Complementary Use of Carbohydrate Antigens Lewis a, Lewis b, and Sialyl-Lewis a (CA19.9 Epitope) in Gastrointestinal Cancers: Biological Rationale towards a Personalized Clinical Application



Abstract
:1. Introduction
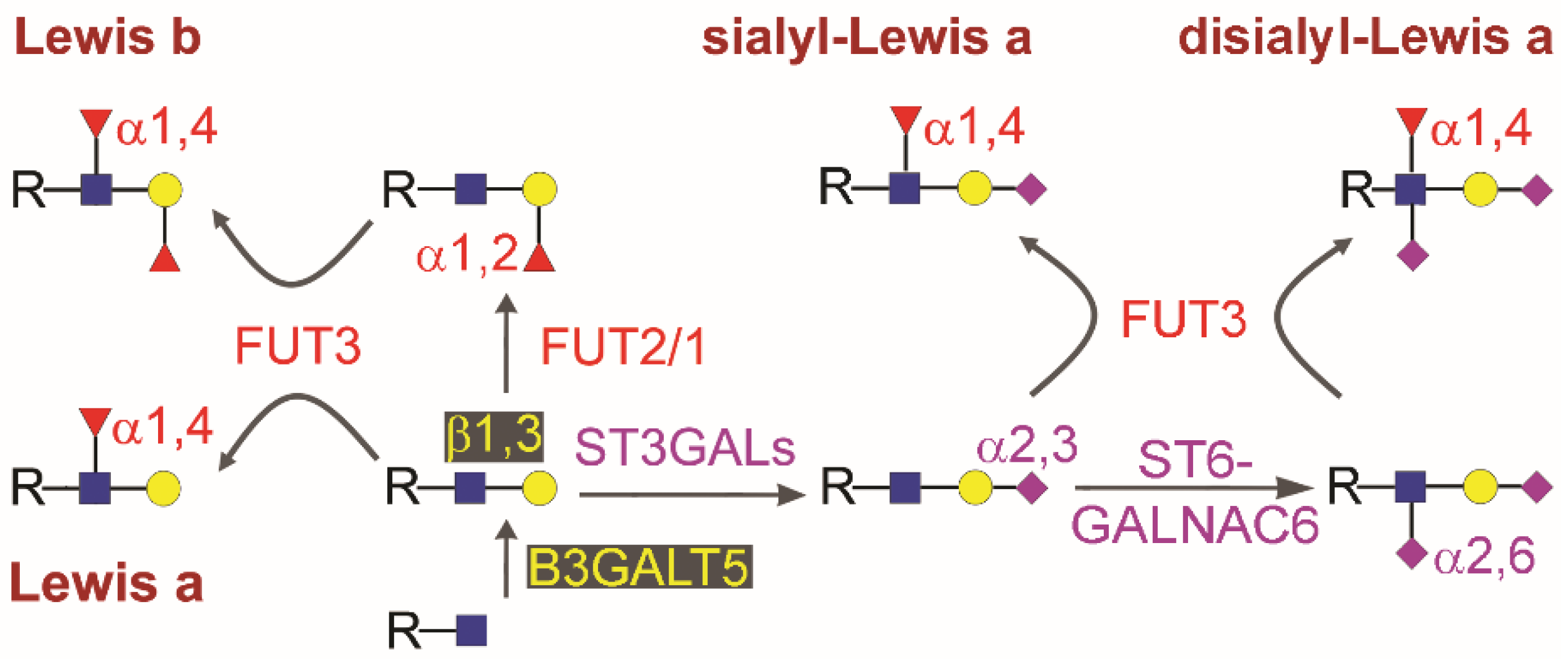
2. Biosynthesis of Type 1 Chain Lewis Antigens in Gastrointestinal Cancers
3. Cancer Cells in Colon Cancer and Pancreatic Ductal Adenocarcinoma Do Not Overproduce CA19.9
4. A Proposed Model of Circulating CA19.9 and Type 1 Chain Lewis Antigen in Pancreatic Ductal Adenocarcinoma (PDAC)
5. CA19.9, Lea, and Leb in Colon Cancer and Other Gastrointestinal Pathologies
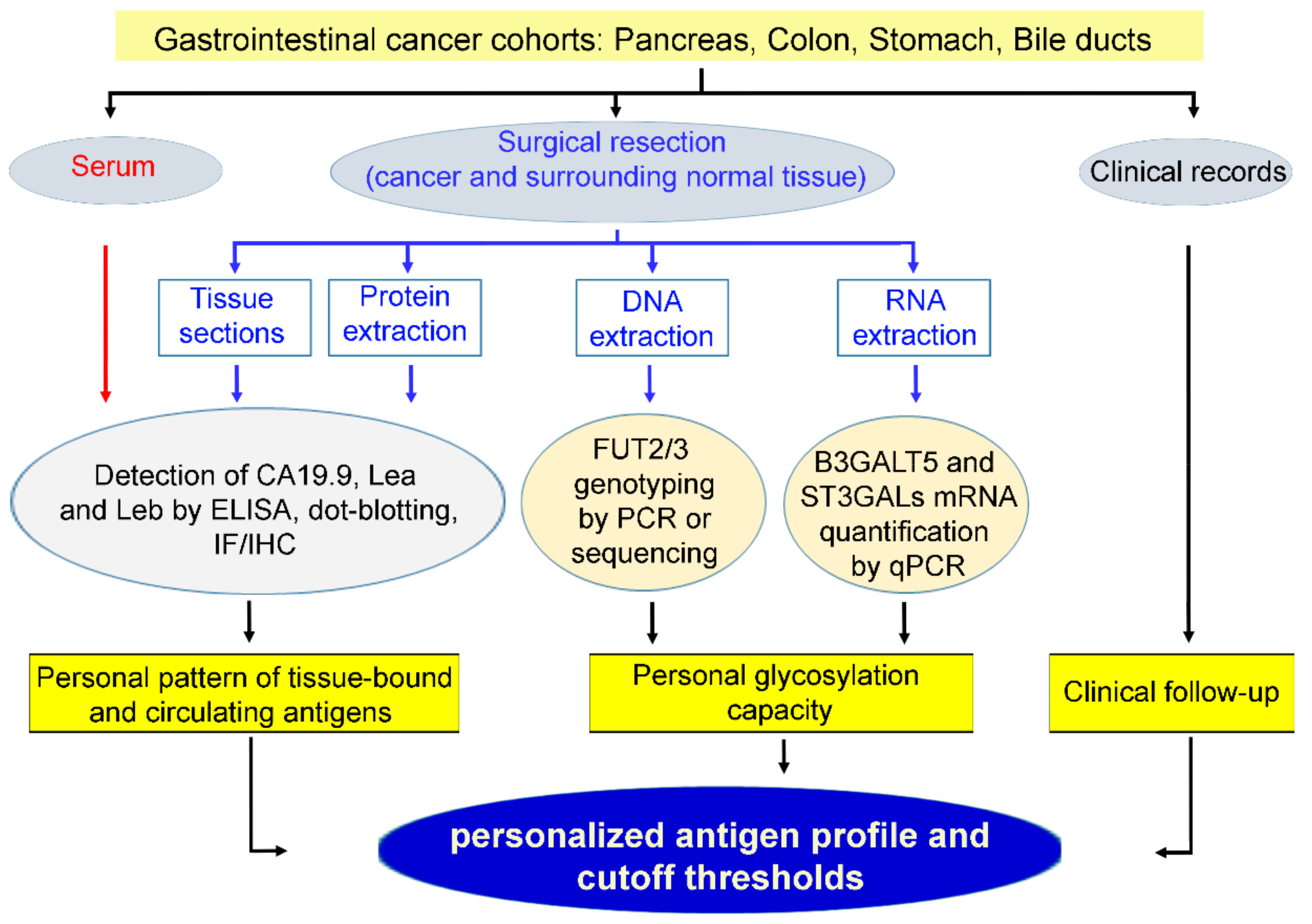
6. Future Applications and Limit of Type 1 Chain Lewis Antigens as Personalized Tumor Markers in Gastrointestinal Cancers
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Das, V.; Kalita, J.; Pal, M. Predictive and prognostic biomarkers in colorectal cancer: A systematic review of recent advances and challenges. Biomed. Pharm. 2017, 87, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Genkinger, J.M.; Su, G.H.; Santella, R.M. Identifying Novel Genetic Markers Through Transcription-Wide Association Study: Can This Be A Path To Reducing The Burden of Pancreatic cancer? J. Natl. Cancer Inst. 2020. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Jermusyk, A.; Wu, L.; Hoskins, J.W.; Collins, I.; Mocci, E.; Zhang, M.; Song, L.; Chung, C.C.; Zhang, T.; et al. A Transcriptome-Wide Association Study (TWAS) Identifies Novel Candidate Susceptibility Genes for Pancreatic Cancer. J. Natl. Cancer Inst. 2020. [Google Scholar] [CrossRef] [PubMed]
- Dell’Aquila, E.; Fulgenzi, C.A.M.; Minelli, A.; Citarella, F.; Stellato, M.; Pantano, F.; Russano, M.; Cursano, M.C.; Napolitano, A.; Zeppola, T.; et al. Prognostic and predictive factors in pancreatic cancer. Oncotarget 2020, 11, 924–941. [Google Scholar] [CrossRef] [Green Version]
- Duffy, M.J.; Sturgeon, C.; Lamerz, R.; Haglund, C.; Holubec, V.L.; Klapdor, R.; Nicolini, A.; Topolcan, O.; Heinemann, V. Tumor markers in pancreatic cancer: A European Group on Tumor Markers (EGTM) status report. Ann. Oncol. Off. J. Eur. Soc. Med Oncol. 2010, 21, 441–447. [Google Scholar] [CrossRef]
- Ge, L.; Pan, B.; Song, F.; Ma, J.; Zeraatkar, D.; Zhou, J.; Tian, J. Comparing the diagnostic accuracy of five common tumour biomarkers and CA19-9 for pancreatic cancer: A protocol for a network meta-analysis of diagnostic test accuracy. BMJ Open 2017, 7, e018175. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.; Yu, X. Selecting chemotherapy for pancreatic cancer: Far away or so close? Semin. Oncol. 2019, 46, 39–47. [Google Scholar] [CrossRef]
- Al Abbas, A.I.; Hogg, M.E. ASO Author Reflections: Serum CA19-9 Utility in Pancreatic Adenocarcinoma in Patients Undergoing Neoadjuvant Therapy. Ann. Surg. Oncol. 2020. [Google Scholar] [CrossRef]
- Capello, M.; Bantis, L.E.; Scelo, G.; Zhao, Y.; Li, P.; Dhillon, D.S.; Patel, N.J.; Kundnani, D.L.; Wang, H.; Abbruzzese, J.L.; et al. Sequential Validation of Blood-Based Protein Biomarker Candidates for Early-Stage Pancreatic Cancer. J. Natl. Cancer Inst. 2017, 109. [Google Scholar] [CrossRef]
- Kim, J.; Bamlet, W.R.; Oberg, A.L.; Chaffee, K.G.; Donahue, G.; Cao, X.J.; Chari, S.; Garcia, B.A.; Petersen, G.M.; Zaret, K.S. Detection of early pancreatic ductal adenocarcinoma with thrombospondin-2 and CA19-9 blood markers. Sci. Transl. Med. 2017, 9. [Google Scholar] [CrossRef] [Green Version]
- Fahrmann, J.F.; Bantis, L.E.; Capello, M.; Scelo, G.; Dennison, J.B.; Patel, N.; Murage, E.; Vykoukal, J.; Kundnani, D.L.; Foretova, L.; et al. A Plasma-Derived Protein-Metabolite Multiplexed Panel for Early-Stage Pancreatic Cancer. J. Natl. Cancer Inst. 2019, 111, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Al Abbas, A.I.; Zenati, M.; Reiser, C.J.; Hamad, A.; Jung, J.P.; Zureikat, A.H.; Zeh, H.J., 3rd; Hogg, M.E. Serum CA19-9 Response to Neoadjuvant Therapy Predicts Tumor Size Reduction and Survival in Pancreatic Adenocarcinoma. Ann. Surg. Oncol. 2020. [Google Scholar] [CrossRef]
- Ragupathi, G.; Damani, P.; Srivastava, G.; Srivastava, O.; Sucheck, S.J.; Ichikawa, Y.; Livingston, P.O. Synthesis of sialyl Lewis(a) (sLe (a), CA19-9) and construction of an immunogenic sLe(a) vaccine. Cancer Immunol. Immunother. 2009, 58, 1397–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawada, R.; Sun, S.M.; Wu, X.; Hong, F.; Ragupathi, G.; Livingston, P.O.; Scholz, W.W. Human monoclonal antibodies to sialyl-Lewis (CA19.9) with potent CDC, ADCC, and antitumor activity. Clin. Cancer Res. 2011, 17, 1024–1032. [Google Scholar] [CrossRef] [Green Version]
- Weitzenfeld, P.; Bournazos, S.; Ravetch, J.V. Antibodies targeting sialyl Lewis A mediate tumor clearance through distinct effector pathways. J. Clin. Investig. 2019, 129, 3952–3962. [Google Scholar] [CrossRef]
- Engle, D.D.; Tiriac, H.; Rivera, K.D.; Pommier, A.; Whalen, S.; Oni, T.E.; Alagesan, B.; Lee, E.J.; Yao, M.A.; Lucito, M.S.; et al. The glycan CA19-9 promotes pancreatitis and pancreatic cancer in mice. Science 2019, 364, 1156–1162. [Google Scholar] [CrossRef]
- Yamashita, K.; Watanabe, M. Clinical significance of tumor markers and an emerging perspective on colorectal cancer. Cancer Sci. 2009, 100, 195–199. [Google Scholar] [CrossRef]
- Galli, C.; Basso, D.; Plebani, M. CA 19-9: Handle with care. Clin. Chem. Lab. Med. 2013, 51, 1369–1383. [Google Scholar] [CrossRef]
- Scarà, S.; Bottoni, P.; Scatena, R. CA 19-9: Biochemical and Clinical Aspects. Adv. Exp. Med. Biol. 2015, 867, 247–260. [Google Scholar] [CrossRef]
- Acharya, A.; Markar, S.R.; Matar, M.; Ni, M.; Hanna, G.B. Use of Tumor Markers in Gastrointestinal Cancers: Surgeon Perceptions and Cost-Benefit Trade-Off Analysis. Ann. Surg. Oncol. 2017, 24, 1165–1173. [Google Scholar] [CrossRef] [Green Version]
- Saengboonmee, C.; Sawanyawisuth, K.; Chamgramol, Y.; Wongkham, S. Prognostic biomarkers for cholangiocarcinoma and their clinical implications. Expert Rev. Anticancer Ther. 2018, 18, 579–592. [Google Scholar] [CrossRef]
- Zhou, W.; Yang, F.; Peng, J.; Wang, F.; Lin, Y.; Jiang, W.; Yang, X.; Li, L.; Lu, Z.; Wan, D.; et al. High pretreatment serum CA19-9 level predicts a poor prognosis for patients with stage III colon cancer after curative resection and adjuvant chemotherapy. J. Cancer 2019, 10, 3810–3818. [Google Scholar] [CrossRef] [Green Version]
- Uhlenbruck, G.; van Meensel-Maene, U.; Hanisch, F.G.; Dienst, C. Unexpected occurrence of the Ca 19-9 tumor marker in normal human seminal plasma. Biol. Chem. 1984, 365, 613–617. [Google Scholar] [CrossRef]
- Baeckstrom, D.; Karlsson, N.; Hansson, G.C. Purification and characterization of sialyl-Le(a)-carrying mucins of human bile; evidence for the presence of MUC1 and MUC3 apoproteins. J. Biol. Chem. 1994, 269, 14430–14437. [Google Scholar] [PubMed]
- Kalthoff, H.; Kreiker, C.; Schmiegel, W.H.; Greten, H.; Thiele, H.G. Characterization of CA 19-9 bearing mucins as physiological exocrine pancreatic secretion products. Cancer Res. 1986, 46, 3605–3607. [Google Scholar]
- Salvini, R.; Bardoni, A.; Valli, M.; Trinchera, M. beta 1,3-Galactosyltransferase beta 3Gal-T5 acts on the GlcNAcbeta 1-->3Galbeta 1-->4GlcNAcbeta 1-->R sugar chains of carcinoembryonic antigen and other N-linked glycoproteins and is down-regulated in colon adenocarcinomas. J. Biol. Chem. 2001, 276, 3564–3573. [Google Scholar] [CrossRef] [Green Version]
- Isshiki, S.; Kudo, T.; Nishihara, S.; Ikehara, Y.; Togayachi, A.; Furuya, A.; Shitara, K.; Kubota, T.; Watanabe, M.; Kitajima, M.; et al. Lewis type 1 antigen synthase (beta3Gal-T5) is transcriptionally regulated by homeoproteins. J. Biol. Chem. 2003, 278, 36611–36620. [Google Scholar] [CrossRef] [Green Version]
- Zulueta, A.; Caretti, A.; Signorelli, P.; Dall’olio, F.; Trinchera, M. Transcriptional control of the B3GALT5 gene by a retroviral promoter and methylation of distant regulatory elements. Faseb J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2014, 28, 946–955. [Google Scholar] [CrossRef] [Green Version]
- Kloppel, G.; Lingenthal, G.; von Bulow, M.; Kern, H.F. Histological and fine structural features of pancreatic ductal adenocarcinomas in relation to growth and prognosis: Studies in xenografted tumours and clinico-histopathological correlation in a series of 75 cases. Histopathology 1985, 9, 841–856. [Google Scholar] [CrossRef]
- Koprowski, H.; Steplewski, Z.; Mitchell, K.; Herlyn, M.; Herlyn, D.; Fuhrer, P. Colorectal carcinoma antigens detected by hybridoma antibodies. Somat. Cell Genet. 1979, 5, 957–971. [Google Scholar] [CrossRef]
- Magnani, J.L.; Nilsson, B.; Brockhaus, M.; Zopf, D.; Steplewski, Z.; Koprowski, H.; Ginsburg, V. A monoclonal antibody-defined antigen associated with gastrointestinal cancer is a ganglioside containing sialylated lacto-N-fucopentaose II. J. Biol. Chem. 1982, 257, 14365–14369. [Google Scholar] [PubMed]
- Magnani, J.L.; Steplewski, Z.; Koprowski, H.; Ginsburg, V. Identification of the gastrointestinal and pancreatic cancer-associated antigen detected by monoclonal antibody 19-9 in the sera of patients as a mucin. Cancer Res. 1983, 43, 5489–5492. [Google Scholar] [PubMed]
- Yue, T.; Partyka, K.; Maupin, K.A.; Hurley, M.; Andrews, P.; Kaul, K.; Moser, A.J.; Zeh, H.; Brand, R.E.; Haab, B.B. Identification of blood-protein carriers of the CA 19-9 antigen and characterization of prevalence in pancreatic diseases. Proteomics 2011, 11, 3665–3674. [Google Scholar] [CrossRef] [Green Version]
- Uozumi, N.; Gao, C.; Yoshioka, T.; Nakano, M.; Moriwaki, K.; Nakagawa, T.; Masuda, T.; Tanabe, M.; Miyoshi, E. Identification of a novel type of CA19-9 carrier in human bile and sera of cancer patients: An implication of the involvement in nonsecretory exocytosis. J. Proteome Res. 2010, 9, 6345–6353. [Google Scholar] [CrossRef] [PubMed]
- Trinchera, M.; Aronica, A.; Dall’Olio, F. Selectin Ligands Sialyl-Lewis a and Sialyl-Lewis x in Gastrointestinal Cancers. Biology 2017, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Glavey, S.V.; Huynh, D.; Reagan, M.R.; Manier, S.; Moschetta, M.; Kawano, Y.; Roccaro, A.M.; Ghobrial, I.M.; Joshi, L.; O’Dwyer, M.E. The cancer glycome: Carbohydrates as mediators of metastasis. Blood Rev. 2015, 29, 269–279. [Google Scholar] [CrossRef]
- Starzonek, S.; Maar, H.; Labitzky, V.; Wicklein, D.; Rossdam, C.; Buettner, F.F.R.; Wolters-Eisfeld, G.; Guengoer, C.; Wagener, C.; Schumacher, U.; et al. Systematic analysis of the human tumor cell binding to human vs. murine E- and P-selectin under static vs. dynamic conditions. Glycobiology 2020, cwaa019. [Google Scholar] [CrossRef] [Green Version]
- Jin, F.; Wang, F. The physiological and pathological roles and applications of sialyl Lewis x, a common carbohydrate ligand of the three selectins. Glycoconj. J. 2020, 37, 277–291. [Google Scholar] [CrossRef]
- Carrascal, M.A.; Talina, C.; Borralho, P.; Goncalo Mineiro, A.; Henriques, A.R.; Pen, C.; Martins, M.; Braga, S.; Sackstein, R.; Videira, P.A. Staining of E-selectin ligands on paraffin-embedded sections of tumor tissue. BMC Cancer 2018, 18, 495. [Google Scholar] [CrossRef]
- Kudo, T.; Ikehara, Y.; Togayachi, A.; Morozumi, K.; Watanabe, M.; Nakamura, M.; Nishihara, S.; Narimatsu, H. Up-regulation of a set of glycosyltransferase genes in human colorectal cancer. Lab. Investig. A J. Tech. Methods Pathol. 1998, 78, 797–811. [Google Scholar]
- Nishihara, S.; Hiraga, T.; Ikehara, Y.; Kudo, T.; Iwasaki, H.; Morozumi, K.; Akamatsu, S.; Tachikawa, T.; Narimatsu, H. Molecular mechanisms of expression of Lewis b antigen and other type I Lewis antigens in human colorectal cancer. Glycobiology 1999, 9, 607–616. [Google Scholar] [CrossRef]
- Carvalho, A.S.; Harduin-Lepers, A.; Magalhaes, A.; Machado, E.; Mendes, N.; Costa, L.T.; Matthiesen, R.; Almeida, R.; Costa, J.; Reis, C.A. Differential expression of alpha-2,3-sialyltransferases and alpha-1,3/4-fucosyltransferases regulates the levels of sialyl Lewis a and sialyl Lewis x in gastrointestinal carcinoma cells. Int. J. Biochem. Cell Biol. 2010, 42, 80–89. [Google Scholar] [CrossRef]
- Gomes, C.; Osorio, H.; Pinto, M.T.; Campos, D.; Oliveira, M.J.; Reis, C.A. Expression of ST3GAL4 leads to SLe(x) expression and induces c-Met activation and an invasive phenotype in gastric carcinoma cells. PLoS ONE 2013, 8, e66737. [Google Scholar] [CrossRef] [PubMed]
- Indellicato, R.; Domenighini, R.; Malagolini, N.; Cereda, A.; Mamoli, D.; Pezzani, L.; Iascone, M.; dall’Olio, F.; Trinchera, M. A novel nonsense and inactivating variant of ST3GAL3 in two infant siblings suffering severe epilepsy and expressing circulating CA19.9. Glycobiology 2020, 30, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, K.; Ohmori, K.; Izawa, M.; Koike, T.; Kumamoto, K.; Furukawa, K.; Ando, T.; Kiso, M.; Yamaji, T.; Hashimoto, Y.; et al. Loss of disialyl Lewis(a), the ligand for lymphocyte inhibitory receptor sialic acid-binding immunoglobulin-like lectin-7 (Siglec-7) associated with increased sialyl Lewis(a) expression on human colon cancers. Cancer Res. 2004, 64, 4498–4505. [Google Scholar] [CrossRef] [Green Version]
- Ito, H.; Hiraiwa, N.; Sawada-Kasugai, M.; Akamatsu, S.; Tachikawa, T.; Kasai, Y.; Akiyama, S.; Ito, K.; Takagi, H.; Kannagi, R. Altered mRNA expression of specific molecular species of fucosyl- and sialyl-transferases in human colorectal cancer tissues. Int. J. Cancer 1997, 71, 556–564. [Google Scholar] [CrossRef]
- Aronica, A.; Avagliano, L.; Caretti, A.; Tosi, D.; Bulfamante, G.P.; Trinchera, M. Unexpected distribution of CA19.9 and other type 1 chain Lewis antigens in normal and cancer tissues of colon and pancreas: Importance of the detection method and role of glycosyltransferase regulation. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 3210–3220. [Google Scholar] [CrossRef]
- Hayashi, N.; Nakamori, S.; Okami, J.; Nagano, H.; Dono, K.; Umeshita, K.; Sakon, M.; Narimatsu, H.; Monden, M. Association between expression levels of CA 19-9 and N-acetylglucosamine-beta;1,3-galactosyltransferase 5 gene in human pancreatic cancer tissue. Pathobiol. J. Immunopathol. Mol. Cell. Biol. 2004, 71, 26–34. [Google Scholar] [CrossRef]
- Perez-Garay, M.; Arteta, B.; Llop, E.; Cobler, L.; Pages, L.; Ortiz, R.; Ferri, M.J.; de Bolos, C.; Figueras, J.; de Llorens, R.; et al. alpha2,3-Sialyltransferase ST3Gal IV promotes migration and metastasis in pancreatic adenocarcinoma cells and tends to be highly expressed in pancreatic adenocarcinoma tissues. Int. J. Biochem. Cell Biol. 2013, 45, 1748–1757. [Google Scholar] [CrossRef]
- Trinchera, M.; Zulueta, A.; Caretti, A.; Dall’Olio, F. Control of Glycosylation-Related Genes by DNA Methylation: The Intriguing Case of the B3GALT5 Gene and Its Distinct Promoters. Biology 2014, 3, 484–497. [Google Scholar] [CrossRef] [Green Version]
- Cai, B.H.; Lee, H.Y.; Chou, C.K.; Wu, P.H.; Huang, H.C.; Chao, C.C.; Chung, H.Y.; Kannagi, R. SSEA3 and Sialyl Lewis a Glycan Expression Is Controlled by B3GALT5 LTR through Lamin A-NFYA and SIRT1-STAT3 Signaling in Human ES Cells. Cells 2020, 9, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohshio, G.; Ogawa, K.; Kudo, H.; Yamabe, H.; Nakashima, Y.; Kim, Y.C.; Endo, K.; Watanabe, Y.; Manabe, T.; Tobe, T. Immunohistochemical studies on the localization of cancer associated antigens DU-PAN-2 and CA19-9 in carcinomas of the digestive tract. J. Gastroenterol. Hepatol. 1990, 5, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Itai, S.; Nishikata, J.; Yoneda, T.; Ohmori, K.; Yamabe, H.; Arii, S.; Tobe, T.; Kannagi, R. Tissue distribution of 2-3 and 2-6 sialyl Lewis A antigens and significance of the ratio of two antigens for the differential diagnosis of malignant and benign disorders of the digestive tract. Cancer 1991, 67, 1576–1587. [Google Scholar] [CrossRef]
- Portela, S.V.; Martin, C.V.; Romay, L.M.; Cuevas, E.; Martin, E.G.; Briera, A.F. sLea and sLex expression in colorectal cancer: Implications for tumourigenesis and disease prognosis. Histol. Histopathol. 2011, 26, 1305–1316. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Takahashi, S.; Gotohda, N.; Konishi, M. Prognostic Impact of the Initial Postoperative CA19-9 Level in Patients with Extrahepatic Bile Duct Cancer. J. Gastrointest. Surg. Off. J. Soc. Surg. Aliment. Tract 2016, 20, 1435–1443. [Google Scholar] [CrossRef] [PubMed]
- Furuya, N.; Kawa, S.; Hasebe, O.; Tokoo, M.; Mukawa, K.; Maejima, S.; Oguchi, H. Comparative study of CA242 and CA19-9 in chronic pancreatitis. Br. J. Cancer 1996, 73, 372–376. [Google Scholar] [CrossRef] [Green Version]
- Zhong, A.; Qin, R.; Qin, W.; Han, J.; Gu, Y.; Zhou, L.; Zhang, H.; Ren, S.; Lu, R.; Guo, L.; et al. Diagnostic Significance of Serum IgG Galactosylation in CA19-9-Negative Pancreatic Carcinoma Patients. Front. Oncol. 2019, 9, 114. [Google Scholar] [CrossRef]
- Wang, T.; Matsuda, Y.; Nonaka, K.; Kakizaki, M.; Ishiwata, T.; Kanazawa, N.; Uegaki, S.; Muramatsu, M.; Sawabe, M.; Mori, S.; et al. Clinicopathological characteristics of gastric cancer with carbohydrate antigen 19-9 expression occurring in elderly individuals: An autopsy study. Pathol. Int. 2020, 70, 92–100. [Google Scholar] [CrossRef]
- Han, E.S.; Lee, H.H.; Lee, J.S.; Song, K.Y.; Park, C.H.; Jeon, H.M. At which stage of gastric cancer progression do levels of carcinoembryonic antigen and carbohydrate antigen 19-9 increase? Application in advanced gastric cancer treatment. J. Gastric Cancer 2014, 14, 123–128. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Dong, A.; Xia, H.; He, G.; Cui, J. Associations between CA19-9 and CA125 levels and human epidermal growth factor receptor 2 overexpression in patients with gastric cancer. Oncol. Lett. 2018, 16, 1079–1086. [Google Scholar] [CrossRef]
- Sigel, C.S.; Drill, E.; Zhou, Y.; Basturk, O.; Askan, G.; Pak, L.M.; Vakiani, E.; Wang, T.; Boerner, T.; Do, R.K.G.; et al. Intrahepatic Cholangiocarcinomas Have Histologically and Immunophenotypically Distinct Small and Large Duct Patterns. Am. J. Surg. Pathol. 2018, 42, 1334–1345. [Google Scholar] [CrossRef] [PubMed]
- Mare, L.; Caretti, A.; Albertini, R.; Trinchera, M. CA19.9 antigen circulating in the serum of colon cancer patients: Where is it from? Int. J. Biochem. Cell Biol. 2013, 45, 792–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Greca, G.; Sofia, M.; Lombardo, R.; Latteri, S.; Ricotta, A.; Puleo, S.; Russello, D. Adjusting CA19-9 values to predict malignancy in obstructive jaundice: Influence of bilirubin and C-reactive protein. World J. Gastroenterol. 2012, 18, 4150–4155. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Guan, X.; Zeng, T.; Wu, S.; Zheng, W.; Fu, H.; Long, T.; Wang, Q.; Meng, Y.; Tian, Y.; et al. The significance of serum IgG4 and CA19-9, autoantibodies in diagnosis and differential diagnosis of IgG4-related sclerosing cholangitis. Scand. J. Gastroenterol. 2018, 53, 206–211. [Google Scholar] [CrossRef]
- Terraneo, L.; Avagliano, L.; Caretti, A.; Bianciardi, P.; Tosi, D.; Bulfamante, G.P.; Samaja, M.; Trinchera, M. Expression of carbohydrate-antigen sialyl-Lewis a on colon cancer cells promotes xenograft growth and angiogenesis in nude mice. Int. J. Biochem. Cell Biol. 2013, 45, 2796–2800. [Google Scholar] [CrossRef] [Green Version]
- Wolf, B.C.; Salem, R.R.; Sears, H.F.; Horst, D.A.; Lavin, P.T.; Herlyn, M.; Itzkowitz, S.H.; Schlom, J.; Steele, G.D., Jr. The expression of colorectal carcinoma-associated antigens in the normal colonic mucosa. An immunohistochemical analysis of regional distribution. Am. J. Pathol. 1989, 135, 111–119. [Google Scholar]
- Nudelman, E.; Fukushi, Y.; Levery, S.B.; Higuchi, T.; Hakomori, S. Novel fucolipids of human adenocarcinoma: Disialosyl Lea antigen (III4FucIII6NeuAcIV3NeuAcLc4) of human colonic adenocarcinoma and the monoclonal antibody (FH7) defining this structure. J. Biol. Chem. 1986, 261, 5487–5495. [Google Scholar]
- Miyazaki, K.; Sakuma, K.; Kawamura, Y.I.; Izawa, M.; Ohmori, K.; Mitsuki, M.; Yamaji, T.; Hashimoto, Y.; Suzuki, A.; Saito, Y.; et al. Colonic epithelial cells express specific ligands for mucosal macrophage immunosuppressive receptors siglec-7 and -9. J. Immunol. 2012, 188, 4690–4700. [Google Scholar] [CrossRef] [Green Version]
- Dupuy, F.; Germot, A.; Marenda, M.; Oriol, R.; Blancher, A.; Julien, R.; Maftah, A. Alpha1,4-fucosyltransferase activity: A significant function in the primate lineage has appeared twice independently. Mol. Biol. Evol. 2002, 19, 815–824. [Google Scholar] [CrossRef] [Green Version]
- Rock, J.; Bloomston, M.; Lozanski, G.; Frankel, W.L. The spectrum of hematologic malignancies involving the pancreas: Potential clinical mimics of pancreatic adenocarcinoma. Am. J. Clin. Pathol. 2012, 137, 414–422. [Google Scholar] [CrossRef] [Green Version]
- Prevot, P.P.; Simion, A.; Grimont, A.; Colletti, M.; Khalaileh, A.; Van den Steen, G.; Sempoux, C.; Xu, X.; Roelants, V.; Hald, J.; et al. Role of the ductal transcription factors HNF6 and Sox9 in pancreatic acinar-to-ductal metaplasia. Gut 2012, 61, 1723–1732. [Google Scholar] [CrossRef]
- Bonastre, E.; Brambilla, E.; Sanchez-Cespedes, M. Cell adhesion and polarity in squamous cell carcinoma of the lung. J. Pathol. 2016, 238, 606–616. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.S.; Cho, Y.M.; Kim, G.H.; Kee, K.H.; Kim, H.S.; Kim, K.M.; Kim, J.H.; Choi, C. Recurrent KRAS mutations identified in papillary renal neoplasm with reverse polarity-a comparative study with papillary renal cell carcinoma. Mod. Pathol. Off. J. U. S. Can. Acad. Pathol. Inc. 2020, 33, 690–699. [Google Scholar] [CrossRef]
- Pareja, F.; da Silva, E.M.; Frosina, D.; Geyer, F.C.; Lozada, J.R.; Basili, T.; Da Cruz Paula, A.; Zhong, E.; Derakhshan, F.; D’Alfonso, T.; et al. Immunohistochemical analysis of IDH2 R172 hotspot mutations in breast papillary neoplasms: Applications in the diagnosis of tall cell carcinoma with reverse polarity. Mod. Pathol. Off. J. U. S. Can. Acad. Pathol. Inc. 2020. [Google Scholar] [CrossRef]
- Narimatsu, H.; Iwasaki, H.; Nakayama, F.; Ikehara, Y.; Kudo, T.; Nishihara, S.; Sugano, K.; Okura, H.; Fujita, S.; Hirohashi, S. Lewis and secretor gene dosages affect CA19-9 and DU-PAN-2 serum levels in normal individuals and colorectal cancer patients. Cancer Res. 1998, 58, 512–518. [Google Scholar]
- Wannhoff, A.; Rupp, C.; Friedrich, K.; Brune, M.; Knierim, J.; Flechtenmacher, C.; Sauer, P.; Stremmel, W.; Hov, J.R.; Schirmacher, P.; et al. Inflammation But Not Biliary Obstruction Is Associated With Carbohydrate Antigen 19-9 Levels in Patients With Primary Sclerosing Cholangitis. Clin. Gastroenterol. Hepatol. Off. Clin. Pract. J. Am. Gastroenterol. Assoc. 2015, 13, 2372–2379. [Google Scholar] [CrossRef]
- Luo, G.; Guo, M.; Jin, K.; Liu, Z.; Liu, C.; Cheng, H.; Lu, Y.; Long, J.; Liu, L.; Xu, J.; et al. Optimize CA19-9 in detecting pancreatic cancer by Lewis and Secretor genotyping. Pancreatology 2016, 16, 1057–1062. [Google Scholar] [CrossRef] [Green Version]
- Abe, T.; Koi, C.; Kohi, S.; Song, K.B.; Tamura, K.; Macgregor-Das, A.; Kitaoka, N.; Chuidian, M.; Ford, M.; Dbouk, M.; et al. Gene Variants That Affect Levels of Circulating Tumor Markers Increase Identification of Patients With Pancreatic Cancer. Clin. Gastroenterol. Hepatol. Off. Clin. Pract. J. Am. Gastroenterol. Assoc. 2020, 18, 1161–1169.e1165. [Google Scholar] [CrossRef]
- Tang, H.; Singh, S.; Partyka, K.; Kletter, D.; Hsueh, P.; Yadav, J.; Ensink, E.; Bern, M.; Hostetter, G.; Hartman, D.; et al. Glycan motif profiling reveals plasma sialyl-lewis x elevations in pancreatic cancers that are negative for sialyl-lewis A. Mol. Cell. Proteom. MCP 2015, 14, 1323–1333. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Partyka, K.; Hsueh, P.; Sinha, J.Y.; Kletter, D.; Zeh, H.; Huang, Y.; Brand, R.E.; Haab, B.B. Glycans related to the CA19-9 antigen are elevated in distinct subsets of pancreatic cancers and improve diagnostic accuracy over CA19-9. Cell. Mol. Gastroenterol. Hepatol. 2016, 2, 201–221. [Google Scholar] [CrossRef] [Green Version]
- Trinchera, M.; Malagolini, N.; Chiricolo, M.; Santini, D.; Minni, F.; Caretti, A.; Dall’olio, F. The biosynthesis of the selectin-ligand sialyl Lewis x in colorectal cancer tissues is regulated by fucosyltransferase VI and can be inhibited by an RNA interference-based approach. Int. J. Biochem. Cell Biol. 2011, 43, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Dall’Olio, F.; Malagolini, N.; Chiricolo, M.; Trinchera, M.; Harduin-Lepers, A. The expanding roles of the Sd(a)/Cad carbohydrate antigen and its cognate glycosyltransferase B4GALNT2. Biochim. Biophys. Acta 2014, 1840, 443–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ong, S.L.; Sachdeva, A.; Garcea, G.; Gravante, G.; Metcalfe, M.S.; Lloyd, D.M.; Berry, D.P.; Dennison, A.R. Elevation of carbohydrate antigen 19.9 in benign hepatobiliary conditions and its correlation with serum bilirubin concentration. Dig. Dis. Sci. 2008, 53, 3213–3237. [Google Scholar] [CrossRef]
- Bertino, G.; Ardiri, A.M.; Calvagno, G.S.; Malaguarnera, G.; Interlandi, D.; Vacante, M.; Bertino, N.; Lucca, F.; Madeddu, R.; Motta, M. Carbohydrate 19.9 antigen serum levels in liver disease. Biomed. Res. Int. 2013, 2013, 531640. [Google Scholar] [CrossRef] [Green Version]
- Mayerle, J.; Kalthoff, H.; Reszka, R.; Kamlage, B.; Peter, E.; Schniewind, B.; González Maldonado, S.; Pilarsky, C.; Heidecke, C.D.; Schatz, P.; et al. Metabolic biomarker signature to differentiate pancreatic ductal adenocarcinoma from chronic pancreatitis. Gut 2018, 67, 128–137. [Google Scholar] [CrossRef]
- Ahn, S.; Lee, J.C.; Hwang, J.H.; Yoon, Y.S.; Lee, K.H.; Zen, Y. Duct-obstructive pancreatitis with granulocytic epithelial lesion in a patient with ulcerative colitis: An atypical manifestation of type 2 autoimmune pancreatitis? Pathol. Int. 2019, 69, 420–426. [Google Scholar] [CrossRef]
- Fukasawa, H.; Kaneko, M.; Niwa, H.; Yasuda, H.; Kumagai, H.; Furuya, R. Carbohydrate antigen 19-9 is significantly elevated in autosomal dominant polycystic kidney disease. Nephrology (Carlton) 2018, 23, 210–216. [Google Scholar] [CrossRef]
- Kajbafzadeh, A.M.; Ladi Seyedian, S.S.; Kameli, S.M.; Nabavizadeh, B.; Boroomand, M.; Moghtaderi, M. Urinary carbohydrate antigen 19-9 level as a biomarker in children with acute pyelonephritis. Eur. J. Pediatr. 2020. [Google Scholar] [CrossRef]
- Chachadi, V.B.; Ali, M.F.; Cheng, P.W. Prostatic cell-specific regulation of the synthesis of MUC1-associated sialyl Lewis a. PLoS ONE 2013, 8, e57416. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, E.; Freitas, R.; Ferreira, D.; Soares, J.; Azevedo, R.; Gaiteiro, C.; Peixoto, A.; Oliveira, S.; Cotton, S.; Relvas-Santos, M.; et al. Nucleolin-Sle A Glycoforms as E-Selectin Ligands and Potentially Targetable Biomarkers at the Cell Surface of Gastric Cancer Cells. Cancers 2020, 12, 861. [Google Scholar] [CrossRef] [Green Version]
- Locker, G.Y.; Hamilton, S.; Harris, J.; Jessup, J.M.; Kemeny, N.; Macdonald, J.S.; Somerfield, M.R.; Hayes, D.F.; Bast, R.C., Jr. ASCO 2006 update of recommendations for the use of tumor markers in gastrointestinal cancer. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2006, 24, 5313–5327. [Google Scholar] [CrossRef]
- Bockhorn, M.; Uzunoglu, F.G.; Adham, M.; Imrie, C.; Milicevic, M.; Sandberg, A.A.; Asbun, H.J.; Bassi, C.; Buchler, M.; Charnley, R.M.; et al. Borderline resectable pancreatic cancer: A consensus statement by the International Study Group of Pancreatic Surgery (ISGPS). Surgery 2014, 155, 977–988. [Google Scholar] [CrossRef]
- Gion, M.; Peloso, L.; Trevisiol, C.; Squarcina, E.; Zappa, M.; Fabricio, A.S. An epidemiology-based model as a tool to monitor the outbreak of inappropriateness in tumor marker requests: A national scale study. Clin. Chem. Lab. Med. 2016, 54, 473–482. [Google Scholar] [CrossRef]
- Accordino, M.K.; Wright, J.D.; Vasan, S.; Neugut, A.I.; Tergas, A.; Hu, J.C.; Hershman, D.L. Serum Tumor Marker Use in Patients With Advanced Solid Tumors. J. Oncol. Pract. 2016, 12, 65–66. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Site | Topic | Findings | Reference |
|---|---|---|---|
| Colon | Glycosylation profile | High B3GALT5 1 in normal mucosa, downregulated in cancer, low ST3GALs and high FUT3 in both ST6GALNAC6 downregulated in cancer | [26,27,28,47] [40,46,47] [45] |
| Tissue antigens by immunofluorescence/dot-blot | High Lea 2 and very low CA19.9 and Leb 3 in normal mucosa (luminal staining), none detected in cancer | [47] | |
| Tissue antigens by immunohistochemistry | CA19.9 more elevated in cancer (frequent stromal and cytoplasmic staining) but present in normal mucosa and benign polyps, Lea and Leb abundant in normal mucosa disialyl-Lea more elevated in normal mucosa but present in cancer | [45,52,53,54] [45,53] | |
| Serum | CA19.9 elevated in cancer at low percentage and late stages only; no data available for Lea and Leb | [17,22] | |
| Pancreas | Glycosylation profile | B4GALT5, FUT3, FUT1/2 and ST3GALs well detectable in both normal tissue and PDAC 4 | [47,48,49] |
| Tissue antigens by immunofluorescence/dot-blot | CA19.9, Lea, and Leb detected in both normal and cancerous ducts (luminal staining), no one in normal acinar cells or poorly differentiated cancer cells | [47] | |
| Tissue antigens by immunohistochemistry | CA19.9 detected in cancer (frequent cytoplasmic staining) better than in normal ducts (luminal staining) | [52,53] | |
| Serum | CA19.9 elevation in PDAC Lea and Leb elevation in PDAC (preliminary) CA19.9 elevation in non-malignant diseases | [4,7,12,55] [this article] [56,57] | |
| Stomach | Glycosylation profile | No data available | |
| Tissue antigens by immunofluorescence/dot-blot | No data available | ||
| Tissue antigens by immunohistochemistry | CA19.9 detected in both cancer and normal mucosa, frequent cytoplasmic and stromal staining; no data about Lea and Leb | [52,53,58] | |
| Serum | CA19.9 elevation in cancer, no data about Lea and Leb | [58,59,60] | |
| Bile ducts | Glycosylation profile | No data available | |
| Tissue antigens by immunofluorescence/dot-blot | No data available | ||
| Tissue antigens by immunohistochemistry | CA19.9 detected in both cancer and normal ducts No data about Lea and Leb | [52,61] | |
| Serum | CA19.9 elevation in both cancer and non-malignant diseases No data about Lea and Leb | [21,55,62,63,64] | |
| Mouse tissues | Tissue antigens by immunofluorescence | CA19.9 undetectable in liver, colon, and HCT-15 5 xenograft; HCT-15-T5 6 xenograft very bright | [65] |
| Tissue antigens by immunohistochemistry | CA19.9 detected in liver, colon, adipose tissue, small intestine and both xenografts (poorly, including stromal staining) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Indellicato, R.; Zulueta, A.; Caretti, A.; Trinchera, M. Complementary Use of Carbohydrate Antigens Lewis a, Lewis b, and Sialyl-Lewis a (CA19.9 Epitope) in Gastrointestinal Cancers: Biological Rationale towards a Personalized Clinical Application. Cancers 2020, 12, 1509. https://doi.org/10.3390/cancers12061509
Indellicato R, Zulueta A, Caretti A, Trinchera M. Complementary Use of Carbohydrate Antigens Lewis a, Lewis b, and Sialyl-Lewis a (CA19.9 Epitope) in Gastrointestinal Cancers: Biological Rationale towards a Personalized Clinical Application. Cancers. 2020; 12(6):1509. https://doi.org/10.3390/cancers12061509
Chicago/Turabian StyleIndellicato, Rossella, Aida Zulueta, Anna Caretti, and Marco Trinchera. 2020. "Complementary Use of Carbohydrate Antigens Lewis a, Lewis b, and Sialyl-Lewis a (CA19.9 Epitope) in Gastrointestinal Cancers: Biological Rationale towards a Personalized Clinical Application" Cancers 12, no. 6: 1509. https://doi.org/10.3390/cancers12061509
APA StyleIndellicato, R., Zulueta, A., Caretti, A., & Trinchera, M. (2020). Complementary Use of Carbohydrate Antigens Lewis a, Lewis b, and Sialyl-Lewis a (CA19.9 Epitope) in Gastrointestinal Cancers: Biological Rationale towards a Personalized Clinical Application. Cancers, 12(6), 1509. https://doi.org/10.3390/cancers12061509