Interleukins 4 and 13 and Their Receptors Are Differently Expressed in Gastrointestinal Tract Cancers, Depending on the Anatomical Site and Disease Advancement, and Improve Colon Cancer Cell Viability and Motility

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. Local Expression of Interleukins and Their Receptors
2.1.1. Characteristics of Study Population
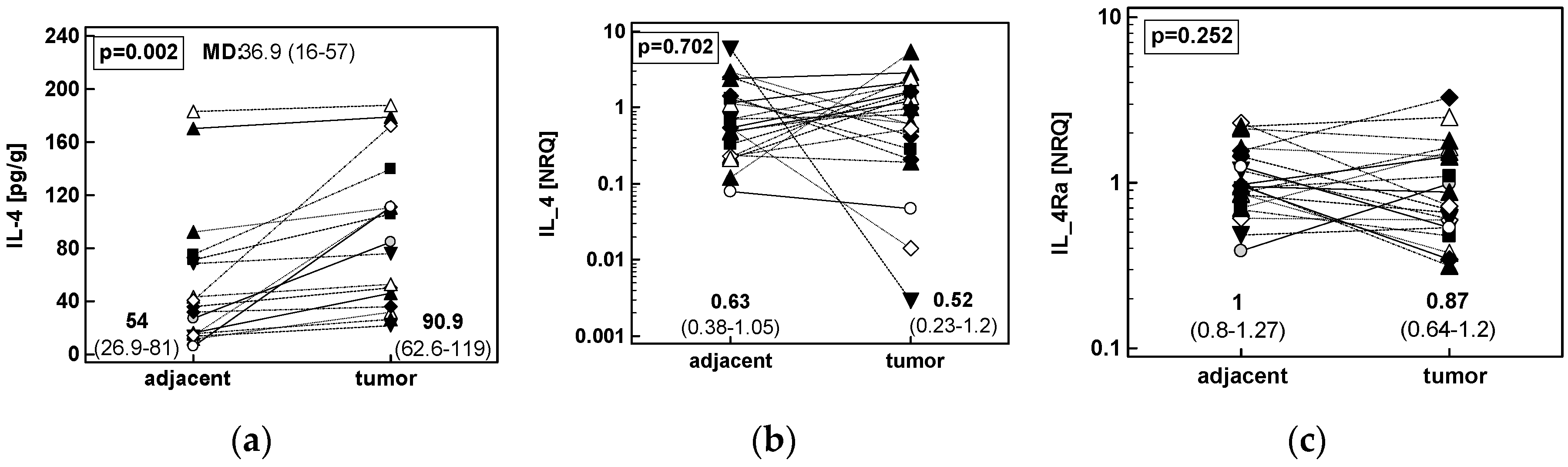
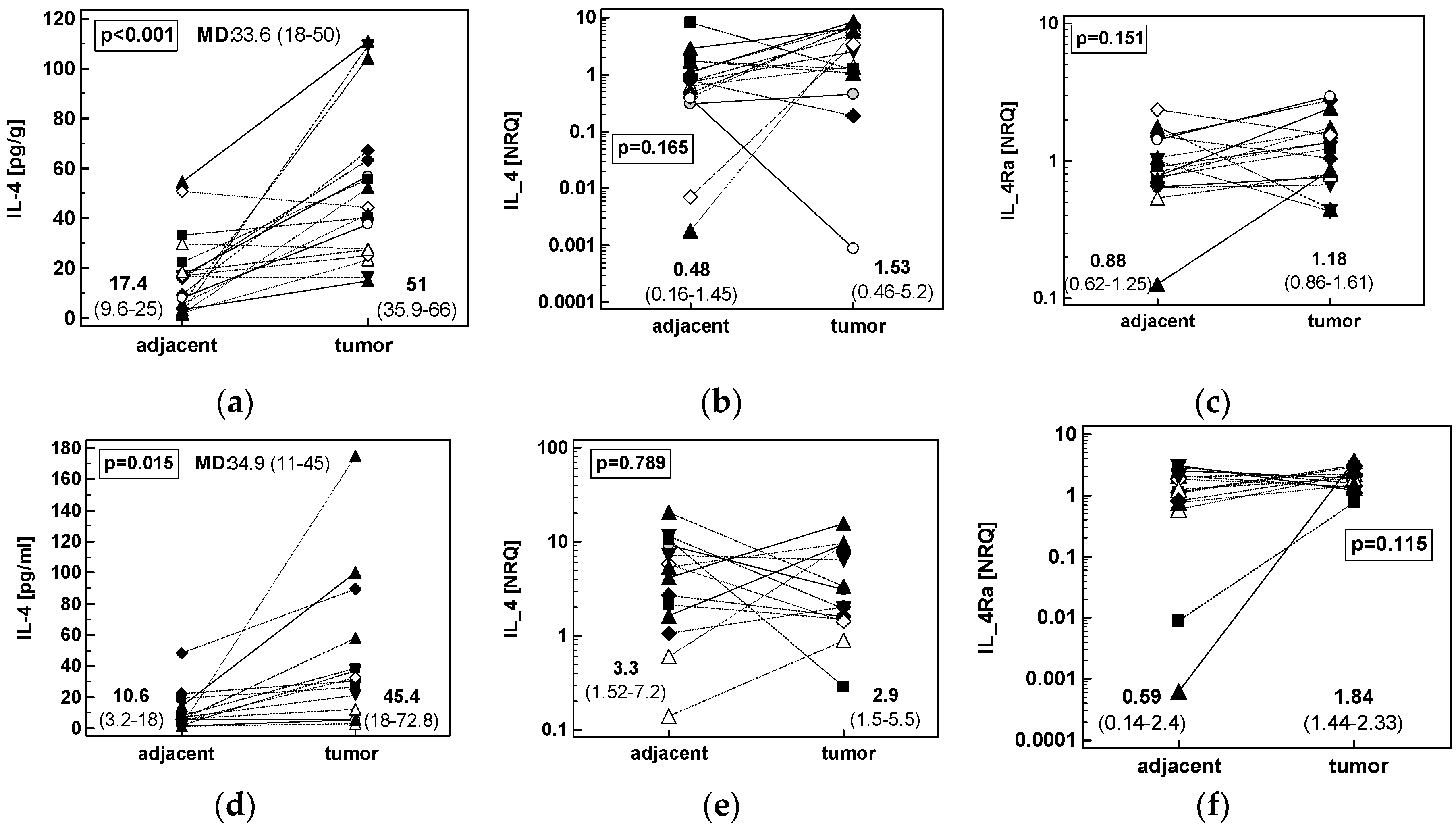
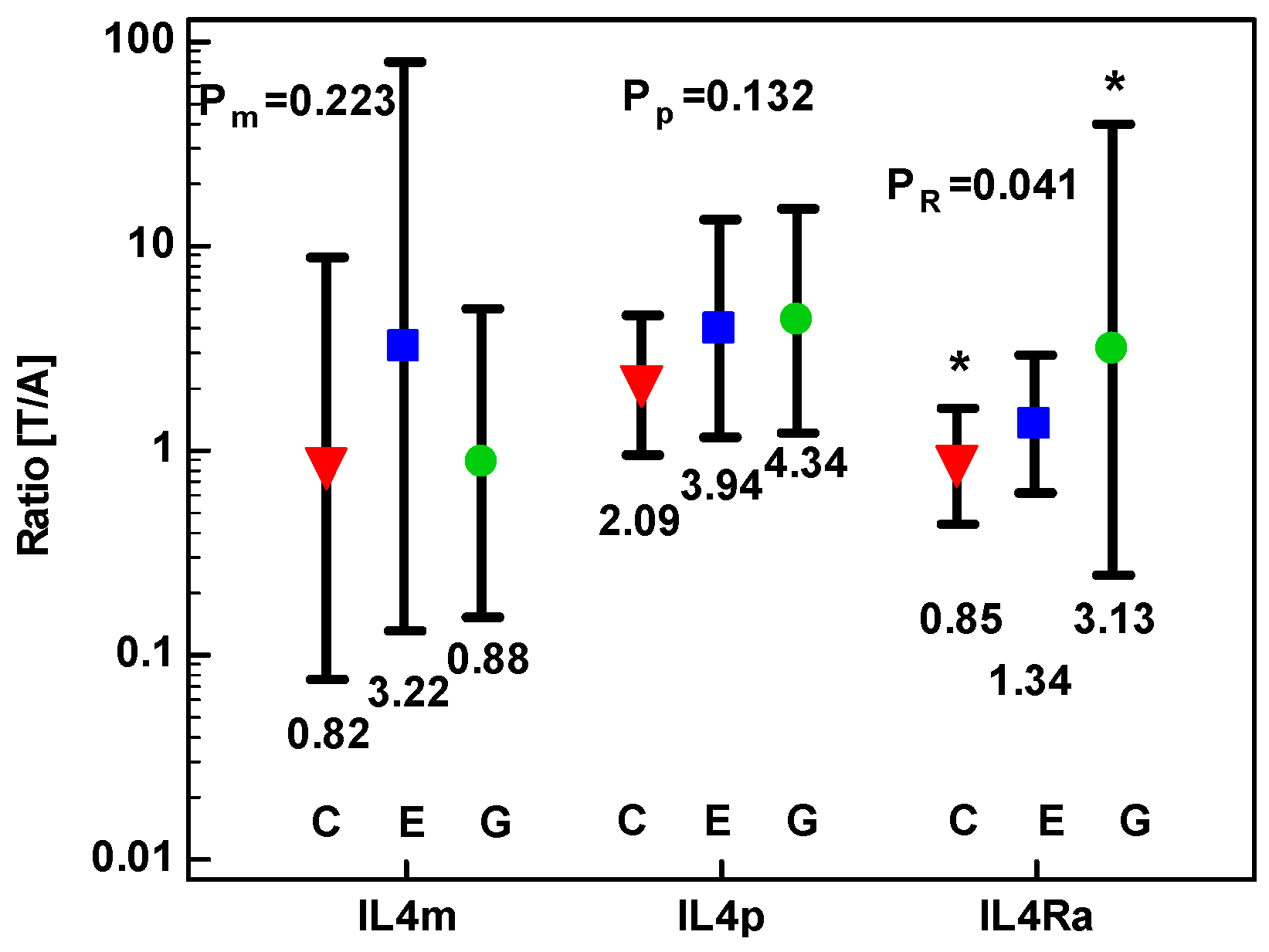
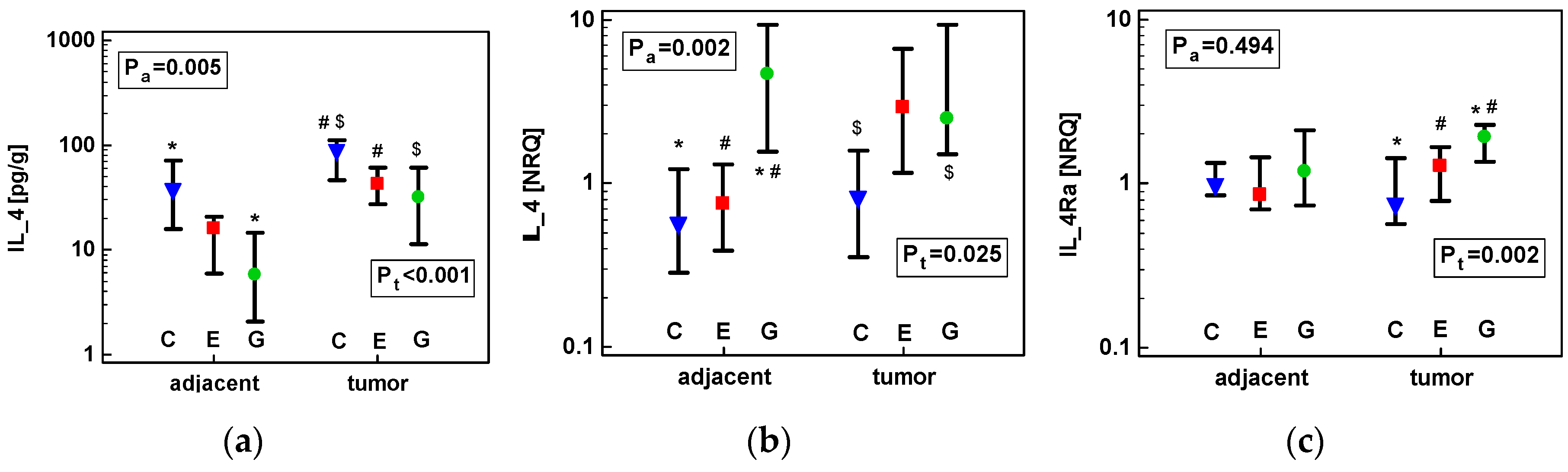
2.1.2. Concentration of IL-4 Protein and Expression of IL4 and IL4Ra Transcripts in CRC as Compared to Upper GIT Cancers
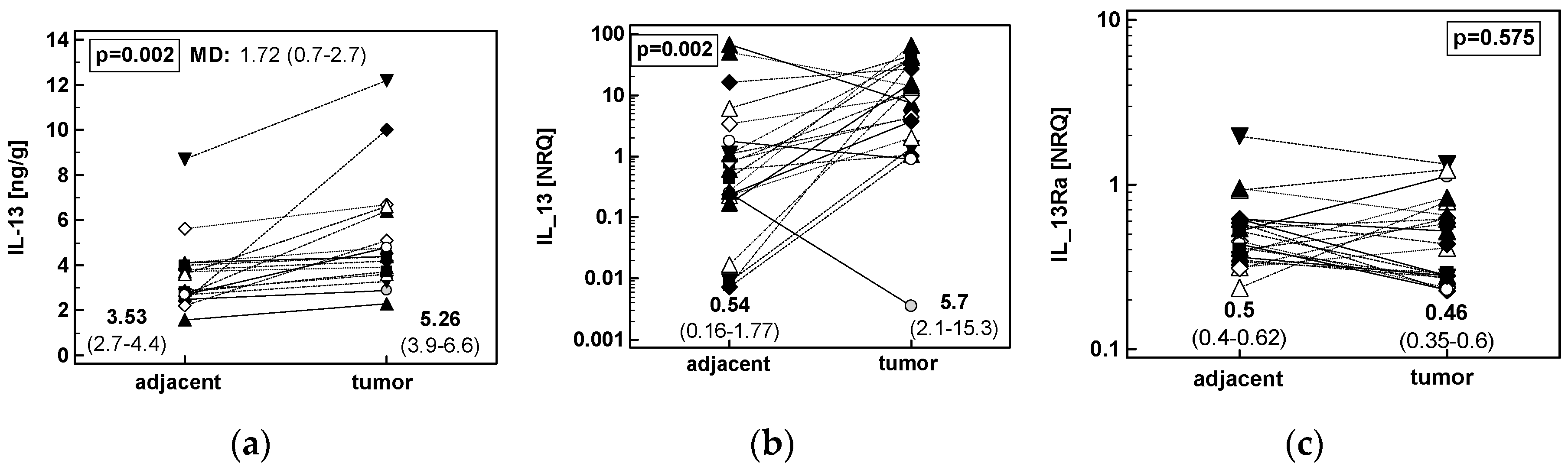
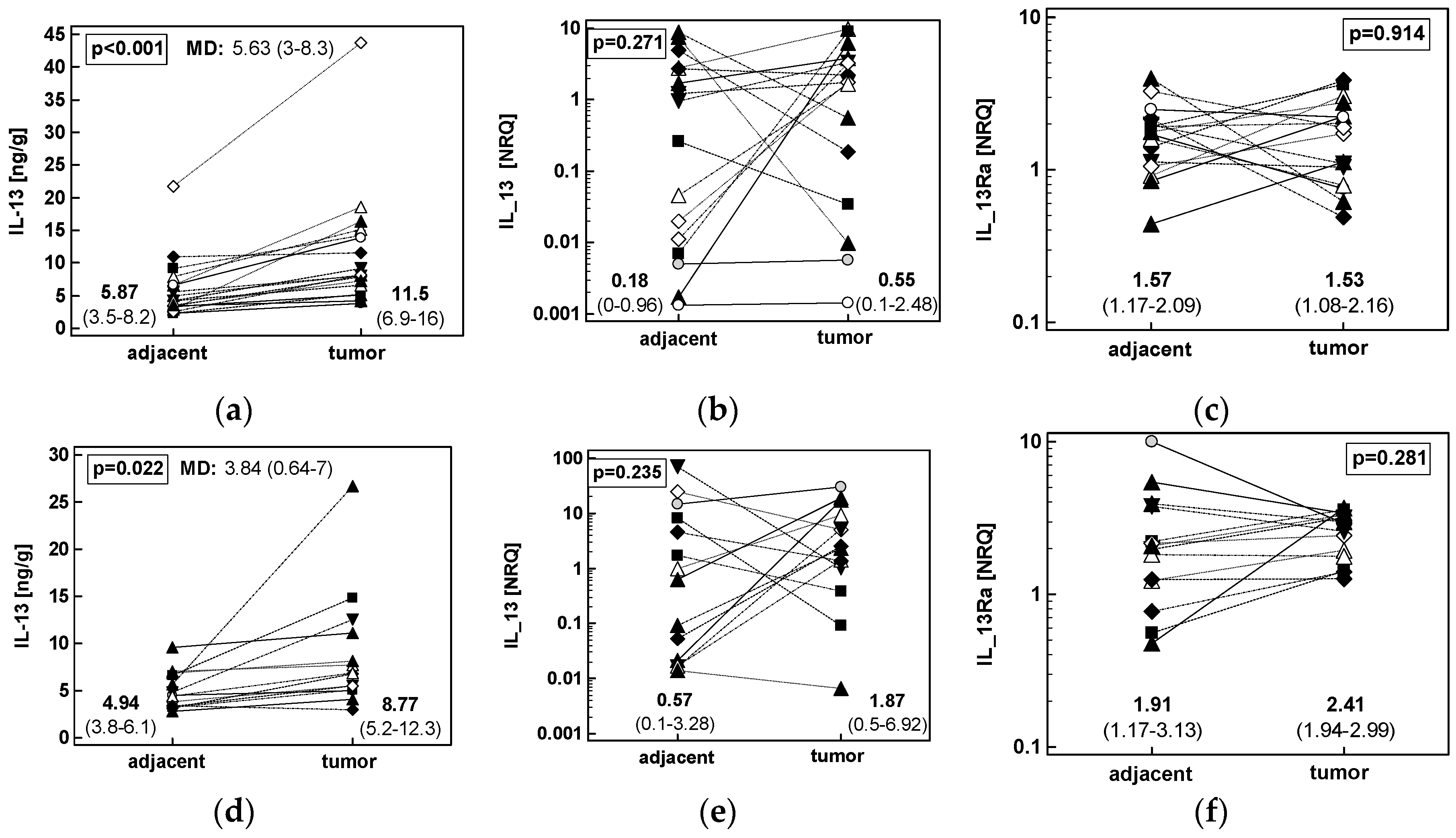
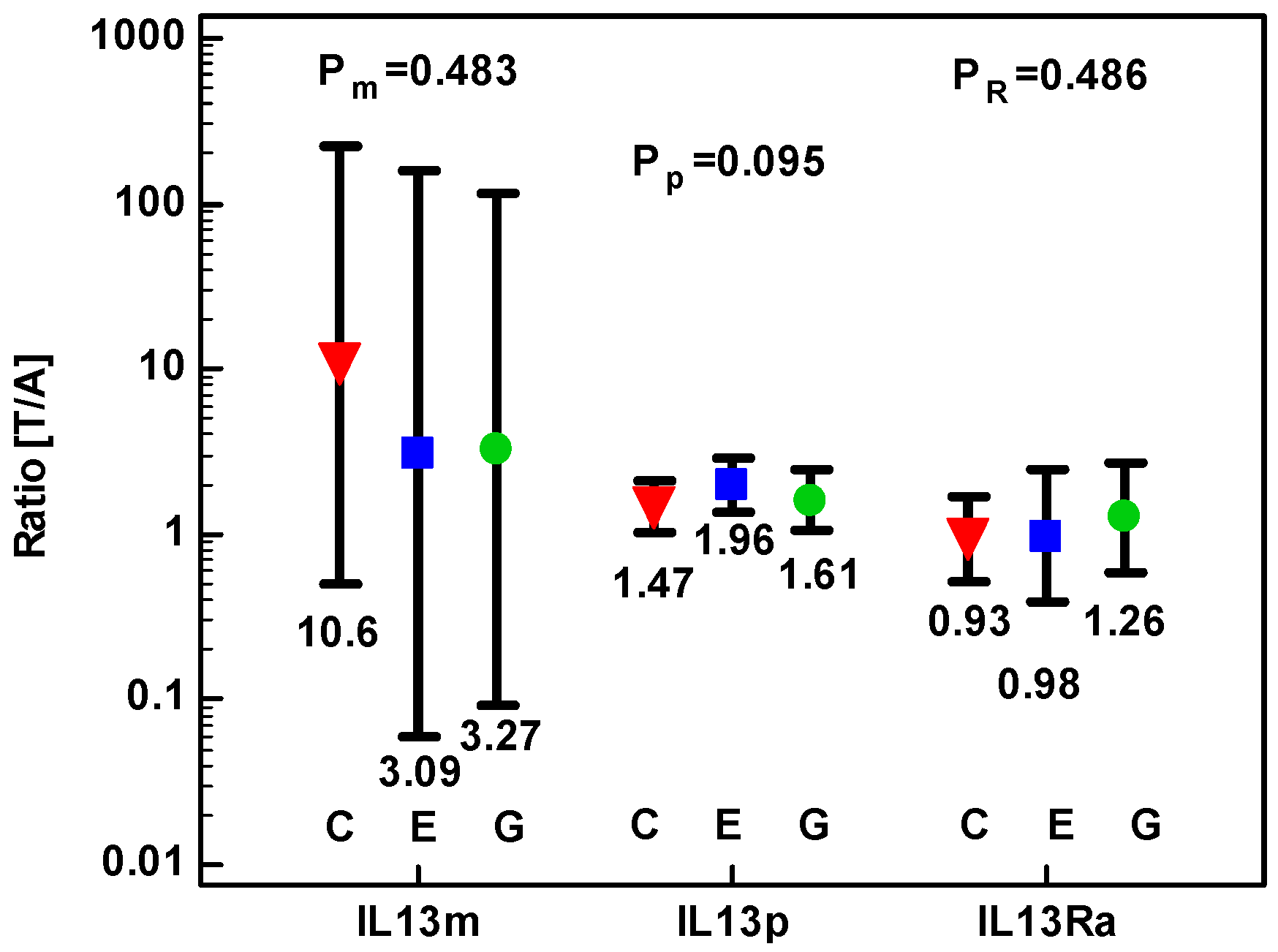
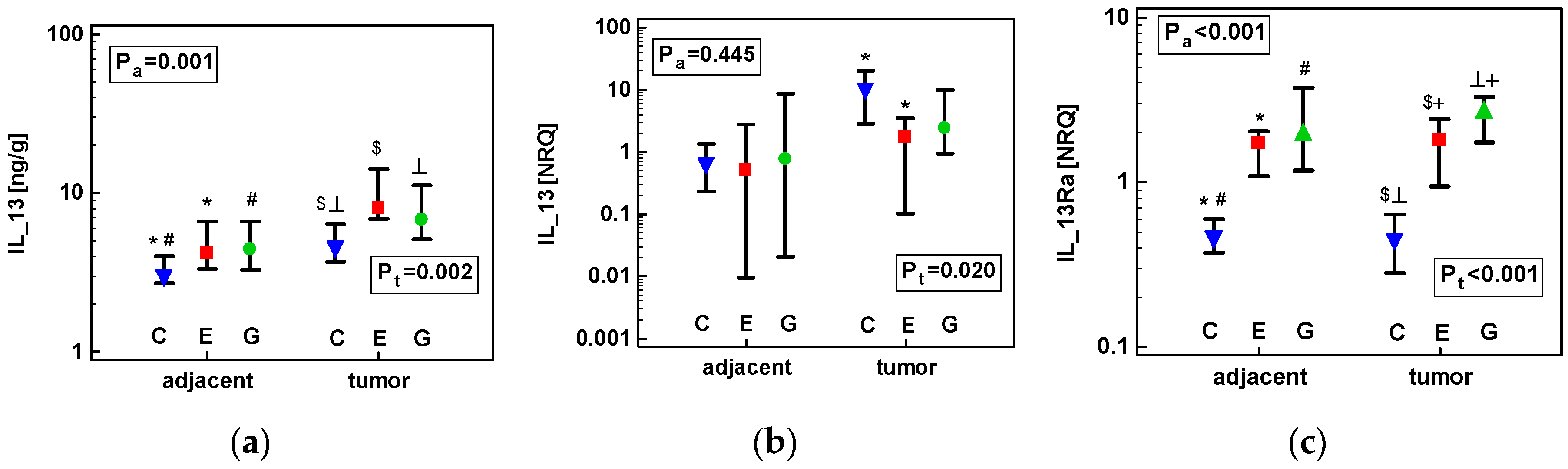
2.1.3. Concentration of IL-13 Protein and Expression of IL13 and IL13Ra1 Transcripts in CRC as Compared to upper GIT Cancers
2.1.4. Correlation Pattern between IL4, IL4Ra, IL13, and IL13Ra1 and the Expression of Mediators Relevant for Cancer Growth in CRC Patients
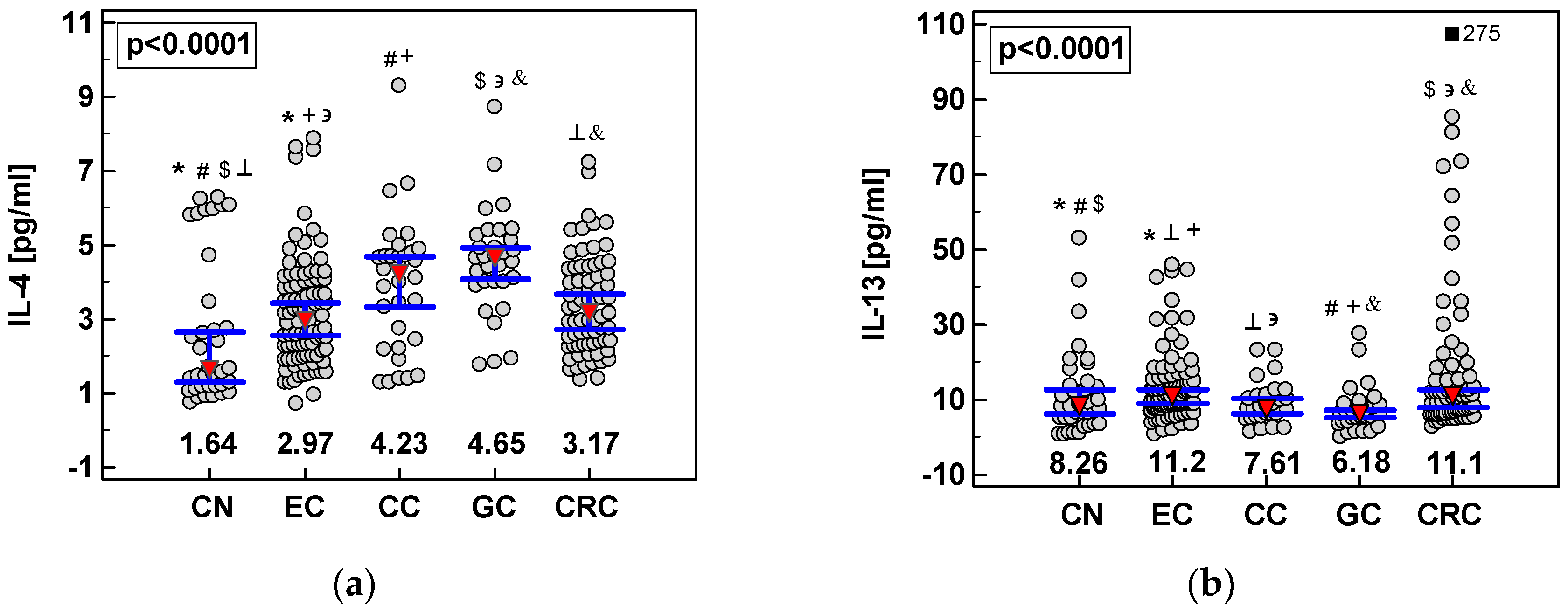
2.2. Systemic Concentrations of IL-4 and IL-13 in CRC and upper GIT Cancers
2.3. Effect of Exogenous IL-4 and IL-13 on Colon Cancer Cells—Preliminary Findings
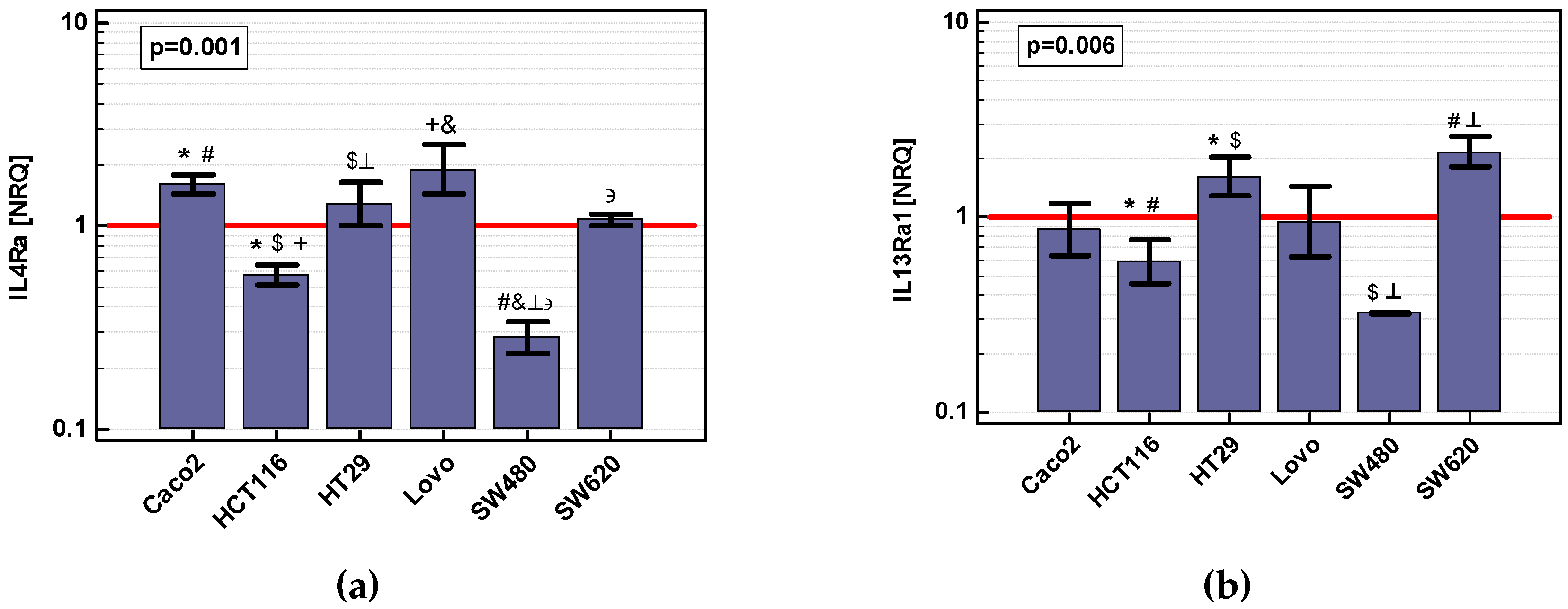
2.3.1. Expression of IL4 and IL13 and Their Receptors (IL4Ra and IL13Ra1) in Colon Cancer Cell Lines: Effect of Nutritional Stress, Chemically Induced Hypoxia, and IL-4 and IL-13
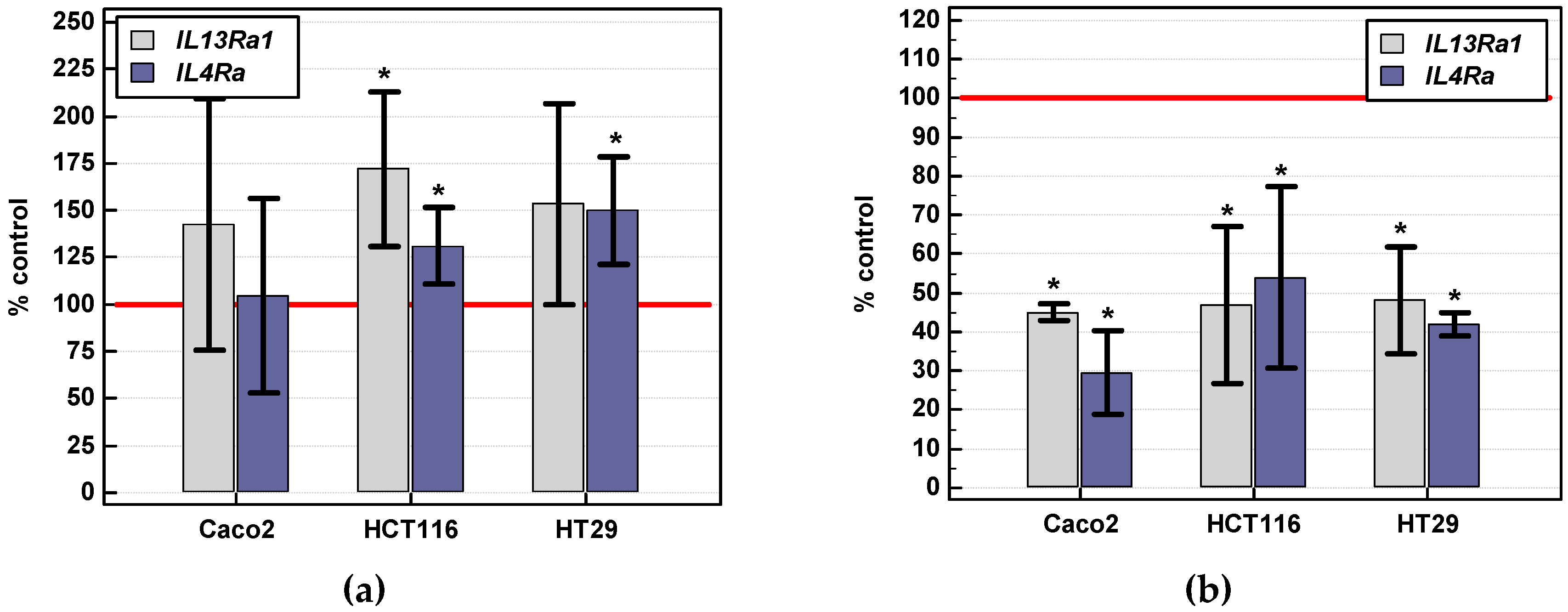
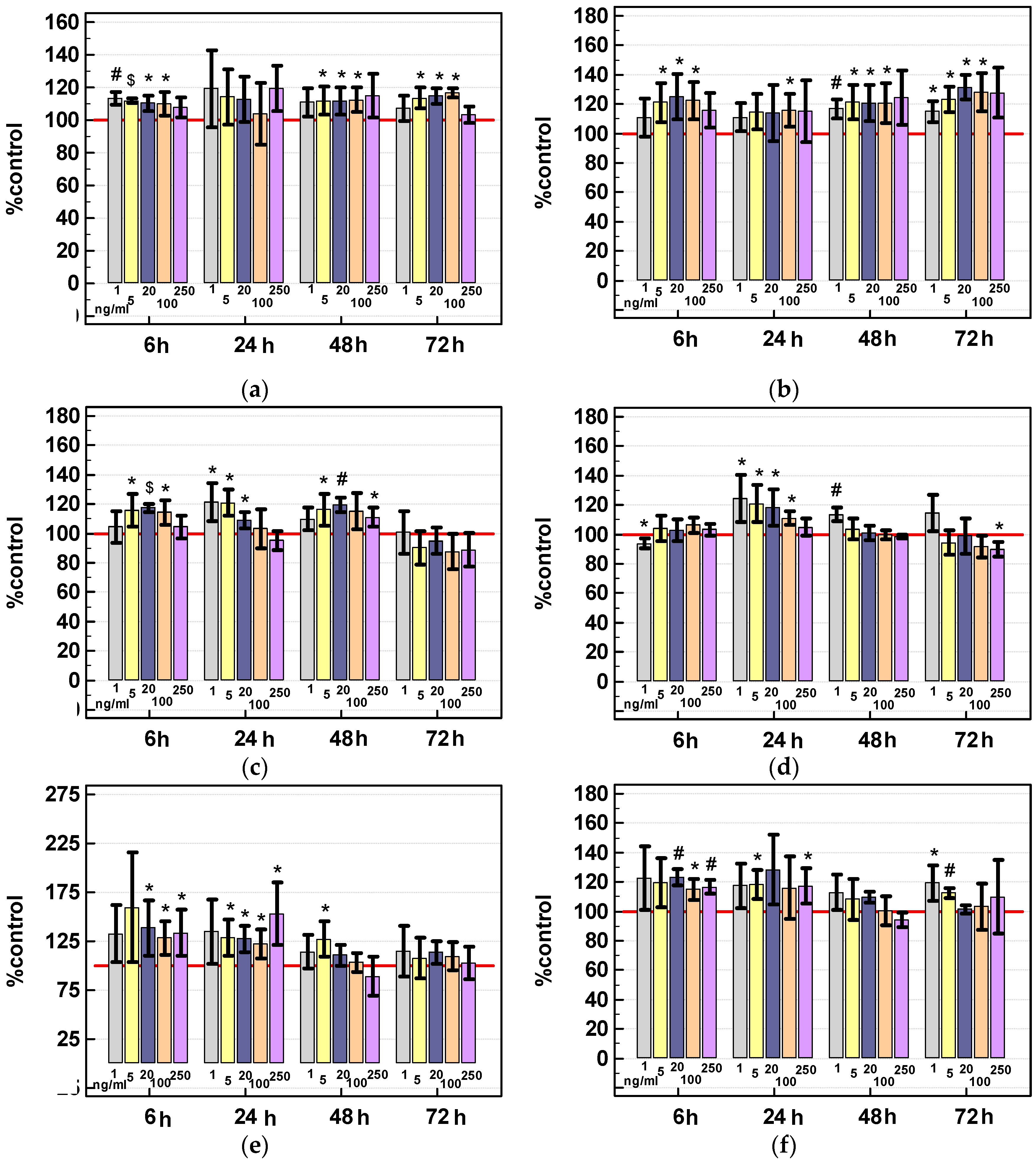
2.3.2. Effect of IL-4 and IL-13 on Viability of Colon Cancer Cells
Viability Assay
Gene Expression
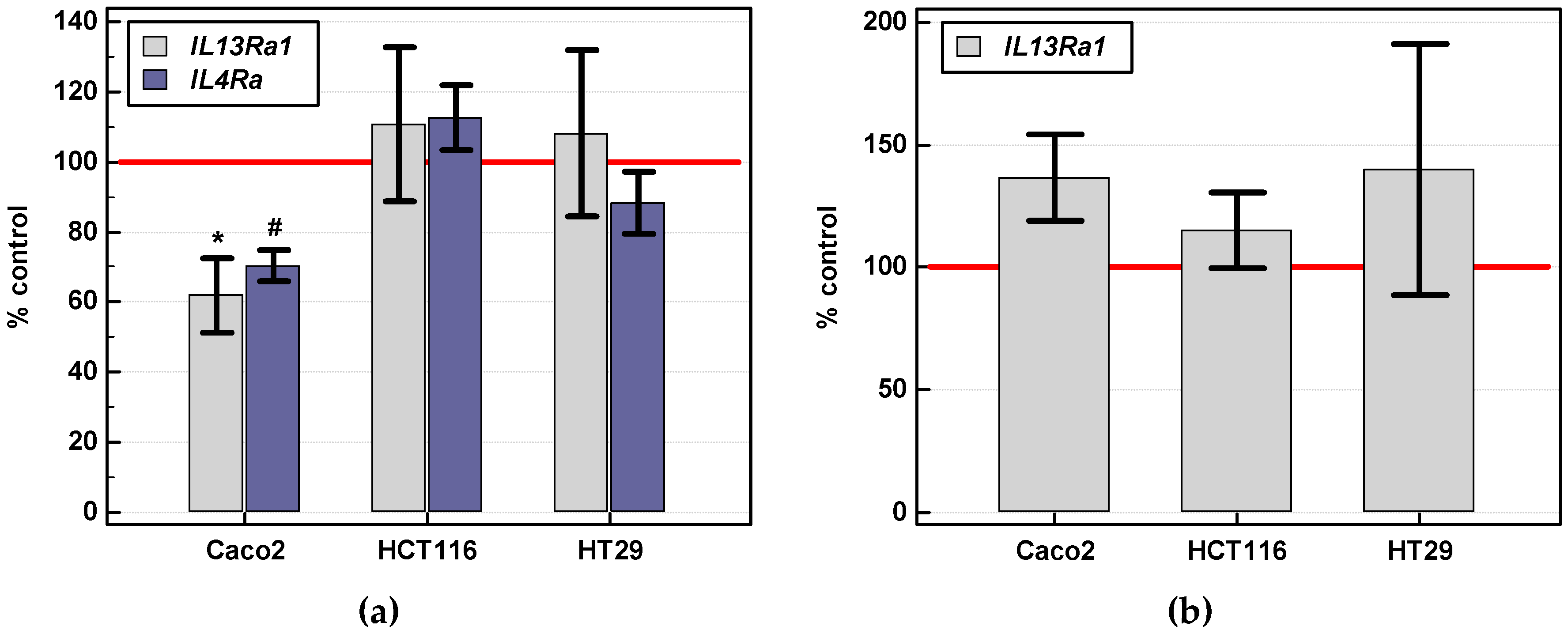
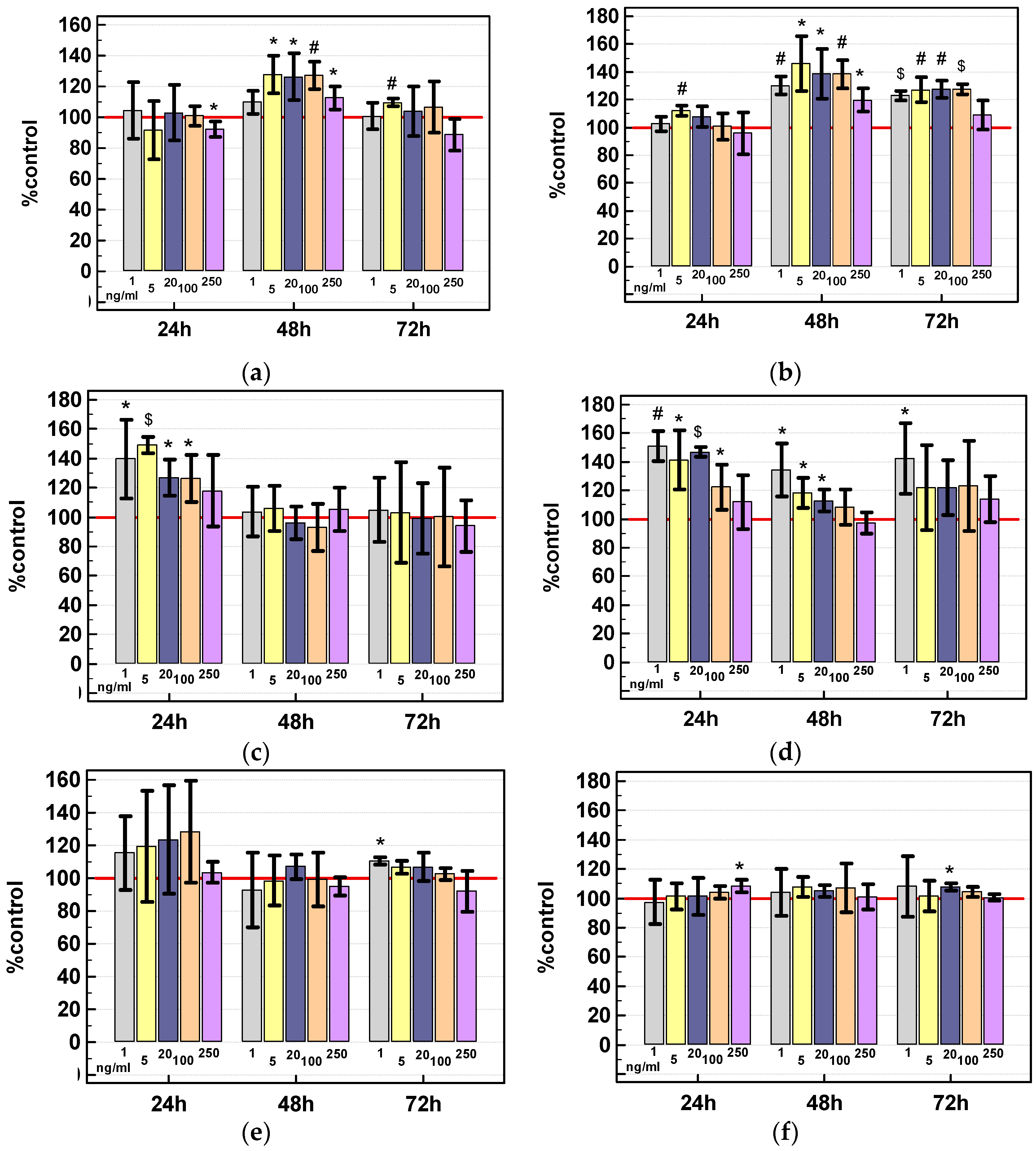
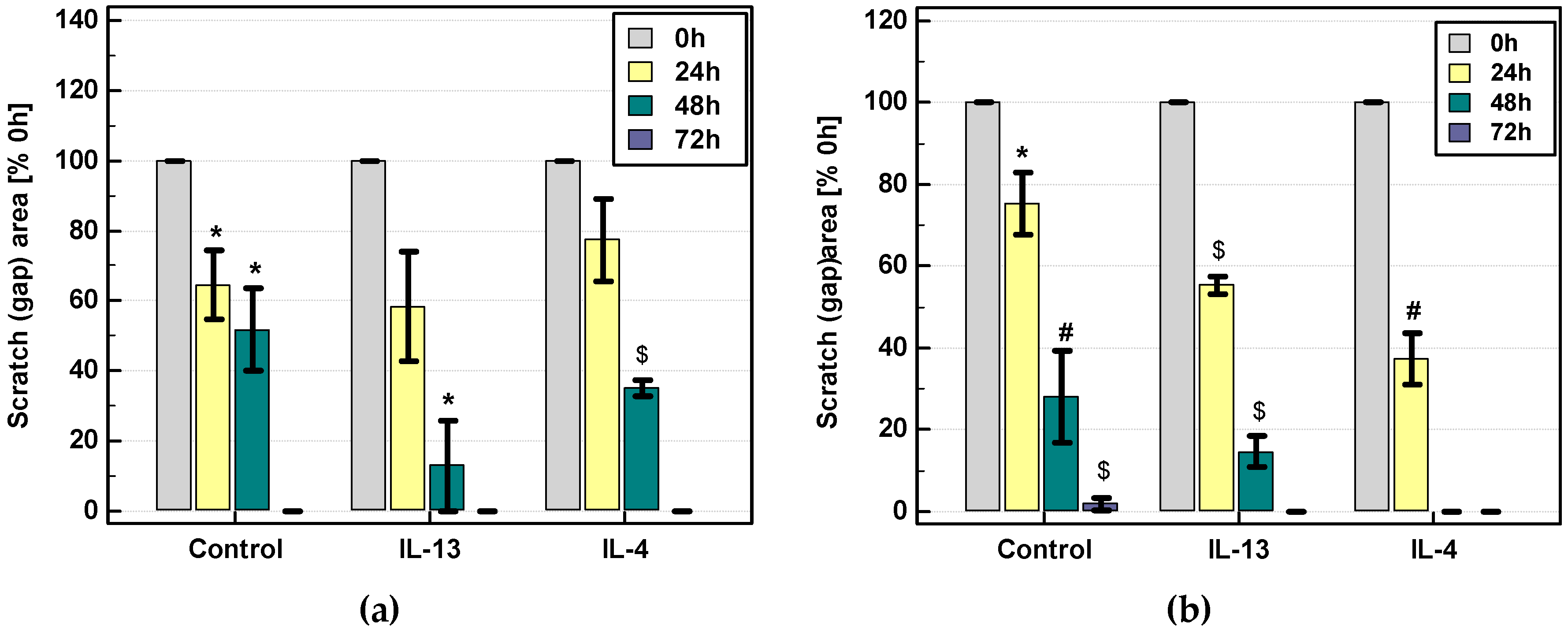
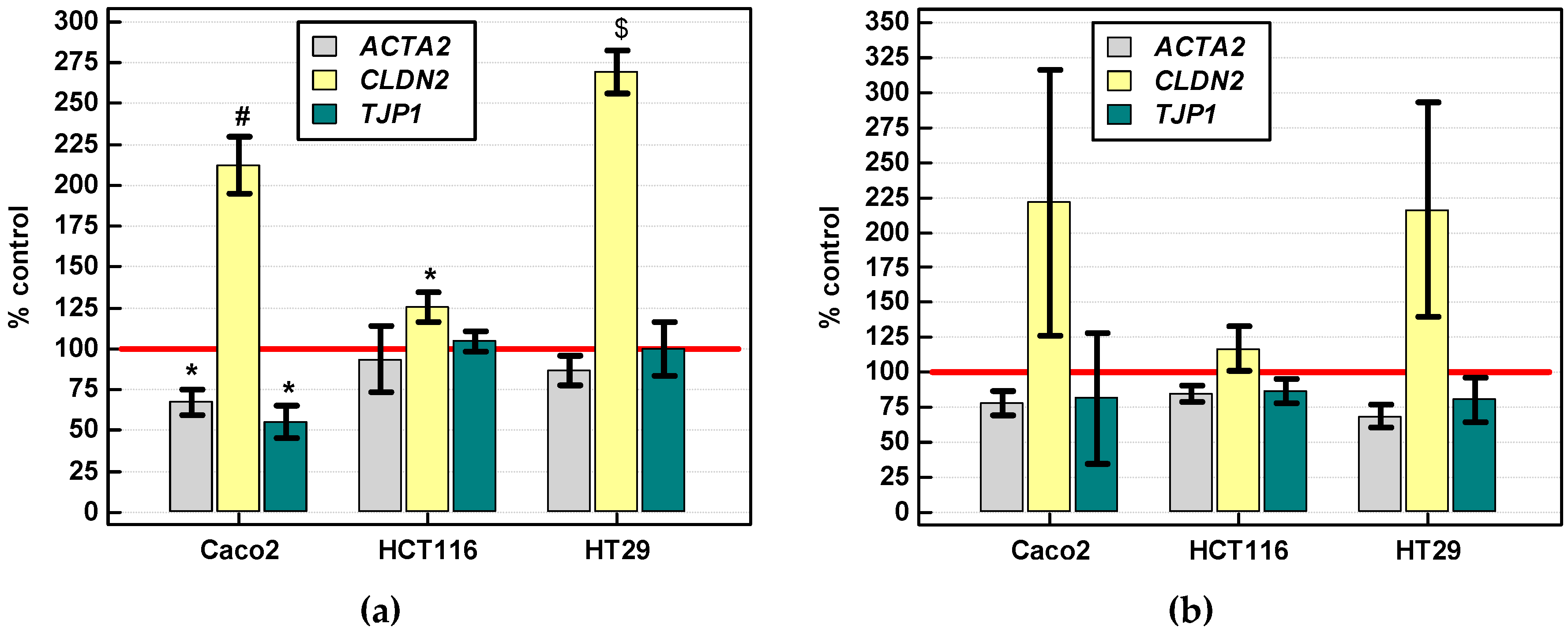
2.3.3. Effect of IL-4 and IL-13 on Motility of Colon Cancer Cells and Expression of Motility-Related Genes
Scratch Assay
Gene Expression
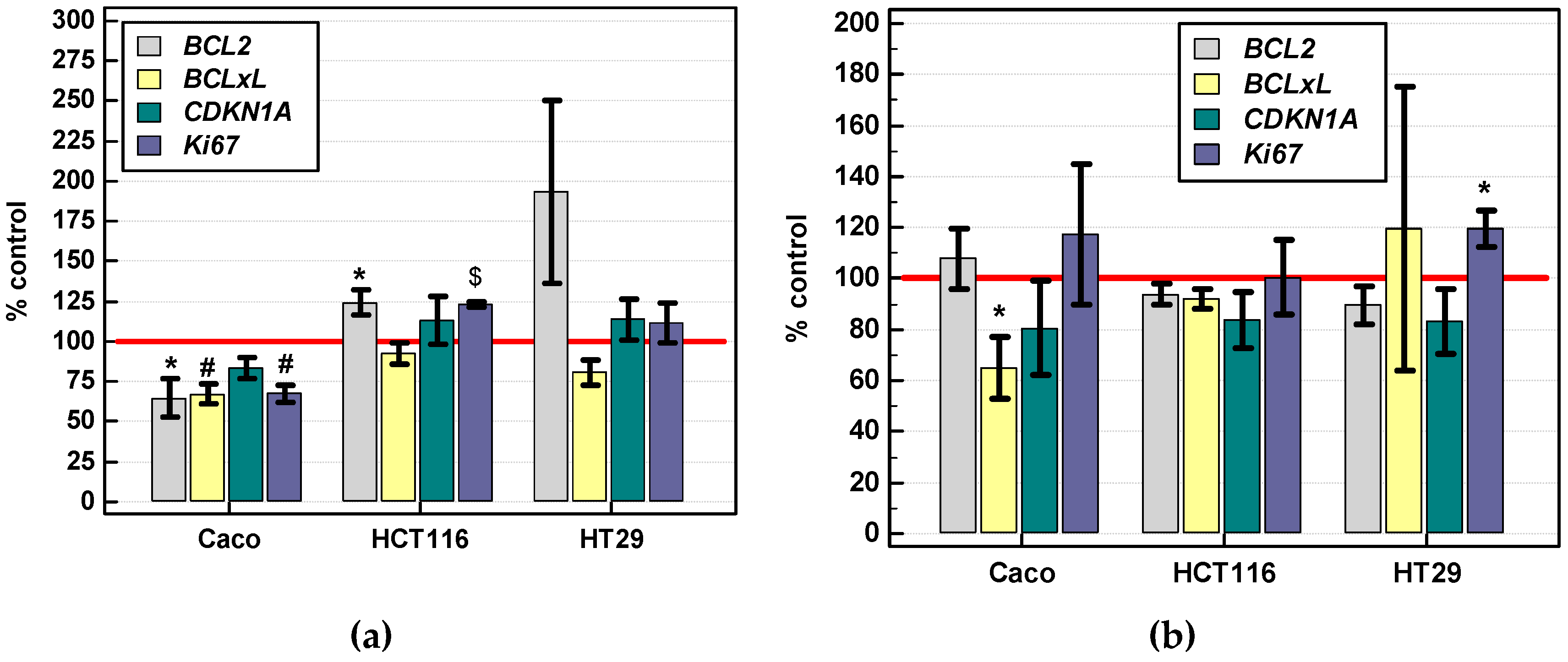
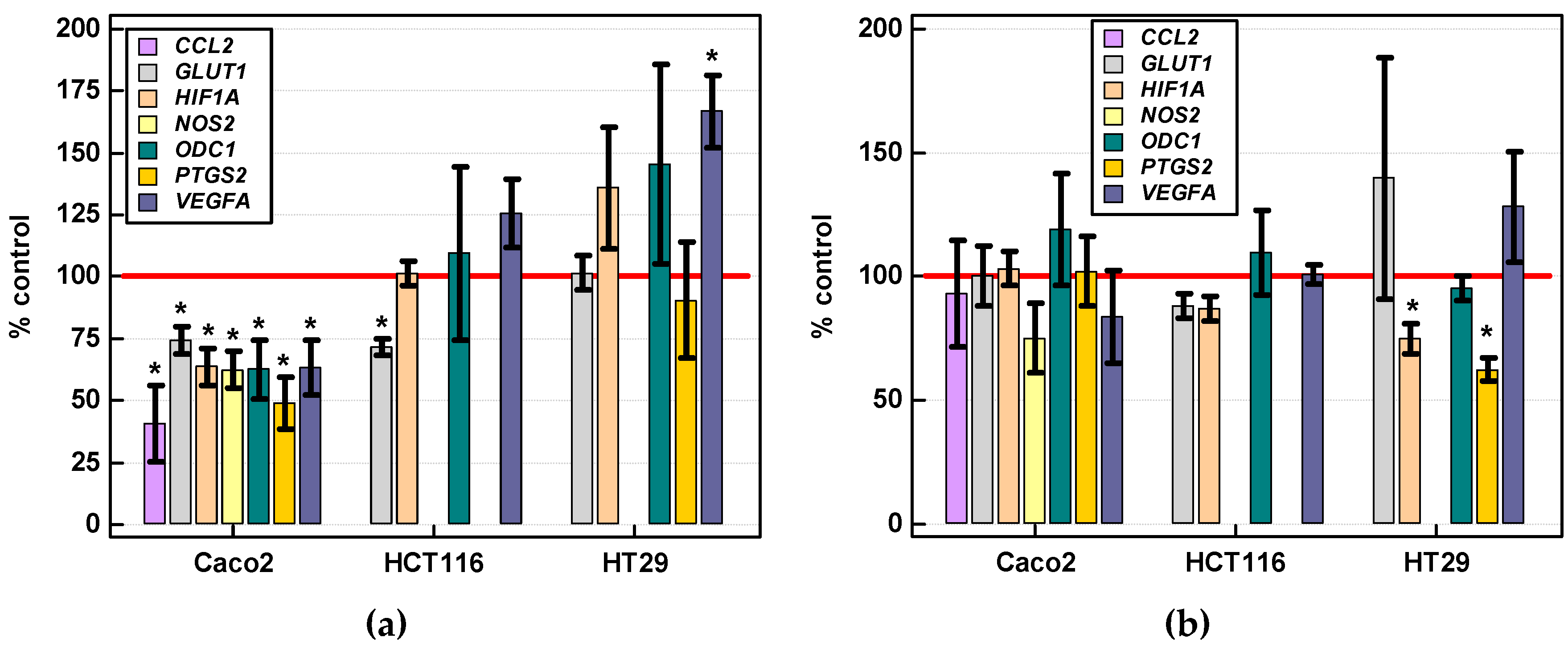
2.3.4. Effect of IL-4 and IL-13 on Expression of Various Cancer Development-Related Genes
3. Discussion
4. Materials and Methods
4.1. Patients
4.1.1. Study Population for Analysis of Local Interleukin Expression
4.1.2. Study Population for Determination of Systemic Interleukin Concentration
4.2. Ethical Considerations
4.3. Analytical Methods
4.3.1. Local Interleukin Concentration (Protein)
4.3.2. Systemic Interleukin Concentration
4.3.3. Transcriptomic Analysis
4.3.4. Cell Cultures and Cell Culture Experiments
Sulforhodamine B (SRB) Viability Assay
Scratch Assay
Cell Culturing for Transcriptional Analysis
4.3.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CC | Adenocarcinoma of gastric cardia |
| CRC | Colorectal adenocarcinoma |
| ESCC FBS | Esophageal squamous cell carcinoma Fetal bovine serum |
| GC | Gastric adenocarcinoma |
| GIT | Gastrointestinal tract |
| IL | Interleukin |
| EMT | Epithelial-mesenchymal transition |
| NRQSRB | Normalized relative quantitiesSulforhodamine B |
| TNM | Tumor-node-metastasis grading system |
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grierson, P.; Lim, K.-H.; Amin, M. Immunotherapy in gastrointestinal cancers. J. Gastrointest. Oncol. 2017, 8, 474–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bever, K.M.; Le, D.T. An expanding role for immunotherapy in colorectal cancer. J. Natl. Compr. Canc. Netw. 2017, 15, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Rabinovich, G.A.; Gabrilovich, D.; Sotomayor, E.M. Immunosuppressive strategies that are mediated by tumor cells. Annu. Rev. Immunol. 2007, 25, 267–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteran, L.; Erez, N. The dark side of fibroblasts: Cancer-associated fibroblasts as mediators of immunosuppression in the tumor microenvironment. Front. Immunol. 2019, 10, 1835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallett, M.A.; Venmar, K.T.; Fingleton, B. Cytokine stimulation of epithelial cancer cells: The similar and divergent functions of IL4 and IL13. Cancer Res. 2012, 72, 6338–6343. [Google Scholar] [CrossRef] [Green Version]
- Bankaitis, K.V.; Fingleton, B. Targeting IL4/IL4R for the treatment of epithelial cancer metastasis. Clin. Exp. Metastasis 2015, 32, 847–856. [Google Scholar] [CrossRef]
- Zhou, R.; Qian, S.; Gu, X.; Chen, Z.; Xiang, J. Interleukin-13 and its receptors in colorectal cancer (Review). Biomed. Rep. 2013, 1, 687–690. [Google Scholar] [CrossRef] [Green Version]
- Barderas, S.; Bartolomé, R.A.; Fernandez-Aceñero, M.J.; Torres, S.; Casal, J.I. High expression of IL-13 receptor α2 in colorectal cancer is associated with invasion, liver metastasis, and poor prognosis. Cancer Res. 2012, 72, 2780–2790. [Google Scholar] [CrossRef] [Green Version]
- Bartolomé, R.A.; Jaén, M.; Casal, J.I. An IL13Rα2 peptide exhibits therapeutic activity against metastatic colorectal cancer. Br. J. Cancer. 2018, 119, 940–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todaro, M.; Lombardo, Y.; Francipane, M.G.; Alea, M.P.; Cammareri, P.; Iovino, F.; Di Stefano, A.B.; Di Bernardo, C.; Agrusa, A.; Condorelli, G.; et al. Apoptosis resistance in epithelial tumors is mediated by tumor-cell-derived interleukin-4. Cell Death Differ. 2008, 15, 762–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, M.; Rabkin, C.S.; Torres, J.; Kemp, T.J.; Zabaleta, J.; Pinto, L.A.; Hildesheim, A.; Sánchez-Figueroa, L.; Guarner, J.; Herrera-Goepfert, R.; et al. Circulating inflammation-related markers and advanced gastric premalignant lesions. J. Gastroenterol. Hepatol. 2019, 34, 852–856. [Google Scholar] [CrossRef] [PubMed]
- Vinagre, R.M.D.F.; Vinagre, I.D.F.; Vilar-E.-Silva, A.; Fecury, A.A.; Martins, L.C. Helicobacter pylori infection and immune profile of patients with different gastroduodenal diseases. Arq. Gastroenterol. 2018, 55, 122–127. [Google Scholar] [CrossRef]
- Diaz Orea, M.A.; Muñoz Perez, V.; Gómez Conde, E.; Castellanos Sánchez, V.O.; Gonzalez Lopez, R.; Flores Alonso, J.C.; Cárdenas, M.E.; Galicia, A.L.; Mendoza, A. Expression of cytokines interleukin-2, interleukin-4, interleukin-10 and transforming growth factor β in gastric adenocarcinoma biopsies obtained from mexican patients. Asian Pac. J. Cancer Prev. 2017, 18, 577–582. [Google Scholar]
- Lin, C.; Liu, H.; Zhang, H.; He, H.; Li, H.; Shen, Z.; Qin, J.; Qin, X.; Xu, J.; Sun, Y. Interleukin-13 receptor α2 is associated with poor prognosis in patients with gastric cancer after gastrectomy. Oncotarget 2016, 7, 49281–49288. [Google Scholar] [CrossRef]
- Liu, H.; Antony, S.; Roy, K.; Juhasz, A.; Wu, Y.; Lu, J.; Meitzler, J.L.; Jiang, G.; Polley, E.; Doroshow, J.H. Interleukin-4 and interleukin-13 increase NADPH oxidase 1-related proliferation of human colon cancer cells. Oncotarget 2017, 8, 38113–38135. [Google Scholar] [CrossRef] [Green Version]
- Matsui, S.; Okabayashi, K.; Tsuruta, M.; Shigeta, K.; Seishima, R.; Ishida, T.; Kondo, T.; Suzuki, Y.; Hasegawa, H.; Shimoda, M.; et al. Interleukin-13 and its signaling pathway is associated with obesity-related colorectal tumorigenesis. Cancer Sci. 2019, 110, 2156–2165. [Google Scholar] [CrossRef]
- Miska, J.; Lui, J.B.; Toomer, K.H.; Devarajan, P.; Cai, X.; Houghton, J.; Lopez, D.M.; Abreu, M.T.; Wang, G.; Chen, Z. Initiation of inflammatory tumorigenesis by CTLA4 insufficiency due to type 2 cytokines. J. Exp. Med. 2018, 215, 841–858. [Google Scholar] [CrossRef] [Green Version]
- Koller, F.L.; Hwang, D.G.; Dozier, E.A.; Fingleton, B. Epithelial interleukin-4 receptor expression promotes colon tumor growth. Carcinogenesis 2010, 31, 1010–1017. [Google Scholar] [CrossRef]
- Li, J.; Chen, X.L.; Shaker, A.; Oshima, T.; Shan, J.; Miwa, H.; Feng, C.; Zhang, J. Contribution of immunomodulators to gastroesophageal reflux disease and its complications: Stromal cells, interleukin 4, and adiponectin. Ann. NY Acad. Sci. 2016, 1380, 183–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, A.R.; Goldenring, J.R. Injury, repair, inflammation and metaplasia in the stomach. J. Physiol. 2018, 596, 3861–3867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Zhang, B.Z.; Qin, Y.R.; Bi, J.; Liu, H.B.; Li, Y.; Cai, M.Y.; Ma, S.; Chan, K.W.; Xie, D.; et al. CD68 and interleukin 13, prospective immune markers for esophageal squamous cell carcinoma prognosis prediction. Oncotarget 2016, 7, 15525–15538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saigusa, S.; Tanaka, K.; Inoue, Y.; Toiyama, Y.; Okugawa, Y.; Iwata, T.; Mohri, Y.; Kusunoki, M. Low serum interleukin-13 levels correlate with poorer prognoses for colorectal cancer patients. Int. Surg. 2014, 99, 223–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Essner, R.; Huynh, Y.; Nguyen, T.; Rose, M.; Kojima, M.; Hoon, D.S. Functional interleukin-4 receptor and interleukin-2 receptor common gamma chain in human gastric carcinoma: A possible mechanism for cytokine-based therapy. J. Gastrointest. Surg. 2001, 5, 81–90. [Google Scholar] [CrossRef]
- Formentini, A.; Braun, P.; Fricke, H.; Link, K.H.; Henne-Bruns, D.; Kornmann, M. Expression of interleukin-4 and interleukin-13 and their receptors in colorectal cancer. Int. J. Colorectal Dis. 2012, 27, 1369–1376. [Google Scholar] [CrossRef] [PubMed]
- Ko, C.W.; Cuthbert, R.J.; Orsi, N.M.; Brooke, D.A.; Perry, S.L.; Markham, A.F.; Coletta, P.L.; Hull, M.A. Lack of interleukin-4 receptor alpha chain-dependent signalling promotes azoxymethane-induced colorectal aberrant crypt focus formation in Balb/c mice. J. Pathol. 2008, 214, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.L.; Peng, X.; Fu, X.Y. Interleukin-4 mediates cell growth inhibition through activation of Stat1. J. Biol. Chem. 2000, 275, 10212–10217. [Google Scholar] [CrossRef] [Green Version]
- Toi, M.; Bicknell, R.; Harris, A.L. Inhibition of colon and breast carcinoma cell growth by interleukin-4. Cancer Res. 1992, 52, 275–279. [Google Scholar]
- Morisaki, T.; Uchiyama, A.; Yuzuki, D.; Essner, R.; Morton, D.L.; Hoon, D.S. Interleukin 4 regulates G1 cell cycle progression in gastric carcinoma cells. Cancer Res. 1994, 54, 1113–1118. [Google Scholar]
- So, E.-Y.; Park, H.-H.; Lee, C.-E. IFN-γ and IFN-α posttranscriptionally down-regulate the IL-4-induced IL-4 receptor gene expression. J. Immunol. 2000, 165, 5472–5479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.T.; Hu, W.T.; Zhou, J.Y.; Tu, Y. Celecoxib enhances the radiosensitivity of HCT116 cells in a COX-2 independent manner by up-regulating BCCIP. Am. J. Transl. Res. 2017, 9, 1088–1100. [Google Scholar] [PubMed]
- Venmar, K.T.; Carter, K.J.; Hwang, D.G.; Dozier, E.A.; Fingleton, B. IL4 receptor ILR4α regulates metastatic colonization by mammary tumors through multiple signaling pathways. Cancer Res. 2014, 74, 4329–4340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, J.; Minoda, Y.; Meredith, T.; Cameron, G.; Philipp, M.S.; Pellicci, D.G.; Corbett, A.J.; Kurts, C.; Gray, D.H.; Godfrey, D.I.; et al. Chronically stimulated human MAIT cells are unexpectedly potent IL-13 producers. Immuno. Cell Biol. 2019, 97, 689–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dougan, M.; Dranoff, G.; Dougan, S.K. Cancer immunotherapy: Beyond checkpoint blockade. Annu. Rev. Cancer Biol. 2019, 3, 4.1–4.21. [Google Scholar] [CrossRef]
- von Bernstorff, W.; Voss, M.; Freichel, S.; Schmid, A.; Vogel, I.; Jöhnk, C.; Henne-Bruns, D.; Kremer, B.; Kalthoff, H. Systemic and local immunosuppression in pancreatic cancer patients. Clin. Cancer Res. 2001, 7 (Suppl. 3), 925s–932s. [Google Scholar]
- Cárdenas, D.M.; Sánchez, A.C.; Rosas, D.A.; Rivero, E.; Paparoni, M.D.; Cruz, M.A.; Suárez, Y.P.; Galvis, N.F. Preliminary analysis of single-nucleotide polymorphisms in IL-10, IL-4, and IL-4Rα genes and profile of circulating cytokines in patients with gastric Cancer. BMC Gastroenterol. 2018, 18, 184. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.W.; Kim, S.K.; Kim, C.S.; Kim, I.Y.; Cho, M.Y.; Kim, N.K. Association of serum and intratumoral cytokine profiles with tumor stage and neutrophil lymphocyte ratio in colorectal cancer. Anticancer Res. 2014, 34, 3481–3487. [Google Scholar]
- Krzystek-Korpacka, M.; Zawadzki, M.; Kapturkiewicz, B.; Lewandowska, P.; Bednarz-Misa, I.; Gorska, S.; Witkiewicz, W.; Gamian, A. Subsite heterogeneity in the profiles of circulating cytokines in colorectal cancer. Cytokine 2018, 110, 435–441. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Okamura, S.; Yamaji, T.; Iwasaki, M.; Tsugane, S.; Shetty, V.; Koizumi, T. Plasma cytokine levels and the presence of colorectal cancer. PLoS ONE 2019, 14, e0213602. [Google Scholar] [CrossRef] [Green Version]
- Johdi, N.A.; Mazlan, L.; Sagap, I.; Jamal, R. Profiling of cytokines, chemokines and other soluble proteins as a potential biomarker in colorectal cancer and polyps. Cytokine 2017, 99, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Olsen, R.S.; Nijm, J.; Andersson, R.E.; Dimberg, J.; Wågsäter, D. Circulating inflammatory factors associated with worse long-term prognosis in colorectal cancer. World. J. Gastroenterol. 2017, 23, 6212–6219. [Google Scholar] [CrossRef] [PubMed]
- Wong, R. Proximal tumors are associated with greater mortality in colon cancer. J. Gen. Intern. Med. 2010, 25, 1157–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.W.; Tsai, H.L.; Huang, M.Y.; Huang, C.M.; Yeh, Y.S.; Ma, C.J.; Wang, J.Y. Different clinicopathologic features and favorable outcomes of patients with stage III left-sided colon cancer. World J. Surg. Oncol. 2015, 13, 257. [Google Scholar]
- Moritani, K.; Hasegawa, H.; Okabayashi, K.; Ishii, Y.; Endo, T.; Kitagawa, Y. Difference in the recurrence rate between right- and left-sided colon cancer: A 17-year experience at a single institution. Surg. Today 2014, 44, 1685–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neubauer, K.; Matusiewicz, M.; Bednarz-Misa, I.; Gorska, S.; Gamian, A.; Krzystek-Korpacka, M. Diagnostic potential of systemic eosinophil-associated cytokines and growth factors in IBD. Gastroenterol. Res. Pract. 2018, 2018, 7265812. [Google Scholar] [CrossRef] [Green Version]
- Gabitass, R.F.; Annels, N.E.; Stocken, D.D.; Pandha, H.A.; Middleton, G.W. Elevated myeloid-derived suppressor cells in pancreatic, esophageal and gastric cancer are an independent prognostic factor and are associated with significant elevation of the Th2 cytokine interleukin-13. Cancer Immunol. Immunother. 2011, 60, 1419–1430. [Google Scholar] [CrossRef] [Green Version]
- Xin, Z.; Wenyu, F.; Shenhua, X. Clinicopathologic significance of cytokine levels in esophageal squamous cell carcinoma. Hepatogastroenterology 2010, 57, 1416–1422. [Google Scholar]
- Zou, Z.; Zhao, L.; Su, S.; Liu, Q.; Yu, L.; Wei, J.; Yang, Y.; Du, J.; Shen, J.; Qian, X.; et al. The plasma levels of 12 cytokines and growth factors in patients with gastric cancer. Medicine (Baltimore) 2018, 97, e0413. [Google Scholar] [CrossRef]
- Zhang, J.; Patel, L.; Pienta, K.J. CC chemokine ligand 2 (CCL2) promotes prostate cancer tumorigenesis and metastasis. Cytokine Growth Factor Rev. 2010, 21, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; Yang, S.C.; Sharma, S.; Heuze-Vourch, N.; Dubinett, S.M. IL-4 regulates COX-2 and PGE2 production in human non-small cell lung cancer. Biochem. Biophys. Res. Commun. 2006, 343, 995–1001. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Gong, C.; Mao, H.; Li, Z.; Fang, Z.; Chen, Q.; Lin, M.; Jiang, X.; Hu, Y.; Wang, W.; et al. E2F1/SP3/STAT6 axis is required for IL-4-induced epithelial-mesenchymal transition of colorectal cancer cells. Int. J. Oncol. 2018, 53, 567–578. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.; Clark, D. Molecular margin of surgical resections—where do we go from here? Cancer 2015, 121, 1914–1916. [Google Scholar] [CrossRef] [PubMed]
- Dakubo, G.D.; Jakupciak, J.P.; Birch-Machin, M.A.; Parr, R.L. Clinical implications and utility of field cancerization. Cancer Cell. Int. 2007, 7, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, A.; Tripathi, G.; Gopalakrishnan, K.; Williams, N.; Arasaradnam, R.P. Field cancerisation in colorectal cancer: A new frontier or pastures past? World J. Gastroenterol. 2015, 21, 3763–3772. [Google Scholar] [CrossRef]
- Neubauer, K.; Bednarz-Misa, I.; Diakowska, D.; Kapturkiewicz, B.; Gamian, A.; Krzystek-Korpacka, M. Nampt/PBEF/visfatin upregulation in colorectal tumors, mirrored in normal tissue and whole blood of colorectal cancer patients, is associated with metastasis, hypoxia, IL1β, and anemia. Biomed. Res. Int. 2015, 2015, 523930. [Google Scholar] [CrossRef] [Green Version]
- Krzystek-Korpacka, M.; Gorska, S.; Diakowska, D.; Kapturkiewicz, B.; Podkowik, M.; Gamian, A.; Bednarz-Misa, I. Midkine is up-regulated in both cancerous and inflamed bowel, reflecting lymph node metastasis in colorectal cancer and clinical activity of ulcerative colitis. Cytokine 2017, 89, 68–75. [Google Scholar] [CrossRef]
- Bednarz-Misa, I.; Diakowska, D.; Krzystek-Korpacka, M. Local and systemic IL-7 concentration in gastrointestinal-tract cancers. Medicina 2019, 55, 262. [Google Scholar] [CrossRef] [Green Version]
- Schlesinger-Raab, A.; Werner, J.; Friess, H.; Hölzel, D.; Engel, J. Age and outcome in gastrointestinal cancers: A population-based evaluation of oesophageal, gastric and colorectal cancer. Visc. Med. 2017, 33, 245–253. [Google Scholar] [CrossRef]
- Greenbaum, D.; Colangelo, C.; Williams, K.; Gerstein, M. Comparing protein abundance and mRNA expression levels on a genomic scale. Genome Biol. 2003, 4, 117. [Google Scholar] [CrossRef] [Green Version]
- Gooch, J.L.; Christy, B.; Yee, D. STAT6 mediates interleukin-4 growth inhibition in human breast cancer cells. Neoplasia 2002, 4, 324–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Topp, M.S.; Koenigsmann, M.; Mire-Sluis, A.; Oberberg, D.; Eitelbach, F.; von Marschall, Z.; Notter, M.; Reufi, B.; Stein, H.; Thiel, E.; et al. Recombinant human interleukin-4 inhibits growth of some human lung tumor cell lines in vitro and in vivo. Blood 1993, 82, 2837–2844. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.R.; Mahdi, R.M.; Ebong, S.; Vistica, B.P.; Chen, J.; Guo, Y.; Gery, I.; Egwuagu, C.E. Cell proliferation and stat6 pathways are negatively regulated in t cells by stat1 and suppressors of cytokine signaling. J. Immunol. 2004, 173, 737–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vella, V.; Mineo, R.; Frasca, F.; Mazzon, E.; Pandini, G.; Vigneri, R.; Belfiore, A. Interleukin-4 stimulates papillary thyroid cancer cell survival: Implications in patients with thyroid cancer and concomitant Graves’ disease. J. Clin. Endocrinol. Metab. 2004, 89, 2880–2889. [Google Scholar] [CrossRef] [Green Version]
- Salvador, E.; Burek, M.; Förster, C.Y. Tight junctions and the tumor microenvironment. Curr. Pathobiol. Rep. 2016, 4, 135–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saatian, B.; Rezaee, F.; Desando, S.; Emo, J.; Chapman, T.; Knowlden, S.; Georas, S.N. Interleukin-4 and interleukin-13 cause barrier dysfunction in human airway epithelial cells. Tissue Barriers 2013, 1, e24333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Sadi, R.; Boivin, M.; Ma, T. Mechanism of cytokine modulation of epithelial tight junction barrier. Front. Biosci. 2009, 14, 2765–2778. [Google Scholar] [CrossRef] [Green Version]
- Hellemans, J.; Vandesompele, J. qPCR data analysis—Unlocking the secret to successful results. In PCR Troubleshooting and Optimization: The Essential Guide, 1st ed.; Kennedy, S., Oswald, N., Eds.; Caister Academic Press: Poole, UK, 2011; pp. 1–13. [Google Scholar]
- Tam, S.Y.; Wu, V.W.C.; Law, H.K.W. JNK pathway mediates low oxygen level induced epithelial–mesenchymal transition and stemness maintenance in colorectal cancer cells. Cancers 2020, 12, 224. [Google Scholar] [CrossRef] [Green Version]





















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | ESCC | GC | CRC | p |
|---|---|---|---|---|
| n | 16 | 14 | 21 | - |
| Sex (F/M), n | 6/10 | 3/11 | 7/14 | 0.619 1 |
| Age [yrs.], mean ± SD | 63.3 ± 7 | 67.3 ± 10 | 67.7 ± 11.4 | 0.431 2 |
| Stage (I/II/III/IV) | 0/5/10/1 | 3/3/5/3 | 5/9/7/0 | 0.064 1 |
| Primary tumor, T (1/2/3/4) | 0/5/8/3 | 2/2/8/2 | 1/5/15/0 | 0.241 1 |
| Lymph node metastasis, N (no/yes) | 8/8 | 6/8 | 14/7 | 0.261 1 |
| Distant metastasis, M (no/yes) | 15/1 | 11/3 | 21/0 | 0.067 1 |
| Characteristics | ESCC | GC | CRC | p |
|---|---|---|---|---|
| n | 18 | 14 | 17 | - |
| Sex (F/M), n | 8/10 | 3/11 | 7/10 | 0.365 1 |
| Age [yrs.], mean ± SD | 64.1 ± 5.6 | 67.3 ± 10 | 66.8 ± 11.3 | 0.567 2 |
| Stage (I/II/III/IV) | 2/6/9/1 | 3/3/5/3 | 4/8/5/0 | 0.248 1 |
| Primary tumor, T (1/2/3/4) | 1/6/9/2 | 2/1/9/2 | 1/5/11/0 | 0.424 1 |
| Lymph node metastasis, N (no/yes) | 9/9 | 6/8 | 12/5 | 0.261 1 |
| Distant metastasis, M (no/yes) | 17/1 | 11/3 | 17/0 | 0.084 1 |
| Data | Fold-Change [Tumor-to-Adjacent] in IL-4 Protein Concentration | ||
|---|---|---|---|
| CRC | ESCC | GC | |
| Age | r =0, p = 0.897 1 | r = 0, p = 0.930 1 | r = 0.46, p = 0.099 1 |
| Sex F vs. M | 1.6 vs. 4.2, p = 0.199 2 | 7.9 vs. 9.2, p = 0.844 2 | 10.1 vs. 3.1, p = 0.392 3 |
| TNM | ρ = 0.11, p = 0.679 4 | ρ = 0.15, p = 0.553 4 | ρ = 0.53, p = 0.051 4 |
| T | ρ = 0.20, p = 0.452 4 | ρ = 0.08, p = 0.762 4 | ρ = 0.46, p = 0.100 4 |
| N0 vs. N ≥ 1 | 3.5 vs. 2.2, p = 0.568 2 | 6.6 vs. 10.7, p = 0.528 2 | 3.7 vs. 19.6, p = 0.349 2 |
| M0 vs. M1 | - | - | 2.1 vs. 17.6, p = 0.024 3 |
| G | ρ = 0.36, p = 0.161 4 | ρ = 0.73, p < 0.001 4 | ρ = 0.32, p = 0.271 4 |
| Data | Fold-Change [Tumor-to-Adjacent] in IL4 Transcript Number | ||
|---|---|---|---|
| CRC | ESCC | GC | |
| Age | r = 16, p = 0.475 1 | r = 21, p = 0.444 1 | r = −0.22, p = 0.440 1 |
| Sex F vs. M | 1.7 vs. 0.6, p = 0.331 2 | 12.2 vs. 1.4, p = 0.206 2 | 0.3 vs. 1.2, p = 0.172 2 |
| TNM | ρ = −0.14, p = 0.548 3 | ρ= −0.21, p = 0.427 3 | ρ = 0.06, p = 0.834 3 |
| T | ρ = −0.37, p = 0.096 3 | ρ= −0.17, p = 0.529 3 | ρ = −0.06, p = 0.829 3 |
| N0 vs. N ≥ 1 | 0.7 vs. 1.3, p = 0.549 2 | 17.4 vs. 0.6, p = 0.030 2 | 0.6 vs. 1.2, p = 0.490 2 |
| M0 vs. M1 | - | - | 1.0 vs. 0.7, p = 0.747 2 |
| G | ρ = 0.01, p = 0.796 3 | ρ= 0.08, p = 0.759 3 | ρ = 0, p = 0.973 3 |
| Data | Fold-Change [Tumor-to-Adjacent] in IL4Ra Transcript Number | ||
|---|---|---|---|
| CRC | ESCC | GC | |
| Age | r = 22, p = 0.347 1 | r = 25, p = 0.355 1 | r = 0, p = 0.999 1 |
| Sex F vs. M | 1.1 vs. 0.7, p = 0.191 2 | 1 vs. 1.6, p = 0.255 2 | 4 vs. 2.9, p = 0.854 2 |
| TNM | ρ = −0.05, p = 0.815 3 | ρ = 0.06, p = 0.839 3 | ρ = −0.25, p = 0.383 3 |
| T | ρ = −0.07, p = 0.772 3 | ρ = 0.13, p = 0.627 3 | ρ = −0.25, p = 0.390 3 |
| N0 vs. N ≥ 1 | 0.9 vs. 0.8, p = 0.729 2 | 1.9 vs. 1, p = 0.074 2 | 3 vs. 3.2, p = 0.958 2 |
| M0 vs. M1 | - | - | 4.1 vs. 0.2, p = 0.492 2 |
| G | ρ = −0.04, p = 0.863 3 | ρ = 0.47, p = 0.077 3 | ρ = 0.05, p = 0.861 3 |
| Data | Fold-Change [Tumor-to-Adjacent] in IL-13 Protein Concentration | ||
|---|---|---|---|
| CRC | ESCC | GC | |
| Age | r = 23, p = 0.366 1 | r = −0.19, p = 0.449 1 | r = 0.34, p = 0.231 1 |
| Sex F vs. M | 1.4 vs. 1.5, p = 0.565 2 | 2 vs. 2.2, p = 0.740 2 | 2.2 vs. 1.4, p = 0.136 2 |
| TNM | ρ = 0.54, p = 0.024 3 | ρ = −0.02, p = 0.940 3 | ρ = −0.13, p = 0.658 3 |
| T | ρ = 0.30, p = 0.251 3 | ρ = 0.12, p = 0.631 3 | ρ = 0.14, p = 0.643 3 |
| N0 vs. N ≥ 1 | 1.3 vs. 1.9, p = 055 2 | 2 vs. 2, p = 0.965 2 | 2 vs. 1.4, p = 0.155 2 |
| M0 vs. M1 | - | - | 1.9 vs. 1.4, p = 0.506 2 |
| G | ρ = −0.07, p = 0.797 3 | ρ = −0.37, p = 0.137 3 | ρ = 0.15, p = 0.601 3 |
| Data | Fold-Change [Tumor-to-Adjacent] in IL13 Expression | ||
|---|---|---|---|
| CRC | ESCC | GC | |
| Age | r = 25, p = 0.266 1 | r = −0.24, p = 0.367 1 | r = 0.1, p = 0.736 1 |
| Sex F vs. M | 14.8 vs. 9, p = 0.736 2 | 1.2 vs. 5.4, p = 0.483 2 | 0.2 vs. 6.8, p = 0.145 2 |
| TNM | ρ = −0.56, p = 0.009 3 | ρ = -0.34, p = 0.192 3 | ρ = 0.13, p = 0.647 3 |
| T | ρ = −0.16, p = 0.490 3 | ρ = -0.24, p = 0.381 3 | ρ = −0.27, p = 0.344 3 |
| N0 vs. N ≥ 1 | 40.7 vs. 0.7, p = 002 2 | 4.9 vs. 1.9, p = 0.650 2 | 1.5 vs. 6, p = 0.485 2 |
| M0 vs. M1 | - | - | 2.2 vs. 13.5, p = 0.460 2 |
| G | ρ = −0.04, p = 0.863 3 | ρ = 0.01, p = 0.968 3 | ρ = −0.24, p = 0.417 3 |
| Data | Fold-Change [Tumor-to-Adjacent] in IL13Ra1 Expression | ||
|---|---|---|---|
| CRC | ESCC | GC | |
| Age | r = 19, p = 0.406 1 | r = −0.21, p = 0.436 1 | r = −0.23, p = 0.434 1 |
| Sex F vs. M | 0.8 vs. 1, p = 0.570 2 | 0.9 vs. 1.1, p = 0.658 2 | 1.1 vs. 1.3, p = 0.670 2 |
| TNM | ρ = 0.03, p = 0.910 3 | ρ = 0.05, p = 0.844 3 | ρ =−0.08, p = 0.774 3 |
| T | ρ = −0.08, p = 0.723 3 | ρ = 0.18, p = 0.517 3 | ρ = 0.14, p = 0.641 3 |
| N0 vs. N ≥ 1 | 0.9 vs. 1, p = 749 2 | 1.4 vs. 0.7, p = 0.144 2 | 1.1 vs. 1.4, p = 0.618 2 |
| M0 vs. M1 | - | - | 1.3 vs. 1, p = 0.547 2 |
| G | ρ = 0, p = 1 3 | ρ = 0.03, p = 0.903 3 | ρ = 0.20, p = 0.490 3 |
| Fold-Change in Expression | IL13 | IL13Ra1 | IL4 | IL4Ra |
|---|---|---|---|---|
| ACTA2 | ns | ns | 0.39 4 | 0.57 2 |
| BCLxL | ns | ns | ns | ns |
| BCL2 | ns | ns | ns | ns |
| CCL2 | ns | ns | ns | 0.67 3 |
| CDKN1A | ns | 0.60 2 | ns | 0.48 1 |
| CLDN2 | ns | ns | ns | −0.44 1 |
| GLUT1 | ns | ns | ns | ns |
| HIF1A | ns | 0.40 4 | ns | 0.42 4 |
| Ki67 | ns | ns | ns | −0.48 1 |
| NOS2 | ns | ns | ns | ns |
| ODC1 | ns | ns | ns | ns |
| PTGS2 | ns | ns | ns | 0.50 1 |
| TJP1 | ns | ns | ns | 0.68 3 |
| VEGFA | ns | 0.53 1 | ns | 0.48 1 |
| Expression | Tumor Tissue | Adjacent non-Cancerous Tissue | ||||||
|---|---|---|---|---|---|---|---|---|
| IL13 | IL13Ra1 | IL4 | IL4Ra | IL13 | IL13Ra1 | IL4 | IL4Ra | |
| ACTA2 | ns | 0.47 1 | ns | 0.64 2 | ns | ns | ns | 0.58 2 |
| BCLxL | ns | ns | ns | 0.40 4 | ns | ns | ns | ns |
| BCL2 | ns | 0.43 4 | ns | 0.57 2 | −0.39 4 | ns | ns | ns |
| CCL2 | ns | 0.51 2 | ns | 0.56 2 | ns | ns | 0.47 1 | ns |
| CDKN1A | ns | 0.47 1 | ns | 0.46 1 | ns | ns | ns | ns |
| CLDN2 | ns | ns | ns | −0.39 4 | ns | ns | ns | ns |
| GLUT1 | ns | ns | ns | ns | −0.41 4 | ns | ns | ns |
| HIF1A | ns | 0.68 3 | ns | 0.51 1 | ns | ns | 0.41 4 | ns |
| Ki67 | ns | ns | ns | ns | ns | 0.42 4 | ns | −0.46 1 |
| NOS2 | ns | ns | −0.42 4 | −0.42 4 | −0.41 4 | ns | ns | ns |
| ODC1 | ns | ns | ns | ns | ns | ns | ns | ns |
| PTGS2 | 0.42 4 | ns | ns | ns | 0.47 1 | ns | 0.41 4 | 0.64 2 |
| TJP1 | ns | 0.63 2 | ns | 0.65 2 | ns | 0.57 2 | ns | 0.44 4 |
| VEGFA | 0.39 4 | 0.63 2 | ns | 0.56 2 | ns | 0.40 4 | ns | ns |
| Characteristics | CN | ESCC | CC | GC | CRC | p |
|---|---|---|---|---|---|---|
| n | 36 | 93 | 32 | 31 | 71 | - |
| Sex (F/M), n | 16/20 | 32/61 | 7/25 | 10/21 | 27/44 | 0.375 1 |
| Age [yrs.], mean ± SD | 65.4 ± 8.1 | 62.4 ± 8.5 | 62.3 ± 8.5 | 64.1 ± 9.7 | 65.8 ± 10.3 | 0.110 2 |
| Stage (I/II/III/IV/x) | - | 9/21/25/38 | 1/2/4/26 | 1/5/5/20 | 7/28/29/6/1 | <0.001 1 |
| Primary tumor, T (1/2/3/4/x) | - | 11/17/24/41 | 0/1/3/28 | 1/2/9/19 | 1/8/39/22/1 | <0.001 1 |
| Lymph node metastasis, N (no/yes/x) | - | 34/59 | 2/30 | 6/25 | 35/35/1 | <0.001 1 |
| Distant metastasis, M (no/yes/x) | - | 55/38 | 6/26 | 11/20 | 64/6/1 | <0.001 1 |
| Symbol | Gene Name | Accession No. | Primer Sequence 5′→3′ | Amp. Size [bp] |
|---|---|---|---|---|
| IL4 1 | Interleukin 4 | NM_000589.4 | F: ccgtaacagacatctttgctgcc R: gagtgtccttctcatggtggct | 108 |
| IL13 1 | Interleukin 13 | NM_002188.3 | F: acggtcattgctctcacttgcc R: ctgtcaggttgatgctccatacc | 160 |
| IL4Ra 1 | Interleukin 4 receptor alpha | NM_000418.4 | F: ctgctcatggatgacgtggtca R: ggtgtgaactgtcaggtttcctg | 138 |
| IL13Ra1 1 | Interleukin 13 receptor subunit alpha 1 | NM_001560.3 | F: cctgaatgagaggatttgtctgc R: cagtcacagcagactcaggatc | 125 |
| ACTA2 1 | Alpha smooth muscle actin | NM_001141945.2 | F: ctatgcctctggacgcacaact R: cagatccagacgcatgatggca | 115 |
| BCL2 1 | B-cell lymphoma 2 | NM_000633.3 | F: atcgccctgtggatgactgagt R: gccaggagaaatcaaacagaggc | 127 |
| BCLxL 1 | B-cell lymphoma-extra large | NM_001317919.2 | F: gccacttacctgaatgaccacc R: aaccagcggttgaagcgttcct | 131 |
| CCL2 1 | Monocyte chemoattractant protein 1 (MCP1) | NM_002982.4 | F: agaatcaccagcagcaagtgtcc R: tcctgaacccacttctgcttgg | 98 |
| CDKN1A 1 | Cyclin Dependent Kinase Inhibitor 1A (p21CIP1/WAF1) | NM_001220777.2 | F: aggtggacctggagactctcag R: tcctcttggagaagatcagccg | 95 |
| GAPDH 2 | Glyceraldehyde-3-phosphate dehydrogenase | NM_001256799.3 | F: tagattattctctgatttggtcgtattgg R: gctcctggaagatggtgatgg | 223 |
| CLDN2 1 | Claudin 2 | NM_020384.4 | F: gtgacagcagttggcttctcca R: ggagattgcactggatgtcacc | 153 |
| GLUT1 1 | Glucose transporter 1 | NM_006516.4 | F: ttgcaggcttctccaactggac R: cagaaccaggagcacagtgaag | 113 |
| HIF1A 1 | Hypoxia-inducible factor 1α | NM_181054.3 | F: tatgagccagaagaacttttaggc R: cacctcttttggcaagcatcctg | 145 |
| Ki67 1 | Proliferation marker Ki67 | NM_001145966.2 | F: gaaagagtggcaacctgccttc R: gcaccaagttttactacatctgcc | 151 |
| NOS2 1 | Inducible nitric oxide synthase | NM_000625.4 | F: gctctacacctccaatgtgacc R: ctgccgagatttgagcctcatg | 136 |
| ODC1 1 | Ornithine decarboxylase | NM_001287189.2 | F: ccaaagcagtctgtcgtctcag R: cagagattgcctgcacgaaggt | 162 |
| PTGS2 1 | Prostaglandin-endoperoxide synthase 2 (COX2) | NM_000963.4 | F: cggtgaaactctggctagacag R: gcaaaccgtagatgctcaggga | 156 |
| TJP1 1 | Tight junction protein 1 | NM_001355014.2 | F: gtccagaatctcggaaaagtgcc R: ctttcagcgcaccataccaacc | 132 |
| VEGFA 1 | Vascular endothelial growth factor A | NM_001025366.3 | F: ttgccttgctgctctacctcca R: gatggcagtagctgcgctgata | 126 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bednarz-Misa, I.; Diakowska, D.; Szczuka, I.; Fortuna, P.; Kubiak, A.; Rosińczuk, J.; Krzystek-Korpacka, M. Interleukins 4 and 13 and Their Receptors Are Differently Expressed in Gastrointestinal Tract Cancers, Depending on the Anatomical Site and Disease Advancement, and Improve Colon Cancer Cell Viability and Motility. Cancers 2020, 12, 1463. https://doi.org/10.3390/cancers12061463
Bednarz-Misa I, Diakowska D, Szczuka I, Fortuna P, Kubiak A, Rosińczuk J, Krzystek-Korpacka M. Interleukins 4 and 13 and Their Receptors Are Differently Expressed in Gastrointestinal Tract Cancers, Depending on the Anatomical Site and Disease Advancement, and Improve Colon Cancer Cell Viability and Motility. Cancers. 2020; 12(6):1463. https://doi.org/10.3390/cancers12061463
Chicago/Turabian StyleBednarz-Misa, Iwona, Dorota Diakowska, Izabela Szczuka, Paulina Fortuna, Agnieszka Kubiak, Joanna Rosińczuk, and Małgorzata Krzystek-Korpacka. 2020. "Interleukins 4 and 13 and Their Receptors Are Differently Expressed in Gastrointestinal Tract Cancers, Depending on the Anatomical Site and Disease Advancement, and Improve Colon Cancer Cell Viability and Motility" Cancers 12, no. 6: 1463. https://doi.org/10.3390/cancers12061463
APA StyleBednarz-Misa, I., Diakowska, D., Szczuka, I., Fortuna, P., Kubiak, A., Rosińczuk, J., & Krzystek-Korpacka, M. (2020). Interleukins 4 and 13 and Their Receptors Are Differently Expressed in Gastrointestinal Tract Cancers, Depending on the Anatomical Site and Disease Advancement, and Improve Colon Cancer Cell Viability and Motility. Cancers, 12(6), 1463. https://doi.org/10.3390/cancers12061463