Clinical Significance of SERPINA1 Gene and Its Encoded Alpha1-antitrypsin Protein in NSCLC

 ,
, 

Abstract
:1. Introduction
2. Results
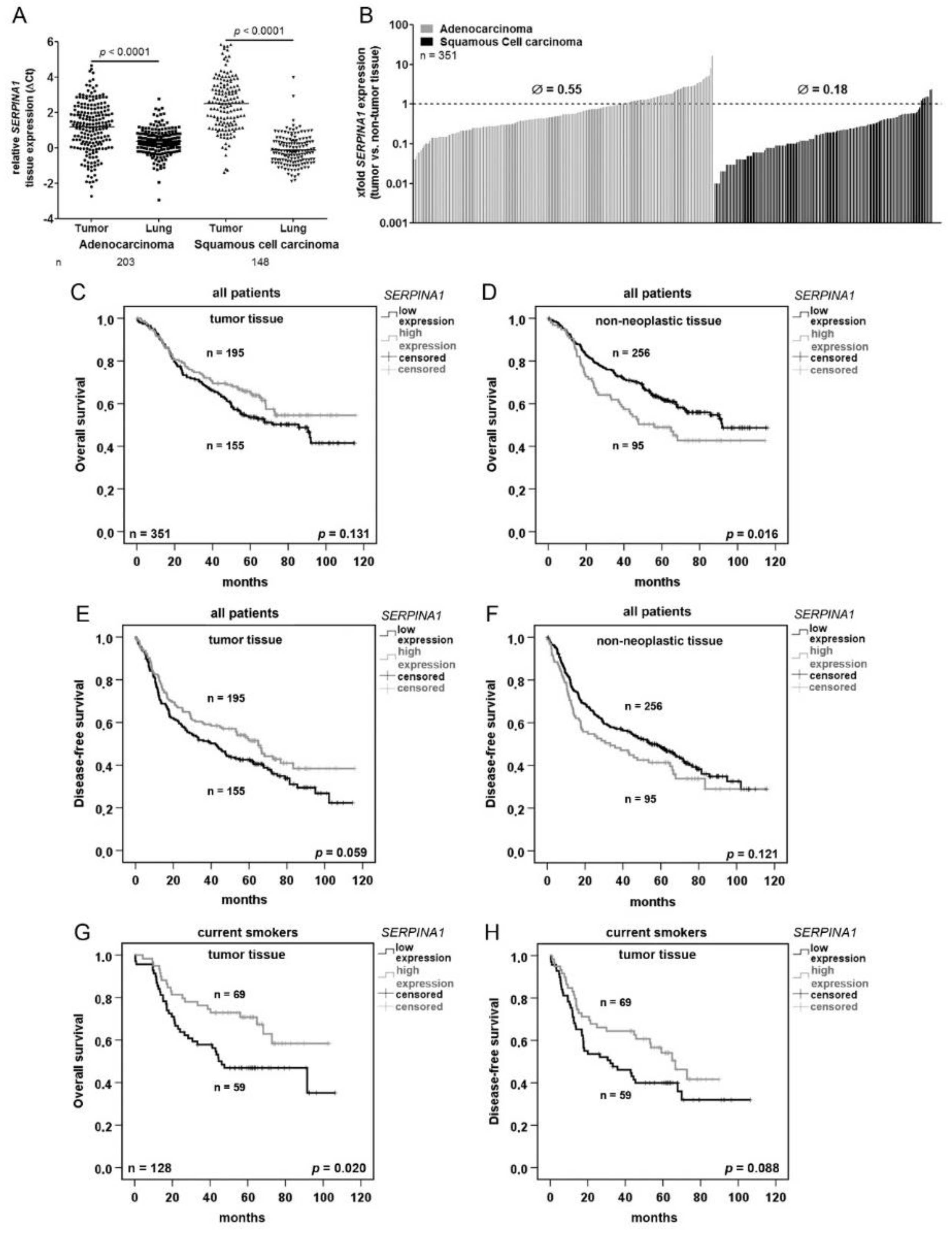
2.1. SERPINA1 Gene Expression Is Lower in NSCLC Tumors as Compared to the Normal Adjacent Lung Tissue and Prognostic for Patients’ Survival
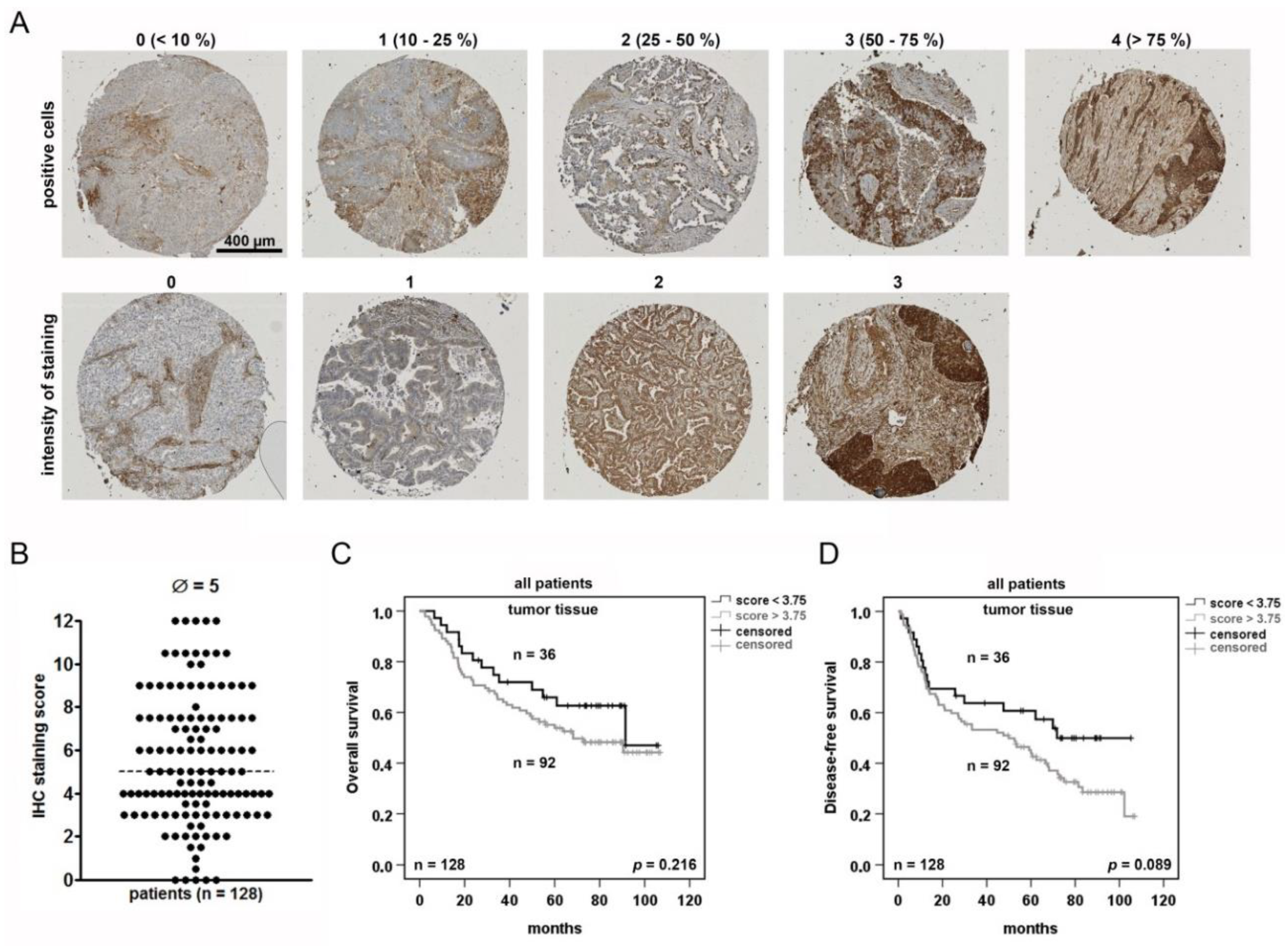
2.2. NSCLC Tumor Cells Are Heterogeneously Stained for AAT and Positive Tumor Cells May Serve as A Prognostic Marker
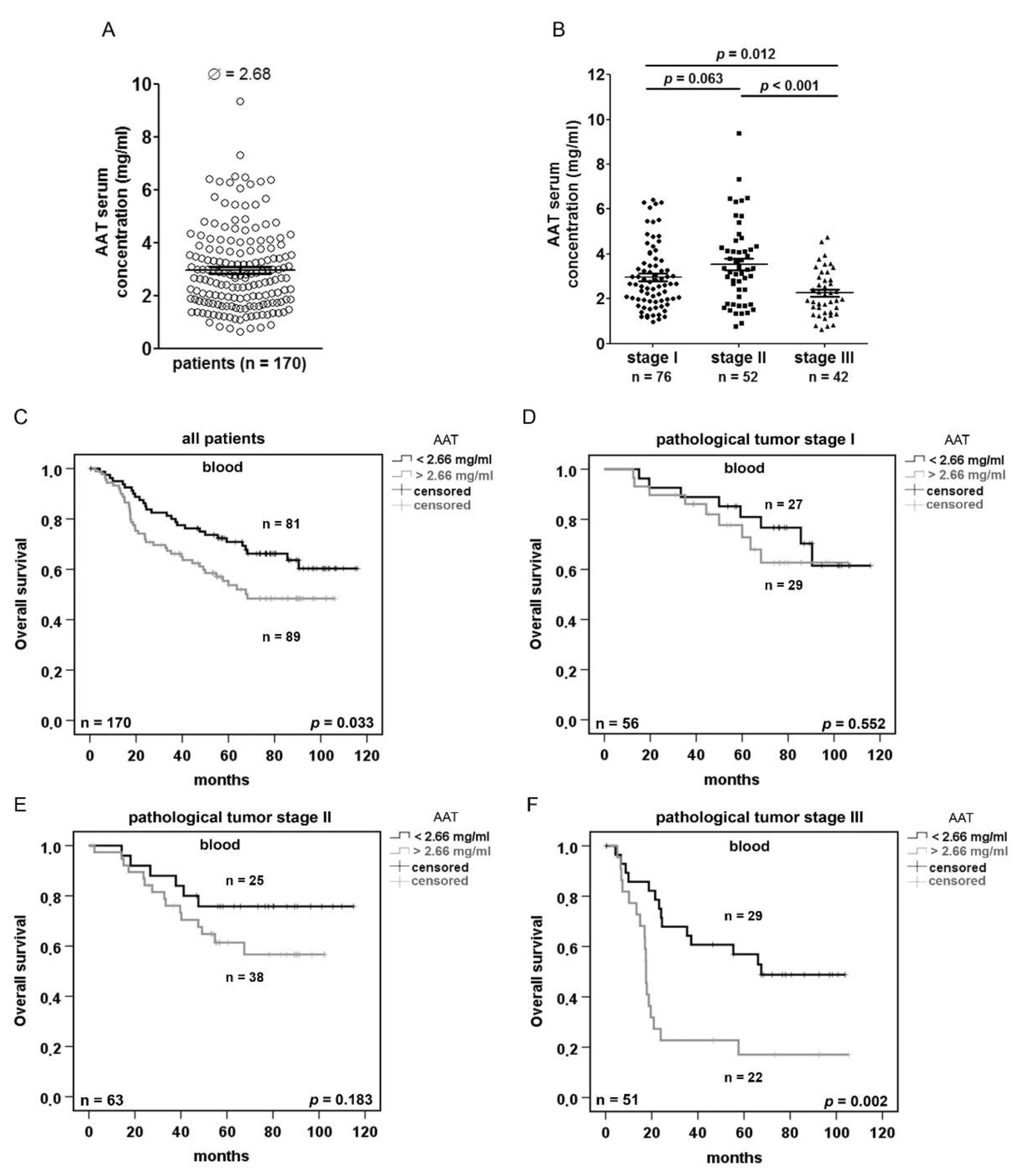
2.3. Serum AAT Concentration does not Correlate With SERPINA1 Expression and AAT IHC Staining but Is Prognostic Depending on the Disease Stage
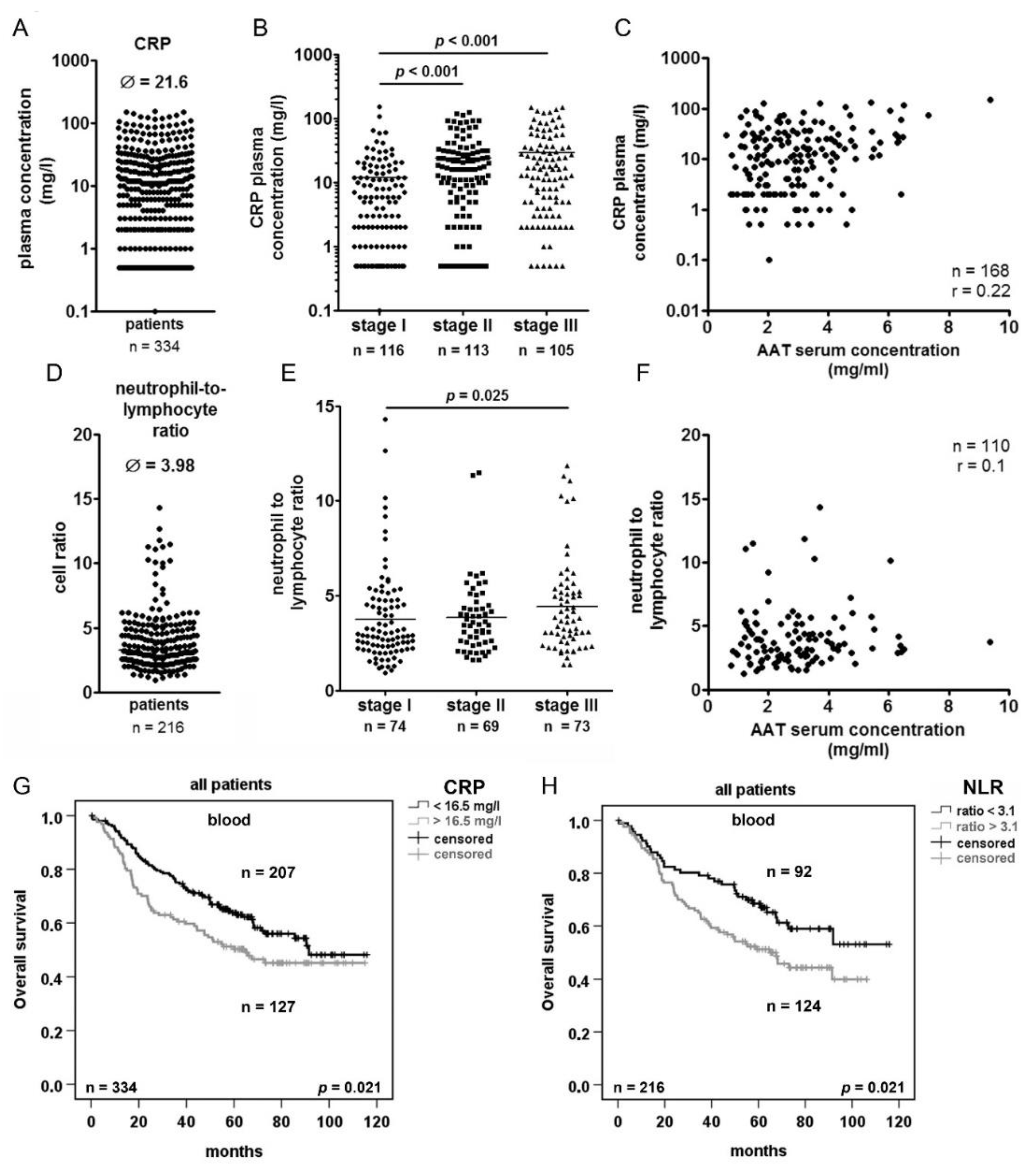
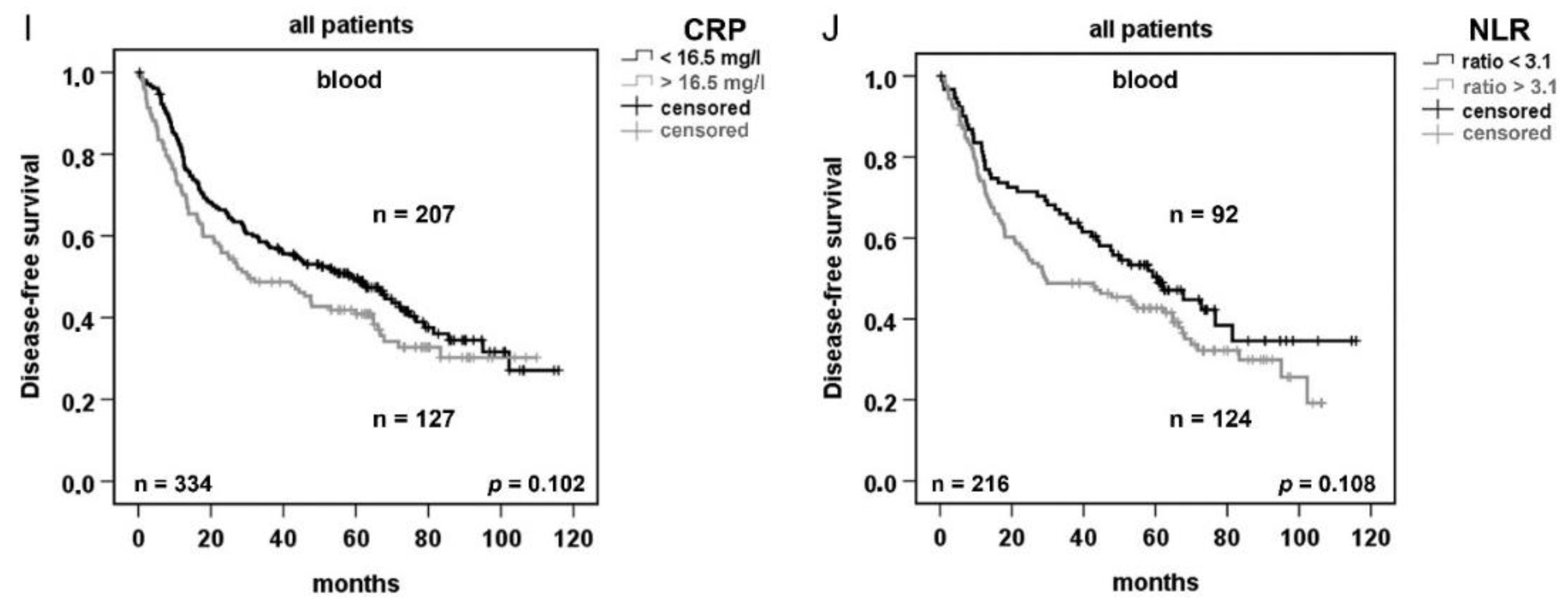
2.4. Serum Concentration of C-reactive Protein (CRP) and Neutrophils/Lymphocytes Ratio (NLR) Increase in Advanced Tumor Stage but do not Correlated with AAT Serum Concentration
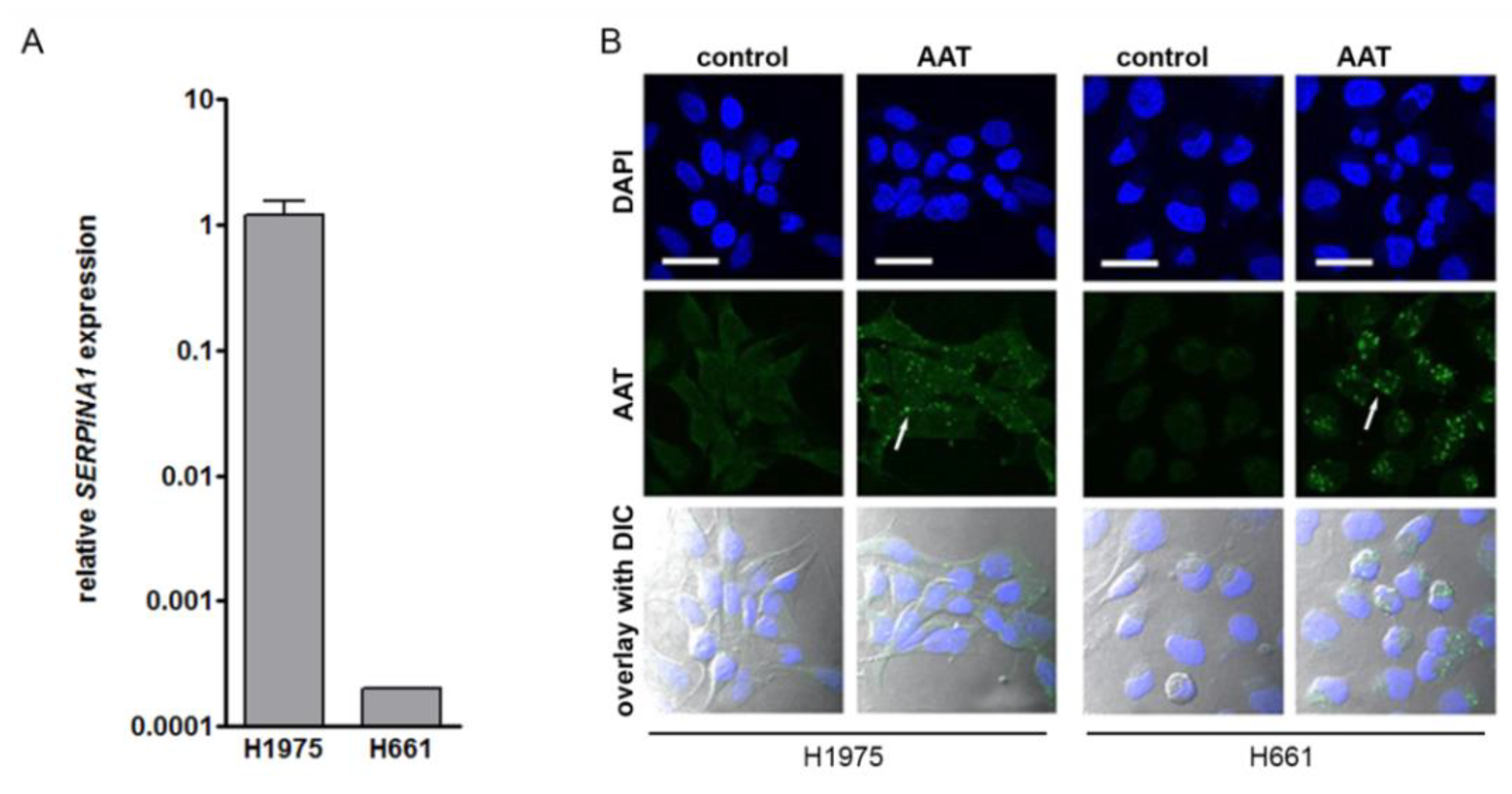
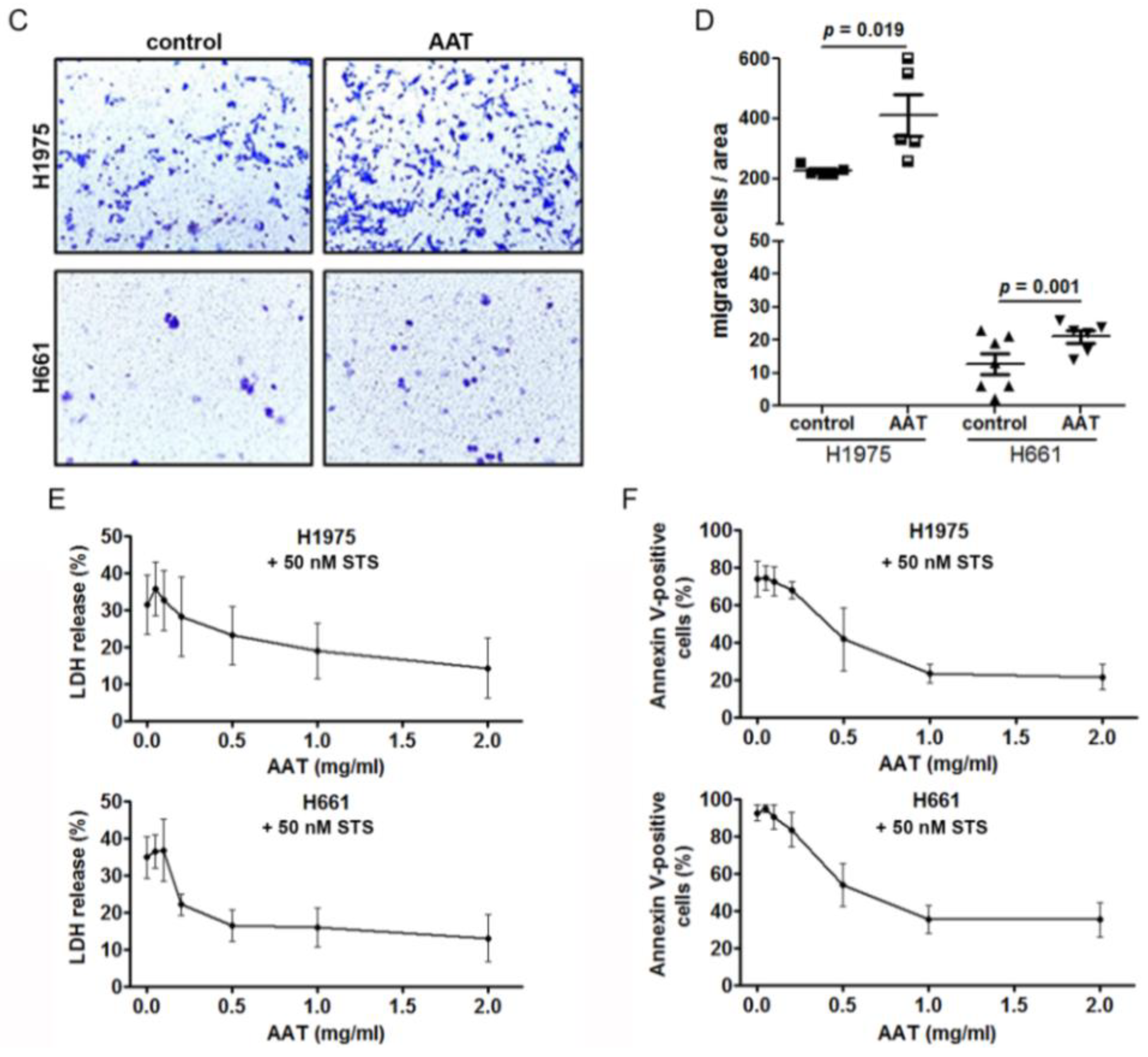
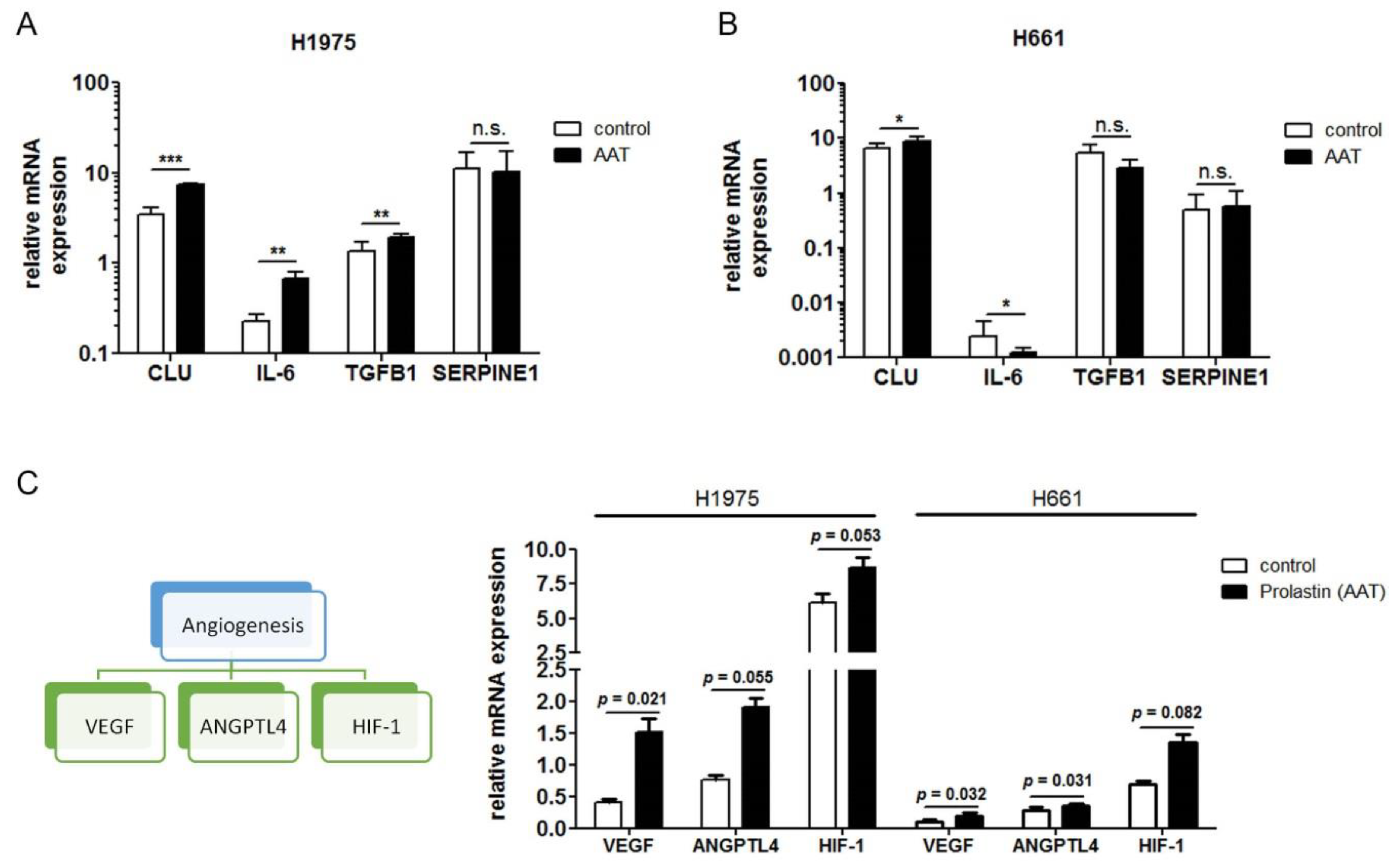
2.5. Exogenous AAT Is Taken Up by NSCLC Cells and Regulates Key Hallmarks of Cancer Cells
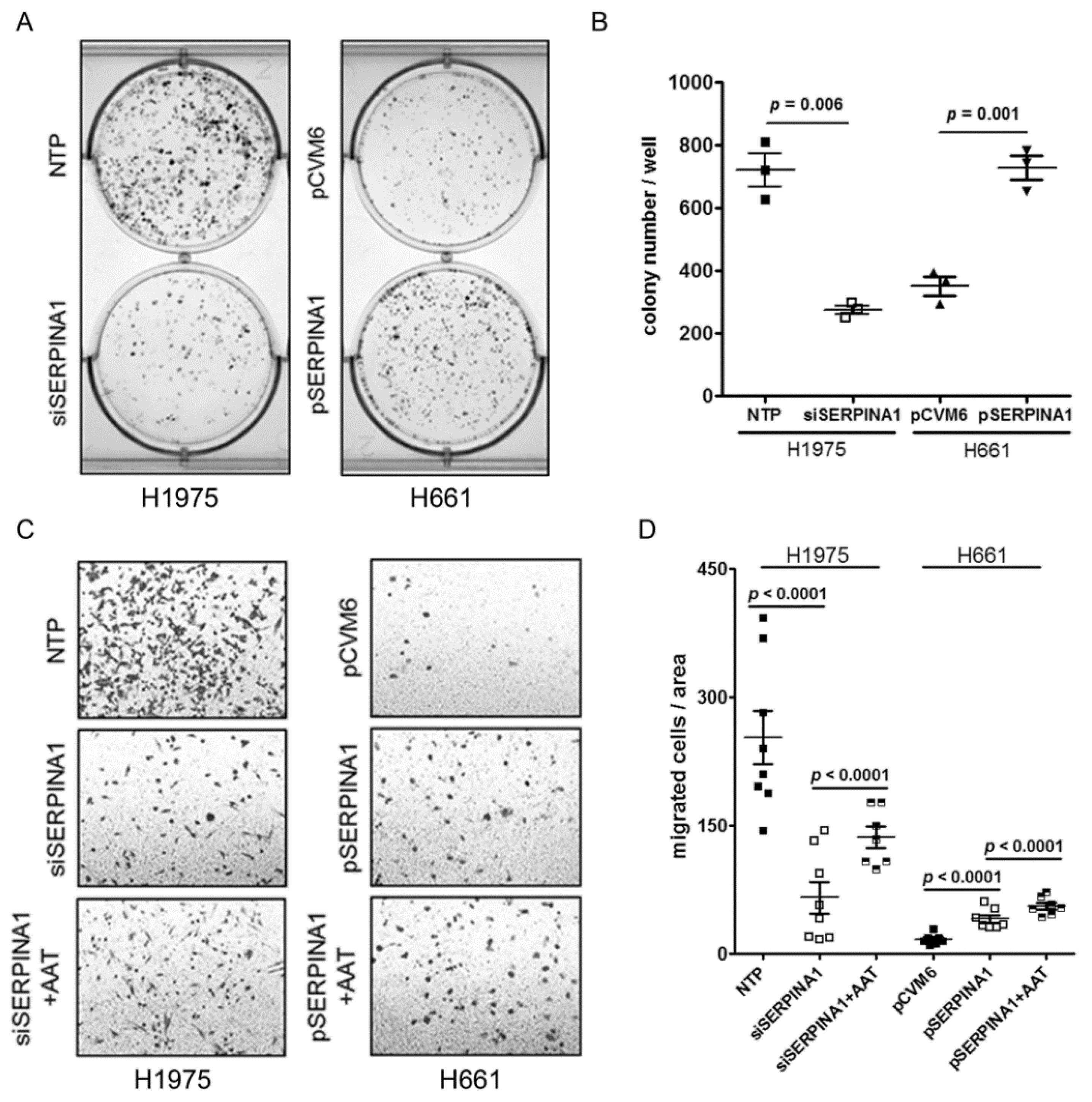
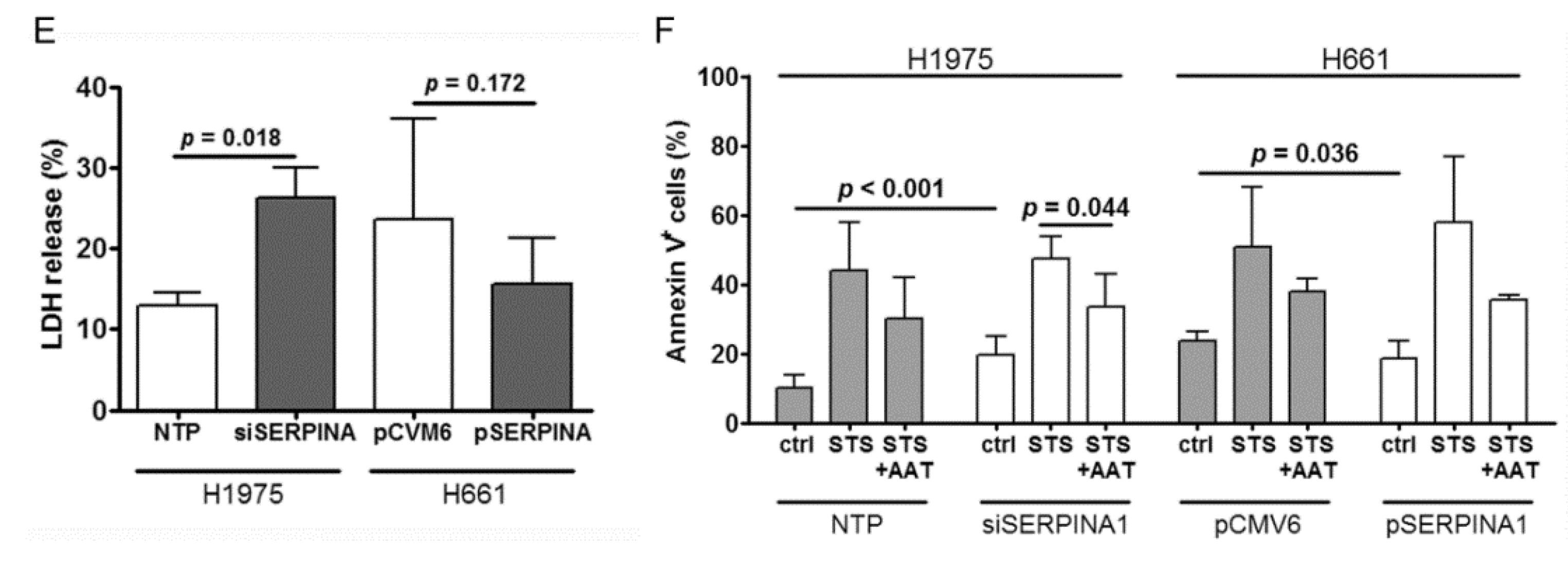
2.6. Silencing or Overexpression of SERPINA1 Regulates Colony Formation, Migration and Survival of NSCLC Cell Lines
2.7. Genes Most Related to SERPINA1 in H1975 and H661 Cells
3. Discussion
4. Materials and Methods
4.1. Sample Collection, Characterization and Preparation
4.2. Total RNA Isolation and cDNA Synthesis
4.3. Real-Time Polymerase Chain Reaction (RT-PCR) Analysis
4.4. Immunohistochemistry (IHC)
4.5. Analysis of Serum AAT Protein Concentration
4.6. CRP, Neutrophil and Lymphocyte Analysis
4.7. Cell Culture Experiments
4.8. SERPINA1 Gene Knockdown and Overexpression
4.9. Western Blots
5. Functional Cell Assays
5.1. Analysis of Intracellular AAT
5.2. Cell Migration
5.3. Lactate Dehydrogenase (LDH) Cytotoxicity Assay
5.4. Apoptosis Assay
5.5. Cancer Colony Forming Assay
5.6. RNA Sequencing (RNA-seq)
5.7. Analysis of RNA-Seq Data from H1975 and H661 Cell Lines
5.8. Statistical Analyses
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Devesa, S.S.; Bray, F.; Vizcaino, A.P.; Parkin, D.M. International lung cancer trends by histologic type: Male:Female differences diminishing and adenocarcinoma rates rising. Int. J. Cancer 2005, 117, 294–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldstraw, P.; Chansky, K.; Crowley, J.; Rami-Porta, R.; Asamura, H.; Eberhardt, W.E.; Nicholson, A.G.; Groome, P.; Mitchell, A.; Bolejack, V.; et al. The iaslc lung cancer staging project: Proposals for revision of the tnm stage groupings in the forthcoming (eighth) edition of the tnm classification for lung cancer. J. Thorac. Oncol. 2016, 11, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Roxburgh, C.S.; McMillan, D.C. Role of systemic inflammatory response in predicting survival in patients with primary operable cancer. Future Oncol. 2010, 6, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Qian, L.; Cui, J. Value of neutrophil-to-lymphocyte ratio for predicting lung cancer prognosis: A meta-analysis of 7219 patients. Mol. Clin. Oncol. 2017, 7, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Ha, M.; Yin, N. Combination of platelet count and lymphocyte to monocyte ratio is a prognostic factor in patients undergoing surgery for non-small cell lung cancer. Oncotarget 2017, 8, 73198–73207. [Google Scholar] [CrossRef] [PubMed]
- Siemes, C.; Visser, L.E.; Coebergh, J.W.; Splinter, T.A.; Witteman, J.C.; Uitterlinden, A.G.; Hofman, A.; Pols, H.A.; Stricker, B.H. C-reactive protein levels, variation in the c-reactive protein gene, and cancer risk: The rotterdam study. J. Clin. Oncol. 2006, 24, 5216–5222. [Google Scholar] [CrossRef]
- Hara, M.; Matsuzaki, Y.; Shimuzu, T.; Tomita, M.; Ayabe, T.; Enomoto, Y.; Onitsuka, T. Preoperative serum c-reactive protein level in non-small cell lung cancer. Anticancer Res. 2007, 27, 3001–3004. [Google Scholar]
- Jones, J.M.; McGonigle, N.C.; McAnespie, M.; Cran, G.W.; Graham, A.N. Plasma fibrinogen and serum c-reactive protein are associated with non-small cell lung cancer. Lung Cancer 2006, 53, 97–101. [Google Scholar] [CrossRef]
- Lee, J.G.; Cho, B.C.; Bae, M.K.; Lee, C.Y.; Park, I.K.; Kim, D.J.; Ahn, S.V.; Chung, K.Y. Preoperative c-reactive protein levels are associated with tumor size and lymphovascular invasion in resected non-small cell lung cancer. Lung Cancer 2009, 63, 106–110. [Google Scholar] [CrossRef]
- Tveitaras, M.K.; Selheim, F.; Sortland, K.; Reed, R.K.; Stuhr, L. Protein expression profiling of plasma and lungs at different stages of metastatic development in a human triple negative breast cancer xenograft model. PLoS ONE 2019, 14, e0215909. [Google Scholar] [CrossRef] [PubMed]
- Boccellino, M.; Pinto, F.; Ieluzzi, V.; Giovane, A.; Quagliuolo, L.; Fariello, C.; Coppola, M.; Carlucci, A.; Santini, M.; Ferati, K.; et al. Proteomics analysis of human serum of patients with non-small-cell lung cancer reveals proteins as diagnostic biomarker candidates. J. Cell. Physiol. 2019, 234, 23798–23806. [Google Scholar] [CrossRef] [PubMed]
- Janciauskiene, S.M.; Bals, R.; Koczulla, R.; Vogelmeier, C.; Kohnlein, T.; Welte, T. The discovery of alpha1-antitrypsin and its role in health and disease. Respir. Med. 2011, 105, 1129–1139. [Google Scholar] [CrossRef] [PubMed]
- Carlson, J.A.; Rogers, B.B.; Sifers, R.N.; Hawkins, H.K.; Finegold, M.J.; Woo, S.L. Multiple tissues express alpha 1-antitrypsin in transgenic mice and man. J. Clin. Investig. 1988, 82, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Janciauskiene, S.; Wrenger, S.; Immenschuh, S.; Olejnicka, B.; Greulich, T.; Welte, T.; Chorostowska-Wynimko, J. The multifaceted effects of alpha1-antitrypsin on neutrophil functions. Front. Pharmacol. 2018, 9, 341. [Google Scholar] [CrossRef] [PubMed]
- El-Akawi, Z.J.; Abu-Awad, A.M.; Sharara, A.M.; Khader, Y. The importance of alpha-1 antitrypsin (alpha1-at) and neopterin serum levels in the evaluation of non-small cell lung and prostate cancer patients. Neuro Endocrinol. Lett. 2010, 31, 113–116. [Google Scholar]
- Zhao, Z.; Ma, J.; Mao, Y.; Dong, L.; Li, S.; Zhang, Y. Silence of alpha1-antitrypsin inhibits migration and proliferation of triple negative breast cancer cells. Med Sci. Monit. 2018, 24, 6851–6860. [Google Scholar] [CrossRef] [PubMed]
- Daddi, G.; Mancini, P.A.; Parola, D.; Contini, A. Alfa-antitrypsin increase in lung cancer. Boll. dell’Istituto Sieroter. Milan. 1976, 55, 510–512. [Google Scholar]
- Zelvyte, I.; Wallmark, A.; Piitulainen, E.; Westin, U.; Janciauskiene, S. Increased plasma levels of serine proteinase inhibitors in lung cancer patients. Anticancer Res. 2004, 24, 241–247. [Google Scholar]
- Higashiyama, M.; Doi, O.; Kodama, K.; Yokouchi, H.; Tateishi, R. An evaluation of the prognostic significance of alpha-1-antitrypsin expression in adenocarcinomas of the lung: An immunohistochemical analysis. Br. J. Cancer 1992, 65, 300–302. [Google Scholar] [CrossRef]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. Gepia: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [PubMed]
- Thompson, D.; Pepys, M.B.; Wood, S.P. The physiological structure of human c-reactive protein and its complex with phosphocholine. Structure 1999, 7, 169–177. [Google Scholar] [CrossRef]
- Diem, S.; Schmid, S.; Krapf, M.; Flatz, L.; Born, D.; Jochum, W.; Templeton, A.J.; Fruh, M. Neutrophil-to-lymphocyte ratio (nlr) and platelet-to-lymphocyte ratio (plr) as prognostic markers in patients with non-small cell lung cancer (nsclc) treated with nivolumab. Lung Cancer 2017, 111, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.H.; Lee, S.H.; Liao, I.C.; Huang, S.H.; Cheng, H.C.; Liao, P.C. Secretomic analysis identifies alpha-1 antitrypsin (a1at) as a required protein in cancer cell migration, invasion, and pericellular fibronectin assembly for facilitating lung colonization of lung adenocarcinoma cells. Mol. Cell. Proteom. 2012, 11, 1320–1339. [Google Scholar] [CrossRef] [PubMed]
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Harris, C.C.; Cohen, M.H.; Connor, R.; Primack, A.; Saccomanno, G.; Talamo, R.C. Serum alpha1-antitrypsin in patients with lung cancer or abnormal sputum cytology. Cancer 1976, 38, 1655–1657. [Google Scholar] [CrossRef]
- Guergova-Kuras, M.; Kurucz, I.; Hempel, W.; Tardieu, N.; Kadas, J.; Malderez-Bloes, C.; Jullien, A.; Kieffer, Y.; Hincapie, M.; Guttman, A.; et al. Discovery of lung cancer biomarkers by profiling the plasma proteome with monoclonal antibody libraries. Mol. Cell. Proteom. 2011, 10, 010298. [Google Scholar] [CrossRef] [PubMed]
- Senn, O.; Russi, E.W.; Schindler, C.; Imboden, M.; von Eckardstein, A.; Brandli, O.; Zemp, E.; Ackermann-Liebrich, U.; Berger, W.; Rochat, T.; et al. Circulating alpha1-antitrypsin in the general population: Determinants and association with lung function. Respir. Res. 2008, 9, 35. [Google Scholar] [CrossRef]
- Tang, W.; Zhou, M.; Dorsey, T.H.; Prieto, D.A.; Wang, X.W.; Ruppin, E.; Veenstra, T.D.; Ambs, S. Integrated proteotranscriptomics of breast cancer reveals globally increased protein-mrna concordance associated with subtypes and survival. Genome Med. 2018, 10, 94. [Google Scholar] [CrossRef]
- Roumeliotis, T.I.; Williams, S.P.; Goncalves, E.; Alsinet, C.; Del Castillo Velasco-Herrera, M.; Aben, N.; Ghavidel, F.Z.; Michaut, M.; Schubert, M.; Price, S.; et al. Genomic determinants of protein abundance variation in colorectal cancer cells. Cell Rep. 2017, 20, 2201–2214. [Google Scholar] [CrossRef]
- Lackey, L.; McArthur, E.; Laederach, A. Increased transcript complexity in genes associated with chronic obstructive pulmonary disease. PLoS ONE 2015, 10, e0140885. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, E.; Fagerberg, L.; Klevebring, D.; Matic, I.; Geiger, T.; Cox, J.; Algenas, C.; Lundeberg, J.; Mann, M.; Uhlen, M. Defining the transcriptome and proteome in three functionally different human cell lines. Mol. Syst. Biol. 2010, 6, 450. [Google Scholar] [CrossRef] [PubMed]
- Ruoslahti, E. Specialization of tumour vasculature. Nat. Rev. Cancer 2002, 2, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Bottaro, D.P.; Liotta, L.A. Cancer: Out of air is not out of action. Nature 2003, 423, 593–595. [Google Scholar] [CrossRef] [PubMed]
- Greenhough, A.; Bagley, C.; Heesom, K.J.; Gurevich, D.B.; Gay, D.; Bond, M.; Collard, T.J.; Paraskeva, C.; Martin, P.; Sansom, O.J.; et al. Cancer cell adaptation to hypoxia involves a hif-gprc5a-yap axis. EMBO Mol. Med. 2018, 10, 8699. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Babapoor-Farrokhran, S.; Rodrigues, M.; Deshpande, M.; Puchner, B.; Kashiwabuchi, F.; Hassan, S.J.; Asnaghi, L.; Handa, J.T.; Merbs, S.; et al. Hypoxia-inducible factor 1 upregulation of both vegf and angptl4 is required to promote the angiogenic phenotype in uveal melanoma. Oncotarget 2016, 7, 7816–7828. [Google Scholar] [CrossRef]
- Frenzel, E.; Wrenger, S.; Brugger, B.; Salipalli, S.; Immenschuh, S.; Aggarwal, N.; Lichtinghagen, R.; Mahadeva, R.; Marcondes, A.M.; Dinarello, C.A.; et al. Alpha1-antitrypsin combines with plasma fatty acids and induces angiopoietin-like protein 4 expression. J. Immunol. 2015, 195, 3605–3616. [Google Scholar] [CrossRef]
- Frenzel, E.; Wrenger, S.; Immenschuh, S.; Koczulla, R.; Mahadeva, R.; Deeg, H.J.; Dinarello, C.A.; Welte, T.; Marcondes, A.M.; Janciauskiene, S. Acute-phase protein alpha1-antitrypsin—A novel regulator of angiopoietin-like protein 4 transcription and secretion. J. Immunol. 2014, 192, 5354–5362. [Google Scholar] [CrossRef]
- Fleixo-Lima, G.; Ventura, H.; Medini, M.; Bar, L.; Strauss, P.; Lewis, E.C. Mechanistic evidence in support of alpha1-antitrypsin as a therapeutic approach for type 1 diabetes. J. Diabetes Sci. Technol. 2014, 8, 1193–1203. [Google Scholar] [CrossRef]
- Chan, H.J.; Li, H.; Liu, Z.; Yuan, Y.C.; Mortimer, J.; Chen, S. Serpina1 is a direct estrogen receptor target gene and a predictor of survival in breast cancer patients. Oncotarget 2015, 6, 25815–25827. [Google Scholar] [CrossRef]
- Shakya, R.; Tarulli, G.A.; Sheng, L.; Lokman, N.A.; Ricciardelli, C.; Pishas, K.I.; Selinger, C.I.; Kohonen-Corish, M.R.J.; Cooper, W.A.; Turner, A.G.; et al. Mutant p53 upregulates alpha-1 antitrypsin expression and promotes invasion in lung cancer. Oncogene 2017, 36, 4469–4480. [Google Scholar] [CrossRef] [PubMed]
- Crystal, R.G. The alpha 1-antitrypsin gene and its deficiency states. Trends Genet. 1989, 5, 411–417. [Google Scholar] [CrossRef]
- Davalieva, K.; Kiprijanovska, S.; Komina, S.; Petrusevska, G.; Zografska, N.C.; Polenakovic, M. Proteomics analysis of urine reveals acute phase response proteins as candidate diagnostic biomarkers for prostate cancer. Proteome Sci. 2015, 13, 2. [Google Scholar] [CrossRef] [PubMed]
- Grivennikov, S.I.; Greten, F.R.; Karin, M. Immunity, inflammation, and cancer. Cell 2010, 140, 883–899. [Google Scholar] [CrossRef] [PubMed]
- Balkwill, F.; Mantovani, A. Inflammation and cancer: Back to virchow? Lancet 2001, 357, 539–545. [Google Scholar] [CrossRef]
- Hara, M.; Yonei, A.; Ayabe, T.; Tomita, M.; Nakamura, K.; Onitsuka, T. Postoperative serum c-reactive protein levels in non-small cell lung cancer patients. Ann. Thorac. Cardiovasc. Surg. 2010, 16, 85–90. [Google Scholar] [PubMed]
- Binnewies, M.; Roberts, E.W.; Kersten, K.; Chan, V.; Fearon, D.F.; Merad, M.; Coussens, L.M.; Gabrilovich, D.I.; Ostrand-Rosenberg, S.; Hedrick, C.C.; et al. Understanding the tumor immune microenvironment (time) for effective therapy. Nat. Med. 2018, 24, 541–550. [Google Scholar] [CrossRef]
- Zhou, X.; Liu, Z.; Shapiro, L.; Yang, J.; Burton, G.F. Low-density lipoprotein receptor-related protein 1 mediates alpha1-antitrypsin internalization in cd4+ t lymphocytes. J. Leukoc. Biol. 2015, 98, 1027–1035. [Google Scholar] [CrossRef]
- Sohrab, S.; Petrusca, D.N.; Lockett, A.D.; Schweitzer, K.S.; Rush, N.I.; Gu, Y.; Kamocki, K.; Garrison, J.; Petrache, I. Mechanism of alpha-1 antitrypsin endocytosis by lung endothelium. FASEB J. 2009, 23, 3149–3158. [Google Scholar] [CrossRef]
- Aldonyte, R.; Tunaitis, V.; Surovas, A.; Suriakaite, K.; Jarmalaviciute, A.; Magnusson, K.E.; Pivoriunas, A. Effects of major human antiprotease alpha-1-antitrypsin on the motility and proliferation of stromal cells from human exfoliated deciduous teeth. Regen. Med. 2010, 5, 633–643. [Google Scholar] [CrossRef]
- Aggarwal, N.; Korenbaum, E.; Mahadeva, R.; Immenschuh, S.; Grau, V.; Dinarello, C.A.; Welte, T.; Janciauskiene, S. Alpha-linoleic acid enhances the capacity of alpha-1 antitrypsin to inhibit lipopolysaccharide induced il-1beta in human blood neutrophils. Mol. Med. 2016, 22, 680–693. [Google Scholar] [CrossRef] [PubMed]
- Kwon, C.H.; Park, H.J.; Lee, J.R.; Kim, H.K.; Jeon, T.Y.; Jo, H.J.; Kim, D.H.; Kim, G.H.; Park, D.Y. Serpin peptidase inhibitor clade a member 1 is a biomarker of poor prognosis in gastric cancer. Br. J. Cancer 2014, 111, 1993–2002. [Google Scholar] [CrossRef] [PubMed]
- Kwon, C.H.; Park, H.J.; Choi, J.H.; Lee, J.R.; Kim, H.K.; Jo, H.J.; Kim, H.S.; Oh, N.; Song, G.A.; Park, D.Y. Snail and serpina1 promote tumor progression and predict prognosis in colorectal cancer. Oncotarget 2015, 6, 20312–20326. [Google Scholar] [CrossRef] [PubMed]
- Normandin, K.; Peant, B.; Le Page, C.; de Ladurantaye, M.; Ouellet, V.; Tonin, P.N.; Provencher, D.M.; Mes-Masson, A.M. Protease inhibitor serpina1 expression in epithelial ovarian cancer. Clin. Exp. Metastasis 2010, 27, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Miao, L.; Yu, M.; Shi, M.; Wang, Y.; Yang, J.; Xiao, Y.; Cai, H. Alpha1-antitrypsin promotes lung adenocarcinoma metastasis through upregulating fibronectin expression. Int. J. Oncol. 2017, 50, 1955–1964. [Google Scholar] [CrossRef]
- Sun, Z.Y.P. Role of imbalance between neutrophil elastase and alpha 1-antitrypsin in cancer development and progression. Lancet Oncol. 2004, 5, 182–190. [Google Scholar] [CrossRef]
- Ehlers, M.R. Immune-modulating effects of alpha-1 antitrypsin. Biol. Chem. 2014, 395, 1187–1193. [Google Scholar] [CrossRef]
- Chang, S.H.; Cho, K.C.; Yu, K.N.; Hong, S.H.; Park, S.; Lee, A.Y.; Kim, S.; Lee, S.; Kang, J.W.; Chae, C.; et al. Alpha 1-antitrypsin activates lung cancer cell survival by acting on cap-dependent protein translation, vesicle-mediated transport, and metastasis. Oncotarget 2016, 19, 5. [Google Scholar] [CrossRef]
- Bergin, D.A.; Hurley, K.; McElvaney, N.G.; Reeves, E.P. Alpha-1 antitrypsin: A potent anti-inflammatory and potential novel therapeutic agent. Arch. Immunol. Ther. Exp. 2012, 60, 81–97. [Google Scholar] [CrossRef]
- Laine, A.; Leroy, A.; Hachulla, E.; Davril, M.; Dessaint, J.P. Comparison of the effects of purified human alpha 1-antichymotrypsin and alpha 1-proteinase inhibitor on nk cytotoxicity: Only alpha 1-proteinase inhibitor inhibits natural killing. Clin. Chim. Acta 1990, 190, 163–173. [Google Scholar] [CrossRef]
- Richtmann, S.; Wilkens, D.; Warth, A.; Lasitschka, F.; Winter, H.; Christopoulos, P.; Herth, F.J.F.; Muley, T.; Meister, M.; Schneider, M.A. Fam83a and fam83b as prognostic biomarkers and potential new therapeutic targets in nsclc. Cancers 2019, 11, 652. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.A.; Granzow, M.; Warth, A.; Schnabel, P.A.; Thomas, M.; Herth, F.J.; Dienemann, H.; Muley, T.; Meister, M. Glycodelin: A new biomarker with immunomodulatory functions in non-small cell lung cancer. Clin. Cancer Res. 2015, 21, 3529–3540. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The miqe guidelines: Minimum information for publication of quantitative real-time pcr experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. Star: Ultrafast universal rna-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Pyl, P.T.; Huber, W. Htseq—A python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Meyer, C.A.; Wang, Q.; Liu, J.S.; Shirley Liu, X.; Zhang, Y. Gfold: A generalized fold change for ranking differentially expressed genes from rna-seq data. Bioinformatics 2012, 28, 2782–2788. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.; Muruganujan, A.; Ebert, D.; Huang, X.; Thomas, P.D. Panther version 14: More genomes, a new panther go-slim and improvements in enrichment analysis tools. Nucleic Acids Res. 2019, 47, D419–D426. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. JSTOR 1995, 57, 289–300. [Google Scholar] [CrossRef]
- McShane, L.M.; Altman, D.G.; Sauerbrei, W.; Taube, S.E.; Gion, M.; Clark, G.M. Reporting recommendations for tumor marker prognostic studies (remark). Breast Cancer Res. Treat. 2006, 100, 229–235. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cohort Description | ||
|---|---|---|
| Parameter | n | (%) |
| Median Age | 65 (38–88) | |
| Total | 351 | 100 |
| Male | 243 | 69 |
| Female | 108 | 31 |
| Histology | ||
| Adeno | 205 | 58 |
| Squamous | 146 | 42 |
| Therapy | ||
| OP | 205 | 58 |
| OP/RT | 13 | 4 |
| OP/ChT | 97 | 27 |
| OP/RT/ChT | 36 | 10 |
| Smoking Status | ||
| Non-Smoker | 37 | 11 |
| Ex-Smoker | 184 | 52 |
| Smoker | 128 | 36 |
| No data | 2 | 1 |
| Pathological Stage (7th TNM Edition) | ||
| IA | 37 | 11 |
| IB | 123 | 35 |
| IIA | 24 | 7 |
| IIB | 74 | 21 |
| IIIA | 83 | 24 |
| IIIB | 10 | 3 |
| ECOG | ||
| 0 | 301 | 86 |
| 1 | 46 | 13 |
| 2 | 4 | 1 |
| BMI | ||
| <18.5 | 9 | 3 |
| 18.5 - <25 | 138 | 39 |
| 25 - <30 | 122 | 35 |
| >30 | 81 | 23 |
| No data | 1 | 0 |
| Univariate Analysis (Gene Expression, Overall Survival) | ||
| Variable (high vs. low) | Significance | Hazard Ratio (95% CI) |
| SERPINA1 (tumor) | 0.132 | 0.781 (0.567–1.077) |
| SERPINA1 (non-tumor) | 0.017 | 1.508 (1.077–2.112) |
| SERPINA1 (tumor vs. non-tumor) | 0.495 | 1.036 (0.935–1.149) |
| Multivariate Analysis (Gene Expression, Overall Survival) | ||
| Variable | Significance | Hazard Ratio (95% CI) |
| SERPINA1 (tumor, high vs. low) | 0.117 | 0.752 (0.526–1.074) |
| SERPINA1 (non-tumor, high vs. low) | 0.028 | 1.540 (1.047–2.265) |
| SERPINA1 (tumor vs. non-tumor, high vs. low) | 0.316 | 1.050 (0.954–1.155) |
| Sex (female vs. male) | 0.105 | 0.727 (0.495–1.069) |
| pathological stage (7th edition) | <0.0001 | 1.069 (1.046–1.093) |
| Histology (ADC vs. SQCC) | 0.389 | 1.175 (0.814–1.697) |
| age | <0.0001 | 1.038 (1.018–1.057) |
| Univariate Analysis (IHC, Overall Survival) | ||
| Variable | Significance | Hazard Ratio (95% CI) |
| AAT (high vs. low) | 0.219 | 1.454 (0.800–2.641) |
| Univariate Analysis (Blood, Overall Survival) | ||
| Variable | Significance | Hazard Ratio (95% CI) |
| AAT (high vs. low) | 0.035 | 1.674 (1.036–2.704) |
| Multivariate Analysis (Blood, Overall Survival) | ||
| Variable | Significance | Hazard Ratio (95% CI) |
| AAT (high vs. low) | 0.009 | 1.938 (1.176–3.194) |
| Sex (female vs. male) | 0.259 | 0.725 (0.415–1.267) |
| pathological stage (7th edition) | <0.0001 | 2.211 (1.552–3.149) |
| Histology (ADC vs. SQCC) | 0.731 | 0.918 (0.564–1.494) |
| age | 0.220 | 1.017 (0.990–1.045) |
| H1975 | |||
| Gene Symbol | Description to the Gene | Regulation Direction | Annotated GO Terms in the Analysis |
| APOE | Apolipoprotein E | Upregulated | inflammatory response, regulation of inflammatory response |
| APP | Amyloid Beta Precursor Protein | Upregulated | inflammatory response, regulation of inflammatory response, positive regulation of apoptotic process, regulation of inflammatory response |
| C3 | Complement C3 | Upregulated | inflammatory response, regulation of inflammatory response |
| CLU | Clusterin | Upregulated | apoptotic process, positive regulation of apoptotic process |
| CTSC | Cathepsin C | Upregulated | apoptotic process, positive regulation of apoptotic process |
| CYR61 | Cysteine-rich angiogenic inducer 61 | Upregulated | apoptotic process, positive regulation of apoptotic process |
| EVA1A | Eva-1 Homolog A, Regulator Of Programmed Cell Death | Upregulated | apoptotic process, positive regulation of apoptotic process |
| IGFBP3 | Insulin Like Growth Factor Binding Protein 3 | Upregulated | apoptotic process, positive regulation of apoptotic process |
| IL6 | Interleukin 6 | Upregulated | inflammatory response, apoptotic process, positive regulation of apoptotic process, regulation of inflammatory response |
| LGALS1 | Galectin 1 | Upregulated | apoptotic process, positive regulation of apoptotic process |
| PCSK9 | Proprotein Convertase Subtilisin/Kexin Type 9 | Downregulated | apoptotic process, positive regulation of apoptotic process |
| PROC | Protein C, Inactivator Of Coagulation Factors Va And VIIIa | Upregulated | inflammatory response, regulation of inflammatory response |
| SERPINE1 | Serpin Family E Member 1 | Upregulated | inflammatory response, regulation of inflammatory response |
| SHISA5 | Shisa Family Member 5 | Upregulated | apoptotic process, positive regulation of apoptotic process |
| TGFB1 | Transforming Growth Factor Beta 1 | Upregulated | apoptotic process, positive regulation of apoptotic process |
| TGFB2 | Transforming Growth Factor Beta 2 | Upregulated | apoptotic process, positive regulation of apoptotic process |
| TMSB4X | Thymosin Beta 4 X-Linked | Upregulated | inflammatory response, regulation of inflammatory response |
| H661 | |||
| Gene Symbol | Description to the Gene | Regulation Direction | Annotated GO Terms in the Analysis |
| APOE | Apolipoprotein E | Upregulated | inflammatory response, regulation of inflammatory response |
| APP | Amyloid Beta Precursor Protein | Upregulated | inflammatory response, regulation of inflammatory response |
| CLU | Clusterin | Upregulated | cell population proliferation, apoptotic process |
| CYR61 | Cysteine-rich angiogenic inducer 61 | Downregulated | apoptotic process |
| IGF1 | Insulin Like Growth Factor 1 | Upregulated | inflammatory response, cell population proliferation |
| LGALS1 | Galectin 1 | Upregulated | apoptotic process |
| SERPINE1 | Serpin Family E Member 1 | Upregulated | inflammatory response, regulation of inflammatory response |
| SHISA5 | Shisa Family Member 5 | Upregulated | apoptotic process |
| TGFB1 | Transforming Growth Factor Beta 1 | Downregulated | cell population proliferation, apoptotic process |
| TGFB2 | Transforming Growth Factor Beta 2 | Downregulated | cell population proliferation, apoptotic process |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ercetin, E.; Richtmann, S.; Delgado, B.M.; Gomez-Mariano, G.; Wrenger, S.; Korenbaum, E.; Liu, B.; DeLuca, D.; Kühnel, M.P.; Jonigk, D.; et al. Clinical Significance of SERPINA1 Gene and Its Encoded Alpha1-antitrypsin Protein in NSCLC. Cancers 2019, 11, 1306. https://doi.org/10.3390/cancers11091306
Ercetin E, Richtmann S, Delgado BM, Gomez-Mariano G, Wrenger S, Korenbaum E, Liu B, DeLuca D, Kühnel MP, Jonigk D, et al. Clinical Significance of SERPINA1 Gene and Its Encoded Alpha1-antitrypsin Protein in NSCLC. Cancers. 2019; 11(9):1306. https://doi.org/10.3390/cancers11091306
Chicago/Turabian StyleErcetin, Evrim, Sarah Richtmann, Beatriz Martinez Delgado, Gema Gomez-Mariano, Sabine Wrenger, Elena Korenbaum, Bin Liu, David DeLuca, Mark P. Kühnel, Danny Jonigk, and et al. 2019. "Clinical Significance of SERPINA1 Gene and Its Encoded Alpha1-antitrypsin Protein in NSCLC" Cancers 11, no. 9: 1306. https://doi.org/10.3390/cancers11091306
APA StyleErcetin, E., Richtmann, S., Delgado, B. M., Gomez-Mariano, G., Wrenger, S., Korenbaum, E., Liu, B., DeLuca, D., Kühnel, M. P., Jonigk, D., Yuskaeva, K., Warth, A., Muley, T., Winter, H., Meister, M., Welte, T., Janciauskiene, S., & Schneider, M. A. (2019). Clinical Significance of SERPINA1 Gene and Its Encoded Alpha1-antitrypsin Protein in NSCLC. Cancers, 11(9), 1306. https://doi.org/10.3390/cancers11091306