
Cellular Aquaculture: Prospects and Challenges

 , , ,
, , ,
Abstract
:1. Introduction
2. Scientific Challenges
2.1. Challenges in Seafood Sustainability
- Overfishing, illegal, unreported, and unregulated fishery activities, which are almost 20% of the total catch ($20 billion) [26].
- Lack of proper monitoring systems to control fraud and mislabeling. Almost 100% of red snapper is mislabeled in sushi bars, 77% in restaurants, and 88% at the retail level [27].
- Lack of sustainable protein sources for fish feed production, which makes aquaculture very dependent on the ocean for providing fishmeal [7].
- Water pollution due to intensive aquaculture activities [26].
- More than 40% of waste during seafood processing, which contains high-value protein, minerals, and oil, is being discarded in many cases [28].
- Lack of proper traceability due to the fragmented industry.
- Lack of accessibility to the workforce [27].
- Being heavily dependent on foreign suppliers; more than 90% of the seafood products in the U.S. are imported from other countries, which becomes more challenging during the pandemic with decreased control of product safety [27].
2.2. Challenges in Seafood Safety
- Microplastics in seafood;
- Harmful algal blooms (HABs) and harvesting region closure;
- Antibiotic-resistant bacteria and antibiotic residues.
2.3. Plant- and Insect-Based Protein Challenges
- Consumer preference, and the fact that plant- and insect-based foods are not currently substitutes for conventionally-produced meat [29].
- A sustainable insect production system depends on food wastes, which may pose several food safety risks, which need to be addressed [30].
- Plants do not contain all the required nutrients for human health and may cause health issues due to nutrient deficiency [29].
3. Cell-Based Seafood Challenges
- Diverse available seafood and lack of available cell line sources: Despite many different available seafood species in the ocean for human consumption, only a few seafood cell lines are available, mainly for toxicology and medical studies. This limitation is due to the concept being very novel, and not that many researchers have access to seafood at different life-stages (eggs to broodstock).
- Lack of proper serum-free media: One of the most important factors in the formulation of cell culture media, which regulates cell growth and proliferation, is serum. Although Fetal Bovine Serum (FBS) is the most commonly used media supplement, its application for cultivated food production is limited due to the cost, possible contamination, high demand, limited supply, variation from batch to batch, inability to grow specific cells, animal suffering and environmental consequences from FBS production, high ratio of protein, and downstream processing difficulties [56].
- Lack of efficient techniques for optimizing the cell culture media to reduce the cost of production: Despite some efforts for optimizing media, machine learning tools have not been applied comprehensively for serum-free media optimization [57].
- All these limitations direct us to develop seafood cell lines from unsustainable, economically critical seafood species, optimize cost-effective serum-free media using innovative machine learning approaches by applying sustainable protein sources, and increase the yield and similarity to phenotypic characters of target seafood cells. For an effective media formulation, establishing the effects of components of media on cells and deciding its concentration accordingly would counter the negative effects of serum.
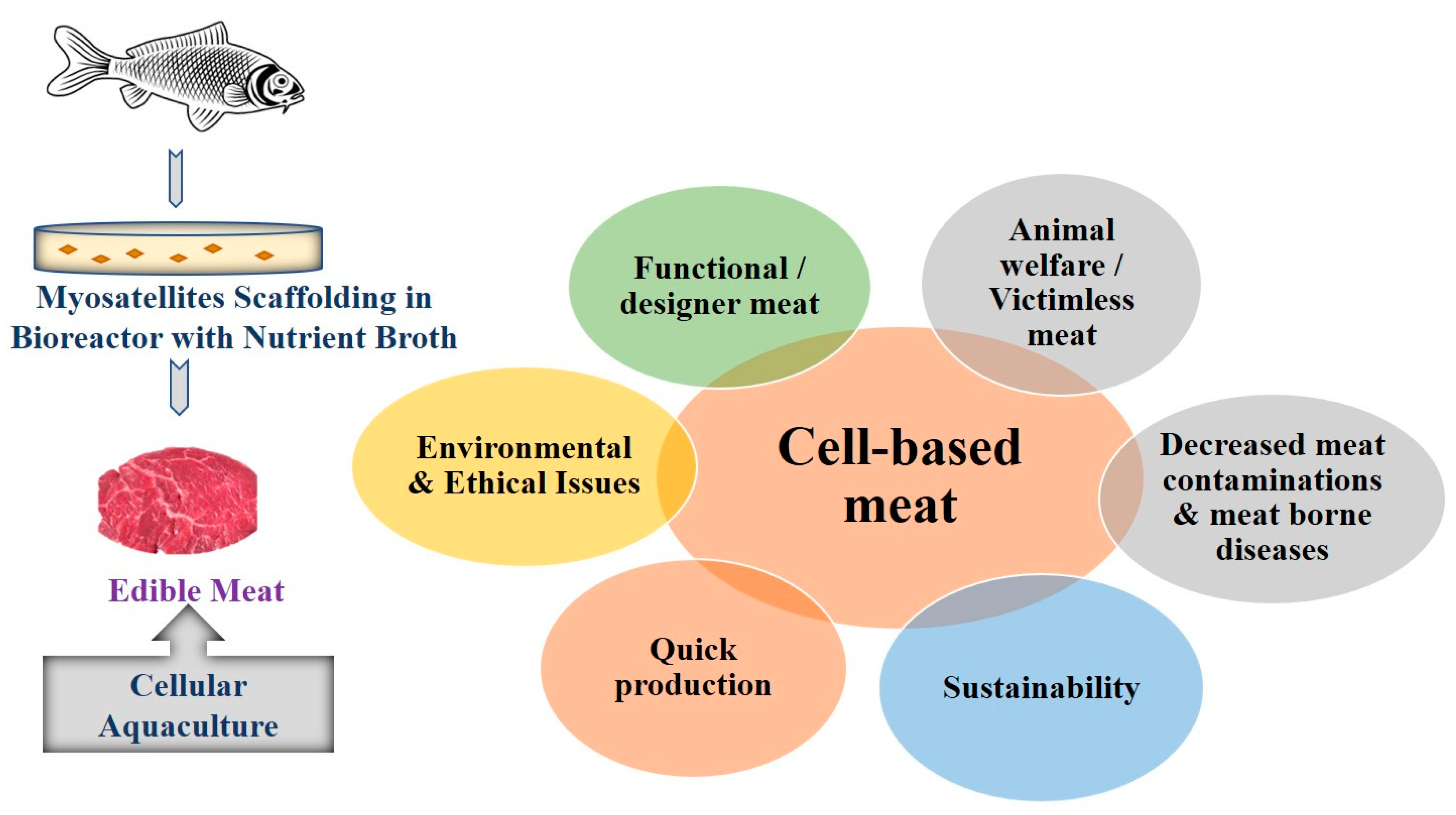
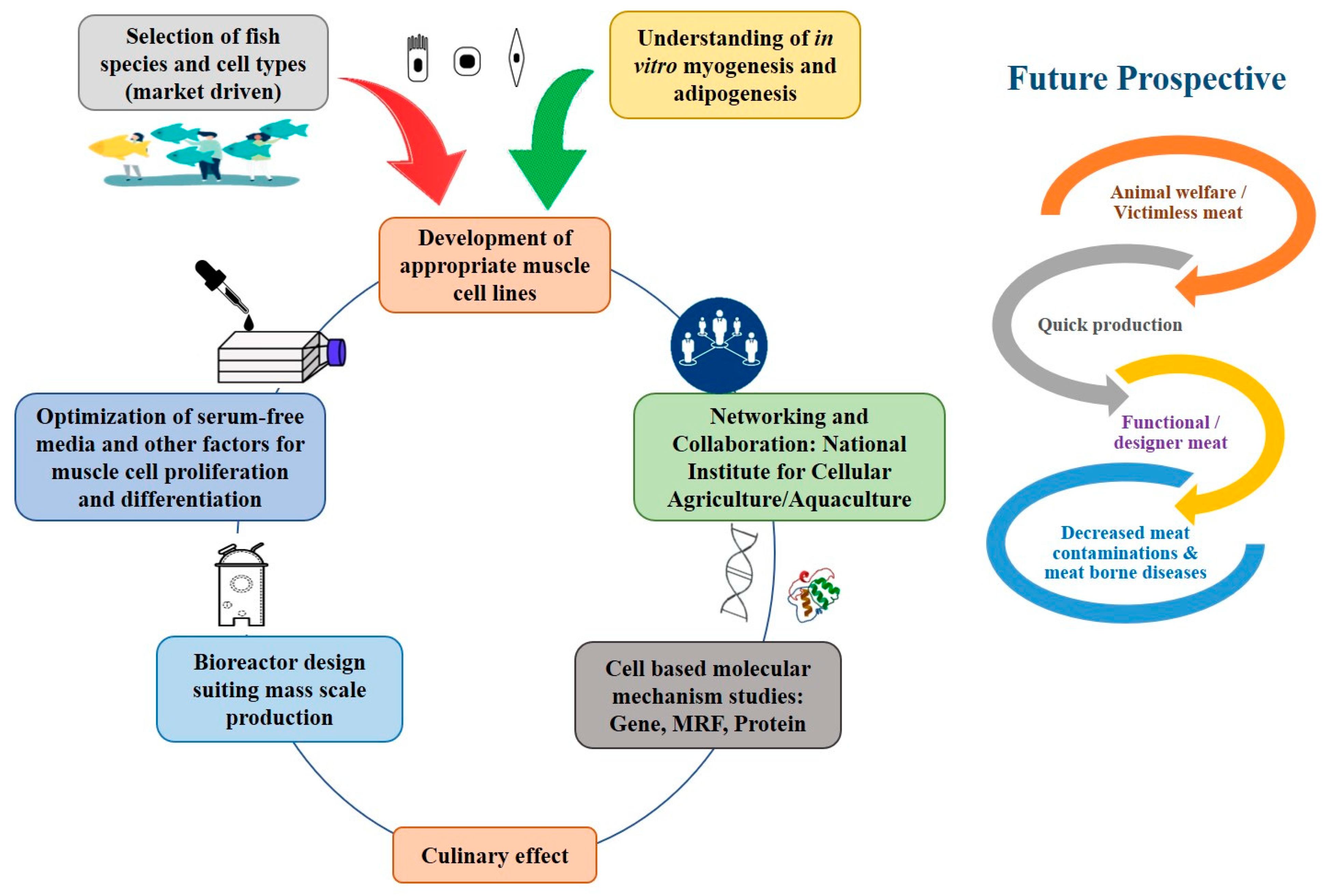
4. Perspectives of Cellular Aquaculture
5. Requirements for In Vitro Meat
5.1. Cell Source and Growth Factors
5.2. Scaffold
5.3. Growth Medium
5.4. Bioreactor
5.5. Characterization of Cell Culture System
6. Genetic and Molecular Markers
Gene Profiling
7. Molecular-Based Studies
8. Sensors, Devices, and Systems Are Available and Used in Cellular Aquaculture Ventures
8.1. Bioreactor Monitor
8.2. Spoilage Detectors
9. Cell Line Repository
Importance of Cell Line Repository
10. Challenges in Cellular Aquaculture
11. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Costello, C.; Cao, L.; Gelcich, S.; Cisneros-Mata, M.Á.; Free, C.M.; Froehlich, H.E.; Golden, C.; Ishimura, G.; Maier, J.; Macadam-Somer, I.; et al. The future of food from the sea. Nature 2020, 588, 95–100. [Google Scholar] [CrossRef] [PubMed]
- The World Bank. Raising More Fish to Meet Rising Demand. Available online: https://www.worldbank.org/en/news/feature/2014/ (accessed on 8 July 2021).
- Éliás, B.; Jámbor, A. Food Security and COVID-19: A Systematic Review of the First-Year Experience. Sustainability 2021, 13, 5294. [Google Scholar] [CrossRef]
- Mardones, F.O.; Rich, K.M.; Boden, L.A.; Moreno-Switt, A.I.; Caipo, M.L.; Zimin-Veselkoff, N.; Alateeqi, A.M.; Baltenweck, I. The COVID-19 Pandemic and Global Food Security. Front. Veter.-Sci. 2020, 7, 578508. [Google Scholar] [CrossRef] [PubMed]
- Organization for Economic Co-Operation and Development/Food and Agricultural Organization. OECD-FAO Agricultural Outlook 2020–2029. 2020. Available online: https://www.fao.org/fishery/en/publications/287024 (accessed on 27 December 2021).
- Food and Agriculture Organization (FAO). The State of World Fisheries and Aquaculture; FAO Fisheries and Aquaculture Department: Rome, Italy, 2018; ISBN 978-92-5-130562-1. [Google Scholar]
- Alexandratos, N.; Bruinsma, J. World Agriculture Towards 2030/2050: 2012 Revision; FAO: Rome, Italy, 2012. [Google Scholar]
- Kanianska, R. Agriculture and its impact on land-use, environment, and ecosystem services. In Landscape Ecology—The Influences of Land Use and Anthropogenic Impacts of Landscape Creation; IntechOpen: London, UK, 2016; pp. 1–26. [Google Scholar]
- Ismail, I.; Hwang, Y.-H.; Joo, S.-T. Meat analog as future food: A review. J. Anim. Sci. Technol. 2020, 62, 111–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Post, M.J. Cultured beef: Medical technology to produce food. J. Sci. Food Agric. 2014, 94, 1039–1041. [Google Scholar] [CrossRef]
- Li, N.; Guo, L.; Guo, H. Establishment, characterization, and transfection potential of a new continuous fish cell line (CAM) derived from the muscle tissue of grass goldfish (Carassius auratus). Vitr. Cell. Dev. Biol. -Anim. 2021, 27, 912–931. [Google Scholar] [CrossRef]
- Wang, L.; Cao, Z.; Liu, Y.; Xiang, Y.; Sun, Y.; Zhou, Y.; Wang, S.; Guo, W. Establishment and characterization of a new cell line from the muscle of humpback grouper (Cromileptes altivelis). Fish Physiol. Biochem. 2020, 46, 1897–1907. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Q.; Guo, Z.; Zhao, Y.; Luo, S.; Yu, T.; Zhang, D.; Wang, G. Identification of the Nrf2 in the fathead minnow muscle cell line: Role for a regulation in response to H2O2 induced the oxidative stress in fish cell. Fish Physiol. Biochem. 2020, 46(5), 1699–1711. [Google Scholar] [CrossRef]
- Vishnolia, K.K.; Martin, N.R.W.; Player, D.J.; Spikings, E.; Lewis, M.P. Zebrafish skeletal muscle cell cultures: Monolayer to three-dimensional tissue engineered collagen constructs. bioRxiv 2020. [Google Scholar]
- Gao, Y.; Zhou, H.; Gao, Z.; Jiang, H.; Wang, X.; Mai, K.; He, G. Establishment and characterization of a fibroblast-like cell line from the muscle of turbot (Scophthalmus maximus L.). Fish Physiol. Biochem. 2019, 45, 1129–1139. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, N.; Goswami, M.; Srivastava, J.K.; Mishra, A.K.; Lakra, W.S. Establishment and Characterization of a New Muscle Cell Line of Zebrafish (Danio rerio) as an In Vitro Model for Gene Expression Studies. Anim. Biotechnol. 2016, 27, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.M.; Zheng, Y.; You, F.; Wu, Z.H.; Tan, X.; Jiao, S.; Zhang, P.J. Comparison of growth characteristics between skeletal muscle satellite cell lines from diploid and triploid olive flounder Paralichthys olivaceus. PeerJ 2016, 4, e1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubey, A.; Goswami, M.; Yadav, K.; Mishra, A.; Kumar, A. Establishment of a novel muscle cell line from Wallago attu for in vitro study of pesticide toxicity. Gene Cell Tissue 2015, 2. [Google Scholar] [CrossRef]
- Lai, Y.S.; Chiou, P.P.; Chen, W.J.; Chen, Y.C.; Chen, C.W.; Chiu, I.S.; Chen, S.D.; Cheng, Y.H.; Chang, C.Y. Characterization of apoptosis induced by grouper iridovirus in two newly established cell lines from barramundi, Lates calcarifer (Bloch). J. Fish Dis. 2008, 31, 825–834. [Google Scholar] [CrossRef]
- Rougée, L.; Ostrander, G.K.; Richmond, R.H.; Lu, Y. Establishment, characterization, and viral susceptibility of two cell lines derived from goldfish Carassius auratus muscle and swim bladder. Dis. Aquat. Org. 2007, 77, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Lu, Y. Establishment and characterization of two cell lines from bluefin trevally Caranx melampygus. Dis. Aquat. Org. 2006, 68, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Montgomery-Brock, D.; Lee, C.S.; Lu, Y. Establishment, characterization and viral susceptibility of 3 new cell lines from snakehead, Channa striatus (Blooch). Methods Cell Sci. 2004, 25, 155–166. [Google Scholar] [CrossRef]
- Wang, G.; Lapatra, S.; Zeng, L.; Zhao, Z.; Lu, Y. Establishment, growth, cryopreservation and species of origin identification of three cell lines from white sturgeon, Acipenser transmontanus. Methods Cell Sci. 2004, 25, 211–220. [Google Scholar] [CrossRef]
- Msangi, S.; Kobayashi, M.; Batka, M.; Vannuccini, S.; Dey, M.M.; Anderson, J.L. Fish to 2030: 655 prospects for fisheries and aquaculture. World Bank Rep. 2013, 83177, 102. [Google Scholar]
- Gephart, J.A.; Froehlich, H.E.; Branch, T.A. To create sustainable seafood industries, the United States needs a better accounting of imports and exports. Proc. Natl. Acad. Sci. USA 2019, 116, 9142–9146. [Google Scholar] [CrossRef] [Green Version]
- Krueck, N.C.; Ahmadia, G.N.; Possingham, H.; Riginos, C.; Treml, E.; Mumby, P.J. Marine Reserve Targets to Sustain and Rebuild Unregulated Fisheries. PLoS Biol. 2017, 15, e2000537. [Google Scholar] [CrossRef] [Green Version]
- Warner, K.; Timme, W.; Lowell, B.; Hirschfield, M. Oceana Study Reveals Seafood Fraud Nationwide; Oceana: Washington, DC, USA, 2013; pp. 1–69. [Google Scholar]
- Ovissipour, M.; Kenari, A.A.; Motamedzadegan, A.; Nazari, R.M. Optimization of Enzymatic Hydrolysis of Visceral Waste Proteins of Yellowfin Tuna (Thunnus albacares). Food Bioprocess Technol. 2010, 5, 696–705. [Google Scholar] [CrossRef]
- Graça, J.; Oliveira, A.; Calheiros, M.M. Meat, beyond the plate. Data-driven hypotheses for understanding consumer willingness to adopt a more plant-based diet. Appetite 2015, 90, 80–90. [Google Scholar] [CrossRef]
- Mézes, M. Insect protein as new alternative of protein supply. Állattenyésztés És Takarmányozás 2018, 67, 287–296. [Google Scholar]
- Batish, I.; Brits, D.; Valencia, P.; Miyai, C.; Rafeeq, S.; Xu, Y.; Galanopoulos, M.; Sismour, E.; Ovissipour, R. Effects of Enzymatic Hydrolysis on the Functional Properties, Antioxidant Activity and Protein Structure of Black Soldier Fly (Hermetia illucens) Protein. Insects 2020, 11, 876. [Google Scholar] [CrossRef] [PubMed]
- FDA. Formal Agreement between FDA and USDA Regarding Oversight of Human Food Produced Using Animal Cell Technology Derived from Cell Lines of USDA-Amenable Species. 2019. Available online: https://www.fda.gov/food/domestic-interagency-agreements-food/formal-agreement-between-fda-and-usda-regardingoversight-human-food-produced-using-animal-cell (accessed on 9 July 2021).
- Zaraska, M. Lab-Grown Beef Taste Test: ‘Almost’ Like a Burger. 2013. Available online: http://www.washingtonpost.com/national/health-science/lab-grown-beef-taste-test-almostlike-a-burger/2013/08/05/921a5996-fdf4-11e2-96a8-d3b921c0924a_story.html (accessed on 22 January 2022).
- Rubio, N.; Datar, I.; Stachura, D.; Kaplan, D.; Krueger, K. Cell-Based Fish: A Novel Approach to Seafood Production and an Opportunity for Cellular Agriculture. Front. Sustain. Food Syst. 2019, 3, 43. [Google Scholar] [CrossRef] [Green Version]
- Bryce, E. Lab-Cultured Seafood Is Coming. But Can It Actually Relieve Pressure on Global Fisheries. Anthopocene 2021. Available online: https://www.anthropocenemagazine.org/2021/04/ (accessed on 21 January 2022).
- Datar, I.; Betti, M. Possibilities for an in vitro meat production system. Innov. Food Sci. Emerg. Technol. 2010, 11, 13–22. [Google Scholar] [CrossRef]
- Waltz. No bones, no scales, no eyeballs: Appetite grows for cell-based seafood. Nat. Biotechnol. 2021, 39, 1483–1485. [Google Scholar] [CrossRef]
- Bhat, Z.F.; Fayaz, H. Prospectus of cultured meat—advancing meat alternatives. J. Food Sci. Technol. 2011, 48, 125–140. [Google Scholar] [CrossRef] [Green Version]
- Ford, B.J. Culturing Meat For The Future: Anti-Death Versus Anti-Life. Death Anti-Death 2010, 7, 55–80. [Google Scholar]
- Ross, R. Growth of Smooth Muscle in Culture and Formation of Elastic Fibers. 1971, 172–186. J. Cell Biol. 1971, 172–186. [Google Scholar] [CrossRef]
- Markowski, J. Moments in Meat History Part IX—In-Vitro Meat. Food Technology, 31 December; 2013.
- Macintyre, B. Test-tube meat science’s next leap. The Australian, 20 January 2007. [Google Scholar]
- Datar, I. Mark Post’s Cultured Beef. New Harvest, 3 November 2015. [Google Scholar]
- Siegelbaum, D.J. In Search of a Test-Tube Hamburger. Time, 23 April 2008. [Google Scholar]
- Harvey, C. This Brooklyn Startup Wowed The Science Community With Lab-Made ’Meat Chips’. Business Insider, 26 September 2014. [Google Scholar]
- Fountain, H. A Lab-Grown Burger Gets a Taste Test. The New York Times, 5 August 2013. [Google Scholar]
- Wohlsen, M. Cow Milk Without the Cow is Coming to Change Food Forever. Wired, 15 April 2015. [Google Scholar]
- Leichman, A.K. Coming soon: Chicken meat without slaughter. Israel21c, 19 November 2015. [Google Scholar]
- Crew, B. Cost of lab-grown burger patty drops from $325,000 to $11.36. Science Alert, 2 April 2015. [Google Scholar]
- Burwood-Taylor, L. New Crop Capital Closes $25m Fund, Invests in Beyond Meat. AgFunderNews, 17 March 2016. [Google Scholar]
- Zacharias, N. The Race to Disrupt Animal Agriculture Just Got a $25 Million Shot in the Arm, and a New Non-Profit. One Green Planet 2016.
- Hanson, H. ‘World’s First’ Lab-Grown Meatball Looks Pretty Damn Tasty. The Huffington Post, 2 February 2016. [Google Scholar]
- Kowitt, B. The First ’Animal-Free’ Ice Cream Hits the Market. Fortune, 11 July 2019. [Google Scholar]
- Wolf, M. USDA Awards $10 Million to Tufts University to Establish a Cultivated Protein Center of Excellence. The Spoon, 13 October 2021. [Google Scholar]
- Van der Weele, C.; Feindt, P.; van der Goot, A.J.; van Mierlo, B.; van Boekel, M. Meat alternatives: An integrative comparison. Trends Food Sci. Technol. 2019, 88, 505–512. [Google Scholar] [CrossRef]
- van der Valk, J.; Brunner, D.; De Smet, K.; Svenningsen, Å.F.; Honegger, P.; Knudsen, L.E.; Lindl, T.; Noraberg, J.; Price, A.; Scarino, M.L.; et al. Optimization of chemically defined cell culture media—Replacing fetal bovine serum in mammalian in vitro methods. Toxicol. Vitr. 2010, 24, 1053–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, H.-H.; Gao, X.; DeKeyser, J.-M.; Fetterman, K.A.; Pinheiro, E.A.; Weddle, C.J.; Fonoudi, H.; Orman, M.V.; Romero-Tejeda, M.; Jouni, M.; et al. Negligible-Cost and Weekend-Free Chemically Defined Human iPSC Culture. Stem Cell Rep. 2020, 14, 256–270. [Google Scholar] [CrossRef] [Green Version]
- Edelman, P.D. In Vitro Meat Production. 2003. Available online: http://new-harvest.org/wp-content/uploads/2013/03/Edelman.pdf (accessed on 15 July 2014).
- Bhat, Z.; Bhat, H. Animal-free Meat Biofabrication. Am. J. Food Technol. 2011, 6, 441–459. [Google Scholar] [CrossRef] [Green Version]
- Collins, C.A.; Zammit, P.S.; Ruiz, A.P.; Morgan, J.E.; Partridge, T.A. Apopulation of myogenic stem cells that survives skeletal muscle aging. Stem Cells 2007, 25, 885–889. [Google Scholar] [CrossRef]
- Kim, M.J.; Choi, Y.S.; Yang, S.H.; Hong, H.N.; Cho, S.W.; Cha, S.M.; Pak, J.H.; Kim, C.W.; Kwon, S.W.; Park, C.J. Muscle regeneration by adipose tissue-derived adult stem cells attached to injectable PLGA spheres. Biochem. Biophys. Res. Commun. 2006, 348, 386–392. [Google Scholar] [CrossRef]
- Weele, C.V.; Driessen, C. Emerging profiles for cultured meat; ethics through and as design. Animals 2013, 3, 647–662. [Google Scholar] [CrossRef] [PubMed]
- Mattick, C.S.; Wetmore, J.M.; Allenby, B.R. An Anticipatory Social Assessment of Factory-Grown Meat. IEEE Technol. Soc. Mag. 2015, 34, 56–64. [Google Scholar] [CrossRef]
- Williams, J. Meat Derived from Stem Cells: How, What and Why. 2012. Available online: http://medlink-uk.net/wp-content/uploads/pathprojectsstemcells2012/WilliamsJ.pdf (accessed on 15 July 2014).
- Van Eelen, W.F.; van Kooten, W.J.; Westerhof, W. Industrial Production of Meat from In Vitro Cell Cultures. Patent Description. International Patent WO/1999/031223, 1999. Available online: http://www.wipo.int/pctdb/en/wo.jsp?wo=1999031223 (accessed on 25 March 2009).
- Madrigal, A. Scientists Flesh out Plans to Grow (and Sell) Test Tube Meat. 2008 [2004-11-4]. Available online: http://www.wired.com/science/discoveries/news/2008/04/invitro_meat (accessed on 27 January 2022).
- Alexander, R. In vitro meat: A vehicle for the ethical rescaling of the factory farming industry and in vivo testing or an intractable enterprise? Intersect 2011, 4, 42–47. [Google Scholar]
- Chriki, S.; Hocquette, J.F. The myth of cultured meat: A review. Front. Nutr. 2020, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.L. Test tube meat on the menu? Nat. Biotechnol. 2009, 27, 873. [Google Scholar] [CrossRef]
- Schneider, Z. In vitro meat: Space travel, cannibalism, and federal regulation. Houst. Law Rev. 2013, 50, 991–1024. [Google Scholar]
- Fox News. 2009. Available online: http://www.foxnews.com/story/2009/05/07/who-meat-from-pigs-infected-with-swine-flu-should-not-be-eaten/ (accessed on 31 July 2014).
- Johnson, R.; Gibbs, C., Jr. Creutzfeldt-Jakob disease and related transmissible spongiform encephalopathies. N. Engl. J. Med. 1998, 339, 1994–2004. [Google Scholar] [CrossRef]
- Gold, M. The Global Benefits of Eating Less Meat; Compassion in World Farming: Godalming, UK, 2004. [Google Scholar]
- Sanders, T.A.B. The nutritional adequacy of plant-based diets. Proc. Nutr. Soc. 1999, 58, 265–269. [Google Scholar] [CrossRef] [Green Version]
- Stephens, N.; Silvioc, L.D.; Dunsfordb, I.; Ellisd, M.; Glencrosse, A.; Sexton, A. Bringing cultured meat to market: Technical, socio-political, and regulatory challenges in cellular agriculture. Trends Food Sci. Technol. 2018, 78, 155–166. [Google Scholar] [CrossRef]
- Péault, B.; Rudnicki, M.; Torrente, Y.; Cossu, G.; Tremblay, J.P.; Partridge, T.; Gussoni, E.; Kunkel, L.M.; Huard, J. Stem and Progenitor Cells in Skeletal Muscle Development, Maintenance, and Therapy. Mol. Ther. 2007, 15, 867–877. [Google Scholar] [CrossRef]
- Yin, H.; Price, F.; Rudnicki, M.A. Satellite Cells and the Muscle Stem Cell Niche. Physiol. Rev. 2013, 93, 23–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burton, N.M.; Vierck, J.; Krabbenhoft, L.; Bryne, K.; Dodson, M.V. Methods for animal satellite cell culture under a variety of conditions. J. Tissue Cult. Methods 2000, 22, 51–61. [Google Scholar] [CrossRef]
- Vitello, L.; Radu, C.; Malerba, A.; Segat, D.; Cantini, M.; Carraro, U.; Baroni, M.D. Enhancing myoblast proliferation by using myogenic factors: A promising approach for improving fiber regeneration in sport medicine and skeletal muscle diseases. Basic Appl. Myol. 2004, 14, 45–51. [Google Scholar]
- Nicola, N.A.; Babon, J.J. Leukemia inhibitory factor (LIF). Cytokine Growth Factor Rev. 2015, 26, 533–544. [Google Scholar] [CrossRef] [Green Version]
- Allen, R.E.; Boxhorn, L.K. Regulation of skeletal muscle satellite cell proliferation and differentiation by transforming growth factor-beta, insulin-like growth factor I, and fibroblast growth factor. J. Cell. Physiol. 1989, 138, 311–315. [Google Scholar] [CrossRef]
- Pawlikowski, B.; Vogler, T.O.; Gadek, K.; Olwin, B.B. Regulation of skeletal muscle stem cells by fibroblast growth factors. Dev. Dyn. 2017, 246, 359–367. [Google Scholar] [CrossRef] [Green Version]
- Kollias, H.D.; McDermott, J.C. Transforming growth factor-β and myostatin signaling in skeletal muscle. J. Appl. Physiol. 2008, 104, 579–587. [Google Scholar] [CrossRef] [Green Version]
- Miao, Z.G.; Zhang, L.P.; Fu, X.; Yang, Q.Y.; Zhu, M.J.; Dodson, M.V.; Du, M. Invited review: Mesenchymal progenitor cells in intramuscular connective tissue development. Animal 2016, 10, 75–81. [Google Scholar] [CrossRef] [Green Version]
- Krieger, J.; Park, B.-W.; Lambert, C.R.; Malcuit, C. 3D skeletal muscle fascicle engineering is improved with TGF-β1 treatment of myogenic cells and their co-culture with myofibroblasts. PeerJ 2018, 6, e4939. [Google Scholar] [CrossRef]
- Veliça, P.; Bunce, C.M. A quick, simple and unbiased method to quantify C2C12 myogenic differentiation. Muscle Nerve 2011, 44, 366–370. [Google Scholar] [CrossRef]
- Bhat, Z.F.; Bhat, H. Tissue engineered meat-future meat. J. Stored Prod. Postharvest Res. 2011, 2, 1–10. [Google Scholar]
- Rozario, T.; DeSimone, D.W. The extracellular matrix in development and morphogenesis: A dynamic view. Dev. Biol. 2010, 341, 126–140. [Google Scholar] [CrossRef] [Green Version]
- DiMaio, T. This Scientist Is Developing New Cell Lines for Slaughter-Free Meat; The Good Food Institute: Washington, DC, USA, 2019; Available online: https://gfi.org/blog/gareth-sullivan-cell-lines-research-grant/ (accessed on 15 February 2022).
- Loh, Q.L.; Choong, C. Three-Dimensional Scaffolds for Tissue Engineering Applications: Role of Porosity and Pore Size. Tissue Eng. Part B Rev. 2013, 19, 485–502. [Google Scholar] [CrossRef] [Green Version]
- Courtenay, J.C.; Sharma, R.I.; Scott, J.L. Recent Advances in Modified Cellulose for Tissue Culture Applications. Molecules 2018, 23, 654. [Google Scholar] [CrossRef] [Green Version]
- Hickey, R.J.; Modulevsky, D.J.; Cuerrier, C.M.; Pelling, A.E. Customizing the Shape and Microenvironment Biochemistry of Biocompatible Macroscopic Plant-Derived Cellulose Scaffolds. ACS Biomater. Sci. Eng. 2018, 4, 3726–3736. [Google Scholar] [CrossRef] [Green Version]
- Reddy, N.; Jiang, Q.; Yang, Y. Novel Wheat Protein Films as Substrates for Tissue Engineering. J. Biomater. Sci. Polym. Ed. 2011, 22, 2063–2077. [Google Scholar] [CrossRef]
- Zeltinger, J.; Sherwood, J.K.; Graham, D.A.; Müeller, R.; Griffith, L.G. Effect of Pore Size and Void Fraction on Cellular Adhesion, Proliferation, and Matrix Deposition. Tissue Eng. 2001, 7, 557–572. [Google Scholar] [CrossRef]
- Bhat, Z.F.; Morton, J.D.; Mason, S.L.; Bekhit, A.E.A.; Bhat, H.F. Technological, Regulatory, and Ethical Aspects ofIn VitroMeat: A Future Slaughter-Free Harvest. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1192–1208. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Ren, G.; Sha, Z.; Shi, C. Establishment of a continuous embryonic cell line from Japanese flounder Paralichthys olivaceus for virus isolation. Dis. Aquat. Org. 2004, 60, 241–246. [Google Scholar] [CrossRef]
- Lakra, W.S.; Bhonde, R.R.; Sivakumar, N.; Ayyappan, S. A new fibroblast like cell line from the fry of golden mahseer Tor putitora (Ham). Aquaculture 2006, 253, 238–243. [Google Scholar] [CrossRef]
- Freshney, R.I. Culture of Animal Cells—A Manual of Basic Technique and Specialized Applications, 6th ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2010. [Google Scholar]
- Aswad, H.; Jalabert, A.; Rome, S. Depleting extracellular vesicles from foetal bovine serum alters proliferation and differentiation of skeletal muscle cells in vitro. BMC Biotechnol. 2016, 16, 32. [Google Scholar] [CrossRef] [Green Version]
- Brunner, D.; Frank, J.; Appl, H.; Schöffl, H.; Pfaller, W.; Gstraunthaler, G. Serum-free cell culture: The serum-free media interactive online database. ALTEX 2010, 27, 53–62. [Google Scholar] [CrossRef]
- Shiozuka, M.; Kimura, I. Improved Serum-free Defined Medium for Proliferation and Differentiation of Chick Primary Myogenic Cells. Zool. Sci. 2000, 17, 201–207. [Google Scholar] [CrossRef] [Green Version]
- Carrier, R.L.; Rupnick, M.; Langer, R.; Schoen, F.J.; Freed, L.E.; Vunjak-Novakovic, G. Effects of oxygen on engineered cardiac muscle. Biotechnol. Bioeng. 2002, 78, 617–625. [Google Scholar] [CrossRef]
- Van der Weele, C.; Tramper, J. Cultured meat: Every village its own factory? Trends Biotechnol. 2014, 32, 294–296. [Google Scholar] [CrossRef]
- Munteanu, C.; Mireşan, V.; Răducu, C.; Ihuţ, A.; Uiuiu, P.; Pop, D.; Neacşu, A.; Cenariu, M.; Groza, I. Can Cultured Meat Be an Alternative to Farm Animal Production for a Sustainable and Healthier Lifestyle? Front. Nutr. 2021, 8, 8. [Google Scholar] [CrossRef]
- Zhu, M.M.; Mollet, M.; Hubert, R.S.; Kyung, Y.S.; Zhang, G.G. Industrial production of therapeutic proteins: Cell lines, cell culture, and purification. In Handbook of Industrial Chemistry and Biotechnology; Springer: Cham, Switzerland, 2017; pp. 1639–1669. [Google Scholar]
- Oyeleye, O.O.; Ogundeji, S.T.; Ola, S.I.; Omitogun, O.G. Basics of animal cell culture: Foundation for modern science. Biotechnol. Mol. Biol. Rev. 2016, 11, 6–16. [Google Scholar] [CrossRef]
- Yoshino, K.; Iimura, E.; Saijo, K.; Iwase, S.; Fukami, K.; Ohno, T.; Obata, Y.; Nakamura, Y. Essential role for gene profiling analysis in the authentication of human cell lines. Hum. Cell 2006, 19, 43–48. [Google Scholar] [CrossRef]
- Hukku, B.; Halton, D.M.; Mally, M.; Peterson, W.D. Cell Characterization by Use of Multiple Genetic Markers. Drug Resist. Leuk. Lymphoma III 1984, 172, 13–31. [Google Scholar] [CrossRef]
- Didion, J.P.; Buus, R.J.; Naghashfar, Z.; Threadgill, D.W.; Morse, H.C.; De Villena, F.P.-M. SNP array profiling of mouse cell lines identifies their strains of origin and reveals cross-contamination and widespread aneuploidy. BMC Genom. 2014, 15, 847. [Google Scholar] [CrossRef] [Green Version]
- Parodi, B.; Aresu, O.; Bini, D.; Lorenzini, R.; Schena, F.; Visconti, P.; Cesaro, M.; Ferrera, D.; Andreotti, V.; Ruzzon, T. Species Identification and Confirmation of Human and Animal Cell Lines: A PCR-Based Method. Biotechniques 2002, 32, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Lorenzi, P.L.; Reinhold, W.; Varma, S.; Hutchinson, A.A.; Pommier, Y.; Chanock, S.J.; Weinstein, J.N. DNA fingerprinting of the NCI-60 cell line panel. Mol. Cancer Ther. 2009, 8, 713–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stulberg, C.S.; Peterson, W.D., Jr.; Simpson, W.F. Identification of cells in culture. Am. J. Hematol. 1976, 1, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, J.; Hukku, B. Chapter 11 Cell Line Characterization and Authentication. Methods Cell Biol. 1998, 57, 203–216. [Google Scholar] [CrossRef]
- Masters, J.R.; Thomson, J.A.; Daly-Burns, B.; Reid, Y.A.; Dirks, W.G.; Packer, P.; Toji, L.H.; Ohno, T.; Tanabe, H.; Arlett, C.F.; et al. Short tandem repeat profiling provides an international reference standard for human cell lines. Proc. Natl. Acad. Sci. USA 2001, 98, 8012–8017. [Google Scholar] [CrossRef] [Green Version]
- Robin, T.; Capes-Davis, A.; Bairoch, A. CLASTR: The Cellosaurus STR similarity search tool-a precious help for cell line authentication. Int. J. Cancer 2020, 146, 1299–1306. [Google Scholar] [CrossRef] [Green Version]
- Freedman, L.P.; Gibson, M.C.; Ethier, S.P.; Soule, H.R.; Neve, R.M.; Reid, Y.A. Reproducibility: Changing the policies and culture of cell line authentication. Nat. Methods 2015, 12, 493–497. [Google Scholar] [CrossRef] [Green Version]
- Wagg, S.K.; Lee, L.E.J. A proteomics approach to identifying fish cell lines. Proteomics 2005, 5, 4236–4244. [Google Scholar] [CrossRef]
- Millan-Cubillo, A.F.; Martin-Perez, M.; Ibarz, A.; Fernandez-Borras, J.; Gutiérrez, J.; Blasco, J. Proteomic characterization of primary cultured myocytes in a fish model at different myogenesis stages. Sci. Rep. 2019, 9, 14126. [Google Scholar] [CrossRef] [Green Version]
- Ong, K.J.; Johnston, J.; Datar, I.; Sewalt, V.; Holmes, D.; Shatkin, J.A. Food safety considerations and research priorities for the cultured meat and seafood industry. Compr. Rev. Food Sci. Food Saf. 2021, 20, 5421–5448. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.; Yoon, J.W.; Kim, M.; Lee, H.J.; Jeong, J.; Ryu, M.; Jo, C.; Lee, C. Muscle stem cell isolation and in vitro culture for meat production: A methodological review. Compr. Rev. Food Sci. Food Saf. 2020, 20, 429–457. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Dalrymple, B. Transcriptomics of Meat Quality. In New Aspects of Meat Quality; Woodhead Publishing: Sawston, UK, 2017; pp. 259–320. [Google Scholar]
- Sun, X.; Li, M.; Sun, Y.; Cai, H.; Lan, X.; Huang, Y.; Bai, Y.; Qi, X.; Chen, H. The developmental transcriptome sequencing of bovine skeletal muscle reveals a long noncoding RNA, lncMD, promotes muscle differentiation by sponging miR-125b. Biochim. Biophys. Acta 2016, 1863, 2835–2845. [Google Scholar] [CrossRef] [PubMed]
- Seiliez, I.; Froehlich, J.M.; Marandel, L.; Gabillard, J.-C.; Biga, P.R. Evolutionary history and epigenetic regulation of the three paralogous pax7 genes in rainbow trout. Cell Tissue Res. 2014, 359, 715–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koganti, P.; Yao, J.; Cleveland, B.M. Molecular Mechanisms Regulating Muscle Plasticity in Fish. Animal 2020, 11, 61. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-F.; Mandel, E.M.; Thomson, J.M.; Wu, Q.; Callis, T.E.; Hammond, S.M.; Conlon, F.L.; Wang, D.-Z. The Role of MicroRNA-1 and MicroRNA-133 in Skeletal Muscle Proliferation and Differentiation. Nat. Genet. 2005, 38, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-F.; Tao, Y.; Li, J.; Deng, Z.; Yan, Z.; Xiao, X.; Wang, D.-Z. microRNA-1 and microRNA-206 regulate skeletal muscle satellite cell proliferation and differentiation by repressing Pax7. J. Cell Biol. 2010, 190, 867–879. [Google Scholar] [CrossRef] [Green Version]
- Latimer, M.; Sabin, N.; Le Cam, A.; Seiliez, I.; Biga, P.; Gabillard, J.-C. miR-210 expression is associated with methionine-induced differentiation of trout satellite cells. J. Exp. Biol. 2017, 220, 2932–2938. [Google Scholar] [CrossRef] [Green Version]
- Picard, B.; Lebret, B.; Cassar-Malek, I.; Liaubet, L.; Berri, C.; Le Bihan-Duval, E.; Hocquette, J.-F.; Renand, G. Recent advances in omic technologies for meat quality management. Meat Sci. 2015, 109, 18–26. [Google Scholar] [CrossRef]
- Poleti, M.D.; Regitano, L.C.A.; Souza, G.H.M.F.; Cesar, A.S.M.; Simas, R.C.; Silva-Vignato, B.; Oliveira, G.B.; Andrade, S.C.S.; Cameron, L.C.; Coutinho, L.L.; et al. Longissimus dorsi muscle label-free quantitative proteomic reveals biological mechanisms associated with intramuscular fat deposition. J. Proteom. 2018, 179, 30–41. [Google Scholar] [CrossRef]
- Chen, L.; Li, Z.; Everaert, N.; Lametsch, R.; Zhang, D. Quantitative phosphoproteomic analysis of ovine muscle with different postmortem glycolytic rates. Food Chem. 2019, 280, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Veiseth-Kent, E.; de Almeida, A.M.; Picard, B.; Hollung, K. Proteomics in Skeletal Muscle Research. In Proteomics in Domestic Animals: From Farm to Systems Biology; Springer: Berlin/Heidelberg, Germany, 2018; pp. 195–217. [Google Scholar]
- Nienow, A.W. Reactor Engineering in Large Scale Animal Cell Culture. Cytotechnology 2006, 50, 9–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freed, L.E.; Guilak, F.; Guo, X.E.; Gray, M.L.; Tranquillo, R.; Holmes, J.W.; Radisic, M.; Sefton, M.V.; Kaplan, D.; Vunjak-Novakovic, G. Advanced Tools for Tissue Engineering: Scaffolds, Bioreactors, and Signaling. Tissue Eng. 2006, 12, 3285–3305. [Google Scholar] [CrossRef] [PubMed]
- Eibl, R.; Werner, S.; Eibl, D. Bag Bioreactor Based on Wave-Induced Motion: Characteristics and Applications. Invertebr. Cytokines Phylogeny Immun. 2009, 115, 55–87. [Google Scholar] [CrossRef]
- Allan, S.J.; De Bank, P.A.; Ellis, M.J. Bioprocess design considerations for cultured meat production with a focus on the expansion bioreactor. Front. Sustain. Food Syst. 2019, 3, 44. [Google Scholar] [CrossRef] [Green Version]
- Grayson, W.L.; Fröhlich, M.; Yeager, K.; Bhumiratana, S.; Chan, M.E.; Cannizzaro, C.; Wan, L.Q.; Liu, X.S.; Guo, X.E.; Vunjak-Novakovic, G. Engineering anatomically shaped human bone grafts. Proc. Natl. Acad. Sci. USA 2009, 107, 3299–3304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu-Absi, N.R.; Kenty, B.M.; Cuellar, M.E.; Borys, M.C.; Sakhamuri, S.; Strachan, D.J.; Hausladen, M.C.; Li, Z.J. Real time monitoring of multiple parameters in mammalian cell culture bioreactors using an in-line Raman spectroscopy probe. Biotechnol. Bioeng. 2011, 108, 1215–1221. [Google Scholar] [CrossRef] [PubMed]
- Bellani, C.F.; Ajeian, J.; Duffy, L.; Miotto, M.; Groenewegen, L.; Connon, C.J. Scale-Up Technologies for the Manufacture of Adherent Cells. Front. Nutr. 2020, 7, 178. [Google Scholar] [CrossRef]
- Shemesh, J.; Jalilian, I.; Shi, A.; Yeoh, G.H.; Tate, M.L.K.; Warkiani, M.E. Flow-induced stress on adherent cells in microfluidic devices. Lab A Chip 2015, 15, 4114–4127. [Google Scholar] [CrossRef]
- Fletcher, B.; Mullane, K.; Platts, P.; Todd, E.; Power, A.; Roberts, J.; Chapman, J.; Cozzolino, D.; Chandra, S. Advances in meat spoilage detection: A short focus on rapid methods and technologies. CyTA—J. Food 2018, 16, 1037–1044. [Google Scholar] [CrossRef] [Green Version]
- Daszczuk, A.; Dessalegne, Y.; Drenth, I.; Hendriks, E.; Jo, E.; van Lente, T.; Oldebesten, A.; Parrish, J.; Poljakova, W.; Purwanto, A.A.; et al. Bacillus subtilis biosensor engineered to assess meat spoilage. ACS Synth. Biol. 2014, 3, 999–1002. [Google Scholar] [CrossRef] [PubMed]
- Lohumi, S.; Lee, S.; Lee, H.; Cho, B.-K. A review of vibrational spectroscopic techniques for the detection of food authenticity and adulteration. Trends Food Sci. Technol. 2015, 46, 85–98. [Google Scholar] [CrossRef]
- Magnaghi, L.R.; Alberti, G.; Quadrelli, P.; Biesuz, R. Development of a Dye-Based Device to Assess Poultry Meat Spoilage. Part I: Building and Testing the Sensitive Array. J. Agric. Food Chem. 2020, 68, 12702–12709. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Gurr, P.A.; Qiao, G.G. An irreversible spoilage sensor for protein-based food. ACS Sens. 2020, 5, 2903–2908. [Google Scholar] [CrossRef]
- Balasubramanian, B.; Liu, W.; Pushparaj, K.; Park, S. The Epic of In Vitro Meat Production—A Fiction into Reality. Foods 2021, 10, 1395. [Google Scholar] [CrossRef]
- Bairoch, A. The Cellosaurus, a Cell-Line Knowledge Resource. J. Biomol. Tech. 2018, 29, 25–38. [Google Scholar] [CrossRef]
- Stachelscheid, H.; Seltmann, S.; Lekschas, F.; Fontaine, J.-F.; Mah, N.; Neves, M.; Andrade-Navarro, M.A.; Leser, U.; Kurtz, A. CellFinder: A cell data repository. Nucleic Acids Res. 2013, 42, D950–D958. [Google Scholar] [CrossRef] [Green Version]
- Goswami, M.; Nagpure, N.S.; Jena, J.K. Fish Cell Line Repository: An enduring effort for conservation. Curr. Sci. 2014, 107, 738–739. [Google Scholar]
- Coriell, L.L. Cell Repository. Science 1973, 180, 427. [Google Scholar] [CrossRef] [Green Version]
- Reiss, J.; Robertson, S.; Suzuki, M. Cell Sources for Cultivated Meat: Applications and Considerations throughout the Production Workflow. Int. J. Mol. Sci. 2021, 22, 7513. [Google Scholar] [CrossRef]
- Specht, L. An Analysis of Culture Medium Costs and Production Volumes for Cultivated Meat; The Good Food Institute: Washington, DC, USA, 2020; Available online: https://gfi.org/wp-content/uploads/2021/01/clean-meat-production-volume-and-medium-cost.pdf (accessed on 10 March 2022).
- Welin, S. Introducing the new meat. Problems and prospects. Etikk Praksis-Nord. J. Appl. Ethics 2013, 7, 24–37. [Google Scholar] [CrossRef] [Green Version]
- Liongue, C.; Ward, A.C. Evolution of Class I cytokine receptors. BMC Evol. Biol. 2007, 7, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopkins, P.D.; Dacey, A. Vegetarian Meat: Could Technology Save Animals and Satisfy Meat Eaters? J. Agric. Environ. Ethics 2008, 21, 579–596. [Google Scholar] [CrossRef]
- Tomiyama, A.J.; Kawecki, N.S.; Rosenfeld, D.L.; Jay, J.A.; Rajagopal, D.; Rowat, A.C. Bridging the gap between the science of cultured meat and public perceptions. Trends Food Sci. Technol. 2020, 104, 144–152. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| SL. No. | Cell Line | Species | Reference |
|---|---|---|---|
| 1 | CAM | Carassius auratus | [11] |
| 2 | CAM | Cromileptes altivelis | [12] |
| 3 | FHM | Pimephales promelas | [13] |
| 4 | MPCs | Danio rerio | [14] |
| 5 | TMF | Scophthalmus maximus | [15] |
| 6 | DRM | Danio rerio | [16] |
| 7 | MSCs | Paralichthys olivaceus | [17] |
| 8 | WAM | Wallagu attu | [18] |
| 9 | BM | Lates calcarifer | [19] |
| 10 | GFM | Carassius auratus | [20] |
| 11 | BTMS | Caranx melampygus | [21] |
| 12 | SHMS | Channa striatus | [22] |
| 13 | WSBM | Acipenser transmontanus | [23] |
| SL. No. | Company | Headquarters | Fish Species Considered for Cultivated Seafood Production |
|---|---|---|---|
| 1 | Another Fish | Montreal | Whitefish |
| 2 | Avant Meats | Hong Kong | Fish maw, sea cucumber, whitefish |
| 3 | Cell Ag Tech | Ontario, Canada | Whitefish |
| 4 | Bluefin Foods | Los Angeles | Bluefin tuna |
| 5 | BlueNalu | San Diego | Tuna, mahi mahi, red snapper |
| 6 | Bluu Biosciences | Berlin | Salmon, trout, carp |
| 7 | Cultured Decadence | Madison, Wisconsin, USA | Lobster |
| 8 | Finless Foods | Emeryville, California, USA | Bluefin tuna |
| 9 | Magic Caviar | Amsterdam | Caviar |
| 10 | Memphis Meats | Berkeley, California, USA | Coho salmon |
| 11 | Sea-Stematic | Johannesburg, South Africa | – |
| 12 | Shiok Meats | Singapore | Crab, lobster, shrimp |
| 13 | SoundEats | Seattle | Whitefish, zebrafish |
| 14 | Umami Meats | Singapore | Japanese eel, red snapper, grouper, yellowfin tuna |
| 15 | Wildtype | San Francisco | Salmon |
| Year | Development | Reference |
|---|---|---|
| 1912 | French biologist Alexis Carrel keeps a piece of chick heart muscle alive in a Petri dish, demonstrating the possibility of keeping muscle tissue alive outside of the body. | [38] |
| 1930 | Frederick Edwin Smith, 1st Earl of Birkenhead predicts that “It will no longer be necessary to go to the extravagant length of rearing a bullock to eat its steak. From one ‘parent’ steak of choice tenderness, it will be possible to grow as large and as juicy a steak as can be desired.” | [39] |
| 1932 | Winston Churchill writes “Fifty years hence we shall escape the absurdity of growing a whole chicken to eat the breast or wing by growing these parts separately under a suitable medium.” | [39] |
| The early 1950s | Willem van Eelen recognizes the possibility of generating meat from tissue culture. | [38] |
| 1971 | Russell Ross achieves the in vitro cultivation of muscular fibers. | [40] |
| 1995 | The U.S. Food and Drug Administration approves the use of commercial in vitro meat production. | [41] |
| 1999 | Willem van Eelen secures the first patent for cultured meat. | [38] |
| 2001 | NASA begins in vitro meat experiments, producing cultured turkey meat. | [42] |
| 2002 | Researchers culture muscle tissue of the common goldfish in Petri dishes. The meat was judged by a test-panel to be acceptable as food. | [38] |
| 2004 | Jason Matheny founds New Harvest, the first nonprofit to work for the development of cultured meat. | [39] |
| 2005 | Dutch government agency SenterNovem begins funding cultured meat research. | [43] |
| 2008 | The In Vitro Meat Consortium holds the first international conference on the production of in vitro meat. | [44] |
| 2008 | People for the Ethical Treatment of Animals offers a $1 million prize to the first group to make a commercially viable lab-grown chicken by 2012. | [41] |
| 2011 | The company Modern Meadow, aimed at producing cultured leather and meat, is founded. | [45] |
| 2013 | The first cultured hamburger, developed by Dutch researcher Mark Post’s lab, is taste-tested. | [46] |
| 2014 | Muufri and Clara Foods, companies aimed at producing cultured dairy and eggs, respectively, are founded with the assistance of New Harvest. | [43] |
| 2014 | Real Vegan Cheese, a startup aimed at creating cultured cheese, is founded. | [47] |
| 2014 | Modern Meadow presents “steak chips”, discs of lab-grown meat that could be produced at a relatively low cost. | [45] |
| 2015 | The Modern Agriculture Foundation, which focuses on developing cultured chicken meat (as chickens make up the large majority of land animals killed for food, is founded in Israel). | [48] |
| 2015 | According to Mark Post’s lab, the cost of producing a cultured hamburger patty drops from $325,000 in 2013 to less than $12 | [49] |
| 2016 | New Crop Capital, a private venture capital fund investing in alternatives to animal agriculture—including cellular agriculture—is founded. Its $25 million portfolio includes cultured meat company Memphis Meats and cultured collagen company Gelzen, along with Lighter, a software platform designed to facilitate plant-based eating, a plant-based meal delivery service called Purple Carrot, a dairy alternative called Lyrical Foods, the New Zealand plant-based meat company SunFed Foods, and alternative cheese company Miyoko’s Kitchen. | [50] |
| 2016 | The Good Food Institute, an organization devoted to promoting alternatives to animal food products—including cellular agriculture—is founded. | [51] |
| 2016 | Memphis Meats announces the creation of the first cultured meatball. | [52] |
| 2019 | Perfect Day (formerly Muufri) sells 1000 3-pint bundles of ice cream made with non-animal whey protein. | [53] |
| 2021 | Tufts University is awarded US$10 million by the USDA to establish the National Institute for Cellular Agriculture | [54] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goswami, M.; Belathur Shambhugowda, Y.; Sathiyanarayanan, A.; Pinto, N.; Duscher, A.; Ovissipour, R.; Lakra, W.S.; Chandragiri Nagarajarao, R. Cellular Aquaculture: Prospects and Challenges. Micromachines 2022, 13, 828. https://doi.org/10.3390/mi13060828
Goswami M, Belathur Shambhugowda Y, Sathiyanarayanan A, Pinto N, Duscher A, Ovissipour R, Lakra WS, Chandragiri Nagarajarao R. Cellular Aquaculture: Prospects and Challenges. Micromachines. 2022; 13(6):828. https://doi.org/10.3390/mi13060828
Chicago/Turabian StyleGoswami, Mukunda, Yashwanth Belathur Shambhugowda, Arjunan Sathiyanarayanan, Nevil Pinto, Alexandrea Duscher, Reza Ovissipour, Wazir Singh Lakra, and Ravishankar Chandragiri Nagarajarao. 2022. "Cellular Aquaculture: Prospects and Challenges" Micromachines 13, no. 6: 828. https://doi.org/10.3390/mi13060828
APA StyleGoswami, M., Belathur Shambhugowda, Y., Sathiyanarayanan, A., Pinto, N., Duscher, A., Ovissipour, R., Lakra, W. S., & Chandragiri Nagarajarao, R. (2022). Cellular Aquaculture: Prospects and Challenges. Micromachines, 13(6), 828. https://doi.org/10.3390/mi13060828