Small Non-Coding RNA Profiling in Plasma Extracellular Vesicles of Bladder Cancer Patients by Next-Generation Sequencing: Expression Levels of miR-126-3p and piR-5936 Increase with Higher Histologic Grades

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Results
2.1. Sample Characteristics
2.2. Sequencing Results
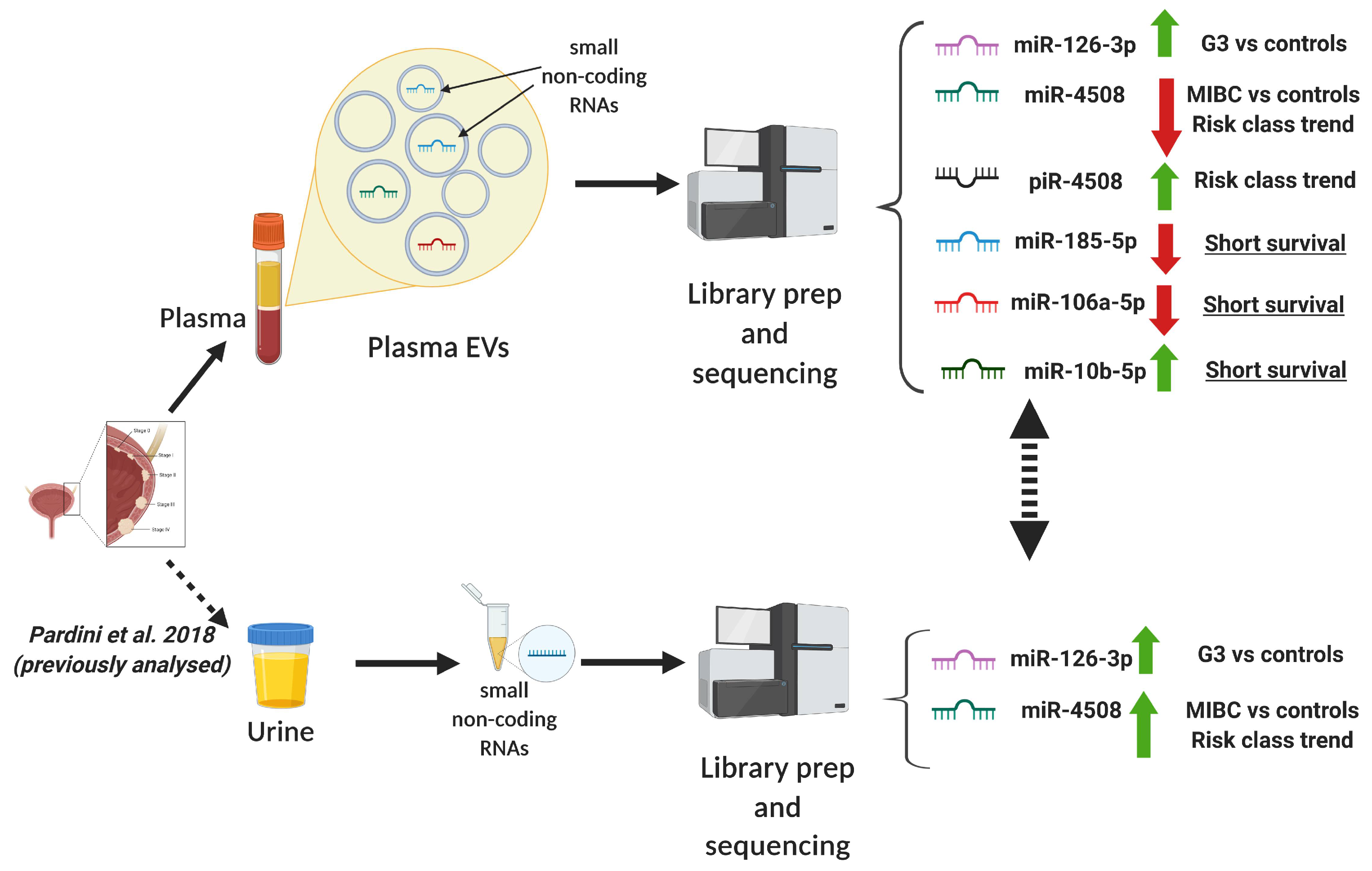
2.3. Differentially expressed sncRNAs in Extracellular Vesicles
2.4. Replication of the Differentially Expressed sncRNAs in Urine of the Same Subjects
2.5. Comparison with miRNA Profiles in Primary Tissues from TCGA
2.6. Functional Implications of Dysregulated miRNAs
2.7. Predictivity of sncRNAs in BC Diagnosis
2.8. Bladder Cancer Prognosis and Survival
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Histological Grading of Bladder Cancers
4.3. Risk Group Stratification
4.4. Plasma Separation and Extracellular Vesicles Precipitation
4.5. RNA Extraction and Quality Control
4.6. Library Preparation for Small RNA-Seq
4.7. Computational and Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA: A Cancer, J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Antoni, S.; Ferlay, J.; Soerjomataram, I.; Znaor, A.; Jemal, A.; Bray, F. Bladder Cancer Incidence and Mortality: A Global Overview and Recent Trends. Eur. Urol. 2017, 71, 96–108. [Google Scholar] [CrossRef]
- Matullo, G.; Naccarati, A.; Pardini, B. MicroRNA expression profiling in bladder cancer: The challenge of next-generation sequencing in tissues and biofluids. Int. J. Cancer 2015, 138, 2334–2345. [Google Scholar] [CrossRef]
- Cumberbatch, M.G.K.; Noon, A.P. Epidemiology, aetiology and screening of bladder cancer. Transl. Androl. Urol. 2019, 8, 5–11. [Google Scholar] [CrossRef] [PubMed]
- European Association of Urology. European Association of Urology Guidelines; 2018 Edition; European Association of Urolog: Arnhem, The Netherlands, 2018. [Google Scholar]
- Babjuk, M.; Burger, M.; Compérat, E.; Gontero, P.; Mostafid, A.H.; Palou, J.; van Rhijn, B.W.G.; Rouprêt, M.; Shariat, S.F.; Sylvester, R.; et al. EAU Guidelines on Non-Muscle-Invasive Bladder Cancer (TaT1 and CIS) 2018. In European Association of Urology Guidelines; 2018 Edition; European Association of Urolog: Arnhem, The Netherlands, 2018. [Google Scholar]
- Lotan, Y.; Roehrborn, C.G. Sensitivity and specificity of commonly available bladder tumor markers versus cytology: Results of a comprehensive literature review and meta-analyses. Urology 2003, 61, 109–118. [Google Scholar] [CrossRef]
- Stav, K.; Leibovici, D.; Goren, E.; Livshitz, A.; Siegel, Y.I.; Lindner, A.; Zisman, A. Adverse effects of cystoscopy and its impact on patients’ quality of life and sexual performance. Isr. Med Assoc. J. 2004, 6, 474–478. [Google Scholar]
- Linxweiler, J.; Junker, K. Extracellular vesicles in urological malignancies: An update. Nat. Rev. Urol. 2019, 17, 11–27. [Google Scholar] [CrossRef] [PubMed]
- Dragomir, M.P.; Knutsen, E.; Calin, G.A. SnapShot: Unconventional miRNA Functions. Cell 2018, 174, 1038. [Google Scholar] [CrossRef]
- Ozata, D.M.; Gainetdinov, I.; Zoch, A.; O’Carroll, D.; Zamore, P. PIWI-interacting RNAs: Small RNAs with big functions. Nat. Rev. Genet. 2018, 20, 89–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anfossi, S.; Babayan, A.; Pantel, K.; Calin, G.A. Clinical utility of circulating non-coding RNAs: An update. Nat. Rev. Clin. Oncol. 2018, 15, 541–563. [Google Scholar] [CrossRef] [PubMed]
- Pardini, B.; Calin, G.A. MicroRNAs and Long Non-Coding RNAs and Their Hormone-Like Activities in Cancer. Cancers 2019, 11, 378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pardini, B.; Cordero, F.; Naccarati, A.; Viberti, C.; Birolo, G.; Oderda, M.; Di Gaetano, C.; Arigoni, M.; Martina, F.; Calogero, R.A.; et al. microRNA profiles in urine by next-generation sequencing can stratify bladder cancer subtypes. Oncotarget 2018, 9, 20658–20669. [Google Scholar] [CrossRef] [PubMed]
- Tölle, A.; Blobel, C.C.; Jung, K. Circulating miRNAs in blood and urine as diagnostic and prognostic biomarkers for bladder cancer: An update in 2017. Biomarkers Med. 2018, 12, 667–676. [Google Scholar] [CrossRef]
- Zeuschner, P.; Linxweiler, J.; Junker, K. Non-coding RNAs as biomarkers in liquid biopsies with a special emphasis on extracellular vesicles in urological malignancies. Expert Rev. Mol. Diagn. 2019, 20, 151–167. [Google Scholar] [CrossRef] [PubMed]
- Santoni, G.; Morelli, M.B.; Amantini, C.; Battelli, N. Urinary Markers in Bladder Cancer: An Update. Front. Oncol. 2018, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuzaki, K.; Fujita, K.; Jingushi, K.; Kawashima, A.; Ujike, T.; Nagahara, A.; Ueda, Y.; Tanigawa, G.; Yoshioka, I.; Ueda, K.; et al. MiR-21-5p in urinary extracellular vesicles is a novel biomarker of urothelial carcinoma. Oncotarget 2017, 8, 24668–24678. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, D.A.; Green, B.B.; Seigne, J.D.; Schned, A.R.; Marsit, C.J. MicroRNA molecular profiling from matched tumor and bio-fluids in bladder cancer. Mol. Cancer 2015, 14, 194. [Google Scholar] [CrossRef] [Green Version]
- De Long, J.; Sullivan, T.B.; Humphrey, J.; Logvinenko, T.; Summerhayes, K.A.; Kozinn, S.; Harty, N.; Summerhayes, I.C.; Libertino, J.A.; Holway, A.H.; et al. A non-invasive miRNA based assay to detect bladder cancer in cell-free urine. Am. J. Transl. Res. 2015, 7, 2500–2509. [Google Scholar]
- Andreu, Z.; Oshiro, R.O.; Redruello, A.; López-Martín, S.; Gutierrez-Vazquez, C.; Morato, E.; Marina, A.I.; Gómez, C.O.; Yáñez-Mó, M. Extracellular vesicles as a source for non-invasive biomarkers in bladder cancer progression. Eur. J. Pharm. Sci. 2017, 98, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Baumgart, S.; Meschkat, P.; Edelmann, P.; Heinzelmann, J.; Pryalukhin, A.; Bohle, R.; Heinzelbecker, J.; Stöckle, M.; Junker, K. MicroRNAs in tumor samples and urinary extracellular vesicles as a putative diagnostic tool for muscle-invasive bladder cancer. J. Cancer Res. Clin. Oncol 2019, 145, 2725–2736. [Google Scholar] [CrossRef]
- Usuba, W.; Urabe, F.; Yamamoto, Y.; Matsuzaki, J.; Sasaki, H.; Ichikawa, M.; Takizawa, S.; Aoki, Y.; Niida, S.; Kato, K.; et al. Circulating miRNA panels for specific and early detection in bladder cancer. Cancer Sci. 2018, 110, 408–419. [Google Scholar] [CrossRef]
- Khetrapal, P.; Lee, M.W.L.; Tan, W.S.; Dong, L.; De Winter, P.; Feber, A.; Kelly, J.D. The role of circulating tumour cells and nucleic acids in blood for the detection of bladder cancer: A systematic review. Cancer Treat. Rev. 2018, 66, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Hurd, P.; Nelson, C.J. Advantages of next-generation sequencing versus the microarray in epigenetic research. Briefings Funct. Genom. Proteom. 2009, 8, 174–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanke, M.; Hoefig, K.; Merz, H.; Feller, A.C.; Kausch, I.; Jocham, D.; Warnecke, J.M.; Sczakiel, G. A robust methodology to study urine microRNA as tumor marker: microRNA-126 and microRNA-182 are related to urinary bladder cancer. Urol. Oncol. Semin. Orig. Investig. 2010, 28, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Pardini, B.; Sabo, A.A.; Birolo, G.; Calin, G.A. Noncoding RNAs in Extracellular Fluids as Cancer Biomarkers: The New Frontier of Liquid Biopsies. Cancers 2019, 11, 1170. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Anderson, C.; Hanus, J.; Zhao, F.; Ma, J.; Yoshimura, A.; Wang, S. Strand and Cell Type-specific Function of microRNA-126 in Angiogenesis. Mol. Ther. 2016, 24, 1823–1835. [Google Scholar] [CrossRef] [Green Version]
- Pitzler, L.; Auler, M.; Probst, K.; Frie, C.; Bergmeier, V.; Holzer, T.; Belluoccio, D.; Bergen, J.V.D.; Etich, J.; Ehlen, H.; et al. miR-126-3p Promotes Matrix-Dependent Perivascular Cell Attachment, Migration and Intercellular Interaction. Stem Cells 2016, 34, 1297–1309. [Google Scholar] [CrossRef] [Green Version]
- Grimolizzi, F.; Monaco, F.; Leoni, F.; Bracci, M.; Staffolani, S.; Bersaglieri, C.; Gaetani, S.; Valentino, M.; Amati, M.; Rubini, C.; et al. Exosomal miR-126 as a circulating biomarker in non-small-cell lung cancer regulating cancer progression. Sci. Rep. 2017, 7, 15277. [Google Scholar] [CrossRef]
- Meister, J.; Schmidt, M.H.H. miR-126 and miR-126*: New Players in Cancer. Sci. World, J. 2010, 10, 2090–2100. [Google Scholar] [CrossRef]
- Jin, Y.; Jiang, Z.; Guan, X.; Chen, Y.; Tang, Q.; Wang, G.; Wang, X. miR-450b-5p Suppresses Stemness and the Development of Chemoresistance by Targeting SOX2 in Colorectal Cancer. DNA Cell Biol. 2016, 35, 249–256. [Google Scholar] [CrossRef]
- Ye, Y.-P.; Wu, P.; Gu, C.-C.; Deng, D.-L.; Jiao, H.-L.; Li, T.-T.; Wang, S.-Y.; Wang, Y.-X.; Xiao, Z.-Y.; Wei, W.-T.; et al. miR-450b-5p induced by oncogenic KRAS is required for colorectal cancer progression. Oncotarget 2016, 7, 61312–61324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Chen, J.-J.; Xia, S.-H.; Zhao, H.; Yang, J.-B.; Zhang, H.; He, B.; Jiao, J.; Zhan, B.-T.; Sun, C.-C. Long intergenic non-protein coding RNA 319 aggravates lung adenocarcinoma carcinogenesis by modulating miR-450b-5p/EZH2. Gene 2018, 650, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Svoboda, M.; Sana, J.; Fabian, P.; Kocáková, I.; Gombosova, J.; Nekvindova, J.; Radova, L.; Vyzula, R.; Slaby, O. MicroRNA expression profile associated with response to neoadjuvant chemoradiotherapy in locally advanced rectal cancer patients. Radiat. Oncol. 2012, 7, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, A.C.; Vira, M.; Shen, J.; Sanda, M.G.; Raman, J.; Liao, J.; Patil, D.; Taioli, E. Circulating microRNAs in plasma as potential biomarkers for the early detection of prostate cancer. Prostate 2018, 78, 411–418. [Google Scholar] [CrossRef]
- Zhang, H.; Jin, K. Peripheral Circulating Exosomal miRNAs Potentially Contribute to the Regulation of Molecular Signaling Networks in Aging. Int. J. Mol. Sci. 2020, 21, 1908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Gao, J.; Yu, Y.; Zhao, Z.; Pan, Y. Long non-coding RNA UCA1 targets miR-185-5p and regulates cell mobility by affecting epithelial-mesenchymal transition in melanoma via Wnt/β-catenin signaling pathway. Gene 2018, 676, 298–305. [Google Scholar] [CrossRef]
- Ostadrahimi, S.; Fayaz, S.; Parvizhamidi, M.; Abedi-Valugerdi, M.; Hassan, M.; Kadivar, M.; Teimoori-Toolabi, L.; Asgari, M.; Shahrokh, H.; Abolhasani, M.; et al. Downregulation of miR-1266-5P, miR-185-5P and miR-30c-2 in prostatic cancer tissue and cell lines. Oncol. Lett. 2018, 15, 8157–8164. [Google Scholar] [CrossRef]
- Mearini, E.; Poli, G.; Cochetti, G.; Boni, A.; Egidi, M.G.; Brancorsini, S. Expression of urinary miRNAs targeting NLRs inflammasomes in bladder cancer. OncoTargets Ther. 2017, 10, 2665–2673. [Google Scholar] [CrossRef] [Green Version]
- Shin, S.-S.; Park, S.-S.; Hwang, B.; Kim, W.T.; Choi, Y.H.; Kim, W.-J.; Moon, S.-K. MicroRNA-106a suppresses proliferation, migration, and invasion of bladder cancer cells by modulating MAPK signaling cell cycle regulators, and Ets-1-mediated MMP-2 expression. Oncol. Rep. 2016, 36, 2421–2429. [Google Scholar] [CrossRef] [Green Version]
- Sheedy, P.; Medarova, Z. The fundamental role of miR-10b in metastatic cancer. Am. J. Cancer Res. 2018, 8, 1674–1688. [Google Scholar]
- Xiao, H.; Li, H.; Yu, G.; Xiao, W.; Hu, J.; Tang, K.; Zeng, J.; He, W.; Zeng, G.; Zhang, X.; et al. MicroRNA-10b promotes migration and invasion through KLF4 and HOXD10 in human bladder cancer. Oncol. Rep. 2014, 31, 1832–1838. [Google Scholar] [CrossRef] [Green Version]
- Zaravinos, A.; Radojičić, J.; Lambrou, G.I.; Volanis, D.; Delakas, D.; Stathopoulos, E.N.; Spandidos, D. Expression of miRNAs Involved in Angiogenesis, Tumor Cell Proliferation, Tumor Suppressor Inhibition, Epithelial-Mesenchymal Transition and Activation of Metastasis in Bladder Cancer. J. Urol. 2012, 188, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Tracey, E.; Watt, H.; Currow, D.; Young, J.; Armstrong, B. Investigation of poorer bladder cancer survival in women in NSW, Australia: A data linkage study. BJU Int. 2014, 113, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Sharples, R.A.; Scicluna, B.J.; Hill, A.F. Exosomes provide a protective and enriched source of miRNA for biomarker profiling compared to intracellular and cell-free blood. J. Extracell. Vesicles 2014, 3, 263. [Google Scholar] [CrossRef]
- Schageman, J.; Zeringer, E.; Li, M.; Barta, T.; Lea, K.; Gu, J.; Magdaleno, S.; Setterquist, R.; Vlassov, A. The Complete Exosome Workflow Solution: From Isolation to Characterization of RNA Cargo. BioMed Res. Int. 2013, 2013, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Turinetto, V.; Pardini, B.; Allione, A.; Fiorito, G.; Viberti, C.; Guarrera, S.; Russo, A.; Anglesio, S.; Redda, M.G.R.; Casetta, G.; et al. H2AX phosphorylation level in peripheral blood mononuclear cells as an event-free survival predictor for bladder cancer. Mol. Carcinog. 2015, 55, 1833–1842. [Google Scholar] [CrossRef]
- Sacerdote, C.; Guarrera, S.; Ricceri, F.; Pardini, B.; Polidoro, S.; Allione, A.; Critelli, R.; Russo, A.; Andrew, A.S.; Ye, Y.; et al. Polymorphisms in the XRCC1 gene modify survival of bladder cancer patients treated with chemotherapy. Int. J. Cancer 2013, 133, 2004–2009. [Google Scholar] [CrossRef] [Green Version]
- Compérat, E.; Burger, M.; Gontero, P.; Mostafid, A.H.; Palou, J.; Rouprêt, M.; Van Rhijn, B.W.; Shariat, S.F.; Sylvester, R.J.; Zigeuner, R.; et al. Grading of Urothelial Carcinoma and The New “World Health Organisation Classification of Tumours of the Urinary System and Male Genital Organs 2016”. Eur. Urol. Focus 2019, 5, 457–466. [Google Scholar] [CrossRef]
- Ferrero, G.; Cordero, F.; Tarallo, S.; Arigoni, M.; Riccardo, F.; Gallo, G.; Ronco, G.; Allasia, M.; Kulkarni, N.; Matullo, G.; et al. Small non-coding RNA profiling in human biofluids and surrogate tissues from healthy individuals: Description of the diverse and most represented species. Oncotarget 2017, 9, 3097–3111. [Google Scholar] [CrossRef] [Green Version]
- MIQE guidelines. Available online: http://miqe.gene-quantification.info/ (accessed on 7 June 2019).
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet.journal 2011, 17, 10. [Google Scholar] [CrossRef]
- Li, H. Exploring single-sample SNP and INDEL calling with whole-genome de novo assembly. Bioinformatics 2012, 28, 1838–1844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sticht, C.; De La Torre, C.; Parveen, A.; Gretz, N. miRWalk: An online resource for prediction of microRNA binding sites. PLoS ONE 2018, 13, e0206239. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’Ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [Green Version]
- R Package. Available online: http://www.sthda.com/english/wiki/survminer-r-package-survival-data-analysis-and-visualization (accessed on 7 June 2019).
- Eissa, S.; Safwat, M.; Matboli, M.; Zaghloul, A.; El-Sawalhi, M.; Shaheen, A. Measurement of Urinary Level of a Specific Competing endogenous RNA network (FOS and RCAN mRNA/ miR-324-5p, miR-4738-3p, /lncRNA miR-497-HG) Enables Diagnosis of Bladder Cancer. Urol. Oncol. Semin. Orig. Investig. 2019, 37, 292.e19–292.e27. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Covariates | Categories | Cases n = 47 | Controls n = 46 | p-Value |
|---|---|---|---|---|
| Age | Mean (Median) | 65.0 (66.4) | 64.3 (65.7) | 0.91 |
| Range | 47.7–73.7 | 46.4–74.5 | ||
| Smoking | Non-smoker | 5 | 5 | 0.95 |
| Former smoker | 26 | 24 | ||
| Current smoker | 16 | 17 | ||
| WHO 1973 | G1 | 12 | ||
| G2 | 16 | |||
| G3 | 11 | |||
| WHO 2004/2016 | HG ** | 30 | ||
| LG | 17 | |||
| Tumor type | NMIBC | 39 | ||
| MIBC | 8 | |||
| Risk * | 1 | 11 | ||
| 2 | 10 | |||
| 3 | 18 | |||
| 4 (MIBC) | 8 |
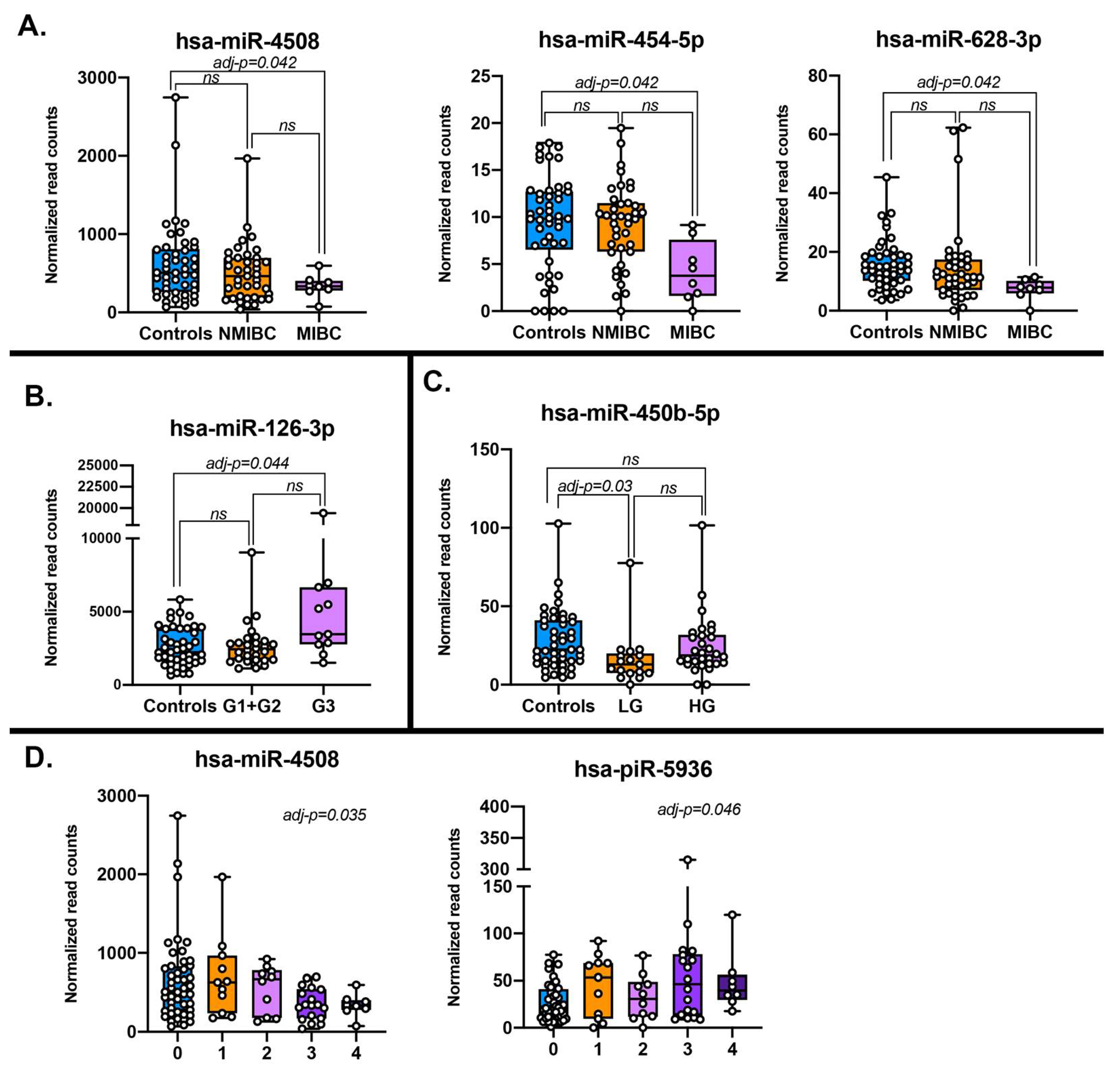
| sncRNAs | Source | Comparison | Base Mean | log2 FC | Nominal p-Value | Adjusted p-Value * |
|---|---|---|---|---|---|---|
| miR-126-3p | Plasma EVs | G3 vs CO | 3571.00 | 0.83 | 8.46 × 10−5 | 4.43 × 10−2 |
| miR-126-3p | Urine | G3 vs CO | 101.00 | 2.41 | 2.11 × 10−7 | 1.63 × 10−6 |
| miR-126 | Tissue (TCGA) | Tumor vs Normal | 5504.14 | 0.26 | 9.35 × 10−2 | 1.31 × 10−1 |
| miR-3140-3p | Plasma EVs | MIBC vs CO | 3.16 | −3.46 | 2.05 × 10−5 | 1.06 × 10−2 |
| miR-4508 | Plasma EVs | MIBC vs CO | 653.24 | −1.23 | 1.62 × 10−4 | 4.16 × 10−2 |
| miR-4508 | Urine | MIBC vs CO | 54.28 | 1.24 | 9.38 × 10−3 | 2.48 × 10−2 |
| miR-4508 | Plasma EVs | Risk class trend | 542.91 | −0.20 | 1.12 × 10−4 | 3.54 × 10−2 |
| miR-4508 | Urine | Risk class trend | 231.79 | 0.32 | 1.98 × 10−3 | 6.40 × 10−3 |
| miR-450b-5p | Plasma EVs | LG vs CO | 26.10 | −1.10 | 5.92 × 10−5 | 3.05 × 10−2 |
| miR-450b-5p | Urine | LG vs CO | 5.26 | −0.07 | 8.96 × 10−1 | 1.00 |
| miR-450b | Tissue (TCGA) | Tumor vs Normal | 53.7 | −0.59 | 3.87 × 10−2 | 5.85 × 10−2 |
| miR-454-5p | Plasma EVs | MIBC vs CO | 9.99 | −1.36 | 3.22 × 10−4 | 4.16 × 10−2 |
| miR-454 | Tissue (TCGA) | Tumor vs Normal | 26.37 | −1.05 | 1.48 × 10−7 | 6.74 × 10−7 |
| miR-628-3p | Plasma EVs | MIBC vs CO | 16.58 | −1.22 | 3.09 × 10−4 | 4.16 × 10−2 |
| miR-628-3p | Urine | MIBC vs CO | 11.27 | 2.32 | 7.84 × 10−5 | 3.76 × 10−4 |
| miR-628 | Tissue (TCGA) | Tumor vs Normal | 31.68 | 0.23 | 4.18 × 10−1 | 4.87 × 10−1 |
| piR-hsa-5936 | Plasma EVs | Risk class trend | 37.01 | 0.29 | 2.92 × 10−4 | 4.61 × 10−2 |
| piR-hsa-5936 | Urine | Risk class trend | 540.58 | −0.09 | 2.86 × 10−1 | 3.89 × 10−1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sabo, A.A.; Birolo, G.; Naccarati, A.; Dragomir, M.P.; Aneli, S.; Allione, A.; Oderda, M.; Allasia, M.; Gontero, P.; Sacerdote, C.; et al. Small Non-Coding RNA Profiling in Plasma Extracellular Vesicles of Bladder Cancer Patients by Next-Generation Sequencing: Expression Levels of miR-126-3p and piR-5936 Increase with Higher Histologic Grades. Cancers 2020, 12, 1507. https://doi.org/10.3390/cancers12061507
Sabo AA, Birolo G, Naccarati A, Dragomir MP, Aneli S, Allione A, Oderda M, Allasia M, Gontero P, Sacerdote C, et al. Small Non-Coding RNA Profiling in Plasma Extracellular Vesicles of Bladder Cancer Patients by Next-Generation Sequencing: Expression Levels of miR-126-3p and piR-5936 Increase with Higher Histologic Grades. Cancers. 2020; 12(6):1507. https://doi.org/10.3390/cancers12061507
Chicago/Turabian StyleSabo, Alexandru A., Giovanni Birolo, Alessio Naccarati, Mihnea P. Dragomir, Serena Aneli, Alessandra Allione, Marco Oderda, Marco Allasia, Paolo Gontero, Carlotta Sacerdote, and et al. 2020. "Small Non-Coding RNA Profiling in Plasma Extracellular Vesicles of Bladder Cancer Patients by Next-Generation Sequencing: Expression Levels of miR-126-3p and piR-5936 Increase with Higher Histologic Grades" Cancers 12, no. 6: 1507. https://doi.org/10.3390/cancers12061507
APA StyleSabo, A. A., Birolo, G., Naccarati, A., Dragomir, M. P., Aneli, S., Allione, A., Oderda, M., Allasia, M., Gontero, P., Sacerdote, C., Vineis, P., Matullo, G., & Pardini, B. (2020). Small Non-Coding RNA Profiling in Plasma Extracellular Vesicles of Bladder Cancer Patients by Next-Generation Sequencing: Expression Levels of miR-126-3p and piR-5936 Increase with Higher Histologic Grades. Cancers, 12(6), 1507. https://doi.org/10.3390/cancers12061507