Transport-Mediated Oxaliplatin Resistance Associated with Endogenous Overexpression of MRP2 in Caco-2 and PANC-1 Cells

 ,
, 

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Caco-2 and PANC-1 Cells Endogenously Overexpress MRP2
2.2. Reduced MRP2 Expression in ABCC2-siRNA-Transfected Cells
2.3. ABCC2-siRNA Sensitized GI Cells to Oxaliplatin Growth Inhibition
2.4. ABCC2-siRNA Increased CDCF and Platinum Accumulation in GI Cancer Cells
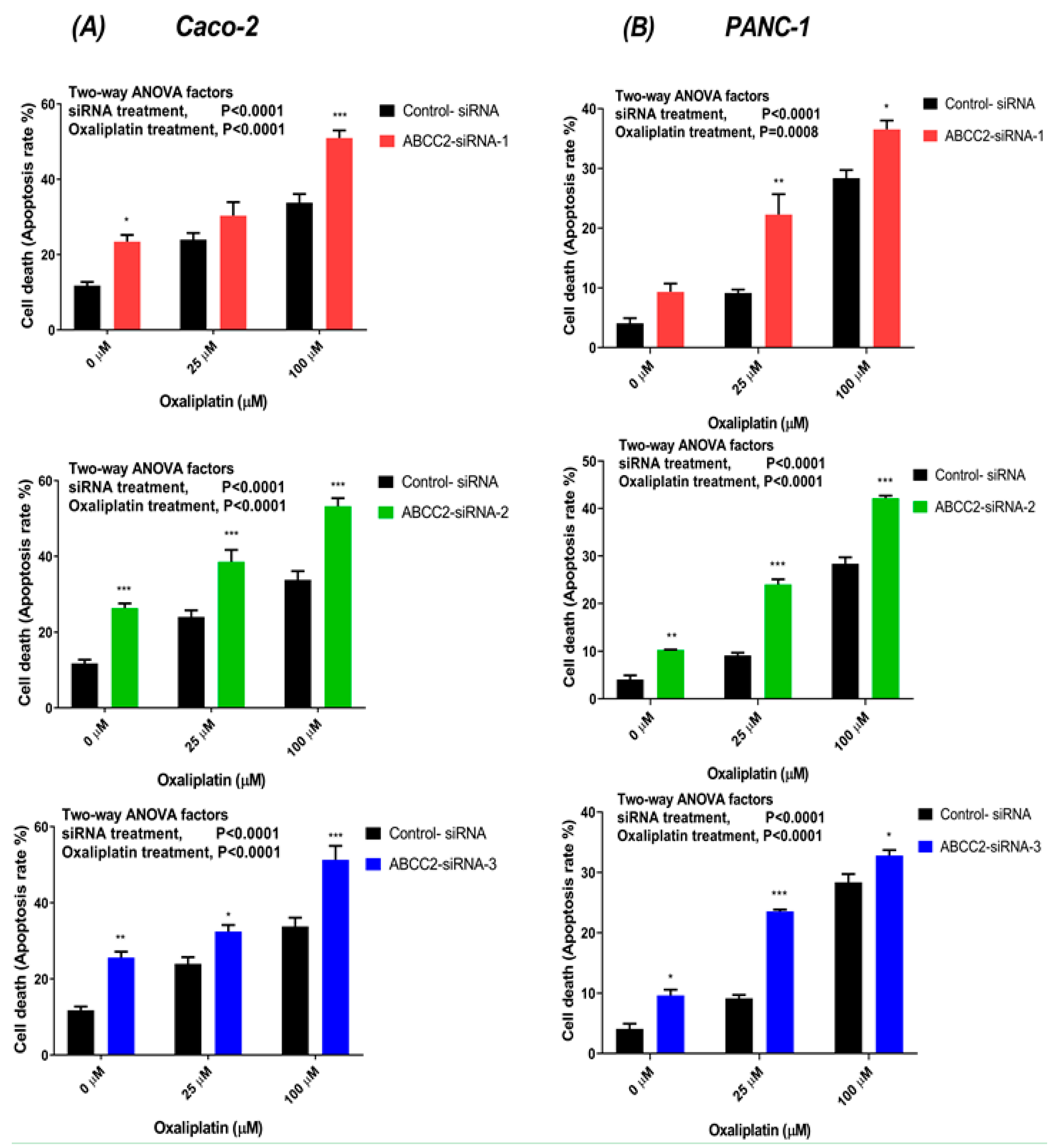
2.5. ABCC2-siRNA Increased Oxaliplatin-Induced Apoptosis Rate
2.6. Effect of Myricetin on Oxaliplatin Accumulation and Growth Inhibition
2.7. Oxaliplatin Stimulated the ATPase Activity of MRP2
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. ATPase Assay
4.4. siRNA Transfection
- a. ABCC2-siRNA-1:
- 5’-GAU CAU GAA UGA GAU UCU UAG UGG A-3’5’-UCC ACU AAG AAU CUC AUU CAU GAU C-3’
- b. ABCC2-siRNA-2:
- 5’-CCA GCA AAG GCA AGA UCC AGU UUA A-3’5’-UUA AAC UGG AUC UUG CCU UUG CUG G-3’
- c. ABCC2-siRNA-3:
- 5’-ACC AAG ACA UUA GUG AGC AAG UUU G-3’5’-CAA ACU UGC UCA CUA AUG UCU UGG U-3’
4.5. Expression of Transporter Genes
4.6. MRP2 Surface Staining
4.7. MRP2 Substrate Accumulation
4.8. Platinum Accumulation
4.9. Cytotoxicity Assay
4.10. Annexin-V-FITC Apoptosis Assay
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Andre, T.; Boni, C.; Mounedji-Boudiaf, L.; Navarro, M.; Tabernero, J.; Hickish, T.; Topham, C.; Zaninelli, M.; Clingan, P.; Bridgewater, J.; et al. Oxaliplatin, fluorouracil, and leucovorin as adjuvant treatment for colon cancer. N. Engl. J. Med. 2004, 350, 2343–2351. [Google Scholar] [CrossRef] [PubMed]
- Ryan, D.P.; Hong, T.S.; Bardeesy, N. Pancreatic adenocarcinoma. N. Engl. J. Med. 2014, 371, 2140–2141. [Google Scholar] [CrossRef] [PubMed]
- ESMO Clinical Practice Guidelines: Gastrointestinal Cancers. 2017. Available online: https://www.esmo.org/Guidelines/Gastrointestinal-Cancers (accessed on 2 September 2019).
- Suker, M.; Beumer, B.R.; Sadot, E.; Marthey, L.; Faris, J.E.; Mellon, E.A.; El-Rayes, B.F.; Wang-Gillam, A.; Lacy, J.; Hosein, P.J.; et al. FOLFIRINOX for locally advanced pancreatic cancer: a systematic review and patient-level meta-analysis. Lancet Oncol. 2016, 17, 801–810. [Google Scholar] [CrossRef] [Green Version]
- Hector, S.; Bolanowska-Higdon, W.; Zdanowicz, J.; Hitt, S.; Pendyala, L. In vitro studies on the mechanisms of oxaliplatin resistance. Cancer Chemother. Pharmacol. 2001, 48, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Martelli, L.; Di Mario, F.; Ragazzi, E.; Apostoli, P.; Leone, R.; Perego, P.; Fumagalli, G. Different accumulation of cisplatin, oxaliplatin and JM216 in sensitive and cisplatin-resistant human cervical tumour cells. Biochem. Pharmacol. 2006, 72, 693–700. [Google Scholar] [CrossRef]
- Chen, C.C.; Chen, L.T.; Tsou, T.C.; Pan, W.Y.; Kuo, C.C.; Liu, J.F.; Yeh, S.C.; Tsai, F.Y.; Hsieh, H.P.; Chang, J.Y. Combined modalities of resistance in an oxaliplatin-resistant human gastric cancer cell line with enhanced sensitivity to 5-fluorouracil. Br. J. Cancer 2007, 97, 334–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.J.; Kim, Y.; Yan, F.; Ding, Q.; Ip, V.; Jong, N.N.; Mercer, J.F.; McKeage, M.J. Contributions of rat Ctr1 to the uptake and toxicity of copper and platinum anticancer drugs in dorsal root ganglion neurons. Biochem. Pharmacol. 2013, 85, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Hall, M.D.; Okabe, M.; Shen, D.W.; Liang, X.J.; Gottesman, M.M. The role of cellular accumulation in determining sensitivity to platinum-based chemotherapy. Annu. Rev. Pharmacol. Toxicol. 2008, 48, 495–535. [Google Scholar] [CrossRef]
- Sprowl, J.A.; Ness, R.A.; Sparreboom, A. Polymorphic transporters and platinum pharmacodynamics. Drug Metab. Pharmacokinet. 2013, 28, 19–27. [Google Scholar] [CrossRef]
- Burger, H.; Loos, W.J.; Eechoute, K.; Verweij, J.; Mathijssen, R.H.; Wiemer, E.A. Drug transporters of platinum-based anticancer agents and their clinical significance. Drug Resist. Updat. 2011, 14, 22–34. [Google Scholar] [CrossRef]
- Jong, N.N.; Nakanishi, T.; Liu, J.J.; Tamai, I.; McKeage, M.J. Oxaliplatin transport mediated by organic cation/carnitine transporters OCTN1 and OCTN2 in overexpressing human embryonic kidney 293 cells and rat dorsal root ganglion neurons. J. Pharmacol. Exp. Ther. 2011, 338, 537–547. [Google Scholar] [CrossRef]
- Yonezawa, A.; Masuda, S.; Yokoo, S.; Katsura, T.; Inui, K. Cisplatin and oxaliplatin, but not carboplatin and nedaplatin, are substrates for human organic cation transporters (SLC22A1-3 and multidrug and toxin extrusion family). J. Pharmacol. Exp. Ther. 2006, 319, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Lovejoy, K.S.; Shima, J.E.; Lagpacan, L.L.; Shu, Y.; Lapuk, A.; Chen, Y.; Komori, T.; Gray, J.W.; Chen, X.; et al. Organic cation transporters are determinants of oxaliplatin cytotoxicity. Cancer Res. 2006, 66, 8847–8857. [Google Scholar] [CrossRef] [PubMed]
- Myint, K.; Li, Y.; Paxton, J.; McKeage, M. Multidrug resistance-associated protein 2 (MRP2) mediated transport of oxaliplatin-derived platinum in membrane vesicles. PLoS ONE 2015, 10, e0130727. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Liu, G.; Fang, T.; Wang, Y.; Zhang, H.; Yang, S.; Wei, J.; Lv, Z.; Tan, L.; Liu, J. Silencing MRP1-4 genes by RNA interference enhances sensitivity of human hepatoma cells to chemotherapy. Am. J. Transl. Res. 2016, 8, 2790–2802. [Google Scholar]
- Shen, K.; Cui, D.; Sun, L.; Lu, Y.; Han, M.; Liu, J. Inhibition of IGF-IR increases chemosensitivity in human colorectal cancer cells through MRP-2 promoter suppression. J. Cell. Biochem. 2012, 113, 2086–2097. [Google Scholar] [CrossRef]
- Wang, Z.; Sun, X.; Feng, Y.; Liu, X.; Zhou, L.; Sui, H.; Ji, Q.; E, Q.; Chen, J.; Wu, L.; et al. Dihydromyricetin reverses MRP2-mediated MDR and enhances anticancer activity induced by oxaliplatin in colorectal cancer cells. Anticancer Drugs 2017, 28, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Mohn, C.; Kalayda, G.; Häcker, H.; Gütschow, M.; Metzger, S.; Jaehde, U. Contribution of glutathione and MRP-mediated efflux to intracellular oxaliplatin accumulation. Int. J. Clin. Pharmacol. Ther. 2010, 48, 445. [Google Scholar] [CrossRef] [PubMed]
- Myint, K.; Biswas, R.; Li, Y.; Jong, N.; Jamieson, S.; Liu, J.; Han, C.; Squire, C.; Merien, F.; Lu, J.; et al. Identification of MRP2 as a targetable factor limiting oxaliplatin accumulation and response in gastrointestinal cancer. Sci. Rep. 2019, 9, 2245. [Google Scholar] [CrossRef]
- Cassidy, J.; Tabernero, J.; Twelves, C.; Brunet, R.; Butts, C.; Conroy, T.; Debraud, F.; Figer, A.; Grossmann, J.; Sawada, N.; et al. XELOX (capecitabine plus oxaliplatin): Active first-line therapy for patients with metastatic colorectal cancer. J. Clin. Oncol. 2004, 22, 2084–2091. [Google Scholar] [CrossRef]
- de Gramont, A.; Figer, A.; Seymour, M.; Homerin, M.; Hmissi, A.; Cassidy, J.; Boni, C.; Cortes-Funes, H.; Cervantes, A.; Freyer, G.; et al. Leucovorin and fluorouracil with or without oxaliplatin as first-line treatment in advanced colorectal cancer. J. Clin. Oncol. 2000, 18, 2938–2947. [Google Scholar] [CrossRef] [PubMed]
- Conroy, T.; Desseigne, F.; Ychou, M.; Bouche, O.; Guimbaud, R.; Becouarn, Y.; Adenis, A.; Raoul, J.L.; Gourgou-Bourgade, S.; de la Fouchardiere, C.; et al. FOLFIRINOX versus gemcitabine for metastatic pancreatic cancer. N. Engl. J. Med. 2011, 364, 1817–1825. [Google Scholar] [CrossRef] [PubMed]
- Oettle, H.; Riess, H.; Stieler, J.M.; Heil, G.; Schwaner, I.; Seraphin, J.; Gorner, M.; Molle, M.; Greten, T.F.; Lakner, V.; et al. Second-line oxaliplatin, folinic acid, and fluorouracil versus folinic acid and fluorouracil alone for gemcitabine-refractory pancreatic cancer: outcomes from the CONKO-003 trial. J. Clin. Oncol. 2014, 32, 2423–2429. [Google Scholar] [CrossRef] [PubMed]
- Al-Batran, S.E.; Hartmann, J.T.; Probst, S.; Schmalenberg, H.; Hollerbach, S.; Hofheinz, R.; Rethwisch, V.; Seipelt, G.; Homann, N.; Wilhelm, G.; et al. Phase III trial in metastatic gastroesophageal adenocarcinoma with fluorouracil, leucovorin plus either oxaliplatin or cisplatin: A study of the arbeitsgemeinschaft internistische onkologie. J. Clin. Oncol. 2008, 26, 1435–1442. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, D.; Starling, N.; Rao, S.; Iveson, T.; Nicolson, M.; Coxon, F.; Middleton, G.; Daniel, F.; Oates, J.; Norman, A.R. Capecitabine and Oxaliplatin for Advanced Esophagogastric Cancer. N. Engl. J. Med. 2008, 358, 36–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, S.; Bai, Y.; Lim, H.Y.; Thongprasert, S.; Chao, Y.; Fan, J.; Yang, T.-S.; Bhudhisawasdi, V.; Kang, W.K.; Zhou, Y.; et al. Randomized, multicenter, open-label study of oxaliplatin plus fluorouracil/leucovorin versus doxorubicin as palliative chemotherapy in patients with advanced hepatocellular carcinoma from asia. J. Clin. Oncol. 2013, 31, 3501–3508. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Mori, Y.; Hayashi, R.; Takada, M.; Ino, Y.; Naishiro, Y.; Kondo, T.; Hirohashi, S. Suppression of intestinal polyposis in Mdr1-deficient ApcMin/+ mice. Cancer Res. 2003, 63, 895–901. [Google Scholar]
- Smyth, M.J.; Krasovskis, E.; Sutton, V.R.; Johnstone, R.W. The drug efflux protein, P-glycoprotein, additionally protects drug-resistant tumor cells from multiple forms of caspase-dependent apoptosis. Proc. Natl. Acad. Sci. USA 1998, 95, 7024–7029. [Google Scholar] [CrossRef] [Green Version]
- Johnstone, R.W.; Cretney, E.; Smyth, M.J. P-glycoprotein protects leukemia cells against caspase-dependent, but not caspase-independent, cell death. Blood 1999, 93, 1075–1085. [Google Scholar]
- Peaston, A.E.; Gardaneh, M.; Franco, A.V.; Hocker, J.E.; Murphy, K.M.; Farnsworth, M.L.; Catchpoole, D.R.; Haber, M.; Norris, M.D.; Lock, R.B.; et al. MRP1 gene expression level regulates the death and differentiation response of neuroblastoma cells. Br. J. Cancer 2001, 85, 1564–1571. [Google Scholar] [CrossRef]
- Kuss, B.J.; Corbo, M.; Lau, W.M.; Fennell, D.A.; Dean, N.M.; Cotter, F.E. In vitro and in vivo downregulation of MRP1 by antisense oligonucleotides: a potential role in neuroblastoma therapy. Int. J. Cancer 2002, 98, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Agullo, G.; Gamet-Payrastre, L.; Manenti, S.; Viala, C.; Rémésy, C.; Chap, H.; Payrastre, B. Relationship between flavonoid structure and inhibition of phosphatidylinositol 3-kinase: A comparison with tyrosine kinase and protein kinase C inhibition. Biochem. Pharmacol. 1997, 53, 1649–1657. [Google Scholar] [CrossRef]
- Sprowl, J.A.; Ong, S.S.; Gibson, A.A.; Hu, S.; Du, G.; Lin, W.; Li, L.; Bharill, S.; Ness, R.A.; Stecula, A.; et al. A phosphotyrosine switch regulates organic cation transporters. Nat. Commun. 2016, 7, 10880. [Google Scholar] [CrossRef] [PubMed]
- Muller, J.; Lips, K.S.; Metzner, L.; Neubert, R.H.; Koepsell, H.; Brandsch, M. Drug specificity and intestinal membrane localization of human organic cation transporters (OCT). Biochem. Pharmacol. 2005, 70, 1851–1860. [Google Scholar] [CrossRef] [PubMed]
- Sarkadi, B.; Price, E.M.; Boucher, R.C.; Germann, U.A.; Scarborough, G.A. Expression of the human multidrug resistance cDNA in insect cells generates a high activity drug-stimulated membrane ATPase. J. Biol. Chem. 1992, 267, 4854–4858. [Google Scholar] [PubMed]
- Heredi-Szabo, K.; Kis, E.; Molnar, E.; Gyorfi, A.; Krajcsi, P. Characterization of 5(6)-carboxy-2,’7’-dichlorofluorescein transport by MRP2 and utilization of this substrate as a fluorescent surrogate for LTC4. J. Biomol. Screen. 2008, 13, 295–301. [Google Scholar] [CrossRef]
- Pedersen, J.M.; Matsson, P.; Bergstrom, C.A.; Norinder, U.; Hoogstraate, J.; Artursson, P. Prediction and identification of drug interactions with the human ATP-binding cassette transporter multidrug-resistance associated protein 2 (MRP2; ABCC2). J. Med. Chem. 2008, 51, 3275–3287. [Google Scholar] [CrossRef]
- Choudhuri, S.; Klaassen, C.D. Structure, function, expression, genomic organization, and single nucleotide polymorphisms of human ABCB1 (MDR1), ABCC (MRP), and ABCG2 (BCRP) efflux transporters. Int. J. Toxicol. 2006, 25, 231–259. [Google Scholar] [CrossRef]
- Graham, M.A.; Lockwood, G.F.; Greenslade, D.; Brienza, S.; Bayssas, M.; Gamelin, E. Clinical pharmacokinetics of oxaliplatin: a critical review. Clin. Cancer Res. 2000, 6, 1205–1218. [Google Scholar]
- Evers, R.; De Haas, M.; Sparidans, R.; Beijnen, J.; Wielinga, P.; Lankelma, J.; Borst, P. Vinblastine and sulfinpyrazone export by the multidrug resistance protein MRP2 is associated with glutathione export. Br. J. Cancer 2000, 83, 375–383. [Google Scholar] [CrossRef] [Green Version]
- Zelcer, N.; Huisman, M.T.; Reid, G.; Wielinga, P.; Breedveld, P.; Kuil, A.; Knipscheer, P.; Schellens, J.H.; Schinkel, A.H.; Borst, P. Evidence for two interacting ligand binding sites in human multidrug resistance protein 2 (ATP binding cassette C2). J. Biol. Chem. 2003, 278, 23538–23544. [Google Scholar] [CrossRef] [PubMed]
- Van Zanden, J.J.; de Mul, A.; Wortelboer, H.M.; Usta, M.; van Bladeren, P.J.; Rietjens, I.; Cnubben, N.H.P. Reversal of in vitro cellular MRP1 and MRP2 mediated vincristine resistance by the flavonoid myricetin. Biochem. Pharmacol. 2005, 69, 1657–1665. [Google Scholar] [CrossRef] [PubMed]
- Van Zanden, J.J.; Wortelboer, H.M.; Bijlsma, S.; Punt, A.; Usta, M.; van Bladeren, P.J.; Rietjens, I.; Cnubben, N.H.P. Quantitative structure activity relationship studies on the flavonoid mediated inhibition of multidrug resistance proteins 1 and 2. Biochem. Pharmacol. 2005, 69, 699–708. [Google Scholar] [CrossRef] [PubMed]
- Gentiluomo, M.; Garcia, P.P.; Galeotti, A.A.; Talar-Wojnarowska, R.; Tjaden, C.; Tavano, F.; Strobel, O.; Kupcinskas, J.; Neoptolemos, J.; Hegyi, P.; et al. Genetic variability of the ABCC2 gene and clinical outcomes in pancreatic cancer patients. Carcinogenesis 2019, 40, 544–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calcagno, A.M.; Ambudkar, S.V. Analysis of expression of drug resistance-linked ABC transporters in cancer cells by quantitative RT-PCR. Methods Mol. Biol. 2010, 637, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Chen, D.; Luo, M.; Ge, M.; Zhu, J. Knockdown of ribosomal protein L39 by RNA interference inhibits the growth of human pancreatic cancer cells in vitro and in vivo. Biotechnol. J. 2014, 9, 652–663. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Lines | Transfected Cells | IC50 * (µM) (Mean ± SEM) | p-Value |
|---|---|---|---|
| Caco-2 | Control-siRNA | 13.82 ± 1.21 | - |
| ABCC2-siRNA-1 | 7.75 ± 0.08 | 0.0001 | |
| ABCC2-siRNA-2 | 8.4 ± 0.13 | 0.0001 | |
| ABCC2-siRNA-3 | 7.03 ± 0.76 | 0.0001 | |
| PANC-1 | Control-siRNA | 35.13 ± 3.19 | - |
| ABCC2-siRNA-1 | 16.48 ± 0.53 | 0.018 | |
| ABCC2-siRNA-2 | 11.91 ± 0.25 | 0.027 | |
| ABCC2-siRNA-3 | 14.60 ± 0.72 | 0.012 |
| Cell Lines | siRNA Transfection | CDCF Accumulation (% of Control) | Platinum Accumulation (pmol/mgprotein) | |
|---|---|---|---|---|
| 25 μM Oxaliplatin | 100 μM Oxaliplatin | |||
| Caco-2 | Control-siRNA | 100 ± 8.4 | 5.8 ± 0.3 | 105.2 ± 0.4 |
| ABCC2-siRNA-1 | 133.1 ± 7.1 ** | 10.6 ± 2.6 | 128.8 ± 3.0 | |
| ABCC2-siRNA-2 | 152.6 ± 3.7 ** | 11.4 ± 1.0 * | 152.5 ± 0.7 * | |
| ABCC2-siRNA-3 | 145.5 ± 4.5 ** | 11.2 ± 1.5 * | 180.7 ± 24.5 ** | |
| PANC-1 | Control-siRNA | 100 ± 3.89 | 5.92 ± 0.26 | 32.6 ± 0.6 |
| ABCC2-siRNA-1 | 153.6 ± 6.3 ** | 78.0 ± 2.4 ** | 260.6 ± 2.7 ** | |
| ABCC2-siRNA-2 | 154.1 ± 10.0 ** | 84.3 ± 10.0 ** | 212.5 ± 14.8 ** | |
| ABCC2-siRNA-3 | 148.8 ± 3.9 ** | 72.3 ± 11.3 ** | 217.4 ± 50.5 ** | |
| Cell Lines | Cellular Platinum Accumulation (pmol/mg Protein) | p-Value | IC50 (µM) | p-Value | ||
|---|---|---|---|---|---|---|
| Myricetin | Control | Myricetin | Control | |||
| Caco-2 | 87 ± 3.35 | 74 ± 2.07 | 0.03 | 7.72 ± 0.36 | 10.4 ± 0.79 | 0.03 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biswas, R.; Bugde, P.; He, J.; Merien, F.; Lu, J.; Liu, D.-X.; Myint, K.; Liu, J.; McKeage, M.; Li, Y. Transport-Mediated Oxaliplatin Resistance Associated with Endogenous Overexpression of MRP2 in Caco-2 and PANC-1 Cells. Cancers 2019, 11, 1330. https://doi.org/10.3390/cancers11091330
Biswas R, Bugde P, He J, Merien F, Lu J, Liu D-X, Myint K, Liu J, McKeage M, Li Y. Transport-Mediated Oxaliplatin Resistance Associated with Endogenous Overexpression of MRP2 in Caco-2 and PANC-1 Cells. Cancers. 2019; 11(9):1330. https://doi.org/10.3390/cancers11091330
Chicago/Turabian StyleBiswas, Riya, Piyush Bugde, Ji He, Fabrice Merien, Jun Lu, Dong-Xu Liu, Khine Myint, Johnson Liu, Mark McKeage, and Yan Li. 2019. "Transport-Mediated Oxaliplatin Resistance Associated with Endogenous Overexpression of MRP2 in Caco-2 and PANC-1 Cells" Cancers 11, no. 9: 1330. https://doi.org/10.3390/cancers11091330
APA StyleBiswas, R., Bugde, P., He, J., Merien, F., Lu, J., Liu, D. -X., Myint, K., Liu, J., McKeage, M., & Li, Y. (2019). Transport-Mediated Oxaliplatin Resistance Associated with Endogenous Overexpression of MRP2 in Caco-2 and PANC-1 Cells. Cancers, 11(9), 1330. https://doi.org/10.3390/cancers11091330