Microcellular Environmental Regulation of Silver Nanoparticles in Cancer Therapy: A Critical Review

 , ,
, ,  ,
, 

Abstract
:1. Introduction
2. Biodistribution of AgNPs and Mechanical Cues Regulating Tumor Growth
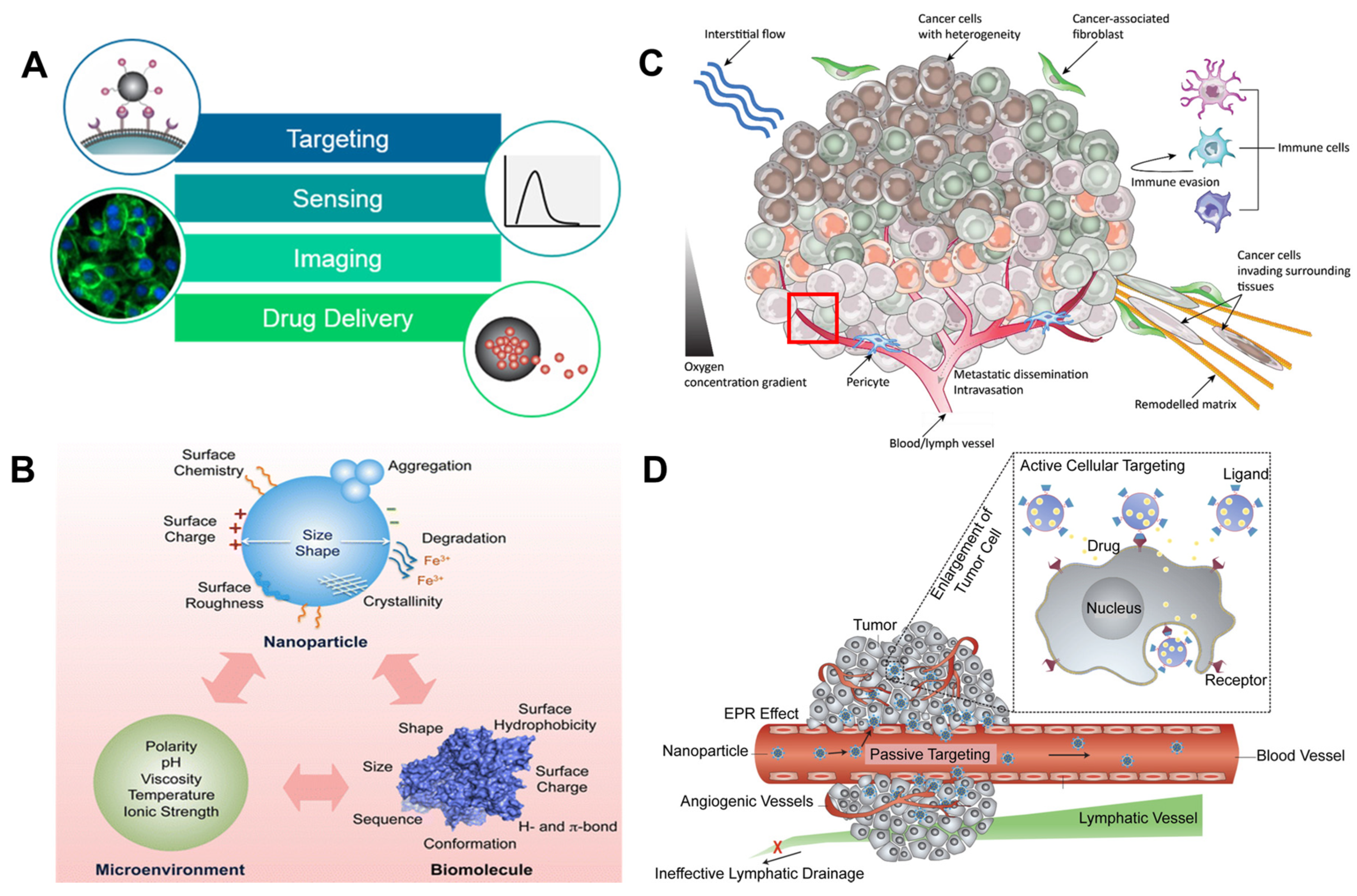
2.1. Heterogeneity of Cancer Cells
2.2. Phagocytosis and ROS Generation
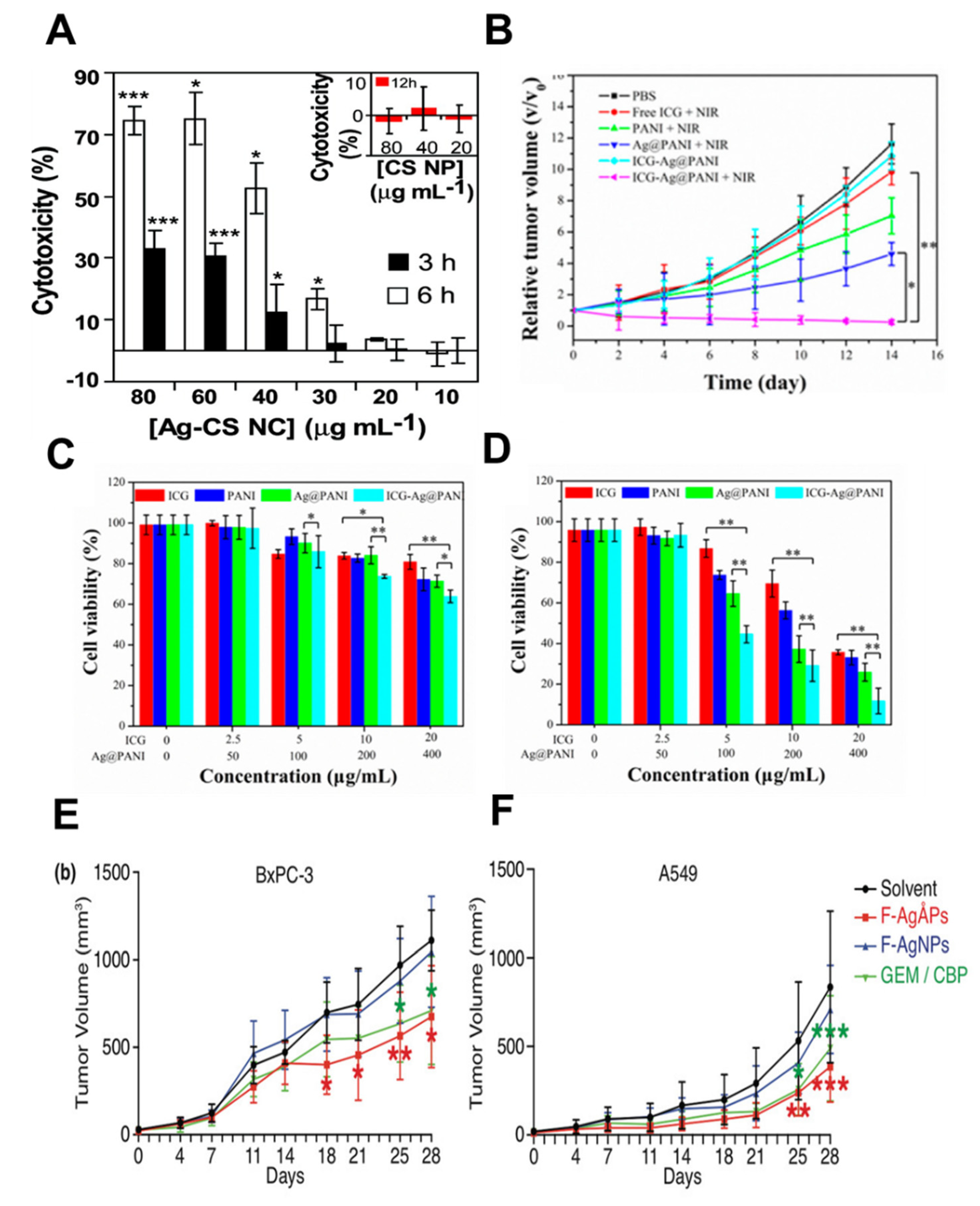
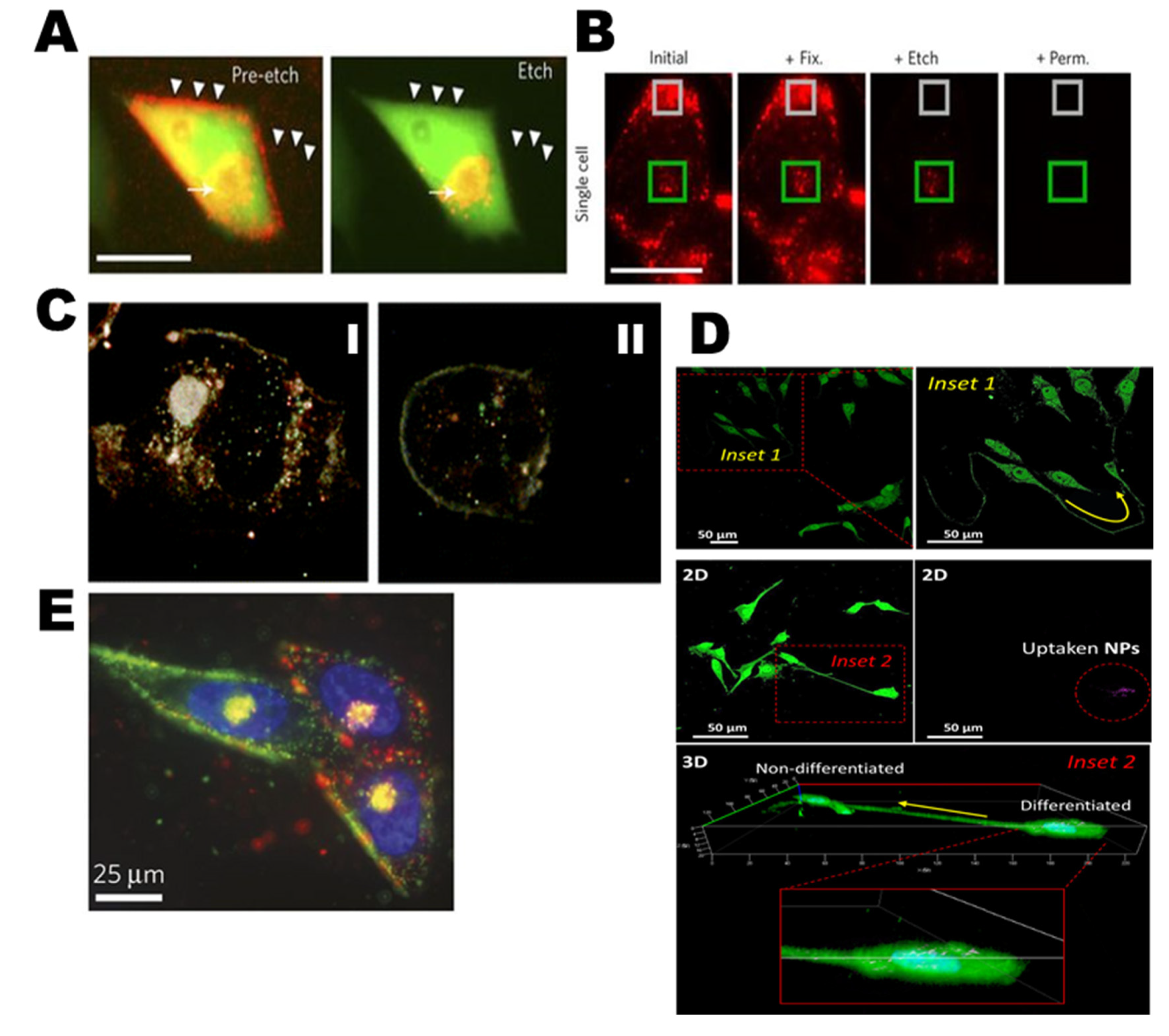
2.3. Biodistribution of AgNPs and Tumor Targeting
2.4. Relative Tumor Thermal Therapies
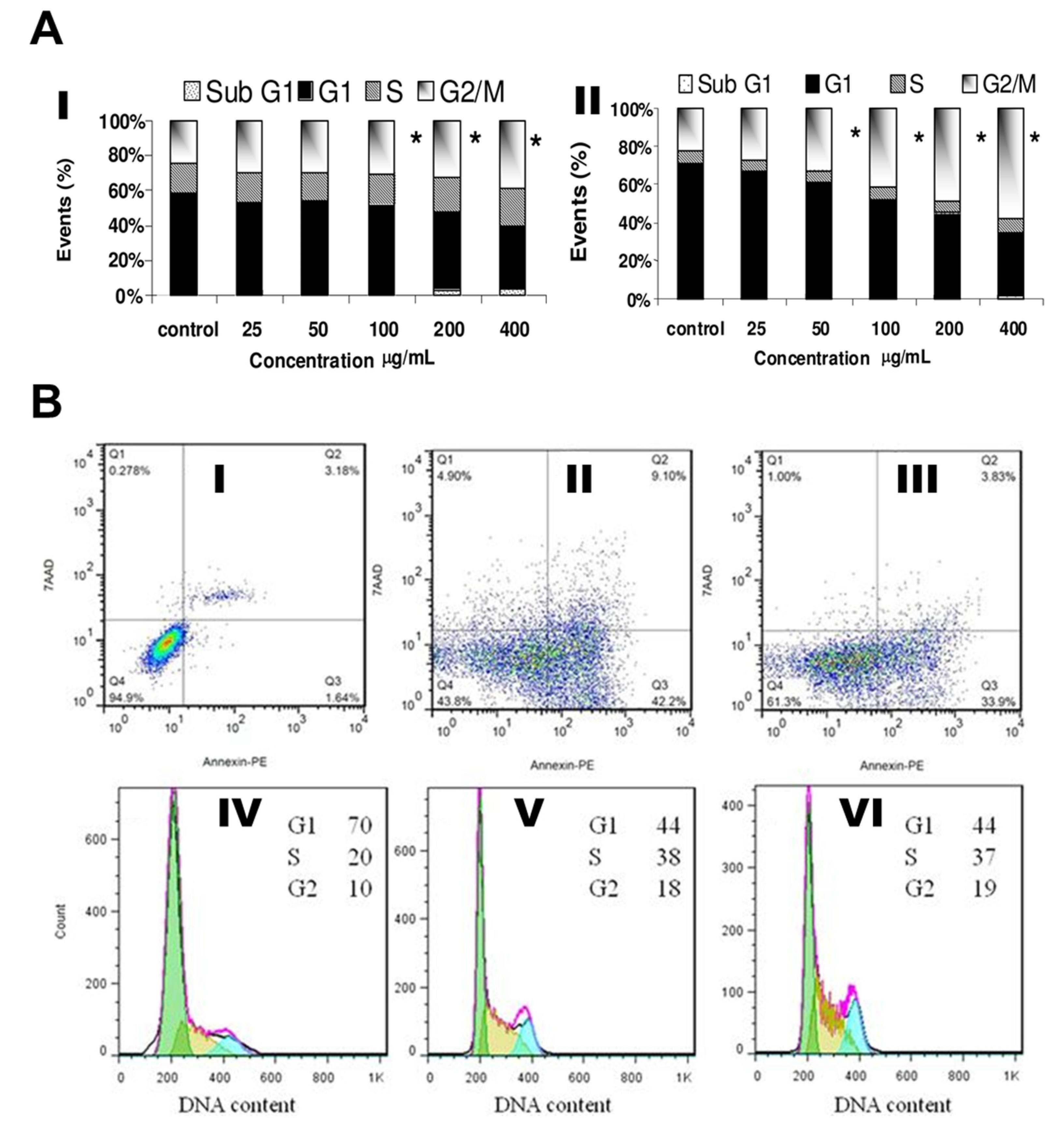
2.5. Cell cycles and Cell Death
3. AgNPs Currently Applied Proteogenomic Imaging and Drug Delivery
3.1. Drug Delivery and Theragnostics
3.2. Mechanisms of Proteogenomic Signal Specificity and Apoptosis
3.3. Fate of Stress-Related Therapeutics
4. AgNPs Currently Applied in Chemical Metabolomics and Suppression of Metabolic Pathways
4.1. NMR Spectra and Assignment of Cancer Metabolites
4.2. Pattern Recognition of Cancer Cells
4.3. Topology of Energy Metabolism
4.4. Topology of Antioxidant Defense Metabolism
4.5. Topology of Membrane Metabolism
5. Conclusion, Challenges, and Future Perspectives
Funding
Acknowledgments
Conflicts of Interest
Ethical Approval
Abbreviations
| Ag | Silver |
| Ag2CO3 | Silver carbonate |
| AP-1 | Activator protein 1 |
| ATP | Adenosine triphosphate |
| Ca2+ | Calcium ions |
| Cit | Citrate |
| Conc | Concentrations |
| CXCR1 | C-X-C Motif Chemokine Receptor 1 |
| DNA | Deoxyribonucleic acid |
| GS | Green synthesis |
| GSH | Glutathione |
| h | Hours |
| H2O2 | Hydrogen peroxide |
| IL | Interleukin |
| JNK | c-Jun N-terminal kinases |
| LDHQ | Lactate dehydrogenase |
| mg/mL | Milligram/milliliter |
| MTT | Methylthiazolyldiphenyl-tetrazolium bromide |
| NAC | N-acetylcysteine |
| NAD+ | Nicotinamide adenine dinucleotide |
| NADP+ | Nicotinamide adenine dinucleotide phosphate |
| NF-kB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| nm | Nanometer |
| NMR | Nuclear magnetic resonance |
| NPs | Nanoparticles |
| OPLS-DA | Orthogonal PLS-DA |
| OS | Oxidative stress |
| p38 MAPK | p38 mitogen-activated protein kinases |
| PCA | Principal component analysis |
| PCR | Polymerase chain reaction |
| Pgp | P-glycoprotein |
| PLS-DA | Partial least squares-discriminant analysis |
| Ref | References |
| RNA | Ribonucleic acid |
| ROS | Reactive oxygen species |
| SOD | Superoxide dismutase |
| SRXN1 | Sulfiredoxin 1 |
| TCA | Tricarboxylic acid |
| TNF | Tumor necrosis factor |
| µg/L | Microgram/Liter |
| μm | Micrometer |
References
- Xu, H.; Qu, F.; Xu, H.; Lai, W.; Andrew Wang, Y.; Aguilar, Z.P.; Wei, H. Role of reactive oxygen species in the antibacterial mechanism of silver nanoparticles on Escherichia coli O157:H7. Biometals 2012, 25, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Quinteros, M.A.; Cano Aristizabal, V.; Dalmasso, P.R.; Paraje, M.G.; Paez, P.L. Oxidative stress generation of silver nanoparticles in three bacterial genera and its relationship with the antimicrobial activity. Toxicol. In Vitro 2016, 36, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Siritongsuk, P.; Hongsing, N.; Thammawithan, S.; Daduang, S.; Klaynongsruang, S.; Tuanyok, A.; Patramanon, R. Two-Phase Bactericidal Mechanism of Silver Nanoparticles against Burkholderia pseudomallei. PLoS ONE 2016, 11, e0168098. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, N.; Ramalingam, C. Silver nanoparticle antimicrobial activity explained by membrane rupture and reactive oxygen generation. Environ. Chem. Lett. 2016, 14, 477–485. [Google Scholar] [CrossRef]
- Seong, M.; Lee, D.G. Silver Nanoparticles against Salmonella enterica Serotype Typhimurium: Role of Inner Membrane Dysfunction. Curr. Microbiol. 2017, 74, 661–670. [Google Scholar] [CrossRef]
- Jain, J.; Arora, S.; Rajwade, J.M.; Omray, P.; Khandelwal, S.; Paknikar, K.M. Silver nanoparticles in therapeutics: Development of an antimicrobial gel formulation for topical use. Mol. Pharm. 2009, 6, 1388–1401. [Google Scholar] [CrossRef]
- Kokura, S.; Handa, O.; Takagi, T.; Ishikawa, T.; Naito, Y.; Yoshikawa, T. Silver nanoparticles as a safe preservative for use in cosmetics. Nanomedicine 2010, 6, 570–574. [Google Scholar] [CrossRef]
- Jung, J.H.; Hwang, G.B.; Lee, J.E.; Bae, G.N. Preparation of airborne Ag/CNT hybrid nanoparticles using an aerosol process and their application to antimicrobial air filtration. Langmuir 2011, 27, 10256–10264. [Google Scholar] [CrossRef]
- Zodrow, K.; Brunet, L.; Mahendra, S.; Li, D.; Zhang, A.; Li, Q.; Alvarez, P.J. Polysulfone ultrafiltration membranes impregnated with silver nanoparticles show improved biofouling resistance and virus removal. Water Res. 2009, 43, 715–723. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Wu, X.L.; Chen, Y.Y.; Lin, H. Application of Silver Nanoparticles to Cotton Fabric as an Antibacterial Textile Finish. Fiber. Polym. 2009, 10, 496–501. [Google Scholar] [CrossRef] [Green Version]
- Slane, J.; Vivanco, J.; Rose, W.; Ploeg, H.L.; Squire, M. Mechanical, material, and antimicrobial properties of acrylic bone cement impregnated with silver nanoparticles. Mater. Sci. Eng. C Mater. Biol. Appl. 2015, 48, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Pollini, M.; Paladini, F.; Catalano, M.; Taurino, A.; Licciulli, A.; Maffezzoli, A.; Sannino, A. Antibacterial coatings on haemodialysis catheters by photochemical deposition of silver nanoparticles. J. Mater. Sci. Mater. Med. 2011, 22, 2005–2012. [Google Scholar] [CrossRef]
- Morris, A.S.; Salem, A.K. Surface Engineered Nanoparticles: Considerations for Biomedical Applications. Adv. Eng. Mater. 2017, 19, 1700302. [Google Scholar] [CrossRef]
- Elinav, E.; Peer, D. Harnessing nanomedicine for mucosal theranostics--a silver bullet at last? ACS Nano 2013, 7, 2883–2890. [Google Scholar] [CrossRef] [PubMed]
- Homan, K.A.; Souza, M.; Truby, R.; Luke, G.P.; Green, C.; Vreeland, E.; Emelianov, S. Silver nanoplate contrast agents for in vivo molecular photoacoustic imaging. ACS Nano 2012, 6, 641–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mu, Q.; Jiang, G.; Chen, L.; Zhou, H.; Fourches, D.; Tropsha, A.; Yan, B. Chemical basis of interactions between engineered nanoparticles and biological systems. Chem. Rev. 2014, 114, 7740–7781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riaz Ahmed, K.B.; Nagy, A.M.; Brown, R.P.; Zhang, Q.; Malghan, S.G.; Goering, P.L. Silver nanoparticles: Significance of physicochemical properties and assay interference on the interpretation of in vitro cytotoxicity studies. Toxicol. In Vitro 2017, 38, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Gao, H.; Bao, G. Physical Principles of Nanoparticle Cellular Endocytosis. ACS Nano 2015, 9, 8655–8671. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Qin, T.; Ingle, T.; Yan, J.; He, W.; Yin, J.J.; Chen, T. Differential genotoxicity mechanisms of silver nanoparticles and silver ions. Arch. Toxicol. 2017, 91, 509–519. [Google Scholar] [CrossRef]
- Arora, S.; Jain, J.; Rajwade, J.M.; Paknikar, K.M. Cellular responses induced by silver nanoparticles: In vitro studies. Toxicol. Lett. 2008, 179, 93–100. [Google Scholar] [CrossRef]
- Piao, M.J.; Kang, K.A.; Lee, I.K.; Kim, H.S.; Kim, S.; Choi, J.Y.; Choi, J.; Hyun, J.W. Silver nanoparticles induce oxidative cell damage in human liver cells through inhibition of reduced glutathione and induction of mitochondria-involved apoptosis. Toxicol. Lett. 2011, 201, 92–100. [Google Scholar] [CrossRef]
- Foldbjerg, R.; Dang, D.A.; Autrup, H. Cytotoxicity and genotoxicity of silver nanoparticles in the human lung cancer cell line, A549. Arch. Toxicol. 2011, 85, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Chairuangkitti, P.; Lawanprasert, S.; Roytrakul, S.; Aueviriyavit, S.; Phummiratch, D.; Kulthong, K.; Chanvorachote, P.; Maniratanachote, R. Silver nanoparticles induce toxicity in A549 cells via ROS-dependent and ROS-independent pathways. Toxicol. In Vitro 2013, 27, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Park, M.V.; Neigh, A.M.; Vermeulen, J.P.; de la Fonteyne, L.J.; Verharen, H.W.; Briede, J.J.; van Loveren, H.; de Jong, W.H. The effect of particle size on the cytotoxicity, inflammation, developmental toxicity and genotoxicity of silver nanoparticles. Biomaterials 2011, 32, 9810–9817. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.; Zhu, L.; Huang, Z.; Zhou, H.; Ge, Y.; Ma, W.; Wu, J.; Zhang, X.; Zhou, X.; Zhang, Y.; et al. Anti-leukemia activity of PVP-coated silver nanoparticles via generation of reactive oxygen species and release of silver ions. Biomaterials 2013, 34, 7884–7894. [Google Scholar] [CrossRef]
- Sanpui, P.; Chattopadhyay, A.; Ghosh, S.S. Induction of apoptosis in cancer cells at low silver nanoparticle concentrations using chitosan nanocarrier. ACS Appl. Mater. Interfaces 2011, 3, 218–228. [Google Scholar] [CrossRef]
- Auffan, M.; Rose, J.; Wiesner, M.R.; Bottero, J.Y. Chemical stability of metallic nanoparticles: A parameter controlling their potential cellular toxicity in vitro. Environ. Pollut. 2009, 157, 1127–1133. [Google Scholar] [CrossRef]
- Fu, P.P.; Xia, Q.; Hwang, H.M.; Ray, P.C.; Yu, H. Mechanisms of nanotoxicity: Generation of reactive oxygen species. J. Food Drug Anal. 2014, 22, 64–75. [Google Scholar] [CrossRef] [Green Version]
- Wei, L.; Lu, J.; Xu, H.; Patel, A.; Chen, Z.S.; Chen, G. Silver nanoparticles: Synthesis, properties, and therapeutic applications. Drug Discov. Today 2015, 20, 595–601. [Google Scholar] [CrossRef] [Green Version]
- Asharani, P.V.; Lian Wu, Y.; Gong, Z.; Valiyaveettil, S. Toxicity of silver nanoparticles in zebrafish models. Nanotechnology 2008, 19, 255102. [Google Scholar] [CrossRef]
- Van Aerle, R.; Lange, A.; Moorhouse, A.; Paszkiewicz, K.; Ball, K.; Johnston, B.D.; de-Bastos, E.; Booth, T.; Tyler, C.R.; Santos, E.M. Molecular mechanisms of toxicity of silver nanoparticles in zebrafish embryos. Environ. Sci. Technol. 2013, 47, 8005–8014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.N.; Habbit, N.L.; Su, C.Y.; Choi, N.; Ahn, E.H.; Lipke, E.A.; Kim, D.H. Microphysiological Systems as Enabling Tools for Modeling Complexity in the Tumor Microenvironment and Accelerating Cancer Drug Development. Adv. Funct. Mater. 2019, 29, 1807553. [Google Scholar] [CrossRef]
- Junttila, M.R.; de Sauvage, F.J. Influence of tumour micro-environment heterogeneity on therapeutic response. Nature 2013, 501, 346–354. [Google Scholar] [CrossRef]
- Abdi, S.; Ali, A. Role of ROS modified human DNA in the pathogenesis and etiology of cancer. Cancer Lett. 1999, 142, 1–9. [Google Scholar] [CrossRef]
- Chompoosor, A.; Saha, K.; Ghosh, P.S.; Macarthy, D.J.; Miranda, O.R.; Zhu, Z.J.; Arcaro, K.F.; Rotello, V.M. The Role of Surface Functionality on Acute Cytotoxicity, ROS Generation and DNA Damage by Cationic Gold Nanoparticles. Small 2010, 6, 2246–2249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, B.; Chen, Y.; Shi, J. Reactive Oxygen Species (ROS)-Based Nanomedicine. Chem. Rev. 2019, 119, 4881–4985. [Google Scholar] [CrossRef] [PubMed]
- Sadat Shandiz, S.A.; Shafiee Ardestani, M.; Shahbazzadeh, D.; Assadi, A.; Ahangari Cohan, R.; Asgary, V.; Salehi, S. Novel imatinib-loaded silver nanoparticles for enhanced apoptosis of human breast cancer MCF-7 cells. Artif. Cells Nanomed. Biotechnol. 2017, 45, 1–10. [Google Scholar] [CrossRef]
- Li, Y.; Guo, M.; Lin, Z.; Zhao, M.; Xiao, M.; Wang, C.; Xu, T.; Chen, T.; Zhu, B. Polyethylenimine-functionalized silver nanoparticle-based co-delivery of paclitaxel to induce HepG2 cell apoptosis. Int. J. Nanomedicine 2016, 11, 6693–6702. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Choi, J.E.; Choi, J.; Chung, K.H.; Park, K.; Yi, J.; Ryu, D.Y. Oxidative stress-dependent toxicity of silver nanoparticles in human hepatoma ells. Toxicol. In Vitro 2009, 23, 1076–1084. [Google Scholar] [CrossRef]
- Yong, T.; Zhang, X.; Bie, N.; Zhang, H.; Zhang, X.; Li, F.; Hakeem, A.; Hu, J.; Gan, L.; Santos, H.A.; et al. Tumor exosome-based nanoparticles are efficient drug carriers for chemotherapy. Nat. Commun. 2019, 10, 3838. [Google Scholar] [CrossRef] [Green Version]
- Bohmert, L.; Niemann, B.; Thunemann, A.F.; Lampen, A. Cytotoxicity of peptide-coated silver nanoparticles on the human intestinal cell line Caco-2. Arch. Toxicol. 2012, 86, 1107–1115. [Google Scholar] [CrossRef] [PubMed]
- Bohmert, L.; Niemann, B.; Lichtenstein, D.; Juling, S.; Lampen, A. Molecular mechanism of silver nanoparticles in human intestinal cells. Nanotoxicology 2015, 9, 852–860. [Google Scholar] [CrossRef]
- Gurunathan, S.; Park, J.H.; Han, J.W.; Kim, J.H. Comparative assessment of the apoptotic potential of silver nanoparticles synthesized by Bacillus tequilensis and Calocybe indica in MDA-MB-231 human breast cancer cells: Targeting p53 for anticancer therapy. Int. J. Nanomedicine 2015, 10, 4203–4222. [Google Scholar] [CrossRef] [Green Version]
- Kawata, K.; Osawa, M.; Okabe, S. In Vitro Toxicity of Silver Nanoparticles at Noncytotoxic Doses to HepG2 Human Hepatoma Cells. Environ. Sci. Technol. 2009, 43, 6046–6051. [Google Scholar] [CrossRef]
- Wang, J.; Rahman, M.F.; Duhart, H.M.; Newport, G.D.; Patterson, T.A.; Murdock, R.C.; Hussain, S.M.; Schlager, J.J.; Ali, S.F. Expression changes of dopaminergic system-related genes in PC12 cells induced by manganese, silver, or copper nanoparticles. Neurotoxicology 2009, 30, 926–933. [Google Scholar] [CrossRef] [PubMed]
- Hsin, Y.H.; Chen, C.F.; Huang, S.; Shih, T.S.; Lai, P.S.; Chueh, P.J. The apoptotic effect of nanosilver is mediated by a ROS- and JNK-dependent mechanism involving the mitochondrial pathway in NIH3T3 cells. Toxicol. Lett. 2008, 179, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Guan, R.; Ye, X.; Jiang, J.; Liu, M.; Huang, G.; Chen, X. Toxicity of nano- and micro- sized silver particles in human hepatocyte cell line L02. J. Phys. Conf. Ser. 2011, 304, 012036. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Wu, Y.; Wang, C.; Li, H.C.; Wang, T.; Liao, C.Y.; Cui, L.; Zhou, Q.F.; Yan, B.; Jiang, G.B. Impact of silver nanoparticles on human cells: Effect of particle size. Nanotoxicology 2010, 4, 319–330. [Google Scholar] [CrossRef]
- Miethling-Graff, R.; Rumpker, R.; Richter, M.; Verano-Braga, T.; Kjeldsen, F.; Brewer, J.; Hoyland, J.; Rubahn, H.G.; Erdmann, H. Exposure to silver nanoparticles induces size- and dose-dependent oxidative stress and cytotoxicity in human colon carcinoma cells. Toxicology Vitro 2014, 28, 1280–1289. [Google Scholar] [CrossRef]
- Miura, N.; Shinohara, Y. Cytotoxic effect and apoptosis induction by silver nanoparticles in HeLa cells. Biochem. Biophys. Res. Commun. 2009, 390, 733–737. [Google Scholar] [CrossRef]
- Williams, K.M.; Gokulan, K.; Cerniglia, C.E.; Khare, S. Size and dose dependent effects of silver nanoparticle exposure on intestinal permeability in an in vitro model of the human gut epithelium. J. Nanobiotechnology 2016, 14, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Y.G.; Gurunathan, S. Combination of graphene oxide-silver nanoparticle nanocomposites and cisplatin enhances apoptosis and autophagy in human cervical cancer cells. Int. J. Nanomedicine 2017, 12, 6537–6558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dziedzic, A.; Kubina, R.; Buldak, R.J.; Skonieczna, M.; Cholewa, K. Silver Nanoparticles Exhibit the Dose-Dependent Anti-Proliferative Effect against Human Squamous Carcinoma Cells Attenuated in the Presence of Berberine. Molecules 2016, 21, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, Z.H.; Li, M.; Feng, Y.X.; Shi, J.C.; Zhang, J.; Shao, B. Hormesis effects of silver nanoparticles at non-cytotoxic doses to human hepatoma cells. PLoS ONE 2014, 9, e102564. [Google Scholar] [CrossRef]
- Gopisetty, M.K.; Kovacs, D.; Igaz, N.; Ronavari, A.; Belteky, P.; Razga, Z.; Venglovecz, V.; Csoboz, B.; Boros, I.M.; Konya, Z.; et al. Endoplasmic reticulum stress: Major player in size-dependent inhibition of P-glycoprotein by silver nanoparticles in multidrug-resistant breast cancer cells. J. Nanobiotechnology 2019, 17, 9. [Google Scholar] [CrossRef]
- Dadashpour, M.; Firouzi-Amandi, A.; Pourhassan-Moghaddam, M.; Maleki, M.J.; Soozangar, N.; Jeddi, F.; Nouri, M.; Zarghami, N.; Pilehvar-Soltanahmadi, Y. Biomimetic synthesis of silver nanoparticles using Matricaria chamomilla extract and their potential anticancer activity against human lung cancer cells. Mater. Sci. Eng. C Mater. Biol. Appl. 2018, 92, 902–912. [Google Scholar] [CrossRef]
- Kang, S.J.; Ryoo, I.G.; Lee, Y.J.; Kwak, M.K. Role of the Nrf2-heme oxygenase-1 pathway in silver nanoparticle-mediated cytotoxicity. Toxicol. Appl. Pharmacol. 2012, 258, 89–98. [Google Scholar] [CrossRef]
- Lee, Y.H.; Cheng, F.Y.; Chiu, H.W.; Tsai, J.C.; Fang, C.Y.; Chen, C.W.; Wang, Y.J. Cytotoxicity, oxidative stress, apoptosis and the autophagic effects of silver nanoparticles in mouse embryonic fibroblasts. Biomaterials 2014, 35, 4706–4715. [Google Scholar] [CrossRef]
- Yin, N.; Yao, X.; Zhou, Q.; Faiola, F.; Jiang, G. Vitamin E attenuates silver nanoparticle-induced effects on body weight and neurotoxicity in rats. Biochem. Biophys. Res. Commun. 2015, 458, 405–410. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.L.; Hsiao, I.L.; Lin, H.C.; Wang, C.F.; Huang, Y.J.; Chuang, C.Y. Silver nanoparticles affect on gene expression of inflammatory and neurodegenerative responses in mouse brain neural cells. Environ. Res. 2015, 136, 253–263. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.J.; Nallathamby, P.D.; Browning, L.M.; Osgood, C.J.; Xu, X.H. In vivo imaging of transport and biocompatibility of single silver nanoparticles in early development of zebrafish embryos. ACS Nano 2007, 1, 133–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sriram, M.I.; Kalishwaralal, K.; Barathmanikanth, S.; Gurunathani, S. Size-based cytotoxicity of silver nanoparticles in bovine retinal endothelial cells. Nanosci. Methods 2012, 1, 56–77. [Google Scholar] [CrossRef]
- Foldbjerg, R.; Irving, E.S.; Hayashi, Y.; Sutherland, D.S.; Thorsen, K.; Autrup, H.; Beer, C. Global gene expression profiling of human lung epithelial cells after exposure to nanosilver. Toxicol. Sci. 2012, 130, 145–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.J.; Lee, S.J.; Yun, S.J.; Jang, J.Y.; Kang, H.; Kim, K.; Choi, I.H.; Park, S. Silver nanoparticles affect glucose metabolism in hepatoma cells through production of reactive oxygen species. Int. J. Nanomedicine 2016, 11, 55–68. [Google Scholar]
- Wan, C.; Tai, J.; Zhang, J.; Guo, Y.; Zhu, Q.; Ling, D.; Gu, F.; Gan, J.; Zhu, C.; Wang, Y.; et al. Silver nanoparticles selectively induce human oncogenic gamma-herpesvirus-related cancer cell death through reactivating viral lytic replication. Cell Death Dis. 2019, 10, 392. [Google Scholar] [CrossRef] [Green Version]
- Dziendzikowska, K.; Gromadzka-Ostrowska, J.; Lankoff, A.; Oczkowski, M.; Krawczynska, A.; Chwastowska, J.; Sadowska-Bratek, M.; Chajduk, E.; Wojewodzka, M.; Dusinska, M.; et al. Time-dependent biodistribution and excretion of silver nanoparticles in male Wistar rats. J. Appl. Toxicol. 2012, 32, 920–928. [Google Scholar] [CrossRef]
- Li, Y.; Bhalli, J.A.; Ding, W.; Yan, J.; Pearce, M.G.; Sadiq, R.; Cunningham, C.K.; Jones, M.Y.; Monroe, W.A.; Howard, P.C.; et al. Cytotoxicity and genotoxicity assessment of silver nanoparticles in mouse. Nanotoxicology 2014, 8, 36–45. [Google Scholar] [CrossRef]
- Loeschner, K.; Hadrup, N.; Qvortrup, K.; Larsen, A.; Gao, X.; Vogel, U.; Mortensen, A.; Lam, H.R.; Larsen, E.H. Distribution of silver in rats following 28 days of repeated oral exposure to silver nanoparticles or silver acetate. Part. Fibre Toxicol. 2011, 8, 18. [Google Scholar] [CrossRef] [Green Version]
- Sthijns, M.M.; Thongkam, W.; Albrecht, C.; Hellack, B.; Bast, A.; Haenen, G.R.; Schins, R.P. Silver nanoparticles induce hormesis in A549 human epithelial cells. Toxicol. In Vitro 2017, 40, 223–233. [Google Scholar] [CrossRef]
- Kim, D.H.; Kim, J.H.; Kim, E.H.; Na, H.K.; Cha, Y.N.; Chung, J.H.; Surh, Y.J. 15-Deoxy-Delta12,14-prostaglandin J2 upregulates the expression of heme oxygenase-1 and subsequently matrix metalloproteinase-1 in human breast cancer cells: Possible roles of iron and ROS. Carcinogenesis 2009, 30, 645–654. [Google Scholar] [CrossRef] [Green Version]
- Tan, X.; Wang, J.; Pang, X.; Liu, L.; Sun, Q.; You, Q.; Tan, F.; Li, N. Indocyanine Green-Loaded Silver Nanoparticle@Polyaniline Core/Shell Theranostic Nanocomposites for Photoacoustic/Near-Infrared Fluorescence Imaging-Guided and Single-Light-Triggered Photothermal and Photodynamic Therapy. ACS Appl. Mater. Interfaces 2016, 8, 34991–35003. [Google Scholar] [CrossRef]
- Wang, Z.X.; Chen, C.Y.; Wang, Y.; Li, F.X.Z.; Huang, J.; Luo, Z.W.; Rao, S.S.; Tan, Y.J.; Liu, Y.W.; Yin, H.; et al. Angstrom-Scale Silver Particles as a Promising Agent for Low-Toxicity Broad-Spectrum Potent Anticancer Therapy. Adv. Funct. Mater. 2019, 29, 1808556. [Google Scholar] [CrossRef]
- Haase, A.; Rott, S.; Mantion, A.; Graf, P.; Plendl, J.; Thunemann, A.F.; Meier, W.P.; Taubert, A.; Luch, A.; Reiser, G. Effects of silver nanoparticles on primary mixed neural cell cultures: Uptake, oxidative stress and acute calcium responses. Toxicol. Sci. 2012, 126, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, K.K.; Awasthi, A.; Kumar, N.; Roy, P.; Awasthi, K.; John, P.J. Silver nanoparticle induced cytotoxicity, oxidative stress, and DNA damage in CHO cells. J. Nanopart. Res. 2013, 15, 1898. [Google Scholar] [CrossRef]
- Cheng, X.L.; Zhang, W.Q.; Ji, Y.L.; Meng, J.; Guo, H.; Liu, J.; Wu, X.C.; Xu, H.Y. Revealing silver cytotoxicity using Au nanorods/Ag shell nanostructures: Disrupting cell membrane and causing apoptosis through oxidative damage. Rsc Adv. 2013, 3, 2296–2305. [Google Scholar] [CrossRef]
- AshaRani, P.V.; Low Kah Mun, G.; Hande, M.P.; Valiyaveettil, S. Cytotoxicity and genotoxicity of silver nanoparticles in human cells. ACS Nano 2009, 3, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Aziz Mousavi, S.M.A.; Mirhosseini, S.A.; Rastegar Shariat Panahi, M.; Mahmoodzadeh Hosseini, H. Characterization of Biosynthesized Silver Nanoparticles Using Lactobacillus rhamnosus GG and its in Vitro Assessment Against Colorectal Cancer Cells. Probiotics Antimicrob. Proteins 2019, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kaba, S.I.; Egorova, E.M. In vitro studies of the toxic effects of silver nanoparticles on HeLa and U937 cells. Nanotechnol. Sci. Appl. 2015, 8, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Azizi, M.; Ghourchian, H.; Yazdian, F.; Bagherifam, S.; Bekhradnia, S.; Nystrom, B. Anti-cancerous effect of albumin coated silver nanoparticles on MDA-MB 231 human breast cancer cell line. Sci. Rep. 2017, 7, 5178. [Google Scholar] [CrossRef]
- Kensler, T.W.; Wakabayash, N.; Biswal, S. Cell survival responses to environmental stresses via the Keap1-Nrf2-ARE pathway. Annu. Rev. Pharmacol. 2007, 47, 89–116. [Google Scholar] [CrossRef]
- Jarak, I.; Carrola, J.; Barros, A.S.; Gil, A.M.; Pereira, M.L.; Corvo, M.L.; Duarte, I.F. From the Cover: Metabolism Modulation in Different Organs by Silver Nanoparticles: An NMR Metabolomics Study of a Mouse Model. Toxicol. Sci. 2017, 159, 422–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, G.B.; Friman, T.; Pang, H.B.; Pallaoro, A.; Hurtado de Mendoza, T.; Willmore, A.M.; Kotamraju, V.R.; Mann, A.P.; She, Z.G.; Sugahara, K.N.; et al. Etchable plasmonic nanoparticle probes to image and quantify cellular internalization. Nat. Mater. 2014, 13, 904–911. [Google Scholar] [CrossRef] [Green Version]
- Nallathamby, P.D.; Xu, X.H. Study of cytotoxic and therapeutic effects of stable and purified silver nanoparticles on tumor cells. Nanoscale 2010, 2, 942–952. [Google Scholar] [CrossRef] [PubMed]
- Foldbjerg, R.; Olesen, P.; Hougaard, M.; Dang, D.A.; Hoffmann, H.J.; Autrup, H. PVP-coated silver nanoparticles and silver ions induce reactive oxygen species, apoptosis and necrosis in THP-1 monocytes. Toxicol. Lett. 2009, 190, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.G.; Zhang, S.; Hwang, J.Y.; Kong, I.K. Silver Nanoparticles Potentiates Cytotoxicity and Apoptotic Potential of Camptothecin in Human Cervical Cancer Cells. Oxid. Med. Cell. Longev. 2018, 2018, 6121328. [Google Scholar] [CrossRef] [PubMed]
- Kocanova, S.; Buytaert, E.; Matroule, J.Y.; Piette, J.; Golab, J.; de Witte, P.; Agostinis, P. Induction of heme-oxygenase 1 requires the p38MAPK and PI3K pathways and suppresses apoptotic cell death following hypericin-mediated photodynamic therapy. Apoptosis 2007, 12, 731–741. [Google Scholar] [CrossRef]
- Thapa, R.K.; Kim, J.H.; Jeong, J.H.; Shin, B.S.; Choi, H.G.; Yong, C.S.; Kim, J.O. Silver nanoparticle-embedded graphene oxide-methotrexate for targeted cancer treatment. Colloids Surf. B Biointerfaces 2017, 153, 95–103. [Google Scholar] [CrossRef]
- Ramezani, T.; Nabiuni, M.; Baharara, J.; Parivar, K.; Namvar, F. Sensitization of Resistance Ovarian Cancer Cells to Cisplatin by Biogenic Synthesized Silver Nanoparticles through p53 Activation. Iran. J. Pharm. Res. 2019, 18, 222–231. [Google Scholar] [PubMed]
- Waugh, D.J.; Wilson, C. The interleukin-8 pathway in cancer. Clin. Cancer Res. 2008, 14, 6735–6741. [Google Scholar] [CrossRef] [Green Version]
- Park, E.J.; Yi, J.; Kim, Y.; Choi, K.; Park, K. Silver nanoparticles induce cytotoxicity by a Trojan-horse type mechanism. Toxicol. In Vitro 2010, 24, 872–878. [Google Scholar] [CrossRef]
- Sun, X.; Shi, J.; Zou, X.; Wang, C.; Yang, Y.; Zhang, H. Silver nanoparticles interact with the cell membrane and increase endothelial permeability by promoting VE-cadherin internalization. J. Hazard. Mater. 2016, 317, 570–578. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Huang, Z.; Wu, H.; Zhou, W.; Jin, P.; Wei, P.; Zhang, Y.; Zheng, F.; Zhang, J.; Xu, J.; et al. Inhibition of autophagy enhances the anticancer activity of silver nanoparticles. Autophagy 2014, 10, 2006–2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarre, A.; Gabrielli, J.; Vial, G.; Leverve, X.M.; Assimacopoulos-Jeannet, F. Reactive oxygen species are produced at low glucose and contribute to the activation of AMPK in insulin-secreting cells. Free Radic. Biol. Med. 2012, 52, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Raja, G.; Kim, S.; Yoon, D.; Yoon, C.; Kim, S. H-1-NMR-based Metabolomics Studies of the Toxicity of Mesoporous Carbon Nanoparticles in Zebrafish (Danio rerio). B Korean Chem. Soc. 2017, 38, 271–277. [Google Scholar] [CrossRef]
- Raja, G.; Kim, S.; Yoon, D.; Yoon, C.; Kim, S. H-1 NMR Based Metabolomics Studies of the Toxicity of Titanium Dioxide Nanoparticles in Zebrafish (Danio rerio). B Korean Chem. Soc. 2018, 39, 33–39. [Google Scholar] [CrossRef]
- Carrola, J.; Pinto, R.J.B.; Nasirpour, M.; Freire, C.S.R.; Gil, A.M.; Santos, C.; Oliveira, H.; Duarte, I.F. NMR Metabolomics Reveals Metabolism-Mediated Protective Effects in Liver (HepG2) Cells Exposed to Subtoxic Levels of Silver Nanoparticles. J. Proteome Res. 2018, 17, 1636–1646. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, Z.; Xu, M.; Wang, X.; Liu, R.; Liu, Q.; Zhang, Z.; Xia, T.; Zhao, J.; Jiang, G.; et al. Nanosilver incurs an adaptive shunt of energy metabolism mode to glycolysis in tumor and nontumor cells. ACS Nano 2014, 8, 5813–5825. [Google Scholar] [CrossRef]
- Carrola, J.; Bastos, V.; Ferreira de Oliveira, J.M.; Oliveira, H.; Santos, C.; Gil, A.M.; Duarte, I.F. Insights into the impact of silver nanoparticles on human keratinocytes metabolism through NMR metabolomics. Arch. Biochem. Biophys. 2016, 589, 53–61. [Google Scholar] [CrossRef]
- Avalos, A.; Haza, A.I.; Mateo, D.; Morales, P. Cytotoxicity and ROS production of manufactured silver nanoparticles of different sizes in hepatoma and leukemia cells. J. Appl. Toxicol. 2014, 34, 413–423. [Google Scholar] [CrossRef]
- Mohammadniaei, M.; Yoon, J.; Choi, H.K.; Placide, V.; Bharate, B.G.; Lee, T.; Choi, J.W. Multifunctional Nanobiohybrid Material Composed of Ag@Bi2Se3/RNA Three-Way Junction/miRNA/Retinoic Acid for Neuroblastoma Differentiation. ACS Appl. Mater. Interfaces 2019, 11, 8779–8788. [Google Scholar] [CrossRef]
- Que, Y.M.; Fan, X.Q.; Lin, X.J.; Jiang, X.L.; Hu, P.P.; Tong, X.Y.; Tan, Q.Y. Size dependent anti-invasiveness of silver nanoparticles in lung cancer cells. Rsc Adv. 2019, 9, 21134–21138. [Google Scholar] [CrossRef] [Green Version]
- Gopinath, P.; Gogoi, S.K.; Sanpui, P.; Paul, A.; Chattopadhyay, A.; Ghosh, S.S. Signaling gene cascade in silver nanoparticle induced apoptosis. Colloids Surf. B Biointerfaces 2010, 77, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Panzarini, E.; Mariano, S.; Vergallo, C.; Carata, E.; Fimia, G.M.; Mura, F.; Rossi, M.; Vergaro, V.; Ciccarella, G.; Corazzari, M.; et al. Glucose capped silver nanoparticles induce cell cycle arrest in HeLa cells. Toxicol. In Vitro 2017, 41, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Mytych, J.; Zebrowski, J.; Lewinska, A.; Wnuk, M. Prolonged Effects of Silver Nanoparticles on p53/p21 Pathway-Mediated Proliferation, DNA Damage Response, and Methylation Parameters in HT22 Hippocampal Neuronal Cells. Mol. Neurobiol. 2017, 54, 1285–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.F.; Shen, W.; Gurunathan, S. Silver Nanoparticle-Mediated Cellular Responses in Various Cell Lines: An in Vitro Model. Int. J. Mol. Sci. 2016, 17, 1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velichkova, M.; Hasson, T. Keap1 regulates the oxidation-sensitive shuttling of Nrf2 into and out of the nucleus via a Crm1-dependent nuclear export mechanism. Mol. Cell. Biol. 2005, 25, 4501–4513. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, T.; Shimizu, H.; Morimatsu, H.; Maeshima, K.; Inoue, K.; Akagi, R.; Matsumi, M.; Katayama, H.; Morita, K. Heme Oxygenase-1 is an Essential Cytoprotective Component in Oxidative Tissue Injury Induced by Hemorrhagic Shock. J. Clin. Biochem. Nutr. 2009, 44, 28–40. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Fan, W.G.; Li, D.P.; Kung, H.F.; Lin, M.C.M. Heme oxygenase-1 system and gastrointestinal inflammation: A short review. World J. Gastroenterol. 2011, 17, 4283–4288. [Google Scholar] [CrossRef] [Green Version]
- Engin, A.B.; Nikitovic, D.; Neagu, M.; Henrich-Noack, P.; Docea, A.O.; Shtilman, M.I.; Golokhvast, K.; Tsatsakis, A.M. Mechanistic understanding of nanoparticles’ interactions with extracellular matrix: The cell and immune system. Part. Fibre Toxicol. 2017, 14, 22. [Google Scholar] [CrossRef]
- Dejana, E. Endothelial cell-cell junctions: Happy together. Nat. Rev. Mol. Cell Biol. 2004, 5, 261–270. [Google Scholar] [CrossRef]
- Setyawati, M.I.; Tay, C.Y.; Chia, S.L.; Goh, S.L.; Fang, W.; Neo, M.J.; Chong, H.C.; Tan, S.M.; Loo, S.C.J.; Ng, K.W.; et al. Titanium dioxide nanomaterials cause endothelial cell leakiness by disrupting the homophilic interaction of VE-cadherin. Nat. Commun. 2013, 4, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.D.; Mizushima, N. Autophagy and human diseases. Cell Res. 2014, 24, 69–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zabirnyk, O.; Yezhelyev, M.; Seleverstov, O. Nanoparticles as a novel class of autophagy activators. Autophagy 2007, 3, 278–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dekkers, S.; Williams, T.D.; Zhang, J.K.; Zhou, J.R.; Vandebriel, R.J.; De la Fonteyne, L.J.J.; Gremmer, E.R.; He, S.; Guggenheim, E.J.; Lynch, I.; et al. Multi-omics approaches confirm metal ions mediate the main toxicological pathways of metal-bearing nanoparticles in lung epithelial A549 cells. Environ. Sci-Nano 2018, 5, 1506–1517. [Google Scholar] [CrossRef] [Green Version]
- Raja, G.; Jang, Y.K.; Suh, J.S.; Prabhakaran, V.S.; Kim, T.J. Advanced understanding of genetic risk and metabolite signatures in construction workers via cytogenetics and metabolomics analysis. Process. Biochem. 2019, 86, 117–126. [Google Scholar] [CrossRef]
- Smolkova, K.; Bellance, N.; Scandurra, F.; Genot, E.; Gnaiger, E.; Plecita-Hlavata, L.; Jezek, P.; Rossignol, R. Mitochondrial bioenergetic adaptations of breast cancer cells to aglycemia and hypoxia. J. Bioenerg. Biomembr. 2010, 42, 55–67. [Google Scholar] [CrossRef]
- Moruzzi, N.; Del Sole, M.; Fato, R.; Gerdes, J.M.; Berggren, P.O.; Bergamini, C.; Brismar, K. Short and prolonged exposure to hyperglycaemia in human fibroblasts and endothelial cells: Metabolic and osmotic effects. Int. J. Biochem. Cell B 2014, 53, 66–76. [Google Scholar] [CrossRef]
- Zheng, J. Energy metabolism of cancer: Glycolysis versus oxidative phosphorylation (Review). Oncol. Lett. 2012, 4, 1151–1157. [Google Scholar] [CrossRef] [Green Version]
- Suchorolski, M.T.; Paulson, T.G.; Sanchez, C.A.; Hockenbery, D.; Reid, B.J. Warburg and Crabtree Effects in Premalignant Barrett’s Esophagus Cell Lines with Active Mitochondria. PLoS ONE 2013, 8, e56884. [Google Scholar] [CrossRef] [Green Version]
- Hagman, A.; Sall, T.; Piskur, J. Analysis of the yeast short-term Crabtree effect and its origin. FEBS J. 2014, 281, 4805–4814. [Google Scholar] [CrossRef] [Green Version]
- Knijn, A.; Brisdelli, F.; Ferretti, A.; Iorio, E.; Marcheggiani, D.; Bozzi, A. Metabolic alterations in K562 cells exposed to taxol and tyrphostin AG957: 1H NMR and biochemical studies. Cell Biol. Int. 2005, 29, 890–897. [Google Scholar] [CrossRef] [PubMed]
- Bury, N.R.; Grosell, M.; Grover, A.K.; Wood, C.M. ATP-dependent silver transport across the basolateral membrane of rainbow trout gills. Toxicol. Appl. Pharm. 1999, 159, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheline, C.T.; Behrens, M.M.; Choi, D.W. Zinc-induced cortical neuronal death: Contribution of energy failure attributable to loss of NAD (+) and inhibition of glycolysis. J. Neurosci. 2000, 20, 3139–3146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrola, J.; Bastos, V.; Jarak, I.; Oliveira-Silva, R.; Malheiro, E.; Daniel-da-Silva, A.L.; Oliveira, H.; Santos, C.; Gil, A.M.; Duarte, I.F. Metabolomics of silver nanoparticles toxicity in HaCaT cells: Structure-activity relationships and role of ionic silver and oxidative stress. Nanotoxicology 2016, 10, 1105–1117. [Google Scholar] [CrossRef]
- Chichova, M.; Shkodrova, M.; Vasileva, P.; Kirilova, K.; Doncheva-Stoimenova, D. Influence of silver nanoparticles on the activity of rat liver mitochondrial ATPase. J. Nanopart. Res. 2014, 16, 2243. [Google Scholar] [CrossRef]
- Costa, C.; Ronconi, J.V.V.; Daufenbach, J.F.; Goncalves, C.L.; Rezin, G.T.; Streck, E.L.; Paula, M.M.D. In vitro effects of silver nanoparticles on the mitochondrial respiratory chain. Mol. Cell. Biochem. 2010, 342, 51–56. [Google Scholar] [CrossRef]
- Teodoro, J.S.; Simoes, A.M.; Duarte, F.V.; Rolo, A.P.; Murdoch, R.C.; Hussain, S.M.; Palmeira, C.M. Assessment of the toxicity of silver nanoparticles in vitro: A mitochondrial perspective. Toxicol. In Vitro 2011, 25, 664–670. [Google Scholar] [CrossRef]
- Jiao, G.Z.; Li, X.; Zhang, N.; Qiu, J.Q.; Xu, H.Y.; Liu, S.M. Metabolomics study on the cytotoxicity of graphene. Rsc Adv. 2014, 4, 44712–44717. [Google Scholar] [CrossRef]
- Alia, M.; Ramos, S.; Mateos, R.; Bravo, L.; Goya, L. Response of the antioxidant Defense system to tert-butyl hydroperoxide and hydrogen peroxide in a human hepatoma cell line (HepG2). J. Biochem. Mol. Toxic. 2005, 19, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Montalvo-Quiros, S.; Aragoneses-Cazorla, G.; Garcia-Alcalde, L.; Vallet-Regi, M.; Gonzalez, B.; Luque-Garcia, J.L. Cancer cell targeting and therapeutic delivery of silver nanoparticles by mesoporous silica nanocarriers: insights into the action mechanisms using quantitative proteomics. Nanoscale 2019, 11, 4531–4545. [Google Scholar] [CrossRef]
- Arora, S.J.; Jain, J.M.; Rajwade, K.M. Paknikar, Interactions of silver nanoparticles with primary mouse fibroblasts and liver cells. Toxicol. Appl. Pharmacol. 2009, 236, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Vrcek, I.V.; Zuntar, I.; Petlevski, R.; Pavicic, I.; Dutour Sikiric, M.; Curlin, M.; Goessler, W. Comparison of in vitro toxicity of silver ions and silver nanoparticles on human hepatoma cells. Environ. Toxicol. 2016, 31, 679–692. [Google Scholar] [CrossRef] [PubMed]
- Avalos Funez, A.; Isabel Haza, A.; Mateo, D.; Morales, P. In vitro evaluation of silver nanoparticles on human tumoral and normal cells. Toxicol. Mech. Methods 2013, 23, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, J.; Levine, B.; Debnath, J. Autophagy and cancer metabolism. Methods Enzymol. 2014, 542, 25–57. [Google Scholar] [PubMed] [Green Version]
- Mishra, A.R.; Zheng, J.; Tang, X.; Goering, P.L. Silver Nanoparticle-Induced Autophagic-Lysosomal Disruption and NLRP3-Inflammasome Activation in HepG2 Cells Is Size-Dependent. Toxicol. Sci. 2016, 150, 473–487. [Google Scholar] [CrossRef] [PubMed]
- Chugh, H.; Sood, D.; Chandra, I.; Tomar, V.; Dhawan, G.; Chandra, R. Role of gold and silver nanoparticles in cancer nano-medicine. Artif Cells Nanomed Biotechnol. 2018, 46 (Suppl. S1), 1210–1220. [Google Scholar] [CrossRef] [Green Version]
- Hammoudi, N.; Ahmed, K.B.; Garcia-Prieto, C.; Huang, P. Metabolic alterations in cancer cells and therapeutic implications. Chin. J. Cancer 2011, 30, 508–525. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.F.; Carman, G.M. CTP synthetase and its role in phospholipid synthesis in the yeast Saccharomyces cerevisiae. Prog. Lipid Res. 2008, 47, 333–339. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Dong, W.; Liu, R.; Wang, Y.; Li, Y. Research on the hepatotoxicity mechanism of citrate-modified silver nanoparticles based on metabolomics and proteomics. Nanotoxicology 2018, 12, 18–31. [Google Scholar] [CrossRef]
- Lindeque, J.Z.; Matthyser, A.; Mason, S.; Louw, R.; Taute, C.J.F. Metabolomics reveals the depletion of intracellular metabolites in HepG2 cells after treatment with gold nanoparticles. Nanotoxicology 2018, 12, 251–262. [Google Scholar] [CrossRef]
- Miranda, R.R.; Gorshkov, V.; Korzeniowska, B.; Kempf, S.J.; Neto, F.F.; Kjeldsen, F. Co-exposure to silver nanoparticles and cadmium induce metabolic adaptation in HepG2 cells. Nanotoxicology 2018, 12, 781–795. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, F.; Wang, X.; Li, W. Toxico-Metabolomics of Engineered Nanomaterials: Progress and Challenges. Adv. Funct. Mater. 2019, 29, 1904268. [Google Scholar] [CrossRef]
- Raja, G.; Cao, S.; Kim, D.H.; Kim, T.J. Mechanoregulation of titanium dioxide nanoparticles in cancer therapy. Mater. Sci. Eng. C 2020, 107, 110303. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, L.; Chen, Q.; Chen, C. Cytotoxic potential of silver nanoparticles. Yonsei Med. J. 2014, 55, 283–291. [Google Scholar] [CrossRef] [Green Version]
- Holmes, A.M.; Lim, J.; Studier, H.; Roberts, M.S. Varying the morphology of silver nanoparticles results in differential toxicity against micro-organisms, HaCaT keratinocytes and affects skin deposition. Nanotoxicology 2016, 10, 1503–1514. [Google Scholar] [CrossRef]
- Mao, B.H.; Tsai, J.C.; Chen, C.W.; Yan, S.J.; Wang, Y.J. Mechanisms of silver nanoparticle-induced toxicity and important role of autophagy. Nanotoxicology 2016, 10, 1021–1040. [Google Scholar] [CrossRef]
- Williams, K.; Milner, J.; Boudreau, M.D.; Gokulan, K.; Cerniglia, C.E.; Khare, S. Effects of subchronic exposure of silver nanoparticles on intestinal microbiota and gut-associated immune responses in the ileum of Sprague-Dawley rats. Nanotoxicology 2015, 9, 279–289. [Google Scholar] [CrossRef]
- Jeong, J.K.; Gurunathan, S.; Kang, M.H.; Han, J.W.; Das, J.; Choi, Y.J.; Kwon, D.N.; Cho, S.G.; Park, C.; Seo, H.G.; et al. Hypoxia-mediated autophagic flux inhibits silver nanoparticle-triggered apoptosis in human lung cancer cells. Sci. Rep. 2016, 6, 21688. [Google Scholar] [CrossRef] [Green Version]
- Johnston, H.J.; Hutchison, G.; Christensen, F.M.; Peters, S.; Hankin, S.; Stone, V. A review of the in vivo and in vitro toxicity of silver and gold particulates: Particle attributes and biological mechanisms responsible for the observed toxicity. Crit. Rev. Toxicol. 2010, 40, 328–346. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
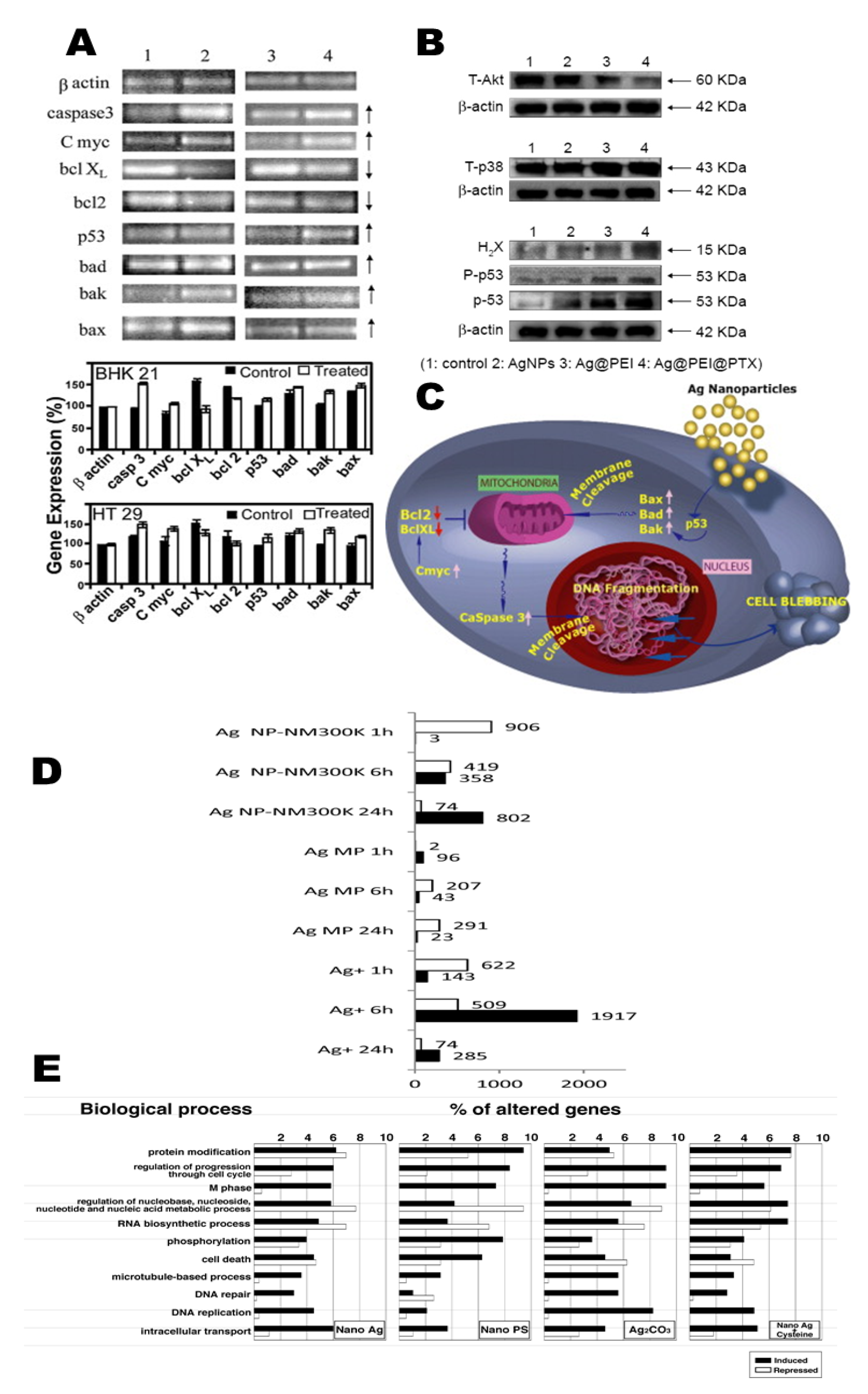{kind=link}
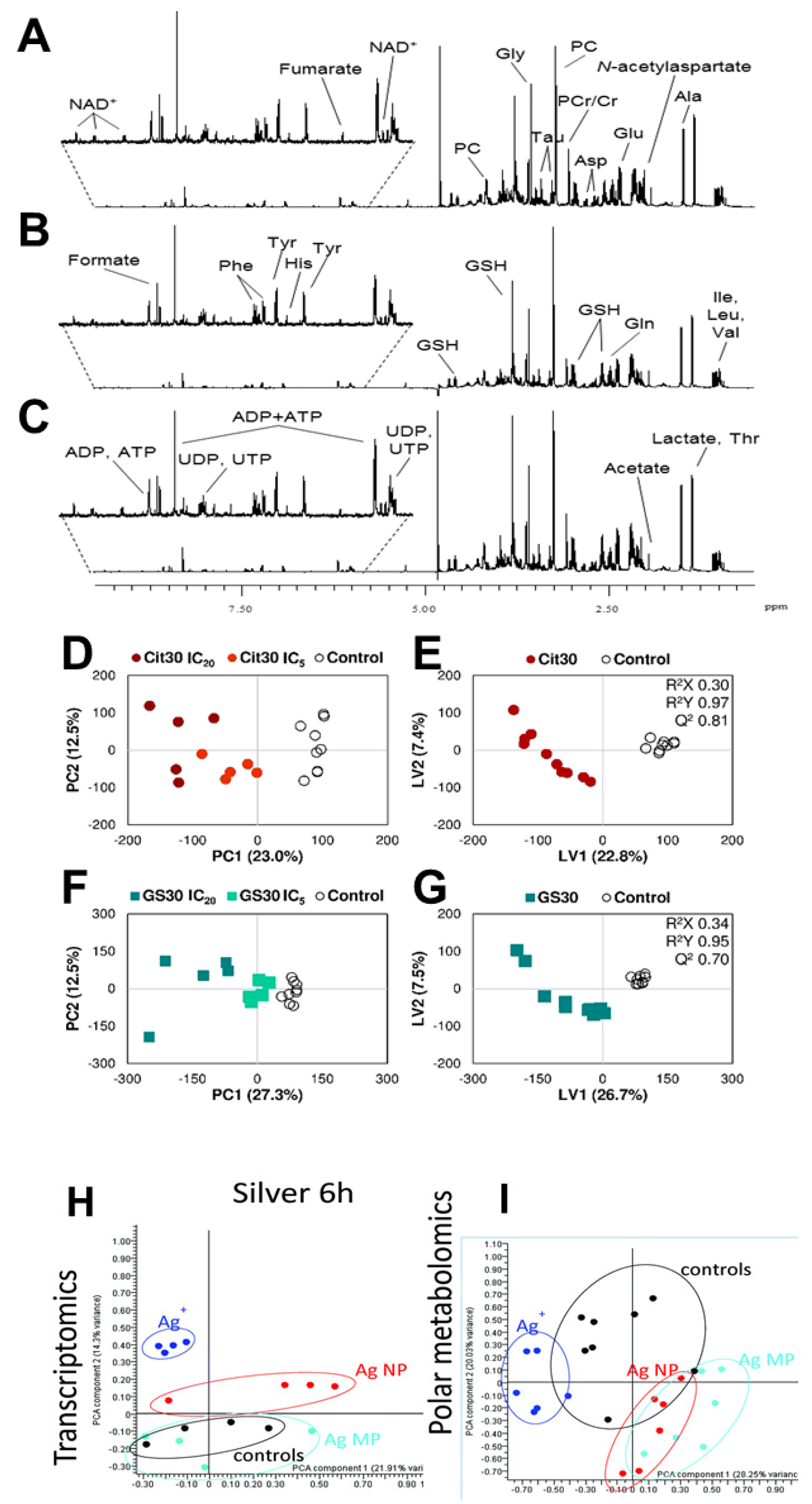{kind=link}
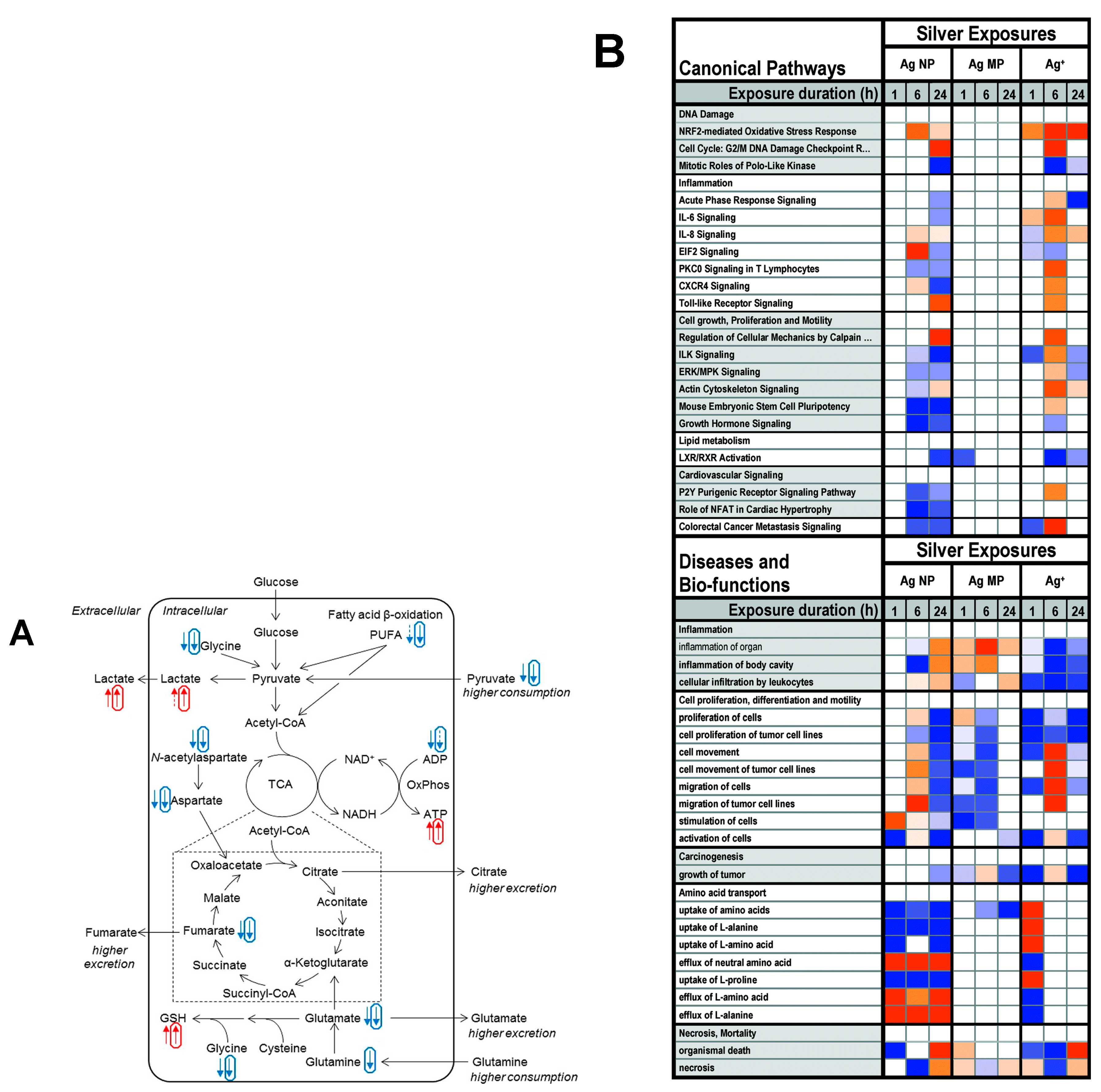{kind=link}
| Year | Cancer Cells | Size (NM) | Concentration | Experimental Time | Mechanobiological Outcomes | Reference |
|---|---|---|---|---|---|---|
| 2009 | Human hepatoma cells (HepG2) | 7–20 | Up to 250 mg/mL | 24 h | Increased glutathione (GSH) level; increased apoptosis (AO-EB staining) and caspase 3 activity | [6] |
| 2009 | A431HT-1080 | 7–20 | 1.56–500 μg/mL | 24 h | Apoptosis induced in both A431 and HT-1080 cell lines | [20] |
| 2009 | Human lung adenocarcinoma (A549 cells) THP-1 monocytes | 30–50 69 | Up to 15 mg/mL | 24 h | ROS generation: formation of DNA adducts Cytotoxicity; ROS; oxidative stress | [22] |
| 2013 | Human lung carcinoma (A549 cells) | <100 | 0, 25, 50, 100, or 200 µg/mL | 24, 48 h | Intracellular ROS levels; cell cycle; proliferating cell nuclear antigen (PCNA) protein expression level | [23] |
| 2011 | Human colon cancer cells (HT29) | 172.6 ± 27.1 | 12–48 μg/mL | 24 h | ROS generation; induced apoptosis (AO-EB staining and annexin V staining); increased DNA fragmentation and expression of caspase 3; mitochondrial membrane depolarization | [26] |
| 2009 | Human hepatoma cells (HepG2) | 5–10 | Up to 2 mg/mL | 24 h | ROS generation and change of oxidative stress-related gene expression (catalase, GPx1, MT1b, and SOD1) Detection of g-H2AX phosphorylation; lactate dehydrogenase (LDH) leakage | [39] |
| 2015 | Human breast cancer cells (MDA-MB-231) | 300 and 600 | 5–25 µg/mL | Targeting p53; Bcl-2; p-p53; ROS; anticancer effects | [43] | |
| 2009 | Human hepatoma cells (HepG2) | 7–10 | 1 mg/mL | 24 h | Increased micronucleus formation | [44] |
| 2009 | PC12 cells | 200 | 2.5–25 µg/mL | 24 h | Gene expression of Gss; Gpx; Txnrd | [45] |
| 2008 | Mouse fibroblast (NIH3T3) and Human colon cancer cells (HCT116) | 1–100 | Up to 100 mg/mL | 72 h | ROS generation; increased apoptosis (annexin V staining); expression of p53 and c-Jun N-terminal kinases (JNK) activation. NIH3T3-induced release of cytochrome c into the cytosol and translocation of Bax gene to mitochondria and decreased Bcl-2 expression | [46] |
| 2011 | Human hepatocyte cell line (L02) | 37.8, 6.7 | Up to 100 mg/mL | 24 h | Increased MDA formation and reduced GSH level and SOD activity; induced DNA breakage (comet assay) | [47] |
| 2010 | Human hepatoma cells (HepG2) | 5.9–3.3, 23.8–6.7, 47.5–22.1 | Up to 50 mg/mL | 24 h | ROS generation; induced apoptosis (annexin V staining and Hoechst 33342 staining) Increased and decreased GSH level; reduced SOD activity; cell cycle arrest in S phase | [48] |
| 2014 | human colon carcinoma cells | 10, 20, 40, 60, and 100 | 0 to 10 µg/mL | 24 h | Intracellular ROS levels; IL-8 release; mitochondrial activity; cell viability; cell proliferation | [49] |
| 2009 | Human carcinoma cell line (HeLa S3) | 2–5 | Up to 120 mg/mL | 3, 4 h | Increased expression of HO-1 and MT-2A expression; induced apoptosis (annexin V staining) | [50] |
| 2016 | Human colorectal carcinoma cell line (T84 cells) | 10, 20, 75, and 110 | 20 and 100 mg/mL | 48 h | Genes related to cell-cell junctions and epithelial barrier functions | [51] |
| 2017 | Human cervical cancer cells (HeLa CCL2) | 420 | 20–100 μg/mL | 24 h | Proapoptotic gene expression, including P53, P21, BAX, BAK, CASP3, and CASP9, | [52] |
| 2016 | Human tongue squamous carcinoma (SCC-25) | 10 | 0.31 to 10 g/mL | 48 h | Reduced proliferation and viability; cytotoxicity; cell cycle arrest; cell morphology analysis; | [53] |
| 2014 | HepG2 cells | 10 and 100 | 2.0 and 4.0 mg/L | 24 h | Non-cytotoxic doses induced p38 MAPK pathway activation and led to the promotion of HepG2 cell proliferation | [54] |
| 2019 | Human breast adenocarcinoma (MCF-7 cells) | 5, 75 | 212 µM | 24, 48 h | ER stress markers; ER calcium levels | [55] |
| 2018 | Human lung adenocarcinoma (A549 cells) | 633 | 10, 40 μg/mL | 24, 48 h | Bcl-2; Bax; caspase-3, 7 expression | [56] |
| 2012 | SK-OV3 cells | 7.5 ± 2.5 | 1, 5, or 10 μg/mL | 24 h | Nrf2 knockdown cells; DNA damage and apoptosis | [57] |
| Omics | PDBe-KB/KEGG | Gene/Proteins | Potential Outcomes | References |
|---|---|---|---|---|
| Genomics and Proteomics | P42574, Q13490, Q8IH92 | Caspase-3, 9; T-Akt; T-p38; P-53 |
| [20,37,38] |
| Q07817 | Bcl-2 |
| ||
| Q8ISJ0, Q07449, P07203 | SOD1, SOD2, GPx-1 |
| [39] | |
| P61604 | HSP60, HSP70 (heat shock proteins) |
| [41,42] | |
| Q07812 | BAX |
| [56] | |
| P06762, A8JBZ0 | HMOX1, HMOX2 |
| [63,70,72] | |
| Q8CHP8 | NA/K ATPase; Pgp; EDEM |
| ||
| Q16611, Q38998, P04049 | Bak, Bax, Bid, Bcl-2, p53, Caspase-3, 6, 9; NF-kB, P38, AKT1, RAF, MEK, |
| [85] | |
| Q9W1M7, P49137 | PI3K, AKT, P38MAPK signaling cascade |
| [86] | |
| Q8IH92 | Tumor protein P53 (TP53), P21, P27 |
| [87,88,89] | |
| P10145, O73909 | IL-8, IL-1beta |
| ||
| P01375 | TNF-α |
| [90] | |
| Q8AYD0 | VE-cadherin; p-VE-cadherin |
| [91] | |
| Q38998 | P-AKT; AKT; LC3-I; LC3-II |
| [92] | |
| Q9BYN0 | SRXN1 (sulfiredoxin 1) |
| [93] | |
| Q16236, P09601 | Nrf2; HO-1 |
| ||
| C00031, C00186, C00158 | Glucose, lactate, citrate |
| [39,84,87,94] | |
| C00157, C00670 | PhosphocholineGlycerophosphocholine |
| ||
| K00016 | LDH leakage |
| ||
| C00051 | Glutathione |
| ||
| C00385, C00262 | Xanthine; hypoxanthine |
| ||
| C01104, C00051 | Trimethylamine N-oxide; GSH |
| [95] | |
| Metabolomics | C00008, C00002, C00003 | ADP, ATP, NAD+ |
| [96] |
| C00422, C00187 | Triglyceride; Cholesterol |
| [97] | |
| C00022, C0004, C00122 | Pyruvate, succinate, fumarate, |
| [98] | |
| C00041, C00183, C00079 | Alanine, valine, phenylalanine, |
| ||
| C00064, C00025 | Glutamine, glutamate, |
| ||
| C00137 | Myo-inositol |
| ||
| C00051, Q8ISJ0 | GSH, SOD; lipid peroxidation; LDH |
| [20,99] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raja, G.; Jang, Y.-K.; Suh, J.-S.; Kim, H.-S.; Ahn, S.H.; Kim, T.-J. Microcellular Environmental Regulation of Silver Nanoparticles in Cancer Therapy: A Critical Review. Cancers 2020, 12, 664. https://doi.org/10.3390/cancers12030664
Raja G, Jang Y-K, Suh J-S, Kim H-S, Ahn SH, Kim T-J. Microcellular Environmental Regulation of Silver Nanoparticles in Cancer Therapy: A Critical Review. Cancers. 2020; 12(3):664. https://doi.org/10.3390/cancers12030664
Chicago/Turabian StyleRaja, Ganesan, Yoon-Kwan Jang, Jung-Soo Suh, Heon-Su Kim, Sang Hyun Ahn, and Tae-Jin Kim. 2020. "Microcellular Environmental Regulation of Silver Nanoparticles in Cancer Therapy: A Critical Review" Cancers 12, no. 3: 664. https://doi.org/10.3390/cancers12030664
APA StyleRaja, G., Jang, Y. -K., Suh, J. -S., Kim, H. -S., Ahn, S. H., & Kim, T. -J. (2020). Microcellular Environmental Regulation of Silver Nanoparticles in Cancer Therapy: A Critical Review. Cancers, 12(3), 664. https://doi.org/10.3390/cancers12030664