Uncovering Tumour Heterogeneity through PKR and nc886 Analysis in Metastatic Colon Cancer Patients Treated with 5-FU-Based Chemotherapy

 ,
,  ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. RNA Extraction from FFPE Tissue and from Plasma Sample
2.3. RT qPCR Assay
2.4. Immunohistochemistry Analysis
2.5. Machine Learning and Statistical Analysis
2.6. Derivation of the Empirical Index
2.7. Feature Selection Process Using Non-Negative Matrix Factorization (NMF) in PGMRA
3. Results
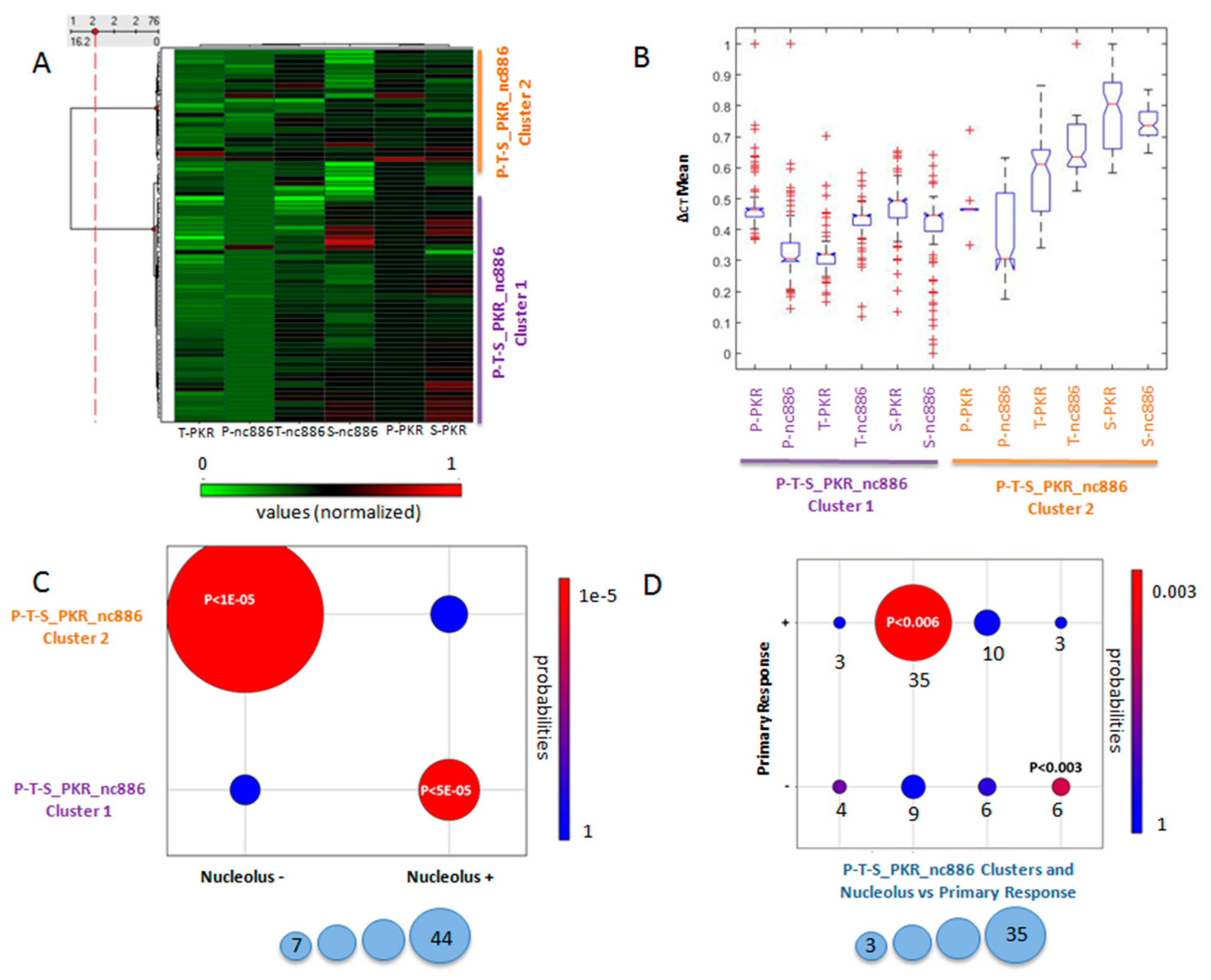
3.1. Normalized Values of Non Coding nc886 in Plasma and Tumor Tissues Predicted the Objective First-Line Chemotherapy Response
3.2. PKR Location Predicted the Objective First-Line Chemotherapy Response
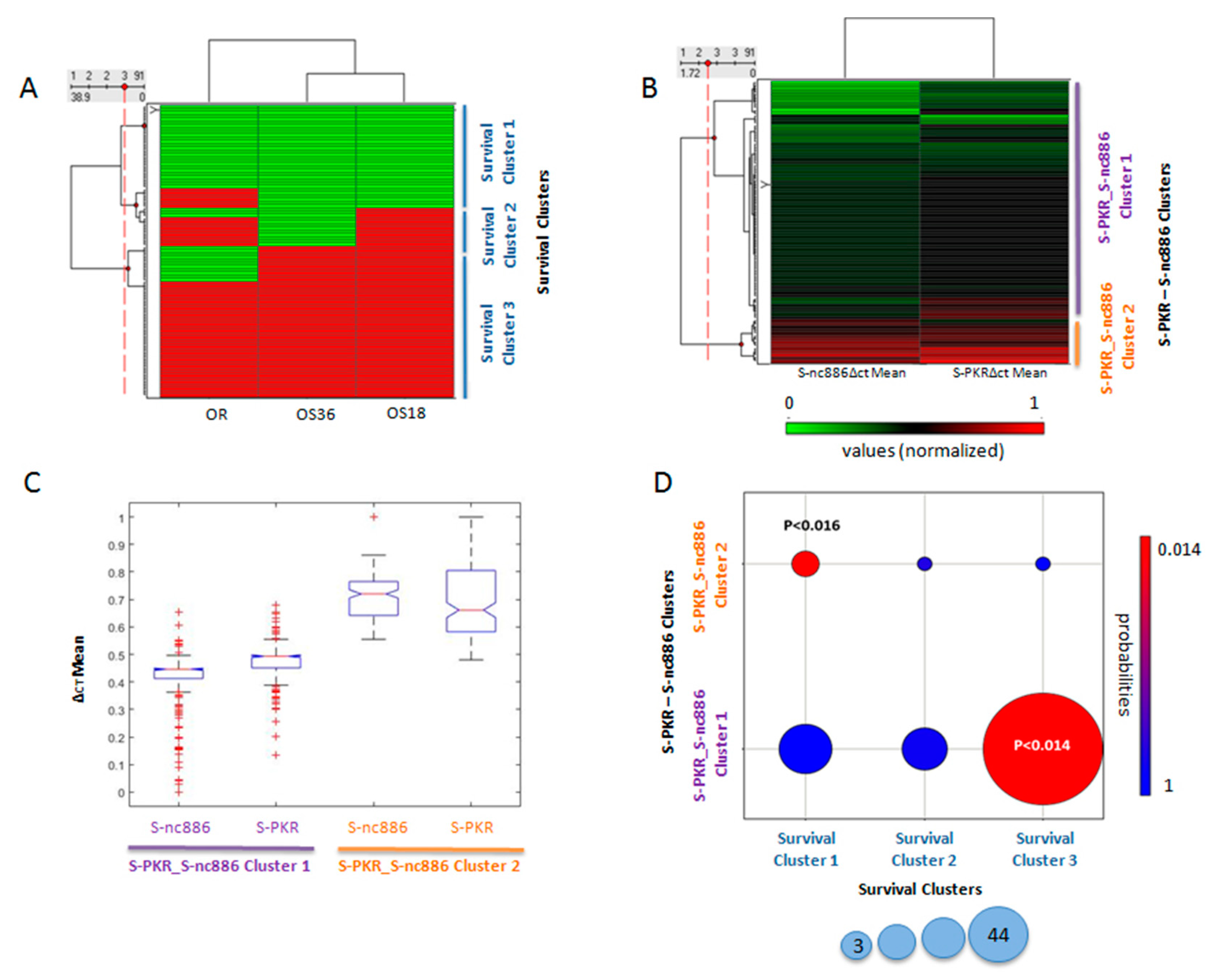
3.3. Final Outcome Was Predicted by the Expression Level of PKR and nc886 in Healthy Tissues
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Keum, N.; Giovannucci, E. Global burden of colorectal cancer: Emerging trends, risk factors and prevention strategies. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 713–732. [Google Scholar] [CrossRef] [PubMed]
- Vacante, M.; Borzì, A.M.; Basile, F.; Biondi, A. Biomarkers in colorectal cancer: Current clinical utility and future perspectives. World J. Clin. Cases 2018, 6, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Van Cutsem, E.; de Haas, S.; Kang, Y.K.; Ohtsu, A.; Tebbutt, N.C.; Xu, J.M.; Yong, W.P.; Langer, B.; Delmar, P.; Scherer, S.J.; et al. Bevacizumab in Combination With Chemotherapy as First-Line Therapy in Advanced Gastric Cancer: A Biomarker Evaluation From the AVAGAST Randomized Phase III Trial. J. Clin. Oncol. 2012, 30, 2119–2127. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.A.; Gil, J.; Ventoso, I.; Guerra, S.; Domingo, E.; Rivas, C.; Esteban, M. The impact of protein kinase PKR in cell biology: From antiviral to antiproliferative action. Microbiol. Mol. Biol. Rev. 2006, 70, 1032–1060. [Google Scholar] [CrossRef] [Green Version]
- Der, S.D.; Yang, Y.L.; Weissmann, C.; Williams, B.R. A double stranded RNA-activated protein kinase-dependent pathway mediating stress-induced apoptosis. Proc. Natl. Acad. Sci. USA 1997, 94, 3279–3283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil, J.; Alcami, J.; Esteban, M. Activation of NFkappa B by the dsRNA-dependent protein kinase, PKR, involves the I kappa B kinase complex. Oncogene 2000, 19, 1369–1378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchal, J.A.; Lopez, G.J.; Peran, M.; Comino, A.; Delgado, J.R.; García-García, J.A.; Conde, V.; Aranda, F.M.; Rivas, C.; Esteban, M.; et al. The Impact of PKR activation:from Neurodegeneration to Cancer. FASEB J. 2014, 28, 1965–1974. [Google Scholar] [CrossRef]
- Garcia-Ortega, M.B.; Lopez, G.J.; Jimenez, G.; Garcia-Garcia, J.A.; Conde, V.; Boulaiz, H.; Carrillo, E.; Perán, M.; Marchal, J.A.; Garcia, M.A. Clinical and therapeutic potential of protein kinase PKR in cancer and metabolism. Expert Rev. Mol. Med. 2017, 19, e9. [Google Scholar] [CrossRef]
- Kunkeaw, N.; Jeon, S.H.; Lee, K.; Johnson, B.H.; Tanasanvimon, S.; Javle, M.; Pairojkul, C.; Chamgramol, Y.; Wongfieng, W.; Gong, B.; et al. Cell death/proliferation roles for nc886, a non-coding RNA, in the protein kinase R pathway in cholangiocarcinoma. Oncogene 2013, 32, 3722–3731. [Google Scholar] [CrossRef] [Green Version]
- Jeon, S.H.; Lee, K.; Lee, K.S.; Kunkeaw, N.; Johnson, B.H.; Holthauzen, L.M.; Gong, B.; Leelayuwat, C.; Lee, Y.S. Characterization of the direct physical interaction of nc886, a cellular non-coding RNA, and PKR. FEBS Lett. 2012, 586, 3477–3484. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.S. A Novel Type of Non-coding RNA, nc886, Implicated in Tumor Sensing and Suppression. Genom. Inf. 2015, 13, 26–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calderon, B.M.; Conn, G.L. Human noncoding RNA 886 (nc886) adopts two structurally distinct conformers that are functionally opposing regulators of PKR. RNA 2017, 23, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Kunkeaw, N.; Lee, Y.S. Protein kinase R and its cellular regulators in cancer: An active player or a surveillant? Wiley Interdiscip. Rev. RNA 2019, e1558. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.A.; Carrasco, E.; Aguilera, M.; Alvarez, P.; Rivas, C.; Campos, J.M.; Prados, J.C.; Calleja, M.A.; Esteban, M.; Marchal, J.A.; et al. The Chemotherapeutic Drug 5-Fluorouracil Promotes PKR-Mediated Apoptosis in a p53-Independent Manner in Colon and Breast Cancer Cells. PLoS ONE 2011, 6, e23887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peidis, P.; Papadakis, A.I.; Muaddi, H.; Richard, S.; and Koromilas, A.E. Doxorubicin bypasses the cytoprotective effects of eIF2alpha phosphorylation and promotes PKR mediated cell death. Cell Death Differ. 2011, 18, 145–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, C.H.; Lee, E.S.; Lim, D.S.; Bae, Y.S. PKR, a p53 target gene, plays a crucial role in the tumor-suppressor function of p53. Proc. Natl. Acad. Sci. USA 2009, 106, 7852–7857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.S.; Park, J.L.; Lee, K.; Richardson, L.E.; Johnson, B.H.; Lee, H.S.; Lee, J.S.; Kim, S.B.; Kwon, O.H.; Song, K.S.; et al. nc886, a non-coding RNA of anti-proliferative role, is suppressed by CpG DNA methylation in human gastric cancer. Oncotarget 2014, 5, 3944–3955. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.S.; Lee, K.; Jang, H.J.; Lee, G.K.; Park, J.L.; Kim, S.Y.; Kim, S.B.; Johnson, B.H.; Zo, J.I.; Lee, J.S.; et al. Epigenetic silencing of the non-coding RNA nc886 provokes oncogenes during human esophageal tumorigenesis. Oncotarget 2014, 5, 3472–3481. [Google Scholar] [CrossRef] [Green Version]
- Fort, R.S.; Matho, C.; Geraldo, M.V.; Ottati, M.C.; Yamashita, A.S.; Saito, K.C.; Leite, K.R.; Méndez, M.; Maedo, N.; Méndez, L.; et al. Nc886 is epigenetically repressed in prostate cancer and acts as a tumor suppressor through the inhibition of cell growth. BMC Cancer 2018, 18, 127. [Google Scholar] [CrossRef] [Green Version]
- Arnedo, J.; del Val, C.; de Erausquin, G.A.; Romero-Zaliz, R.; Svrakic, D.; Cloninger, C.R.; Zwir, I. PGMRA: A web server for (phenotype x genotype) many-to-many relation analysis in GWAS. Nucleic Acids Res. 2013, 41, W142–W149. [Google Scholar] [CrossRef]
- Therasse, P.; Arbuck, S.G.; Eisenhauer, E.A.; Wanders, J.; Kaplan, R.S.; Rubinstein, L.; Verweij, J.; Van, G.M.; van Oosterom, A.T.; Christian, M.C.; et al. New guidelines to evaluate the response to treatment in solid tumors. European Organization for Research and Treatment of Cancer, National Cancer Institute of the United States, National Cancer Institute of Canada. J. Natl. Cancer Inst. 2000, 92, 205–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenhauer, E.A.; Therasse, P.; Bogaerts, J.; Schwartz, L.H.; Sargent, D.; Ford, R.; Dancey, J.; Arbuck, S.; Gwyther, S.; Mooney, M.; et al. New response evaluation criteria in solid tumours: Revised RECIST guideline (version 1.1). Eur. J. Cancer 2009, 45, 228–247. [Google Scholar] [CrossRef] [PubMed]
- Brierley, J.D.; Gospodarowicz, M.K.; Wittekind, C. TNM Classification of Malignant Tumours, 8th ed.; Wiley-Blackwell: New York, NY, USA, 2016; pp. 1–272. [Google Scholar]
- Huggett, J.; Dheda, K.; Bustin, S.; Zumla, A. Real-time RT-PCR normalisation; strategies and considerations. Genes Immun. 2005, 6, 279–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demes, M.; Bartsch, H.; Scheil-Bertram, S.; Mücke, R.; Fisseler-Eckhoff, A. Real-Time PCR Data Processing Shown by the Analysis of Colorectal Specific Candidate Genes, ERCC1, RRM1 and TS in Relation to β2M as Endogenous Control. Appl. Sci. 2012, 2, 139–159. [Google Scholar] [CrossRef]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2ˆ(-delta delta CT) method for quantitative real-time polymerase chain reaction data analysis. Biostat. Bioinforma. Biomath. 2013, 3, 71–85. [Google Scholar]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C (T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Geiger, J.T.; Weininger, F.; Gemmeke, J.F.; Wollmer, M.; Schuller, B.; Rigoll, G. Memory-enhanced neural networks and NMF for robust ASR. IEEE/ACM Trans. Audio Speech Lang. Process. 2014, 22, 1037–1046. [Google Scholar] [CrossRef]
- Hinton, G.E.; Salakhutdinov, R.R. Reducing the dimensionality of data with neural networks. Science 2006, 313, 504–507. [Google Scholar] [CrossRef] [Green Version]
- Arnedo, J.; Svrakic, D.M.; Del Val, C.; Romero-Zaliz, R.; Hernandez-Cuervo, H.; Fanous, A.H.; Pato, M.T.; Pato, C.N.; de Erausquin, G.A.; Cloninger, C.R.; et al. Uncovering the hidden risk architecture of the schizophrenias: Confirmation in three independent genome-wide association studies. Am. J. Psychiatry. 2015, 172, 139–153. [Google Scholar] [CrossRef]
- Romero-Zaliz, R.C.; Rubio-Escudero, C.; Cobb, J.P.; Zwir, I. A Multiobjective Evolutionary Conceptual Clustering Methodology for Gene Annotation within Structural Databases: A Case of Study on the Gene Ontology Database. IEEE Trans. Evol. Comput. 2008, 12, 679–701. [Google Scholar] [CrossRef]
- Arnedo, J.; Mamah, D.; Baranger, D.A.; Harms, M.P.; Barch, D.M.; Svrakic, D.M.; de Erausquin, G.A.; Cloninger, C.R.; Zwir, I. Decomposition of brain diffusion imaging data uncovers latent schizophrenias with distinct patterns of white matter anisotropy. NeuroImage 2015, 120, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Houle, D.; Govindaraju, D.R.; Omholt, S. Phenomics: The next challenge. Nat. Rev. Genet. 2011, 11, 855–866. [Google Scholar] [CrossRef] [PubMed]
- Zwir, I.; Huang, H.; Groisman, E.A. Analysis of differentially-regulated genes within a regulatory network by GPS genome navigation. Bioinformatics 2005, 21, 4073–4083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwir, I.; Shin, D.; Kato, A.; Nishino, K.; Latifi, T.; Solomon, F.; Hare, J.M.; Huang, H.; Groisman, E.A. Dissecting the PhoP regulatory network of Escherichia coli and Salmonella enterica. Proc. Natl. Acad. Sci. USA 2005, 102, 2862–2867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harari, O.; Park, S.Y.; Huang, H.; Groisman, E.A.; Zwir, I. Defining the plasticity of transcription factor binding sites by Deconstructing DNA consensus sequences: The PhoP-binding sites among gamma/enterobacteria. PLoS Comput. Biol. 2010, 6, e1000862. [Google Scholar] [CrossRef] [Green Version]
- Arnedo, J.; Romero-Zaliz, R.; Zwir, I.; Del Val, C. A multiobjective method for robust identification of bacterial small non-coding RNAs. Bioinformatics 2014, 30, 2875–2882. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.C.; Kraft, P.; Epstein, M.P.; Taylor, D.M.; Chanock, S.J.; Hunter, D.J.; Lin, X. Powerful SNP-set analysis for case-control genome-wide association studies. Am. J. Hum. Genet 2010, 86, 929–942. [Google Scholar] [CrossRef] [Green Version]
- Quan, C.; Xiao, J.; Liu, L.; Duan, Q.; Yuan, P.; Zhu, F. Protein Kinases as Tumor Biomarkers and Therapeutic Targets. Curr. Pharm. Des. 2017, 23, 4209–4225. [Google Scholar] [CrossRef]
- Zwir, I.; Del-Val, C.; Arnedo, J.; Pulkki-Råback, L.; Konte, B.; Yang, S.S.; Romero-Zaliz, R.; Hintsanen, M.; Cloninger, K.M.; Garcia, D.; et al. Three genetic–environmental networks for human personality. Mol. Psychiatry 2019, 21. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Kunkeaw, N.; Jeon, S.H.; Lee, I.; Johnson, B.H.; Kang, G.Y.; Bang, J.Y.; Park, H.S.; Leelayuwat, C.; Lee, Y.S. Precursor miR-886, a novel noncoding RNA repressed in cancer, associates with PKR and modulates its activity. RNA 2011, 17, 1076–1089. [Google Scholar] [CrossRef] [Green Version]
- Park, J.L.; Lee, Y.S.; Song, M.J.; Hong, S.H.; Ahn, J.H.; Seo, E.H.; Shin, S.P.; Lee, S.J.; Johnson, B.H.; Stampfer, M.R. Epigenetic regulation of RNA polymerase III transcription in early breast tumorigenesis. Oncogene 2017, 36, 6793–6804. [Google Scholar] [CrossRef] [Green Version]
- Ahn, J.H.; Lee, H.S.; Lee, J.S.; Lee, Y.S.; Park, J.L.; Kim, S.Y.; Hwang, J.A.; Kunkeaw, N.; Jung, S.Y.; Kim, T.J.; et al. Nc886 is induced by TGF-beta and suppresses the micro-RNA pathway in ovarian cancer. Nat. Commun. 2018, 9, 1166. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Song, Y.; Bi, N.; Shen, J.; Liu, W.; Fan, J.; Sun, G.; Tong, T.; He, J.; Shi, Y.; et al. DNA methylation-mediated repression of miR-886-3p predicts poor outcome of human small cell lung cancer. Cancer Res. 2013, 73, 3326–3335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treppendahl, M.B.; Qiu, X.; Sogaard, A.; Yang, X.; Nandrup-Bus, C.; Hother, C.; Andersen, M.K.; Kjeldsen, L.; Möllgård, L.; Hellström-Lindberg, E.; et al. Allelic methylation levels of the noncoding VTRNA2-1 located on chromosome 5q31.1 predict outcome in AML. Blood 2012, 119, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Burnett, S.B.; Vaughn, L.S.; Strom, J.M.; Francois, A.; Patel, R.C. A truncated PACT protein resulting from a frameshift mutation reported in movement disorder DYT16 triggers caspase activation and apoptosis. J. Cell Biochem. 2019, 120, 19004–19018. [Google Scholar] [CrossRef] [PubMed]
- Chukwurah, E.; Patel, R.C. Stress-induced TRBP phosphorylation enhances its interaction with PKR to regulate cellular survival. Sci. Rep. 2018, 8, 1020. [Google Scholar] [CrossRef] [PubMed]
- García, M.A.; Collado, M.; Muñoz-Fontela, C.; Matheu, A.; Marcos-Villar, L.; Arroyo, J.; Esteban, M.; Serrano, M.; Rivas, C. Antiviral action of the tumor suppressor ARF. EMBO J. 2006, 25, 4284–4292. [Google Scholar] [CrossRef]
- De la Cruz-Herrera, C.F.; Campagna, M.; García, M.A.; Marcos-Villar, L.; Lang, V.; Baz-Martínez, M.; Gutiérrez, S.; Vidal, A.; Rodríguez, M.S.; Esteban, M. Activation of the double-stranded RNA-dependent protein kinase PKR by small ubiquitin-like modifier (SUMO). J. Biol. Chem. 2014, 289, 26357–26367. [Google Scholar] [CrossRef] [Green Version]
- Villarroya-Beltri, C.; Guerra, S.; Sánchez-Madrid, F. ISGylation—A key to lock the cell gates for preventing the spread of threats. J. Cell Sci. 2017, 130, 2961–2969. [Google Scholar] [CrossRef] [Green Version]
- Blalock, W.L.; Bavelloni, A.; Piazzi, M.; Tagliavini, F.; Faenza, I.; Martelli, A.M.; Follo, M.Y.; Cocco, L. Multiple forms of PKR present in the nuclei of acute leukemia cells represent an active kinase that is responsive to stress. Leukemia 2011, 25, 236–245. [Google Scholar] [CrossRef] [Green Version]
- Tian, B.; Mathews, M.B. Functional characterization of and cooperation between the double-stranded RNA-binding motifs of the protein kinase PKR. J. Biol. Chem. 2001, 276, 9936–9944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piazzi, M.; Bavelloni, A.; Gallo, A.; Faenza, I.; Blalock, W.L. Signal Transduction in Ribosome Biogenesis: A Recipe to Avoid Disaster. Int. J. Mol. Sci. 2019, 20, 2718. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sex | ||||||||||
| Metastatic Colon Cancer Patients | Total | Male | Female | Age (Mean Years), SD | ||||||
| 197 | 127 | 70 | 65.1 ± 10.5 | |||||||
| Firs-line Chemotherapy Response clusters (OR) | Total | Responders(+) | Non-Responders(-) | ∆ct mean values, SD | ||||||
| 197 | 128 | 69 | ||||||||
| P-nc886_S-nc886 | T-nc886 | P-nc886 | ||||||||
| Cluster 1 | 160 | 110 | 50 | 0.427 ± 0.09 | 0.289 ± 0.05 | |||||
| Cluster 2 | 37 | 18 | 19 | 0.567 ± 0.17 | 0.495 ± 0.14 | |||||
| S-PKR_S-nc886 | S-nc886 | S-PKR | ||||||||
| Cluster 1 | 77 | 45 | 32 | 0.386 ± 0.12 | 0.465 ± 0.09 | |||||
| Cluster 2 | 14 | 6 | 8 | 0.682 ± 0.09 | 0.703 ± 0.16 | |||||
| Survival clusters (OS) | First-line Response | OS 18 m | OS 36 m | |||||||
| Survival | + | - | Survival | Exitus | Survival | Exitus | ||||
| Cluster 1 | 26 | 0 | 26 | 0 | 26 | 0 | 26 | |||
| Cluster 2 | 18 | 15 | 3 | 12 | 6 | 0 | 18 | |||
| Cluster 3 | 47 | 36 | 11 | 47 | 0 | 47 | 0 | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ortega-García, M.B.; Mesa, A.; Moya, E.L.J.; Rueda, B.; Lopez-Ordoño, G.; García, J.Á.; Conde, V.; Redondo-Cerezo, E.; Lopez-Hidalgo, J.L.; Jiménez, G.; et al. Uncovering Tumour Heterogeneity through PKR and nc886 Analysis in Metastatic Colon Cancer Patients Treated with 5-FU-Based Chemotherapy. Cancers 2020, 12, 379. https://doi.org/10.3390/cancers12020379
Ortega-García MB, Mesa A, Moya ELJ, Rueda B, Lopez-Ordoño G, García JÁ, Conde V, Redondo-Cerezo E, Lopez-Hidalgo JL, Jiménez G, et al. Uncovering Tumour Heterogeneity through PKR and nc886 Analysis in Metastatic Colon Cancer Patients Treated with 5-FU-Based Chemotherapy. Cancers. 2020; 12(2):379. https://doi.org/10.3390/cancers12020379
Chicago/Turabian StyleOrtega-García, María Belén, Alberto Mesa, Elisa L.J. Moya, Beatriz Rueda, Gabriel Lopez-Ordoño, Javier Ángel García, Verónica Conde, Eduardo Redondo-Cerezo, Javier Luis Lopez-Hidalgo, Gema Jiménez, and et al. 2020. "Uncovering Tumour Heterogeneity through PKR and nc886 Analysis in Metastatic Colon Cancer Patients Treated with 5-FU-Based Chemotherapy" Cancers 12, no. 2: 379. https://doi.org/10.3390/cancers12020379
APA StyleOrtega-García, M. B., Mesa, A., Moya, E. L. J., Rueda, B., Lopez-Ordoño, G., García, J. Á., Conde, V., Redondo-Cerezo, E., Lopez-Hidalgo, J. L., Jiménez, G., Peran, M., Martínez-González, L. J., del Val, C., Zwir, I., Marchal, J. A., & García, M. Á. (2020). Uncovering Tumour Heterogeneity through PKR and nc886 Analysis in Metastatic Colon Cancer Patients Treated with 5-FU-Based Chemotherapy. Cancers, 12(2), 379. https://doi.org/10.3390/cancers12020379