in the time span between 10 s and 30 s on the color plot panel of
Figure 5f. This event was also captured in the image at 20 s in
Figure 5c.
Subsequently, a rapid decay of the signal corresponding to the transferred amount of bacteria was observed. Fluorescent signal fading can be mainly identified in the gut section extending from the midgut to the rectum, where already 45 s after a food ingestion event and the onset of peristalsis (
t = 0 in the image sequence), the intestine is almost free of fluorescence (
Figure 5c,f for
t ≥ 45 s). The bacteria-associated fluorescent signal becomes negligible after 60 s. Because of the insensitivity of the RFP marker to the relatively strong acidic pH in the mid and posterior region of the intestine, the observed signal decay has to be attributed to the process of nutrient metabolization and absorption, i.e., the activity of digestive enzymes that decompose fluorescent bacterial membrane fragments and nutritive compounds released from the cytoplasm. Defecation did not take place on the time scale of this recording. For these assays, the examined worms were in the YA stage where notable fluorophore/bacterial accumulation in the hindgut did not occur (as was observed for somewhat older worms in
Figure 3a,c). As mentioned above, diffusive fluorescent features can also be identified for the mixed intestinal load monitored in
Figure 5d. In principle, it should also be possible to detect a decay of fluorescence in this color plot panel. However, relatively strong bead signals impede to a certain extent accurate observation of weaker signal variations due to digestion. Furthermore, recording was carried out over a longer time scale in this case, involving successive ingestion events that replenish the bacterial load in the intestine and cyclic peristaltic motion, eventually screening somewhat a digestive decay.
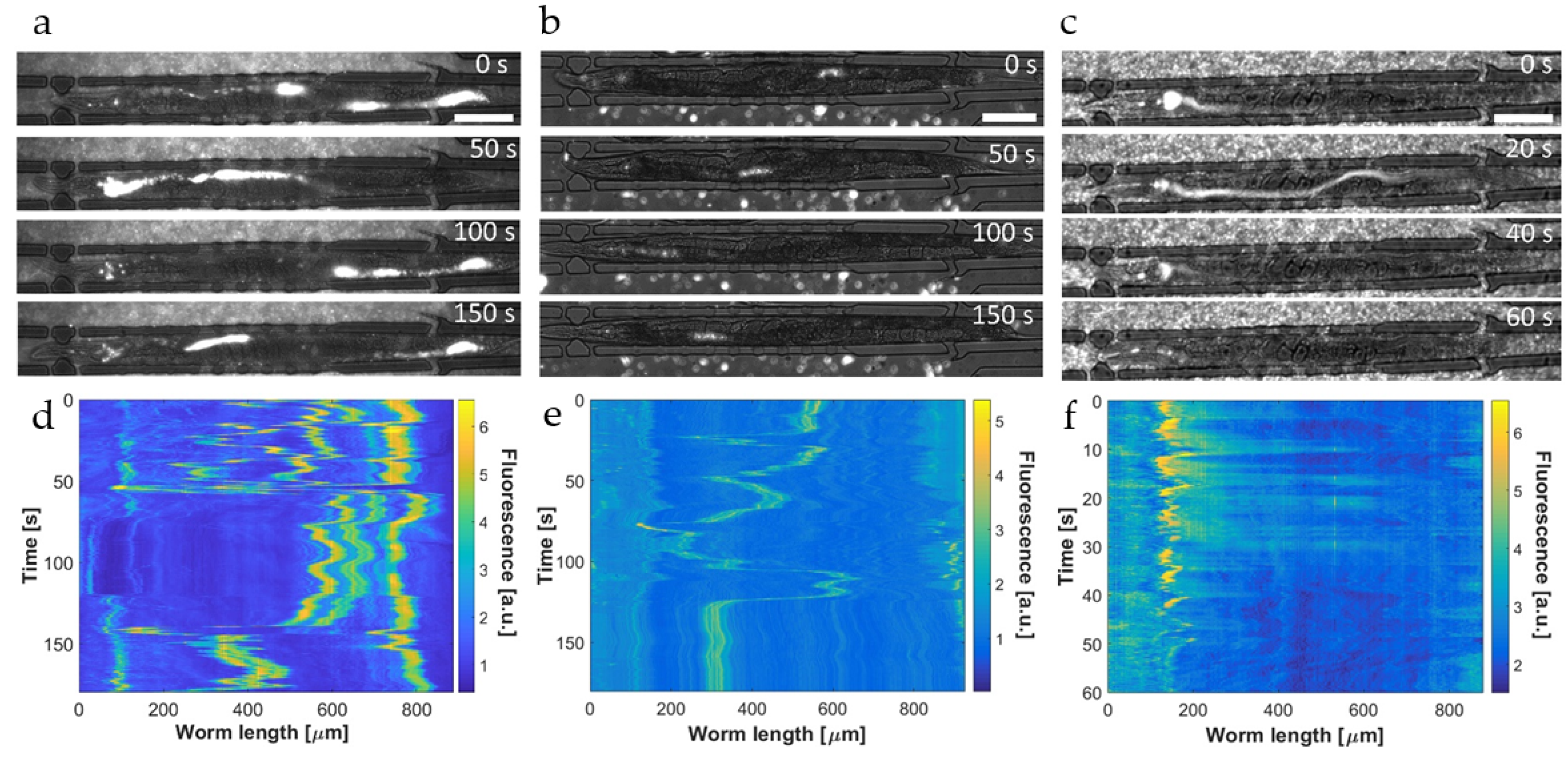
A more detailed investigation of the bacterial transport and digestion processes addressed in
Figure 5 is presented in
Figure 6 based on high-resolution (50×) time-lapse imaging of the hindgut region of immobilized YA worms. As before, the worms were fed a mixture of
E. coli OP50 RFP and red fluorescent melamine microbeads (
Figure 6a), fluorescent microbeads only (
Figure 6b), or
E. coli OP50 RFP only (
Figure 6c). In the mixed state shown in
Figure 6a, single dots and fluorescent clusters, generated by resin microparticles and a small amount of intact bacteria, and diffuse fluorescence associated with grinded bacteria are overlapping and can hardly be discriminated. In this case, the time sequence has been recorded over 180 s in order to focus on the periodicity of motion in the gut due to peristaltic action. The corresponding time evolution of the fluorescent signal in the hindgut region is shown in
Figure 6b. The peak at
t ≈ 70 s and the region extending from
t ≈ 120–160 s (also captured in the image at 150 s in
Figure 6a for instance) are caused by the transient localization of clusters of fluorescent microbeads/intact bacteria in the hindgut that are repeatedly pushed back towards the pharynx.
The somewhat asymmetric shape of the peaks, due to a slower decay of the peaks with respect to the peak rise, might possibly be related to a digestive process superposing the peristaltic activity (see discussion below and
Figure 6f). The cyclic motion of the internal load through the entire worm gut could also be clearly seen in the color plots of
Figure 5d,e. Considering the time scale of these recordings (180 s), it appears that not-grinded intact bacteria and indigestible beads somehow hinder the defecation process, which normally occurs on a time scale of about 50 s [
17].
A closer view on the initial phase after food ingestion is shown in the time-lapse sequence of
Figure 6c (50×, 60 s). In this case, the immobilized YA N2 worm was fed red fluorescent microbeads only for visualizing the dynamic behavior of the initial phase after food uptake where beads have been transported to the posterior gut section after passing the grinder without damage. Since digestion of beads does not occur, the fluorescence intensity averaged over the hindgut region is expected to be stable.
Figure 6d shows that the time evolution of fluorescence approximates a step-like curve, corresponding to the appearance of the first bead cluster in the hindgut (at
t ≈ 20 s in this case) and the temporary stability of the signal (
t ≈ 20–60 s). Subsequently, microbeads are pushed back toward the grinder by a peristaltic contraction, corresponding to the situation shown in
Figure 6a,b. Defecation may also happen but was not observed in this case. On the other hand, additional transitory accumulation in the posterior intestine could be observed if another consecutive ingestion event occurs rapidly. As shown in two of the graphs in
Figure S6, the fluorescence curves feature a two-step shape in the latter case.
In order to accurately quantify the time constant
τ associated with the digestive process of enzymatic and biochemical transformation of disrupted bacterial food, we acquired high-resolution images (50×) of the hindgut area of immobilized YA N2 worms fed with
E. coli OP50 RFP only.
Figure 6e shows a representative image sequence visualizing the evolution of the fluorescent signal. The focal plane was adjusted accurately to the mid-height of the gut in order to correctly capture the possible presence of intact bacteria.
The diffuse fluorescence signal was extracted from the raw images according to the image processing protocol described in
Figure 2c–f. At
t = 0 food indigestion occurred and no bacteria-related signal could be observed in the gut at this stage. At
t = 15 s strong diffuse fluorescence filled the whole intestinal lumen in the posterior region. As was discussed earlier, we assume that this diffuse signal is generated by a mix of small membrane fragments of disrupted bacteria, a significant amount of partly dissociated membrane compounds that still incorporate active RFP markers, and possibly fluorescent cytoplasmic content that was released after disintegration of the bacteria.
Subsequent images (
Figure 6e,
t = 30 s and 40 s) revealed a rapid decay of the fluorescent signal in the intestine. We attributed the fading of intensity to chemical digestion and absorption of nutrients in the worm gut.
Other possible reasons for a loss of signal, in particular pH sensitivity or photobleaching of RFP, relocation of the intestinal load outside the field of view due to peristaltic action (appears only on a longer time scale), and defecation (not observed before the complete fluorescence clearing in this case) can be excluded. In
Figure 6f, we quantitively analyzed the time dependence of the diffuse fluorescent signal intensity averaged over the hindgut, corresponding to the image sequence in
Figure 6e. The initial peak at
t = 15 s indicates the arrival of the bacterial load in the hindgut after food uptake and grinding. In the time span of 15 s to 50 s the plot shows the rapid signal decay, which becomes negligible for
t > 50 s in this case. The exact features of such graphs are subjected to natural variations of the biological processes and a lack of precision of certain assay parameters (e.g., defining the exact time point of food uptake or the transient bacterial accumulation phase in the grinder). Additional typical examples for different worms are reported in
Figure S7. All graphs are clearly different from the step-like shape observed for indigestible beads reflecting a constant fluorescent signal after ingestion (
Figure 6d). By averaging the recorded data, we defined an exponentially decaying curve (dashed line in
Figure 6f) and estimated a time constant
τ = 14 ± 4 s (
n = 5) for biochemical food digestion in the
C. elegans intestine. This result may also be qualitatively compared with
Figure 4d for
eat-2 pharyngeal-pumping defective mutants. In the latter case, the stable baseline of the fluorescent signal, reflecting temporary accumulation of intact bacteria in the hindgut, was superposed by a transient peak due to a new aliquot of grinded bacteria that were rapidly digested.
4.3. Infection Assay Based on M. marinum Accumulation and hsp-6::gfp Expression
A wide variety of bacterial species may infect
C. elegans, which has therefore evolved to an important model for studies of host immunity mechanisms and of pathogen virulence that are analogous to those involved during pathogenesis in humans [
23,
24,
25]. In order to validate our microfluidic platform and experimental methods with respect to specific applications, in particular for infection assays involving studies of pathogenic intestine colonization and for monitoring pathogen-induced mitochondrial stress in
C. elegans, we designed a series of experiments based on the important aquatic pathogen
M. marinum. M. marinum is a slow-growing non-tuberculous mycobacterium that causes a tuberculosis-like illness in fish [
54]. In particular, the zebrafish is a natural host to
M. marinum that was used as a model to study tuberculosis pathogenesis, as well as for antitubercular drug discovery [
55]. In humans,
M. marinum is medically important as it causes infections ranging from cutaneous granulomatous lesions to debilitating disseminated infections, possibly progressing to more invasive diseases such as tenosynovitis, septic arthritis, or osteomyelitis [
56].
M. marinum is also pathogenic to
C. elegans. For instance, it was shown that
C. elegans infected with
M. marinum displayed a highly increased mortality rate within two days post-infection [
57]. Our assays consist of feeding on-chip immobilized worms with the pathogen and measuring (i) the evolution of bacterial accumulation in the worm intestine over several hours, and (ii) quantifying specific fluorescent phenotypic worm features by imaging the time evolution of
hsp-6::gfp expression, which is an important reporter of mitochondrial stress and the activation of the mitochondrial unfolded protein response (UPR
mt) in
C. elegans [
34].
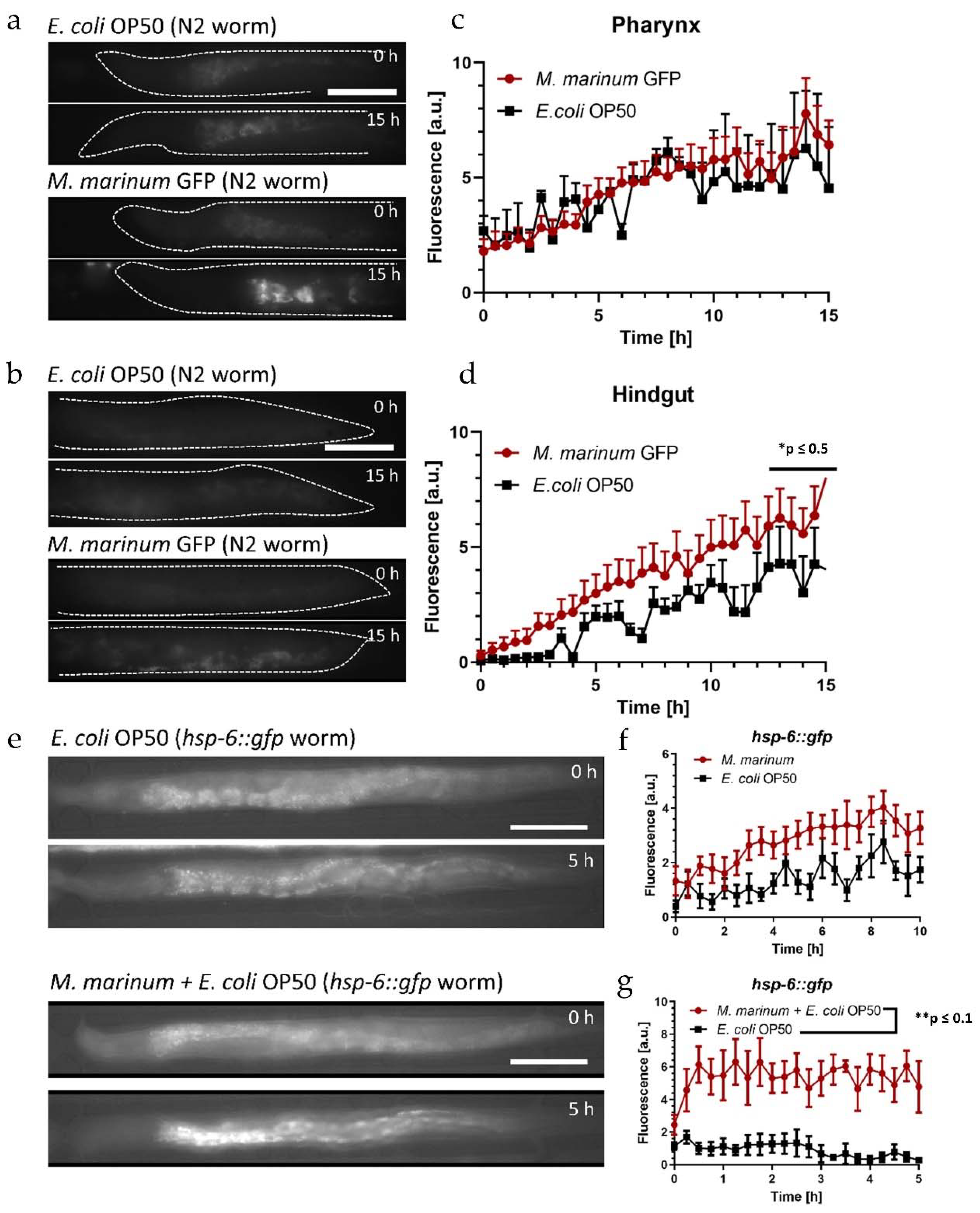
For the colonization assays, approximately 16 N2 worms were loaded in each assay unit of the device. Two units were filled with GFP labeled M. marinum suspended in S-medium. Considering our discussion above, the pH sensitivity of the GFP fluorophore related to disrupted bacteria is not an issue in the present study, as only accumulation of intact live bacteria is of interest, which are not affected by the acidic environment of the intestine. Non-fluorescent E. coli OP50 was used as reference for normal feeding conditions of the worm control groups in the other two on-chip assay units. We acquired high-resolution (50×) fluorescence images of the gut over 15 h, specifically of the pharyngeal and posterior regions, since these sections are most sensitive to bacteria accumulation.
Images for both gut sections immediately after food uptake (
t = 0 h) and after on-chip incubation (
t = 15 h) are shown in
Figure 7a,b, respectively. The evolution of the weak autofluorescent signal in a worm fed
E. coli is hardly visible with exposure conditions optimized for
M. marinum (
Figure 7a, upper panel). On the other hand, bright spots due to localized accumulation of intact
M. marinum GFP in the grinder can be seen after 15 h (
Figure 7a, lower panel). Likewise, the
M. marinum bacteria load increased in the hindgut, but the signal is distributed over a larger portion of the intestine, and thus is relatively weak (
Figure 7b, lower panel, same exposure conditions as in
Figure 7a). The graphs in
Figure 7c,d express more clearly the time evolution of the corresponding signal intensities, averaged over the pharyngeal and posterior regions, respectively. The
E. coli autofluorescence control signal increased over time indicating a progressive accumulation of bacteria in the grinder (
Figure 7c, black curve). The observation of this phenomenon was more evident in
Figure 3b (at
t = 30 h, using
E. coli HT155 GFP). The evolution of the fluorescent signal associated with
M. marinum in the pharynx (
Figure 7c, red curve) was similar to
E. coli. For a better comparison,
M. marinum values were normalized for each time point with respect to the
E. coli OP50 intensity determined in the grinder. Therefore, we deduce from
Figure 7c that no specific additional accumulation occurred in the grinder for the present assay conditions. On the other hand, the normalized
M. marinum signal in the hindgut increased faster than the
E. coli control (
Figure 7d), indicating accumulation and/or colonization possibly due to the pathogenic effect of
M. marinum. In both cases, a rise of the fluorescent signal in the worms could in principle also be partly be due to increasing autofluorescence in aging worms, however, at the early adulthood stage of the worms in the present assays this effect should be negligible [
58].
In order to evaluate the feasibility of recording UPR
mt response with our microfluidic platform, we first measured the mitochondrial stress response in YA
hsp-6::gfp mutants caused by prolonged exposure to tetracycline. For the assay, an
E. coli suspension was mixed with tetracycline to a final concentration of 60 μg/mL. Corresponding brightfield/fluorescence images are shown in
Figure S8, where a bright signal generated by
hsp-6::gfp expression in the pharyngeal region could be observed after tetracycline exposure for 8 h (
Figure S8a, lower panel). The increase in
hsp-6::gfp expression of the treated worms over 15 h is represented in the graph of
Figure S8b. To evaluate the mitochondrial stress response induced by pathogenic
M. marinum, we monitored the fluorescent
hsp-6::gfp signal in YA mutants for different feeding conditions. Fluorescence images of immobilized
hsp-6::gfp mutants acquired after the initial filling of the chip with different bacterial suspensions (
t = 0 h) and after incubation for 5 h are shown in
Figure 7e. Images of a
hsp-6::gfp mutant fed
E. coli OP50 only did not show a significant difference over the time interval of 5 h (
Figure 7e, top panel). The time evolution of fluorescence intensities for
hsp-6::gfp mutants fed
M. marinum only and the
E. coli control group is represented in the graph of
Figure 7f (averaged over the entire worm). Overall, the induced fluorescence response was higher with
M. marinum compared with
E. coli only, showing a slight increase over time, similar to the effect of tetracycline (
Figure S8b). In order to verify that mitochondrial stress in worms was not induced by food avoidance and starvation, the infection assay was repeated with a mixed suspension of
M. marinum +
E. coli OP50 thus delivering the pathogen and providing an adequate food source simultaneously. Fluorescence images of an immobilized
hsp-6::gfp mutant are shown in
Figure 7e (bottom panel). Strong fluorescence due to
hsp-6::gfp expression in the anterior region of the intestinal track is clearly visible after 5 h. The corresponding fluorescence signal averaged over the whole worm is shown in the graph of
Figure 7g (red curve). Similar to
M. marinum only (
Figure 7f), the
hsp-6::gfp induced signal was above the control values for feeding with
E. coli only. However, in the latter case, we observed a faster onset of stress-related
hsp-6::gfp expression and a higher overall fluorescent intensity. This could possibly be due to a higher uptake of
E. coli being a suitable food source, entailing simultaneous ingestion of an enhanced amount of
M. marinum in the mixed feeding suspension, and thus resulting in an enhanced stress response.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






