
Small RNAs in Seminal Plasma as Novel Biomarkers for Germ Cell Tumors

 ,
,  , , , , ,
, , , , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Subjects
2.2. Isolation of Seminal Plasma
2.3. RNA Isolation from Seminal Plasma
2.4. Sequencing Library Preparation
2.5. Small RNA Alignment and Annotation
2.6. Analysis of Read-Counts of Human Small RNAs
2.7. Random Forests Classification
2.8. qPCR Quantification
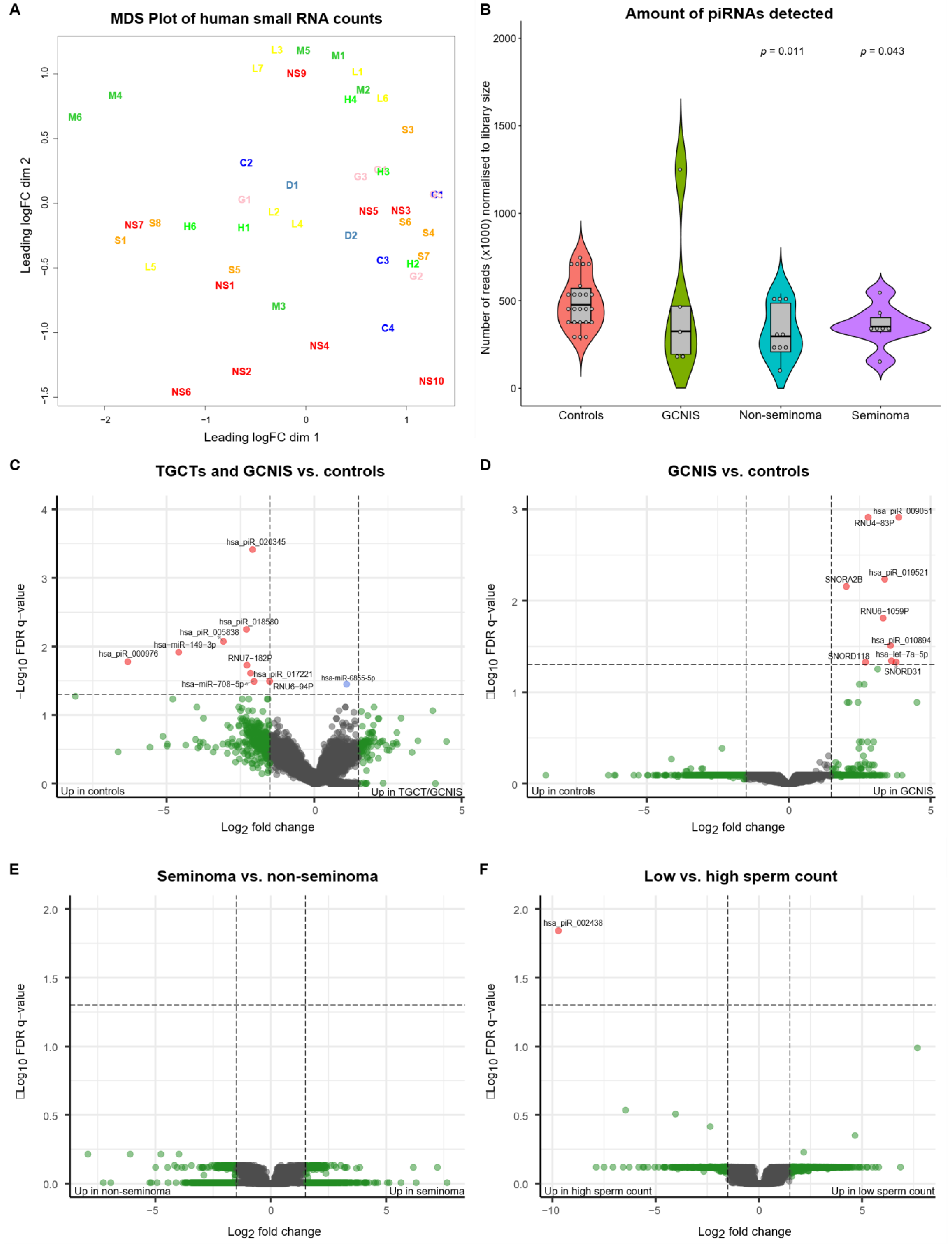
3. Results
3.1. Small RNAs Differentially Present in Seminal Plasma
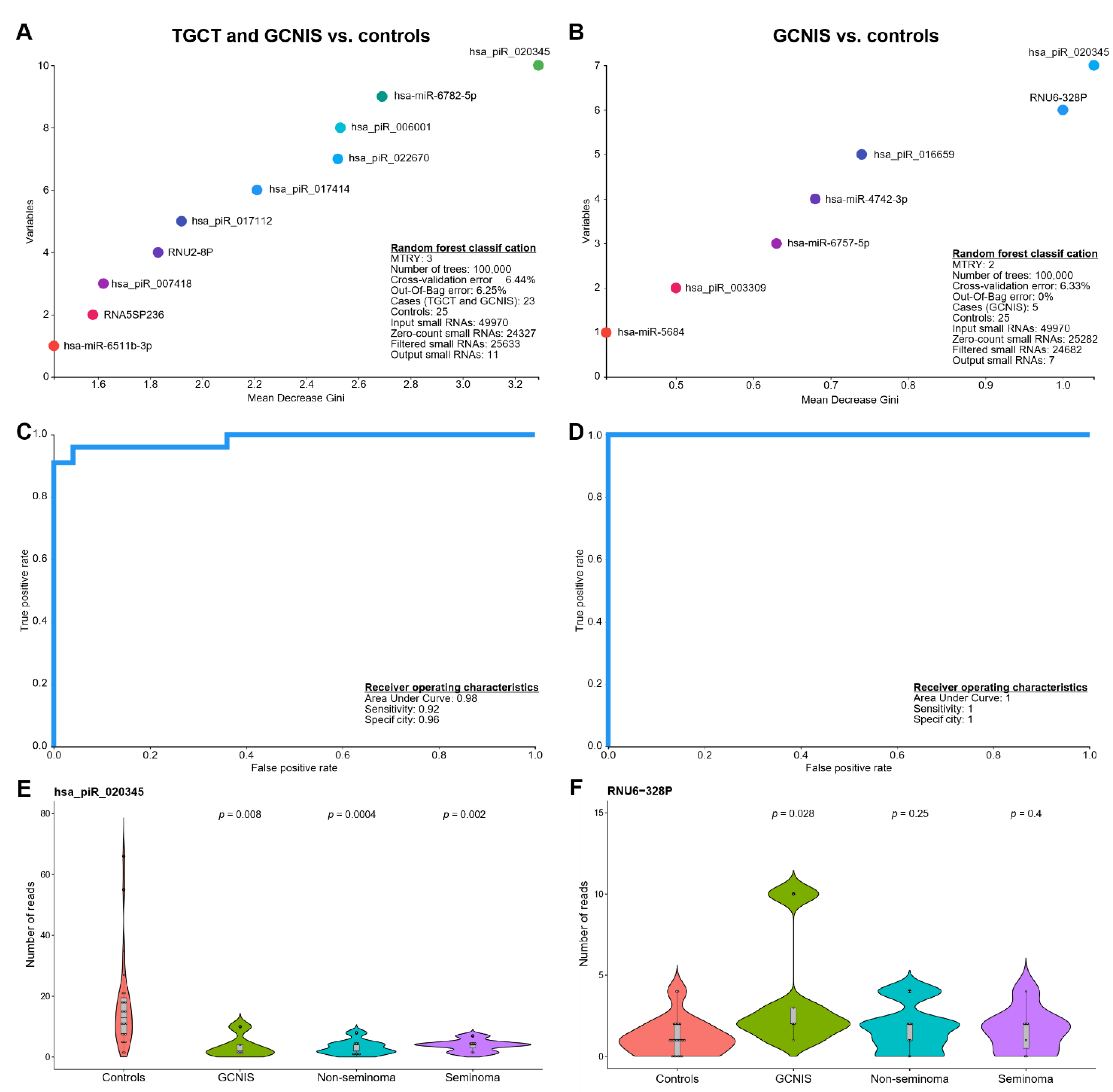
3.2. Random Forests Classification
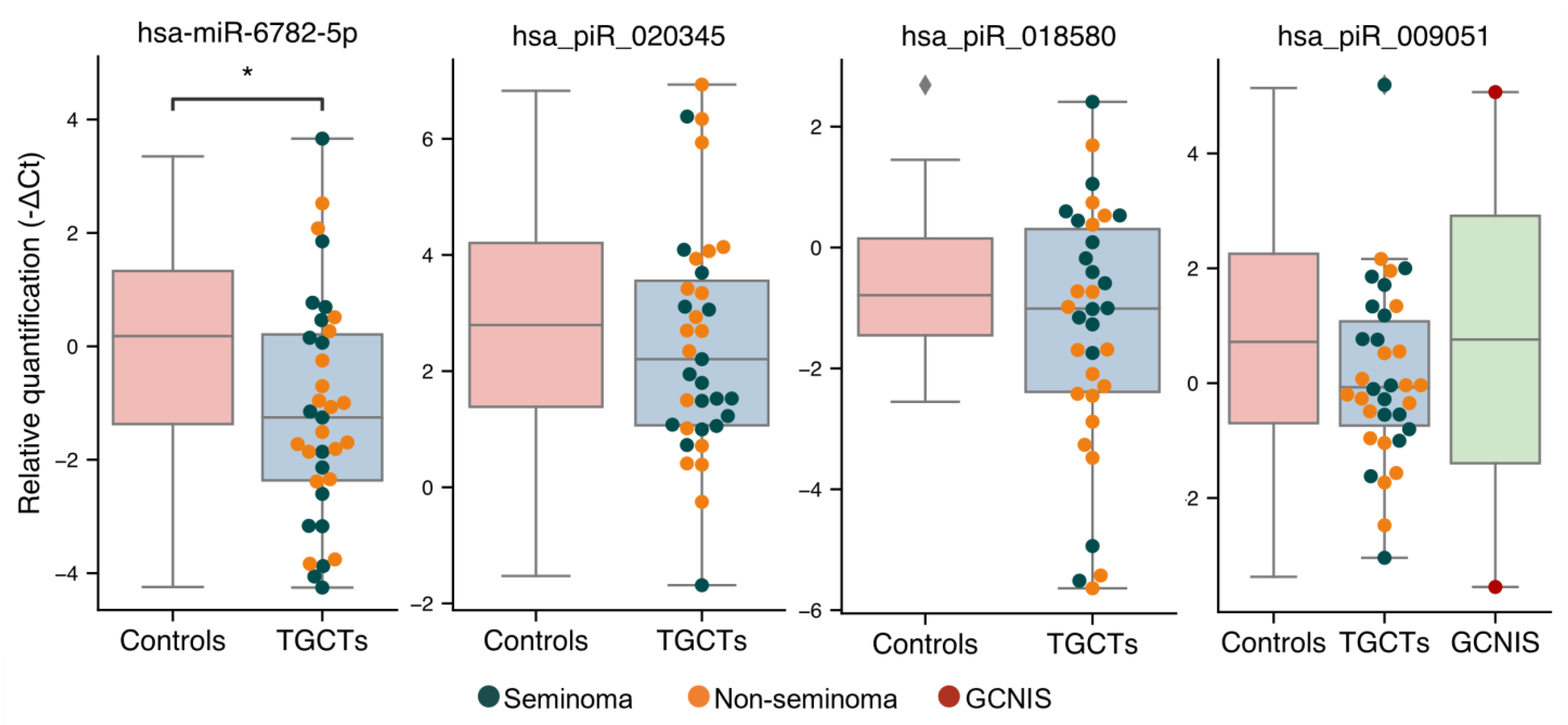
3.3. Validation of Sequencing Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rajpert-De Meyts, E.; McGlynn, K.A.; Okamoto, K.; Jewett, M.A.S.; Bokemeyer, C. Testicular germ cell tumours. Lancet 2016, 387, 1762–1774. [Google Scholar] [CrossRef]
- Almstrup, K.; Hoei-Hansen, C.E.; Wirkner, U.; Blake, J.; Schwager, C.; Ansorge, W.; Nielsen, J.E.; Skakkebaek, N.E.; Rajpert-De Meyts, E.; Leffers, H. Embryonic stem cell-like features of testicular carcinoma in situ revealed by genome-wide gene expression profiling. Cancer Res. 2004, 64, 4736–4743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonne, S.B.; Almstrup, K.; Dalgaard, M.; Juncker, A.S.; Edsgard, D.; Ruban, L.; Harrison, N.J.; Schwager, C.; Abdollahi, A.; Huber, P.E.; et al. Analysis of gene expression profiles of microdissected cell populations indicates that testicular carcinoma in situ is an arrested gonocyte. Cancer Res. 2009, 69, 5241–5250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skakkebaek, N.E.; Berthelsen, J.G.; Giwercman, A.; Müller, J. Carcinoma-in-situ of the testis: Possible origin from gonocytes and precursor of all types of germ cell tumours except spermatocytoma. Int. J. Androl. 1987, 10, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Rajpert-De Meyts, E.; Nielsen, J.E.; Skakkebaek, N.E.; Almstrup, K. Diagnostic markers for germ cell neoplasms: From placental-like alkaline phosphatase to micro-RNAs. Folia Histochem. Cytobiol. 2015, 53, 177–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoei-Hansen, C.E.; Rajpert-De Meyts, E.; Daugaard, G.; Skakkebaek, N.E. Carcinoma in situ testis, the progenitor of testicular germ cell tumours: A clinical review. Ann. Oncol. 2005, 16, 863–868. [Google Scholar] [CrossRef]
- Kier, M.G.G.; Lauritsen, J.; Almstrup, K.; Mortensen, M.S.; Toft, B.G.; Rajpert-De Meyts, E.; Skakkebaek, N.E.; Rørth, M.; von der Maase, H.; Agerbaek, M.; et al. Screening for carcinoma in situ in the contralateral testicle in patients with testicular cancer: A population-based study. Ann. Oncol. 2015, 26, 737–742. [Google Scholar] [CrossRef]
- Hoei-Hansen, C.E.; Olesen, I.A.; Jorgensen, N.; Carlsen, E.; Holm, M.; Almstrup, K.; Leffers, H.; Rajpert-De Meyts, E. Current approaches for detection of carcinoma in situ testis. Int. J. Androl. 2007, 30, 398–404, discussion 404-5. [Google Scholar] [CrossRef]
- Giwercman, A.; Marks, A.; Skakkebaek, N.E. Carcinoma-in-situ germ-cells exfoliated from seminiferous epithelium into seminal fluid. Lancet 1988, 1, 530. [Google Scholar] [CrossRef]
- van Casteren, N.J.; Stoop, H.; Dohle, G.R.; de Wit, R.; Oosterhuis, J.W.; Looijenga, L.H.J. Noninvasive detection of testicular carcinoma in situ in semen using OCT3/4. Eur. Urol. 2008, 54, 153–158. [Google Scholar] [CrossRef]
- Almstrup, K.; Lippert, M.; Mogensen, H.O.; Nielsen, J.E.; Hansen, J.D.; Daugaard, G.; Jørgensen, N.; Foged, N.T.; Skakkebaek, N.E.; Rajpert-De Meyts, E. Screening of subfertile men for testicular carcinoma in situ by an automated image analysis-based cytological test of the ejaculate. Int. J. Androl. 2011, 34, e21–e30, discussion e30-1. [Google Scholar] [CrossRef]
- Nielsen, J.E.; Kristensen, D.M.; Almstrup, K.; Jørgensen, A.; Olesen, I.A.; Jacobsen, G.K.; Horn, T.; Skakkebaek, N.E.; Leffers, H.; Rajpert-De Meyts, E. A novel double staining strategy for improved detection of testicular carcinoma in situ cells in human semen samples. Andrologia 2012, 44, 78–85. [Google Scholar] [CrossRef]
- Almstrup, K.; Lobo, J.; Mørup, N.; Belge, G.; Rajpert-De Meyts, E.; Looijenga, L.H.J.; Dieckmann, K.-P. Application of miRNAs in the diagnosis and monitoring of testicular germ cell tumours. Nat. Rev. Urol. 2020, 17, 201–213. [Google Scholar] [CrossRef]
- Radtke, A.; Cremers, J.-F.; Kliesch, S.; Riek, S.; Junker, K.; Mohamed, S.A.; Anheuser, P.; Belge, G.; Dieckmann, K.-P. Can germ cell neoplasia in situ be diagnosed by measuring serum levels of microRNA371a-3p? J. Cancer Res. Clin. Oncol. 2017, 143, 2383–2392. [Google Scholar] [CrossRef] [Green Version]
- Dieckmann, K.-P.; Radtke, A.; Geczi, L.; Matthies, C.; Anheuser, P.; Eckardt, U.; Sommer, J.; Zengerling, F.; Trenti, E.; Pichler, R.; et al. Serum Levels of MicroRNA-371a-3p (M371 Test) as a New Biomarker of Testicular Germ Cell Tumors: Results of a Prospective Multicentric Study. J. Clin. Oncol. 2019, 37, 1412–1423. [Google Scholar] [CrossRef] [PubMed]
- Dieckmann, K.-P.; Spiekermann, M.; Balks, T.; Flor, I.; Löning, T.; Bullerdiek, J.; Belge, G. MicroRNAs miR-371-3 in serum as diagnostic tools in the management of testicular germ cell tumours. Br. J. Cancer 2012, 107, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Dieckmann, K.-P.; Spiekermann, M.; Balks, T.; Ikogho, R.; Anheuser, P.; Wosniok, W.; Loening, T.; Bullerdiek, J.; Belge, G. MicroRNA miR-371a-3p—A Novel Serum Biomarker of Testicular Germ Cell Tumors: Evidence for Specificity from Measurements in Testicular Vein Blood and in Neoplastic Hydrocele Fluid. Urol. Int. 2016, 97, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Spiekermann, M.; Belge, G.; Winter, N.; Ikogho, R.; Balks, T.; Bullerdiek, J.; Dieckmann, K.-P. MicroRNA miR-371a-3p in serum of patients with germ cell tumours: Evaluations for establishing a serum biomarker. Andrology 2015, 3, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.J.; Bell, E.; Raby, K.L.; Rijlaarsdam, M.A.; Gillis, A.J.M.; Looijenga, L.H.J.; Brown, H.; Destenaves, B.; Nicholson, J.C.; Coleman, N. A pipeline to quantify serum and cerebrospinal fluid microRNAs for diagnosis and detection of relapse in paediatric malignant germ-cell tumours. Br. J. Cancer 2016, 114, 151–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radtke, A.; Dieckmann, K.-P.; Grobelny, F.; Salzbrunn, A.; Oing, C.; Schulze, W.; Belge, G. Expression of miRNA-371a-3p in seminal plasma and ejaculate is associated with sperm concentration. Andrology 2019, 7, 469–474. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Rahman, R.-U.; Gautam, A.; Bethune, J.; Sattar, A.; Fiosins, M.; Magruder, D.S.; Capece, V.; Shomroni, O.; Bonn, S. Oasis 2: Improved online analysis of small RNA-seq data. BMC Bioinform. 2018, 19, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, D.J.; Chen, Y.; Smyth, G.K. Differential expression analysis of multifactor RNA-Seq experiments with respect to biological variation. Nucleic Acids Res. 2012, 40, 4288–4297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kassambara, A. ggplot2. Based Publication Ready Plots [R Package Ggpubr Version 0.2.5] 2020. Available online: https://CRAN.R-project.org/package=ggpubr (accessed on 9 September 2020).
- Blighe, K.; Rana, S.; Lewis, M. EnhancedVolcano: Publication-Ready Volcano Plots with Enhanced Colouring and Labeling. Available online: https://github.com/kevinblighe/EnhancedVolcano (accessed on 9 September 2020).
- Breiman, L. Random forests. Mach. Learn. 2001. [Google Scholar] [CrossRef] [Green Version]
- Cutler, D.R.; Edwards, T.C., Jr.; Beard, K.H.; Cutler, A.; Hess, K.T.; Gibson, J.; Lawler, J.J. Random forests for classification in ecology. Ecology 2007, 88, 2783–2792. [Google Scholar] [CrossRef]
- Schuster, A.; Tang, C.; Xie, Y.; Ortogero, N.; Yuan, S.; Yan, W. SpermBase: A Database for Sperm-Borne RNA Contents. Biol. Reprod. 2016, 95, 99. [Google Scholar] [CrossRef] [Green Version]
- Girard, A.; Sachidanandam, R.; Hannon, G.J.; Carmell, M.A. A germline-specific class of small RNAs binds mammalian Piwi proteins. Nature 2006, 442, 199–202. [Google Scholar] [CrossRef]
- Liu, Y.; Dou, M.; Song, X.; Dong, Y.; Liu, S.; Liu, H.; Tao, J.; Li, W.; Yin, X.; Xu, W. The emerging role of the piRNA/piwi complex in cancer. Mol. Cancer 2019, 18, 123. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, H.J.; Heyn, H.; Garcia del Muro, X.; Vidal, A.; Larriba, S.; Muñoz, C.; Villanueva, A.; Esteller, M. Epigenetic loss of the PIWI/piRNA machinery in human testicular tumorigenesis. Epigenetics 2014, 9, 113–118. [Google Scholar] [CrossRef]
- Gainetdinov, I.V.; Skvortsova, Y.V.; Kondratieva, S.A.; Klimov, A.; Tryakin, A.A.; Azhikina, T.L. Assessment of piRNA biogenesis and function in testicular germ cell tumors and their precursor germ cell neoplasia in situ. BMC Cancer 2018, 18, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, K.; Trzybulska, D.; Tsatsanis, C.; Giwercman, A.; Almstrup, K. Identification of circulating small non-coding RNAs in relation to male subfertility and reproductive hormones. Mol. Cell. Endocrinol. 2019, 492, 110443. [Google Scholar] [CrossRef] [PubMed]
- Pelloni, M.; Coltrinari, G.; Paoli, D.; Pallotti, F.; Lombardo, F.; Lenzi, A.; Gandini, L. Differential expression of miRNAs in the seminal plasma and serum of testicular cancer patients. Endocrine 2017, 57, 518–527. [Google Scholar] [CrossRef]
- Abu-Halima, M.; Galata, V.; Backes, C.; Keller, A.; Hammadeh, M.; Meese, E. MicroRNA signature in spermatozoa and seminal plasma of proven fertile men and in testicular tissue of men with obstructive azoospermia. Andrologia 2020, 52, e13503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Yang, C.; Chen, X.; Yao, B.; Yang, C.; Zhu, C.; Li, L.; Wang, J.; Li, X.; Shao, Y.; et al. Altered profile of seminal plasma microRNAs in the molecular diagnosis of male infertility. Clin. Chem. 2011, 57, 1722–1731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, L.; Wu, C.; Guo, C.; Li, H.; Xiong, C. Identification of microRNAs predominately derived from testis and epididymis in human seminal plasma. Clin. Biochem. 2014, 47, 967–972. [Google Scholar] [CrossRef]
- Vojtech, L.; Woo, S.; Hughes, S.; Levy, C.; Ballweber, L.; Sauteraud, R.P.; Strobl, J.; Westerberg, K.; Gottardo, R.; Tewari, M.; et al. Exosomes in human semen carry a distinctive repertoire of small non-coding RNAs with potential regulatory functions. Nucleic Acids Res. 2014, 42, 7290–7304. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Sample ID | Group | Specification | Pool No. | CATS INDEX | Library Size after Trimming |
|---|---|---|---|---|---|
| C1 | Cryo. control | Leydig cell tumor | 1 | 1 | 10,179,217 |
| C2 | Cryo. control | TC post-orchiectomy | 1 | 10 | 10,450,156 |
| C3 | Cryo. control | Extragonadal GCT | 2 | 3 | 6,312,806 |
| C4 | Cryo. control | Epidermoid cyst | 2 | 11 | 6,248,375 |
| D1 | Donor | Donor | 1 | 3 | 8,279,605 |
| D2 | Donor | Donor | 2 | 2 | 4,786,872 |
| L1 | Low sperm conc. | 13.4 mill./mL | 1 | 15 | 7,064,674 |
| L2 | Low sperm conc. | 15 mill./mL | 1 | 16 | 8,091,871 |
| L3 | Low sperm conc. | 3.4 mill./mL | 1 | 23 | 9,558,667 |
| L4 | Low sperm conc. | 9.2 mill./mL | 2 | 15 | 6,148,043 |
| L5 | Low sperm conc. | 9.7 mill./mL | 2 | 16 | 4,333,736 |
| L6 | Low sperm conc. | 5 mill./mL | 2 | 22 | 6,123,967 |
| L7 | Low sperm conc. | 1.7 mill./mL | 2 | 25 | 8,784,711 |
| M1 | Medium sperm conc. | 38 mill./mL | 1 | 5 | 7,346,572 |
| M2 | Medium sperm conc. | 22 mill./mL | 1 | 12 | 6,260,922 |
| M3 | Medium sperm conc. | 23 mill./mL | 1 | 14 | 5,976,608 |
| M4 | Medium sperm conc. | 36 mill./mL | 2 | 9 | 4,393,984 |
| M5 | Medium sperm conc. | 55 mill./mL | 2 | 10 | 10,054,901 |
| M6 | Medium sperm conc. | 41 mill./mL | 2 | 27 | 3,817,299 |
| H1 | High sperm conc. | 110 mill./mL | 1 | 13 | 6,461,267 |
| H2 | High sperm conc. | 127 mill./mL | 1 | 19 | 5,617,401 |
| H3 | High sperm conc. | 121 mill./mL | 1 | 20 | 6,522,668 |
| H4 | High sperm conc. | 145 mill./mL | 2 | 13 | 6,334,582 |
| H5 * | High sperm conc. | 115 mill./mL | 2 | 14 | 4,762,306 |
| H6 | High sperm conc. | 166 mill./mL | 2 | 21 | 5,564,816 |
| G1 | GCNIS | GCNIS | 1 | 6 | 1,924,818 |
| G2 | GCNIS | GCNIS | 1 | 22 | 8,499,015 |
| G3 | GCNIS | GCNIS | 1 | 25 | 30,972,666 |
| G4 | GCNIS | GCNIS | 2 | 7 | 2,166,453 |
| G5 | GCNIS | GCNIS | 2 | 23 | 10,127,381 |
| NS1 | Non-Seminoma | EC | 1 | 2 | 6,447,654 |
| NS2 | Non-Seminoma | EC, S and YST | 1 | 7 | 4,518,334 |
| NS3 | Non-Seminoma | Unknown NS | 1 | 8 | 9,098,448 |
| NS4 | Non-Seminoma | Unknown NS | 1 | 18 | 7,998,662 |
| NS5 | Non-Seminoma | EC, T, S, CHC and YST | 1 | 27 | 10,326,884 |
| NS6 | Non-Seminoma | Unknown NS | 2 | 4 | 5,249,774 |
| NS7 | Non-Seminoma | EC and T | 2 | 6 | 8,180,864 |
| NS8 * | Non-Seminoma | EC and S | 2 | 8 | 5,302,999 |
| NS9 | Non-Seminoma | Mixed NS | 2 | 12 | 349,856 |
| NS10 | Non-Seminoma | EC | 2 | 20 | 7,490,740 |
| S1 | Seminoma | Seminoma | 1 | 4 | 6,494,618 |
| S2 * | Seminoma | Seminoma | 1 | 9 | 4,800,592 |
| S3 | Seminoma | Seminoma | 1 | 11 | 1,444,440 |
| S4 | Seminoma | Seminoma | 1 | 21 | 7,050,519 |
| S5 | Seminoma | Seminoma | 2 | 1 | 5,277,460 |
| S6 | Seminoma | Seminoma | 2 | 5 | 7,338,222 |
| S7 | Seminoma | Seminoma | 2 | 18 | 10,023,884 |
| S8 | Seminoma | Seminoma | 2 | 19 | 4,732,843 |
| Small RNA | LogFc | LogCPM | LogRatio | p-Value | FDR q-Value |
|---|---|---|---|---|---|
| TGCT/GCNIS vs. controls | |||||
| hsa_piR_020345 # | −2.09 | 3.61 | 31.60 | 1.9 × 10−8 | 0.00039 |
| hsa_piR_018580 # | −2.30 | 1.75 | 25.07 | 5.5 × 10−7 | 0.00564 |
| hsa_piR_005838 | −3.08 | 1.27 | 23.51 | 1.2 × 10−6 | 0.00844 |
| hsa-miR-149-3p § | −4.59 | 7.38 | 22.26 | 2.4 × 10−6 | 0.01217 |
| hsa_piR_000976 | −6.32 | 6.32 | 21.23 | 4.1 × 10−6 | 0.01669 |
| RNU7-182P | −2.28 | 1.50 | 20.65 | 5.5 × 10−6 | 0.01880 |
| hsa_piR_017221 | −2.16 | 1.99 | 19.84 | 8.4 × 10−6 | 0.02457 |
| RNU6-94P | −1.51 | 3.15 | 19.07 | 1.3 × 10−5 | 0.03217 |
| hsa-miR-708-5p § | −2.04 | 1.76 | 18.84 | 1.4 × 10−5 | 0.03231 |
| hsa-miR-6855-5p | 1.10 | 3.80 | 18.46 | 1.7 × 10−5 | 0.03555 |
| GCNIS vs. controls | |||||
| hsa_piR_009051 #,§ | 3.88 | 3.86 | 28.06 | 1.2 × 10−7 | 0.00122 |
| RNU4-83P | 2.80 | 1.85 | 28.02 | 1.2 × 10−7 | 0.00122 |
| hsa_piR_019521 § | 3.39 | 3.97 | 24.24 | 8.5 × 10−7 | 0.00581 |
| SNORA2B | 2.03 | 3.42 | 23.33 | 1.4 × 10−6 | 0.00698 |
| RNU6-1059P | 3.33 | 3.81 | 21.37 | 3.8 × 10−6 | 0.01549 |
| hsa_piR_010894 § | 3.59 | 2.42 | 19.70 | 9.0 × 10−6 | 0.03082 |
| hsa-let-7a-5p § | 3.62 | 1.93 | 18.67 | 1.6 × 10−5 | 0.04548 |
| SNORD31 | 3.79 | 1.60 | 18.31 | 1.9 × 10−5 | 0.04694 |
| SNORD118 | 2.71 | 2.21 | 18.13 | 2.1 × 10−5 | 0.04694 |
| Low vs. high sperm concentrations | |||||
| hsa_piR_002438 | −9.71 | 6.71 | 24.60 | 7.1 × 10−7 | 0.01442 |
| Rank | Small RNA | Type | Mean Decrease Gini | Mean Decrease Accuracy |
|---|---|---|---|---|
| TGCT/GCNIS vs. controls | ||||
| 1 | hsa_piR_020345 #,* | piRNA | 3.29 | 0.05 |
| 2 | hsa-miR-6782-5p # | miRNA | 2.69 | 0.02 |
| 3 | hsa_piR_006001 | piRNA | 2.53 | 0.03 |
| 4 | hsa_piR_022670 | piRNA | 2.52 | 0.04 |
| 5 | hsa_piR_017414 | piRNA | 2.21 | 0.02 |
| 6 | hsa_piR_017112 | piRNA | 1.92 | 0.02 |
| 7 | RNU2-8P | snRNA | 1.83 | 0.02 |
| 8 | hsa_piR_007418 | piRNA | 1.62 | 0.01 |
| 9 | RNA5SP236 | rRNA | 1.58 | 0.01 |
| 10 | hsa-miR-6511b-3p | miRNA | 1.43 | 0.01 |
| 11 | hsa-miR-6796-3p | miRNA | 1.37 | 0.01 |
| GCNIS vs. controls | ||||
| 1 | hsa_piR_020345 #,* | piRNA | 1.04 | 0.02 |
| 2 | RNU6-328P | snRNA | 1.00 | 0.02 |
| 3 | hsa_piR_016659 § | piRNA | 0.74 | 0.01 |
| 4 | hsa-miR-4742-3p | miRNA | 0.68 | 0.01 |
| 5 | hsa-miR-6757-5p | miRNA | 0.63 | 0.01 |
| 6 | hsa_piR_003309 | piRNA | 0.50 | 0.00 |
| 7 | hsa-miR-5684 | miRNA | 0.41 | 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mørup, N.; Stakaitis, R.; Golubickaite, I.; Riera, M.; Dalgaard, M.D.; Schierup, M.H.; Jørgensen, N.; Daugaard, G.; Juul, A.; Almstrup, K. Small RNAs in Seminal Plasma as Novel Biomarkers for Germ Cell Tumors. Cancers 2021, 13, 2346. https://doi.org/10.3390/cancers13102346
Mørup N, Stakaitis R, Golubickaite I, Riera M, Dalgaard MD, Schierup MH, Jørgensen N, Daugaard G, Juul A, Almstrup K. Small RNAs in Seminal Plasma as Novel Biomarkers for Germ Cell Tumors. Cancers. 2021; 13(10):2346. https://doi.org/10.3390/cancers13102346
Chicago/Turabian StyleMørup, Nina, Rytis Stakaitis, Ieva Golubickaite, Meritxell Riera, Marlene Danner Dalgaard, Mikkel H. Schierup, Niels Jørgensen, Gedske Daugaard, Anders Juul, and Kristian Almstrup. 2021. "Small RNAs in Seminal Plasma as Novel Biomarkers for Germ Cell Tumors" Cancers 13, no. 10: 2346. https://doi.org/10.3390/cancers13102346
APA StyleMørup, N., Stakaitis, R., Golubickaite, I., Riera, M., Dalgaard, M. D., Schierup, M. H., Jørgensen, N., Daugaard, G., Juul, A., & Almstrup, K. (2021). Small RNAs in Seminal Plasma as Novel Biomarkers for Germ Cell Tumors. Cancers, 13(10), 2346. https://doi.org/10.3390/cancers13102346