
MicroRNAs Possibly Involved in the Development of Bone Metastasis in Clear-Cell Renal Cell Carcinoma

 , ,
, ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Selection
2.2. Study Objectives and Endpoints
2.3. MiRNA Extraction
2.4. mRNA Extraction
2.5. Selection of miRNAs and Genes
2.6. Clinical Data
2.7. Statistical Analysis
3. Results
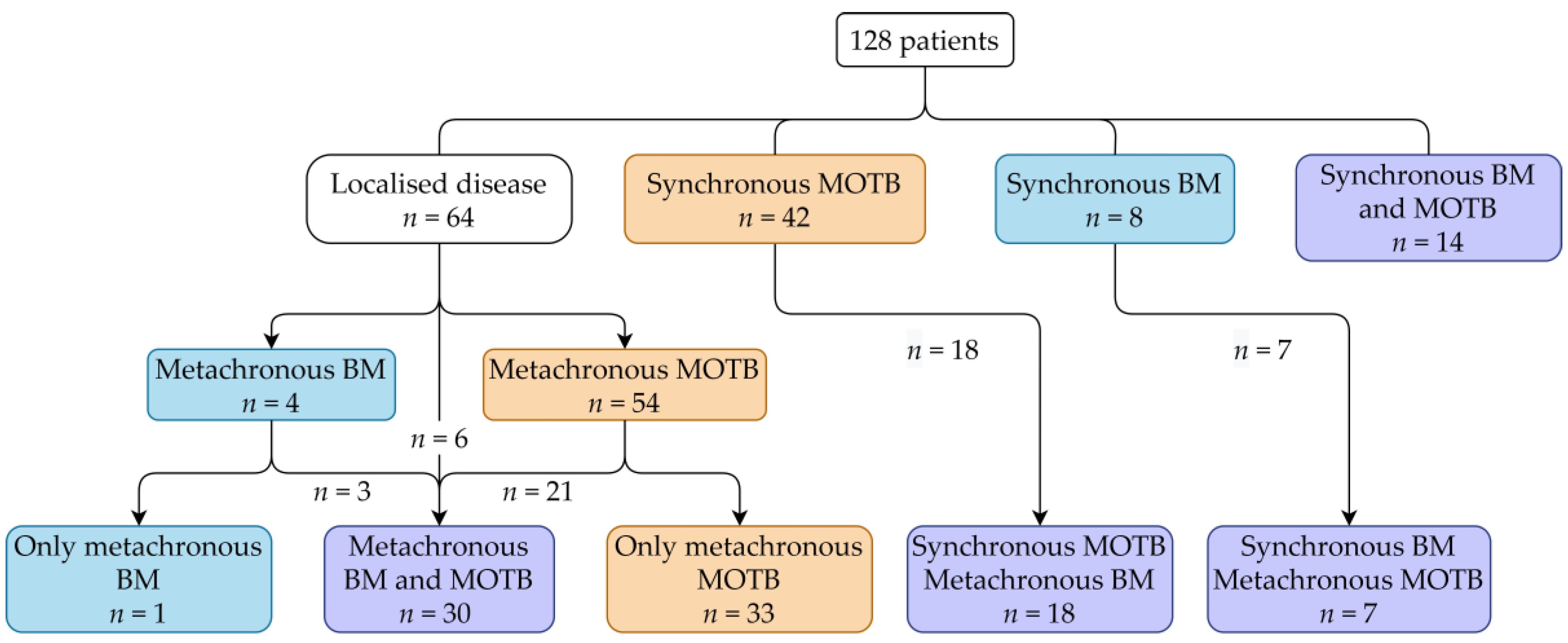
3.1. Included Patients
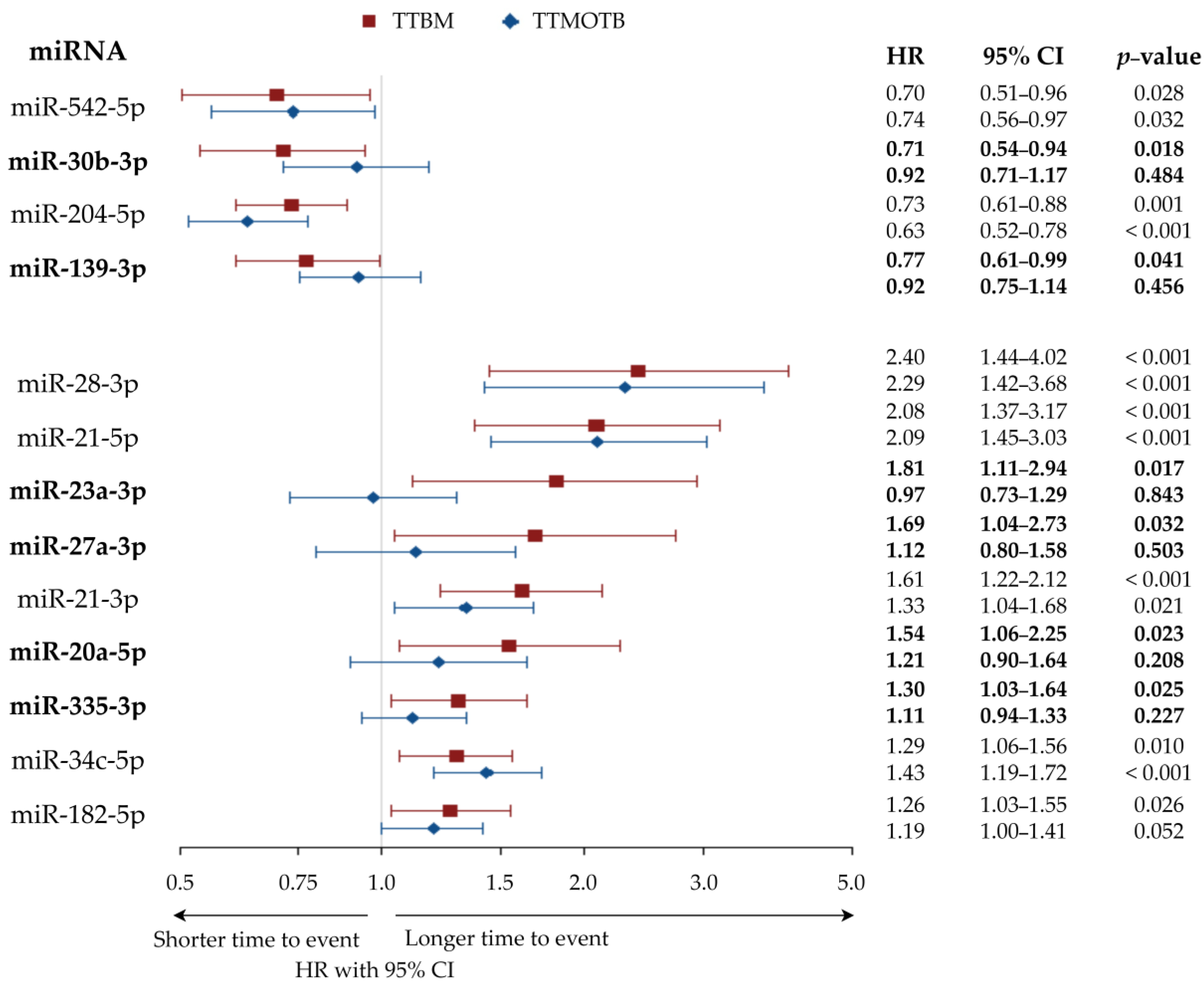
3.2. MiRNAs Associated with Longer or Shorter Time-to-Bone-Metastasis
3.3. Correlation of miRNA with Gene Expression
3.4. Correlation of miRNA with Clinical Outcome
3.5. Correlation of mRNA Expression with TTBM and OS
4. Discussion
4.1. miRNA Associated with Longer TTBM
4.2. miRNA Associated with Shorter TTBM
4.3. miRNA-mRNA Associations
4.4. Limitations and Strengths of Our Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chandrasekar, T.; Klaassen, Z.; Goldberg, H.; Kulkarni, G.S.; Hamilton, R.J.; Fleshner, N.E. Metastatic renal cell carcinoma: Patterns and predictors of metastases—A contemporary population-based series. Urol. Oncol. Semin. Orig. Investig. 2017, 35, 661.e7–661.e14. [Google Scholar] [CrossRef] [PubMed]
- Santini, D.; Procopio, G.; Porta, C.; Ibrahim, T.; Barni, S.; Mazzara, C.; Fontana, A.; Berruti, A.; Berardi, R.; Vincenzi, B.; et al. Natural history of malignant bone disease in renal cancer: Final results of an italian bone metastasis survey. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beuselinck, B.; Oudard, S.; Rixe, O.; Wolter, P.; Blesius, A.; Ayllon, J.; Elaidi, R.; Schöffski, P.; Barrascout, E.; Morel, A.; et al. Negative impact of bone metastasis on outcome in clear-cell renal cell carcinoma treated with sunitinib. Ann. Oncol. 2011, 22, 794–800. [Google Scholar] [CrossRef]
- McKay, R.R.; Kroeger, N.; Xie, W.; Lee, J.L.; Knox, J.J.; Bjarnason, G.A.; MacKenzie, M.J.; Wood, L.; Srinivas, S.; Vaishampayan, U.N.; et al. Impact of bone and liver metastases on patients with renal cell carcinoma treated with targeted therapy. Eur. Urol. 2014, 65, 577–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motzer, R.J.; Escudier, B.; Bukowski, R.; Rini, B.I.; Hutson, T.E.; Barrios, C.H.; Lin, X.; Fly, K.; Matczak, E.; Gore, M.E. Prognostic factors for survival in 1059 patients treated with sunitinib for metastatic renal cell carcinoma. Br. J. Cancer 2013, 108, 2470–2477. [Google Scholar] [CrossRef]
- Beuselinck, B.; Wolter, P.; Karadimou, A.; Elaidi, R.; Dumez, H.; Rogiers, A.; Van Cann, T.; Willems, L.; Body, J.J.; Berkers, J.; et al. Concomitant oral tyrosine kinase inhibitors and bisphosphonates in advanced renal cell carcinoma with bone metastases. Br. J. Cancer 2012, 107, 1665–1671. [Google Scholar] [CrossRef] [Green Version]
- Vrdoljak, E.; Gore, M.; Leyman, S.; Szczylik, C.; Kharkevich, G.; Schöffski, P.; Torday, L.; Mardiak, J.; Zhang, K.; Sajben, P.; et al. Bisphosphonates in patients with renal cell carcinoma and bone metastases: A sunitinib global expanded-access trial subanalysis. Futur. Oncol. 2015, 11, 2831–2840. [Google Scholar] [CrossRef]
- Keizman, D.; Ish-Shalom, M.; Pili, R.; Hammers, H.; Eisenberger, M.A.; Sinibaldi, V.; Boursi, B.; Maimon, N.; Gottfried, M.; Hayat, H.; et al. Bisphosphonates combined with sunitinib may improve the response rate, progression free survival and overall survival of patients with bone metastases from renal cell carcinoma. Eur. J. Cancer 2012, 48, 1031–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beuselinck, B.; Jean-Baptiste, J.; Couchy, G.; Job, S.; De Reynies, A.; Wolter, P.; Théodore, C.; Gravis, G.; Rousseau, B.; Albiges, L.; et al. RANK/OPG ratio of expression in primary clear-cell renal cell carcinoma is associated with bone metastasis and prognosis in patients treated with anti-VEGFR-TKIs. Br. J. Cancer 2015, 113, 1313–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikami, S.; Katsube, K.I.; Oya, M.; Ishida, M.; Kosaka, T.; Mizuno, R.; Mochizuki, S.; Ikeda, T.; Mukai, M.; Okada, Y. Increased RANKL expression is related to tumour migration and metastasis of renal cell carcinomas. J. Pathol. 2009, 218, 530–539. [Google Scholar] [CrossRef]
- Hong, D.S.; Kang, Y.K.; Borad, M.; Sachdev, J.; Ejadi, S.; Lim, H.Y.; Brenner, A.J.; Park, K.; Lee, J.L.; Kim, T.Y.; et al. Phase 1 study of MRX34, a liposomal miR-34a mimic, in patients with advanced solid tumours. Br. J. Cancer 2020, 122, 1630–1637. [Google Scholar] [CrossRef] [PubMed]
- Van Zandwijk, N.; Pavlakis, N.; Kao, S.C.; Linton, A.; Boyer, M.J.; Clarke, S.; Huynh, Y.; Chrzanowska, A.; Fulham, M.J.; Bailey, D.L.; et al. Safety and activity of microRNA-loaded minicells in patients with recurrent malignant pleural mesothelioma: A first-in-man, phase 1, open-label, dose-escalation study. Lancet Oncol. 2017, 18, 1386–1396. [Google Scholar] [CrossRef]
- Reid, G.; Pel, M.E.; Kirschner, M.B.; Cheng, Y.Y.; Mugridge, N.; Weiss, J.; Williams, M.; Wright, C.; Edelman, J.J.B.; Vallely, M.P.; et al. Restoring expression of miR-16: A novel approach to therapy for malignant pleural mesothelioma. Ann. Oncol. 2013, 24, 3128–3135. [Google Scholar] [CrossRef]
- Saliminejad, K.; Khorram Khorshid, H.R.; Soleymani Fard, S.; Ghaffari, S.H. An overview of microRNAs: Biology, functions, therapeutics, and analysis methods. J. Cell. Physiol. 2019, 234, 5451–5465. [Google Scholar] [CrossRef] [PubMed]
- García-Donas, J.; Beuselinck, B.; Inglada-Pérez, L.; Graña, O.; Schöffski, P.; Wozniak, A.; Bechter, O.; Apellániz-Ruiz, M.; Leandro-García, L.J.; Esteban, E.; et al. Deep sequencing reveals microRNAs predictive of antiangiogenic drug response. JCI Insight 2019, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karagkouni, D.; Paraskevopoulou, M.D.; Chatzopoulos, S.; Vlachos, I.S.; Tastsoglou, S.; Kanellos, I.; Papadimitriou, D.; Kavakiotis, I.; Maniou, S.; Skoufos, G.; et al. DIANA-TarBase v8: A decade-long collection of experimentally supported miRNA-gene interactions. Nucleic Acids Res. 2018, 46, D239–D245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Wang, X. MiRDB: An online database for prediction of functional microRNA targets. Nucleic Acids Res. 2020, 48, D127–D131. [Google Scholar] [CrossRef] [Green Version]
- Avendanõ-Félix, M.; Fuentes-Mera, L.; Ramos-Payan, R.; Aguilar-Medina, M.; Pérez-Silos, V.; Moncada-Saucedo, N.; Marchat, L.A.; González-Barrios, J.A.; Ruiz-Garciá, E.; Astudillo-De La Vega, H.; et al. A novel osteomirs expression signature for osteoblast differentiation of human amniotic membrane-derived mesenchymal stem cells. Biomed Res. Int. 2019, 2019. [Google Scholar] [CrossRef] [Green Version]
- Dangi-Garimella, S.; Yun, J.; Eves, E.M.; Newman, M.; Erkeland, S.J.; Hammond, S.M.; Minn, A.J.; Rosner, M.R. Raf kinase inhibitory protein suppresses a metastasis signalling cascade involving LIN28 and let-7. EMBO J. 2009, 28, 347–358. [Google Scholar] [CrossRef] [Green Version]
- Okuda, H.; Xing, F.; Pandey, P.R.; Sharma, S.; Watabe, M.; Pai, S.K.; Mo, Y.Y.; Iiizumi-Gairani, M.; Hirota, S.; Liu, Y.; et al. MiR-7 suppresses brain metastasis of breast cancer stem-like cells by modulating KLF4. Cancer Res. 2013, 73, 1434–1444. [Google Scholar] [CrossRef] [Green Version]
- Zhaoa, F.L.; Hua, G.D.; Wang, X.F.; Zhang, X.H.; Zhang, Y.K.; Yu, Z.S. Serum overexpression of microRNA-10b in patients with bone metastatic primary breast cancer. J. Int. Med. Res. 2012, 40, 859–866. [Google Scholar] [CrossRef]
- Croset, M.; Goehrig, D.; Frackowiak, A.; Bonnelye, E.; Ansieau, S.; Puisieux, A.; Clézardin, P. TWIST1 expression in breast cancer cells facilitates bone metastasis formation. J. Bone Miner. Res. 2014, 29, 1886–1899. [Google Scholar] [CrossRef] [PubMed]
- Vimalraj, S.; Partridge, N.C.; Selvamurugan, N. A positive role of microRNA-15b on regulation of osteoblast differentiation. J. Cell. Physiol. 2014, 229, 1236–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ell, B.; Mercatali, L.; Ibrahim, T.; Campbell, N.; Schwarzenbach, H.; Pantel, K.; Amadori, D.; Kang, Y. Tumor-Induced Osteoclast miRNA Changes as Regulators and Biomarkers of Osteolytic Bone Metastasis. Cancer Cell 2013, 24, 542–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, N.; Wu, D.; Li, H.; Liu, Y.; Yang, H. MiR-17-3p inhibits osteoblast differentiation by downregulating Sox6 expression. FEBS Open Bio. 2020, 10, 2499–2506. [Google Scholar] [CrossRef] [PubMed]
- Vimalraj, S.; Miranda, P.J.; Ramyakrishna, B.; Selvamurugan, N. Regulation of breast cancer and bone metastasis by MicroRNAs. Dis. Markers 2013, 35, 369–387. [Google Scholar] [CrossRef] [Green Version]
- Wotschofsky, Z.; Liep, J.; Meyer, H.A.; Jung, M.; Wagner, I.; Disch, A.C.; Schaser, K.D.; Melcher, I.; Kilic, E.; Busch, J.; et al. Identification of metastamirs as metastasis-associated microRNAs in clear cell renal cell carcinomas. Int. J. Biol. Sci. 2012, 8, 1363–1374. [Google Scholar] [CrossRef]
- Guo, L.; Zhu, Y.; Li, L.; Zhou, S.; Yin, G.; Yu, G.; Cui, H. Breast cancer cell-derived exosomal miR-20a-5p promotes the proliferation and differentiation of osteoclasts by targeting SRCIN1. Cancer Med. 2019, 8, 5687–5701. [Google Scholar] [CrossRef] [Green Version]
- Gong, Y.; Xu, F.; Zhang, L.; Qian, Y.; Chen, J.; Huang, H.; Yu, Y. MicroRNA expression signature for Satb2-induced osteogenic differentiation in bone marrow stromal cells. Mol. Cell. Biochem. 2014, 387, 227–239. [Google Scholar] [CrossRef]
- Xu, Z.; Liu, X.; Wang, H.; Li, J.; Dai, L.; Li, J.; Dong, C. Lung adenocarcinoma cell-derived exosomal miR-21 facilitates osteoclastogenesis. Gene 2018, 666, 116–122. [Google Scholar] [CrossRef]
- Hou, N.; Guo, Z.; Zhao, G.; Jia, G.; Luo, B.; Shen, X.; Bai, Y. Inhibition of microRNA-21-3p suppresses proliferation as well as invasion and induces apoptosis by targeting RNA-binding protein with multiple splicing through Smad4/extra cellular signal-regulated protein kinase signalling pathway in human colorectal can. Clin. Exp. Pharmacol. Physiol. 2018, 45, 729–741. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Zheng, C.; Li, H. Inhibition of miR-23a-3p promotes osteoblast proliferation and differentiation. J. Cell. Biochem. 2019. [Google Scholar] [CrossRef]
- Quan, J.; Pan, X.; Li, Y.; Hu, Y.; Tao, L.; Li, Z.; Zhao, L.; Wang, J.; Li, H.; Lai, Y.; et al. MiR-23a-3p acts as an oncogene and potential prognostic biomarker by targeting PNRC2 in RCC. Biomed. Pharmacother. 2019, 110, 656–666. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.Q.; Gordon, J.A.R.; Beloti, M.M.; Croce, C.M.; Van Wijnen, A.J.; Stein, J.L.; Stein, G.S.; Lian, J.B. A network connecting Runx2, SATB2, and the miR-23a∼27a∼24-2 cluster regulates the osteoblast differentiation program. Proc. Natl. Acad. Sci. USA 2010, 107, 19879–19884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Z.; Wen, Y.; Xuan, C.; Chen, Q.; Xiang, Q.; Wang, J.; Liu, Y.; Luo, L.; Zhao, S.; Deng, Y.; et al. Identifying the key genes and microRNAs in prostate cancer bone metastasis by bioinformatics analysis. FEBS Open Bio. 2020, 10, 674–688. [Google Scholar] [CrossRef] [PubMed]
- Croset, M.; Pantano, F.; Kan, C.W.S.; Bonnelye, E.; Descotes, F.; Alix-Panabieres, C.; Lecellier, C.H.; Bachelier, R.; Allioli, N.; Hong, S.S.; et al. miRNA-30 family members inhibit breast cancer invasion, osteomimicry, and bone destruction by directly targeting multiple bone metastasis–associated genes. Cancer Res. 2018, 78, 5259–5273. [Google Scholar] [CrossRef] [Green Version]
- Kuo, P.L.; Liao, S.H.; Hung, J.Y.; Huang, M.S.; Hsu, Y.L. MicroRNA-33a functions as a bone metastasis suppressor in lung cancer by targeting parathyroid hormone related protein. Biochim. Biophys. Acta Gen. Subj. 2013, 1830, 3756–3766. [Google Scholar] [CrossRef]
- Chen, W.Y.; Liu, S.Y.; Chang, Y.S.; Yin, J.J.; Yeh, H.L.; Mouhieddine, T.H.; Hadadeh, O.; Abou-Kheir, W.; Liu, Y.N. MicroRNA-34a regulates WNT/TCF7 signaling and inhibits bone metastasis in Ras-activated prostate cancer. Oncotarget 2015, 6, 441–457. [Google Scholar] [CrossRef] [Green Version]
- Gu, J.; Wang, G.; Liu, H.; Xiong, C. SATB2 targeted by methylated miR-34c-5p suppresses proliferation and metastasis attenuating the epithelial-mesenchymal transition in colorectal cancer. Cell Prolif. 2018, 51, e12455. [Google Scholar] [CrossRef]
- Clézardin, P. Pathophysiology of bone metastases from solid malignancies. Jt. Bone Spine 2017, 84, 677–684. [Google Scholar] [CrossRef]
- Tang, Y.; Pan, J.; Huang, S.; Peng, X.; Zou, X.; Luo, Y.; Ren, D.; Zhang, X.; Li, R.; He, P.; et al. Downregulation of miR-133a-3p promotes prostate cancer bone metastasis via activating PI3K/AKT signaling. J. Exp. Clin. Cancer Res. 2018, 37, 160. [Google Scholar] [CrossRef]
- Taipaleenmäki, H.; Browne, G.; Akech, J.; Zustin, J.; Van Wijnen, A.J.; Stein, J.L.; Hesse, E.; Stein, G.S.; Lian, J.B. Targeting of Runx2 by miR-135 and miR-203 impairs progression of breast cancer and metastatic bone disease. Cancer Res. 2015, 75, 1433–1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eskildsen, T.; Taipaleenmäki, H.; Stenvang, J.; Abdallah, B.M.; Ditzel, N.; Nossent, A.Y.; Bak, M.; Kauppinen, S.; Kassem, M. MicroRNA-138 regulates osteogenic differentiation of human stromal (mesenchymal) stem cells in vivo. Proc. Natl. Acad. Sci. USA 2011, 108, 6139–6144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, K.; Hu, Z.; Zhou, H.; Zhang, L.; Wang, H.; Li, G.; Zhang, S.; Cao, X.; Shi, F. MicroRNA-139-3p regulates osteoblast differentiation and apoptosis by targeting ELK1 and interacting with long noncoding RNA ODSM. Cell Death Dis. 2018, 9, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Xu, J.; Wang, K.; Tang, X.; He, J. MIR-139-3p suppresses the invasion and migration properties of breast cancer cells by targeting RAB1A. Oncol. Rep. 2019, 42, 1699–1708. [Google Scholar] [CrossRef]
- Xu, S.; Yang, F.; Liu, R.; Li, X.; Fan, H.; Liu, J.; Wei, S.; Chen, G.; Chen, J.; Da, Y. Serum microRNA-139-5p is downregulated in lung cancer patients with lytic bone metastasis. Oncol. Rep. 2018, 39, 2376–2384. [Google Scholar] [CrossRef]
- Ell, B.; Kang, Y. MicroRNAs as regulators of bone homeostasis and bone metastasis. Bonekey Rep. 2014, 3, 549. [Google Scholar] [CrossRef] [Green Version]
- Shabani, P.; Izadpanah, S.; Aghebati-Maleki, A.; Baghbani, E.; Baghbanzadeh, A.; Fotouhi, A.; Bakhshinejad, B.; Aghebati-Maleki, L.; Baradaran, B. Role of miR-142 in the pathogenesis of osteosarcoma and its potential as therapeutic approach. J. Cell. Biochem. 2019, 120, 4783–4793. [Google Scholar] [CrossRef]
- Fordham, J.B.; Guilfoyle, K.; Naqvi, A.R.; Nares, S. MiR-142-3p is a RANKL-dependent inducer of cell death in osteoclasts. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef]
- Peng, X.; Guo, W.; Liu, T.; Wang, X.; Tu, X.; Xiong, D.; Chen, S.; Lai, Y.; Du, H.; Chen, G.; et al. Identification of miRs-143 and -145 that Is Associated with Bone Metastasis of Prostate Cancer and Involved in the Regulation of EMT. PLoS ONE 2011, 6, e20341. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Chen, X.; Deng, W.; Zhong, G.; Cai, Q.; Lin, T. Up-regulated microRNA-143 in cancer stem cells differentiation promotes prostate cancer cells metastasis by modulating FNDC3B expression. BMC Cancer 2013, 13, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Ren, D.; Chen, X.; Tu, X.; Huang, S.; Wang, M.; Song, L.; Zou, X.; Peng, X. HEF1 promotes epithelial mesenchymal transition and bone invasion in prostate cancer under the regulation of microRNA-145. J. Cell. Biochem. 2013, 114, 1606–1615. [Google Scholar] [CrossRef]
- Banerjee, S.; Kalyani Yabalooru, S.R.; Karunagaran, D. Identification of mRNA and non-coding RNA hubs using network analysis in organ tropism regulated triple negative breast cancer metastasis. Comput. Biol. Med. 2020, 127, 104076. [Google Scholar] [CrossRef]
- Eguchi, T.; Watanabe, K.; Hara, E.S.; Ono, M.; Kuboki, T.; Calderwood, S.K. OstemiR: A Novel Panel of MicroRNA Biomarkers in Osteoblastic and Osteocytic Differentiation from Mesencymal Stem Cells. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Ye, Y.; Chang, D.W.; Lin, S.H.; Huang, M.; Tannir, N.M.; Matin, S.; Karam, J.A.; Wood, C.G.; Chen, Z.N.; et al. Global and Targeted miRNA Expression Profiling in Clear Cell Renal Cell Carcinoma Tissues Potentially Links miR-155-5p and miR-210-3p to both Tumorigenesis and Recurrence. Am. J. Pathol. 2018, 188, 2487–2496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.Y.; Des Marais, T.; Costa, M. Deregulation of SATB2 in carcinogenesis with emphasis on miRNA-mediated control. Carcinogenesis 2019, 40, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.M.; Park, S.J.; Jung, S.H.; Kim, E.J.; Jogeswar, G.; Ajita, J.; Rhee, Y.; Kim, C.H.; Lim, S.K. MiR-182 is a negative regulator of osteoblast proliferation, differentiation, and skeletogenesis through targeting FoxO1. J. Bone Miner. Res. 2012, 27, 1669–1679. [Google Scholar] [CrossRef] [PubMed]
- Mock, K.; Preca, B.T.; Brummer, T.; Brabletz, S.; Stemmler, M.P.; Brabletz, T. The EMT-activator ZEB1 induces bone metastasis associated genes including BMP-inhibitors. Oncotarget 2015, 6, 14399–14412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollari, S.; Leivonen, S.K.; Perälä, M.; Fey, V.; Käkönen, S.M.; Kallioniemi, O. Identification of microRNAs inhibiting TGF-β-induced IL-11 production in bone metastatic breast cancer cells. PLoS ONE 2012, 7, e37361. [Google Scholar] [CrossRef] [Green Version]
- Ren, D.; Yang, Q.; Dai, Y.; Guo, W.; Du, H.; Song, L.; Peng, X. Oncogenic miR-210-3p promotes prostate cancer cell EMT and bone metastasis via NF-ΚB signaling pathway. Mol. Cancer 2017, 16, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Li, D.; Dang, L.; Liang, C.; Guo, B.; Lu, C.; He, X.; Cheung, H.Y.S.; He, B.; Liu, B.; et al. Osteoclastic miR-214 targets TRAF3 to contribute to osteolytic bone metastasis of breast cancer. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.Q.; Maeda, Y.; Taipaleenmaki, H.; Zhang, W.; Jafferji, M.; Gordon, J.A.R.; Li, Z.; Croce, C.M.; Van Wijnen, A.J.; Stein, J.L.; et al. miR-218 directs a Wnt signaling circuit to promote differentiation of osteoblasts and osteomimicry of metastatic cancer cells. J. Biol. Chem. 2012, 287, 42084–42092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valencia, K.; Martín-Fernández, M.; Zandueta, C.; Ormazábal, C.; Martínez-Canarias, S.; Bandrés, E.; de la Piedra, C.; Lecanda, F. MiR-326 associates with biochemical markers of bone turnover in lung cancer bone metastasis. Bone 2013, 52, 532–539. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Tang, Y.; Zhu, X.; Tu, T.; Sui, L.; Han, Q.; Yu, L.; Meng, S.; Zheng, L.; Valverde, P.; et al. Overexpression of MiR-335-5p Promotes Bone Formation and Regeneration in Mice. J. Bone Miner. Res. 2017, 32, 2466–2475. [Google Scholar] [CrossRef]
- Gururajan, M.; Josson, S.; Chu, G.C.Y.; Lu, C.L.; Lu, Y.T.; Haga, C.L.; Zhau, H.E.; Liu, C.; Lichterman, J.; Duan, P.; et al. MiR-154∗ and miR-379 in the DLK1-DIO3 MicroRNA mega-cluster regulate epithelial to mesenchymal transition and bone metastasis of prostate cancer. Clin. Cancer Res. 2014, 20, 6559–6569. [Google Scholar] [CrossRef] [Green Version]
- Kureel, J.; Dixit, M.; Tyagi, A.M.; Mansoori, M.N.; Srivastava, K.; Raghuvanshi, A.; Maurya, R.; Trivedi, R.; Goel, A.; Singh, D. MiR-542-3p suppresses osteoblast cell proliferation and differentiation, targets BMP-7 signaling and inhibits bone formation. Cell Death Dis. 2014, 5, e1050. [Google Scholar] [CrossRef]
- He, R.Q.; Li, X.J.; Liang, L.; Xie, Y.; Luo, D.Z.; Ma, J.; Peng, Z.G.; Hu, X.H.; Chen, G. The suppressive role of miR-542-5p in NSCLC: The evidence from clinical data and in vivo validation using a chick chorioallantoic membrane model. BMC Cancer 2017, 17, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Fu, Q.; Liu, X.; Liu, Y.; Yang, J.; Lv, G.; Dong, S. MicroRNA-335 and -543 suppress bone metastasis in prostate cancer via targeting endothelial nitric oxide synthase. Int. J. Mol. Med. 2015, 36, 1417–1425. [Google Scholar] [CrossRef] [Green Version]
- Sosa, M.S.; Bragado, P.; Aguirre-Ghiso, J.A. Mechanisms of disseminated cancer cell dormancy: An awakening field. Nat. Rev. Cancer 2014, 14, 611–622. [Google Scholar] [CrossRef]
- Kobayashi, A.; Okuda, H.; Xing, F.; Pandey, P.R.; Watabe, M.; Hirota, S.; Pai, S.K.; Liu, W.; Fukuda, K.; Chambers, C.; et al. Bone morphogenetic protein 7 in dormancy and metastasis of prostate cancer stem-like cells in bone. J. Exp. Med. 2011, 208, 2641–2655. [Google Scholar] [CrossRef] [Green Version]
- Sugatani, T.; Vacher, J.; Hruska, K.A. A microRNA expression signature of osteoclastogenesis. Blood 2011, 117, 3648–3657. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Xiong, D.; Yao, X.; Gu, W.; Zhang, H.; Yang, B.; Peng, B.; Liu, M.; Zheng, J. Decreased SATB2 expression is associated with metastasis and poor prognosis in human clear cell renal cell carcinoma. Int. J. Clin. Exp. Pathol. 2015, 8, 3710–3718. [Google Scholar] [PubMed]
- Petersen, M.; Pardali, E.; Van Der Horst, G.; Cheung, H.; Van Den Hoogen, C.; Van Der Pluijm, G.; Ten Dijke, P. Smad2 and Smad3 have opposing roles in breast cancer bone metastasis by differentially affecting tumor angiogenesis. Oncogene 2010, 29, 1351–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deckers, M.; Van Dinther, M.; Buijs, J.; Que, I.; Löwik, C.; Van Der Pluijm, G.; Ten Dijke, P. The tumor suppressor Smad4 is required for transforming growth factor β-induced epithelial to mesenchymal transition and bone metastasis of breast cancer cells. Cancer Res. 2006, 66, 2202–2209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, X.; Gong, K.; Zhang, X.; Wu, S.; Cui, Y.; Qian, B.Z. Osteopontin as a multifaceted driver of bone metastasis and drug resistance. Pharmacol. Res. 2019, 144, 235–244. [Google Scholar] [CrossRef]
- Morein, D.; Erlichman, N.; Ben-Baruch, A. Beyond Cell Motility: The Expanding Roles of Chemokines and Their Receptors in Malignancy. Front. Immunol. 2020, 11, 952. [Google Scholar] [CrossRef]
- Zlotnik, A.; Burkhardt, A.M.; Homey, B. Homeostatic chemokine receptors and organ-specific metastasis. Nat. Rev. Immunol. 2011, 11, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.L.; Hou, M.F.; Kuo, P.L.; Huang, Y.F.; Tsai, E.M. Breast tumor-associated osteoblast-derived CXCL5 increases cancer progression by ERK/MSK1/Elk-1/Snail signaling pathway. Oncogene 2013, 32, 4436–4447. [Google Scholar] [CrossRef] [Green Version]
- Li, X.Q.; Lu, J.T.; Tan, C.C.; Wang, Q.S.; Feng, Y.M. RUNX2 promotes breast cancer bone metastasis by increasing integrin α5-mediated colonization. Cancer Lett. 2016, 380, 78–86. [Google Scholar] [CrossRef]
- Pécheur, I.; Peyruchaud, O.; Serre, C.M.; Guglielmi, J.; Voland, C.; Bourre, F.; Margue, C.; Cohen-Solal, M.; Buffet, A.; Kieffer, N.; et al. Integrin alpha(v)beta3 expression confers on tumor cells a greater propensity to metastasize to bone. FASEB J. 2002, 16, 1266–1268. [Google Scholar] [CrossRef] [PubMed]
- Katsuno, Y.; Hanyu, A.; Kanda, H.; Ishikawa, Y.; Akiyama, F.; Iwase, T.; Ogata, E.; Ehata, S.; Miyazono, K.; Imamura, T. Bone morphogenetic protein signaling enhances invasion and bone metastasis of breast cancer cells through Smad pathway. Oncogene 2008, 27, 6322–6333. [Google Scholar] [CrossRef] [Green Version]
- Croset, M.; Kan, C.; Clézardin, P. Tumour-derived miRNAs and bone metastasis. Bonekey Rep. 2015, 4. [Google Scholar] [CrossRef] [Green Version]
- Deligiorgi, M.V.; Panayiotidis, M.I.; Griniatsos, J.; Trafalis, D.T. Harnessing the versatile role of OPG in bone oncology: Counterbalancing RANKL and TRAIL signaling and beyond. Clin. Exp. Metastasis 2020, 37, 13–30. [Google Scholar] [CrossRef]
- Heng, D.Y.C.; Xie, W.; Regan, M.M.; Warren, M.A.; Golshayan, A.R.; Sahi, C.; Eigl, B.J.; Ruether, J.D.; Cheng, T.; North, S.; et al. Prognostic factors for overall survival in patients with metastatic renal cell carcinoma treated with vascular endothelial growth factor-targeted agents: Results from a large, multicenter study. J. Clin. Oncol. 2009, 27, 5794–5799. [Google Scholar] [CrossRef]
- Huang, J.; Zhao, L.; Xing, L.; Chen, D. MicroRNA-204 regulates Runx2 protein expression and mesenchymal progenitor cell differentiation. Stem Cells 2010, 28, 357–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Liu, T.; Sun, L.; Zhang, S.; Dong, G. Long noncoding RNA SNHG4 promotes renal cell carcinoma tumorigenesis and invasion by acting as ceRNA to sponge miR-204-5p and upregulate RUNX2. Cancer Cell Int. 2020, 20. [Google Scholar] [CrossRef]
- Shuai, F.; Wang, B.; Dong, S. MicroRNA-204 inhibits the growth and motility of colorectal cancer cells by downregulation of CXCL8. Oncol. Res. 2018, 26, 1295–1305. [Google Scholar] [CrossRef] [PubMed]
- Garros, R.F.; Paul, R.; Connolly, M.; Lewis, A.; Garfield, B.E.; Natanek, S.A.; Bloch, S.; Mouly, V.; Griffiths, M.J.; Polkey, M.I.; et al. MicroRNA-542 promotes mitochondrial dysfunction and SMAD activity and is elevated in intensive care unit–acquired weakness. Am. J. Respir. Crit. Care Med. 2017, 196, 1422–1433. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Sui, B.D.; Hu, C.H.; Cao, J.; Zheng, C.X.; Hou, R.; Yang, Z.K.; Zhao, P.; Chen, Q.; Yang, Q.J.; et al. MicroRNA-21 contributes to orthodontic tooth movement. J. Dent. Res. 2016, 95, 1425–1433. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.; Zhao, B.; Shi, Y.; Yao, C.; Jin, L.; Jin, Y. BMPRII is a direct target of miR-21. Acta Biochim. Biophys. Sin. 2009, 41, 618–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loboda, A.; Sobczak, M.; Jozkowicz, A.; Dulak, J. TGF-β 1/Smads and miR-21 in Renal Fibrosis and Inflammation. Mediators Inflamm. 2016, 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.; Shi, Y.; Zheng, L.; Zhou, B.; Inose, H.; Wang, J.; Guo, X.E.; Grosschedl, R.; Karsenty, G. miR-34s inhibit osteoblast proliferation and differentiation in the mouse by targeting SATB2. J. Cell Biol. 2012, 197, 509–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.F.; Fu, W.M.; He, M.L.; Xie, W.D.; Lv, Q.; Wan, G.; Li, G.; Wang, H.; Lu, G.; Hu, X.; et al. MiRNA-20a promotes osteogenic differentiation of human mesenchymal stem cells by co-regulating BMP signaling. RNA Biol. 2011, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.H.; Yu, J.; Jiang, D.M.; Li, W.L.; Wang, S.; Ding, Y.Q. MicroRNA-182 targets special AT-rich sequence-binding protein 2 to promote colorectal cancer proliferation and metastasis. J. Transl. Med. 2014, 12, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Choueiri, T.K.; Garcia, J.A.; Elson, P.; Khasawneh, M.; Usman, S.; Golshayan, A.R.; Baz, R.C.; Wood, L.; Rini, B.I.; Bukowski, R.M. Clinical factors associated with outcome in patients with metastatic clear-cell renal cell carcinoma treated with vascular endothelial growth factor-targeted therapy. Cancer 2007, 110, 543–550. [Google Scholar] [CrossRef]
- Santoni, M.; Conti, A.; Procopio, G.; Porta, C.; Ibrahim, T.; Barni, S.; Guida, F.M.; Fontana, A.; Berruti, A.; Berardi, R.; et al. Bone metastases in patients with metastatic renal cell carcinoma: Are they always associated with poor prognosis? J. Exp. Clin. Cancer Res. 2015, 34, 10. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Zhou, H.; Hong, Y.; Li, J.; Jiang, X.; Huang, H. miR-30 family members negatively regulate osteoblast differentiation. J. Biol. Chem. 2012, 287, 7503–7511. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Li, G.; Wang, K.; Wang, Y.; Dong, J.; Wang, H.; Xu, L.; Shi, F.; Cao, X.; Hu, Z.; et al. MiR-30 family members inhibit osteoblast differentiation by suppressing Runx2 under unloading conditions in MC3T3-E1 cells. Biochem. Biophys. Res. Commun. 2020, 522, 164–170. [Google Scholar] [CrossRef]
- Yi, J.; Liu, D.; Xiao, J. LncRNA MALAT1 sponges miR-30 to promote osteoblast differentiation of adipose-derived mesenchymal stem cells by promotion of Runx2 expression. Cell Tissue Res. 2019, 376, 113–121. [Google Scholar] [CrossRef]
- Yu, F.; Deng, H.; Yao, H.; Liu, Q.; Su, F.; Song, E. Mir-30 reduction maintains self-renewal and inhibits apoptosis in breast tumor-initiating cells. Oncogene 2010, 29, 4194–4204. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.J.; Lin, J.; Zhu, D.; Wang, X.; Brooks, D.; Chen, M.; Chu, Z.B.; Takada, K.; Ciccarelli, B.; Admin, S.; et al. MiR-30-5p functions as a tumor suppressor and novel therapeutic tool by targeting the oncogenic Wnt/ b-Catenin/BCL9 pathway. Cancer Res. 2014, 74, 1801–1813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, I.; Tivnan, A.; Bryan, K.; Foley, N.H.; Watters, K.M.; Tracey, L.; Davidoff, A.M.; Stallings, R.L. MicroRNA-542-5p as a Novel Tumor Suppressor in Neuroblastoma. Cancer Lett. 2011, 303, 56–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Song, S.; Ni, G.; Li, Y.; Wang, X. Serum miR-542-3p as a prognostic biomarker in osteosarcoma. Cancer Biomark. 2018, 21, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Yuan, P.; Yuan, H.; Wang, Z.; Run, Z.; Chen, G.; Zhao, P.; Xu, B. miR-542-3p inhibits colorectal cancer cell proliferation, migration and invasion by targeting OTUB1. Am. J. Cancer Res. 2017, 7, 159–172. [Google Scholar]
- Wu, H.X.; Wang, G.M.; Lu, X.; Zhang, L. miR-542-3p targets sphingosine-1-phosphate receptor 1 and regulates cell proliferation and invasion of breast cancer cells. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 108–114. [Google Scholar]
- Tao, J.; Liu, Z.; Wang, Y.; Wang, L.; Yao, B.; Li, Q.; Wang, C.; Tu, K.; Liu, Q. MiR-542-3p inhibits metastasis and epithelial-mesenchymal transition of hepatocellular carcinoma by targeting UBE3C. Biomed. Pharmacother. 2017, 93, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.D.; Yu, T.; Hu, T.; Yao, M.; Fan, C.Y.; Yang, Q.C. MiR-542-5p is a negative prognostic factor and promotes osteosarcoma tumorigenesis by targeting HUWE1. Oncotarget 2015, 6, 42761–42772. [Google Scholar] [CrossRef] [Green Version]
- Okada, R.; Goto, Y.; Yamada, Y.; Kato, M.; Asai, S.; Moriya, S.; Ichikawa, T.; Seki, N. Regulation of oncogenic targets by the tumor-suppressive mir-139 duplex (Mir-139-5p and mir-139-3p) in renal cell carcinoma. Biomedicines 2020, 8, 599. [Google Scholar] [CrossRef] [PubMed]
- Yonemori, M.; Seki, N.; Yoshino, H.; Matsushita, R.; Miyamoto, K.; Nakagawa, M.; Enokida, H. Dual tumor-suppressors miR-139-5p and miR-139-3p targeting matrix metalloprotease 11 in bladder cancer. Cancer Sci. 2016, 107, 1233–1242. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Dong, Y.; Zhu, N.; Tsoi, H.; Zhao, Z.; Wu, C.W.; Wang, K.; Zheng, S.; Ng, S.S.M.; Chan, F.K.L.; et al. MicroRNA-139-5p exerts tumor suppressor function by targeting NOTCH1 in colorectal cancer. Mol. Cancer 2014, 13. [Google Scholar] [CrossRef] [Green Version]
- Dai, S.; Wang, X.; Li, X.; Cao, Y. MicroRNA-139-5p acts as a tumor suppressor by targeting ELTD1 and regulating cell cycle in glioblastoma multiforme. Biochem. Biophys. Res. Commun. 2015, 467, 204–210. [Google Scholar] [CrossRef]
- Zou, Z.C.; Dai, M.; Huang, Z.Y.; Lu, Y.; Xie, H.P.; Li, Y.F.; Li, Y.; Tan, Y.; Wang, F.L. MicroRNA-139-3p suppresses tumor growth and metastasis in hepatocellular carcinoma by repressing ANXA2R. Oncol. Res. 2018, 26, 1391–1399. [Google Scholar] [CrossRef]
- Wang, Y.; Fang, Q.; Tian, L.; Yuan, Z.; Tian, L.; Zhou, Z. Expression and Regulatory Network Analysis of MiR-139-3p, a New Potential Serum Biomarker for Esophageal Squamous Cell Carcinoma Based on Bioinformatics Analysis. Technol. Cancer Res. Treat. 2020, 19. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, M.; Liu, J.; Li, X.; Yang, M.; Su, B.; Lin, Y. 27-Hydroxycholesterol enhanced osteoclastogenesis in lung adenocarcinoma microenvironment. J. Cell. Physiol. 2019, 234, 12692–12700. [Google Scholar] [CrossRef]
- Jedroszka, D.; Orzechowska, M.; Bednarek, A.K. Predictive values of Notch signalling in renal carcinoma. Arch. Med. Sci. 2017, 13, 1249–1254. [Google Scholar] [CrossRef] [Green Version]
- Shu, X.; Hildebrandt, M.A.; Gu, J.; Tannir, N.M.; Matin, S.F.; Karam, J.A.; Wood, C.G.; Wu, X. MicroRNA profiling in clear cell renal cell carcinoma tissues potentially links tumorigenesis and recurrence with obesity. Br. J. Cancer 2017, 116, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Osanto, S.; Qin, Y.; Buermans, H.P.; Berkers, J.; Lerut, E.; Goeman, J.J.; van Poppel, H. Genome-wide microRNA expression analysis of clear cell renal cell carcinoma by next generation deep sequencing. PLoS ONE 2012, 7, 38298. [Google Scholar] [CrossRef] [Green Version]
- Jung, M.; Mollenkopf, H.J.; Grimm, C.; Wagner, I.; Albrecht, M.; Waller, T.; Pilarsky, C.; Johannsen, M.; Stephan, C.; Lehrach, H.; et al. MicroRNA profiling of clear cell renal cell cancer identifies a robust signature to define renal malignancy. J. Cell. Mol. Med. 2009, 13, 3918–3928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verbiest, A.; van Hoef, V.; Rodriguez-Antona, C.; García-Donas, J.; Graña-Castro, O.; Albersen, M.; Baldewijns, M.; Laenen, A.; Roussel, E.; Schöffski, P.; et al. MicroRNA expression profiles in molecular subtypes of clear-cell renal cell carcinoma are associated with clinical outcome and repression of specific mRNA targets. PLoS ONE 2020, 15. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Wu, Y.; He, X.; Zhang, C.; Zhu, M.; Chen, B.; Liu, Q.; Qu, X.; Li, W.; Wen, S.; et al. MicroRNA-204-5p inhibits invasion and metastasis of laryngeal squamous cell carcinoma by suppressing forkhead box C1. J. Cancer 2017, 8, 2356–2368. [Google Scholar] [CrossRef] [PubMed]
- Hong, B.S.; Ryu, H.S.; Kim, N.; Kim, J.; Lee, E.; Moon, H.; Kim, K.H.; Jin, M.S.; Kwon, N.H.; Kim, S.; et al. Tumor suppressor miRNA-204-5p regulates growth, metastasis, and immune microenvironment remodeling in breast cancer. Cancer Res. 2019, 79, 1520–1534. [Google Scholar] [CrossRef]
- Wang, X.; Li, F.; Zhou, X. miR-204-5p regulates cell proliferation and metastasis through inhibiting CXCR4 expression in OSCC. Biomed. Pharmacother. 2016, 82, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Zhang, Z.; Peng, T.; Wang, G.; Xu, Q.; Li, G. MiR-204 inhibits the osteogenic differentiation of mesenchymal stem cells by targeting bone morphogenetic protein 2. Mol. Med. Rep. 2020, 21, 43–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, L.; Xu, G.; Zhu, Y.; Wang, Y. Expression of miR-23a and miR-135 and tumor markers in gastric cancer patients and the significance in diagnosis. Oncol. Lett. 2019, 18, 5853–5858. [Google Scholar] [CrossRef] [Green Version]
- Karimi, N.; Ali Hosseinpour Feizi, M.; Safaralizadeh, R.; Hashemzadeh, S.; Baradaran, B.; Shokouhi, B.; Teimourian, S. Serum overexpression of miR-301a and miR-23a in patients with colorectal cancer. J. Chinese Med. Assoc. 2019, 82, 215–220. [Google Scholar] [CrossRef]
- Wu, Q.; Lu, Z.; Li, H.; Lu, J.; Guo, L.; Ge, Q. Next-generation sequencing of microRNAs for breast cancer detection. J. Biomed. Biotechnol. 2011, 2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Liu, X.; Xu, W.; Zhou, P.; Gao, P.; Jiang, S.; Lobie, P.E.; Zhu, T. c-MYC-regulated miR-23a/24-2/27a Cluster Promotes Mammary Carcinoma Cell Invasion and Hepatic Metastasis by Targeting Sprouty2 * and the. J. Biol. Chem. 2013. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Li, H.; Wang, Y.; Li, T.; Fan, J.; Xiao, K.; Zhao, R.C.; Weng, X. MicroRNA-23a inhibits osteogenic differentiation of human bone marrow-derived mesenchymal stem cells by targeting LRP5. Int. J. Biochem. Cell Biol. 2016, 72, 55–62. [Google Scholar] [CrossRef]
- Moody, L.; Dvoretskiy, S.; An, R.; Mantha, S.; Pan, Y.-X. The Efficacy of miR-20a as a Diagnostic and Prognostic Biomarker for Colorectal Cancer: A Systematic Review and Meta-Analysis. Cancers 2019, 11, 1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, R.; Fu, Y.; Zeng, Y.; Xiang, M.; Yin, Y.; Li, L.; Xu, H.; Zhong, J.; Zeng, X. Serum miR-20a is a promising biomarker for gastric cancer. Biomed. Rep. 2017, 6, 429–434. [Google Scholar] [CrossRef] [Green Version]
- Jung, K.; Lein, M.; Stephan, C.; Von Hösslin, K.; Semjonow, A.; Sinha, P.; Loening, S.A.; Schnorr, D. Comparison of 10 serum bone turnover markers in prostate carcinoma patients with bone metastatic spread: Diagnostic and prognostic implications. Int. J. Cancer 2004, 111, 783–791. [Google Scholar] [CrossRef]
- Zhong, X.L.; Yan, X.; Yang, X.K.; Xiu, H.; Zhao, M.; Wang, X.N.; Liu, J.X. MiR-20a acted as a ceRNA of lncRNA PTENPL and promoted bladder cancer cell proliferation and migration by regulating PDCD4. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 2955–2964. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Li, L.; Hou, Z.; Liu, W.; Wang, H.; Zhou, T.; Li, Y.; Chen, S. LncRNA HAND2-AS1 inhibits 5-fluorouracil resistance by modulating miR-20a/PDCD4 axis in colorectal cancer. Cell. Signal. 2020, 66, 109483. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Liu, T.; Bao, Y.; Zhao, T.; Wang, J.; Wang, H.; Wang, A.; Gan, X.; Wu, Z.; Wang, L. CircRNA cRAPGEF5 inhibits the growth and metastasis of renal cell carcinoma via the miR-27a-3p/TXNIP pathway. Cancer Lett. 2020, 469, 68–77. [Google Scholar] [CrossRef]
- Song, E.L.; Xing, L.; Wang, L.; Song, W.T.; Li, D.B.; Wang, Y.; Gu, Y.W.; Liu, M.M.; Ni, W.J.; Zhang, P.; et al. LncRNA ADAMTS9-AS2 inhibits cell proliferation and decreases chemoresistance in clear cell renal cell carcinoma via the miR-27a-3p/FOXO1 axis. Aging 2019, 11, 5705–5725. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, M.; Liu, X.; Liu, F.; Zhu, J. MiR-27a-3p promotes the malignant phenotypes of osteosarcoma by targeting ten-eleven translocation 1. Int. J. Oncol. 2018, 52, 1295–1304. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.; Tang, J.; Shi, H.; Li, H.; Zhen, T.; Duan, J.; Kang, L.; Zhang, F.; Dong, Y.; Han, A. miR-27a-3p targeting RXRa promotes colorectal cancer progression by activating Wnt/β-catenin pathway. Oncotarget 2017, 8, 82991–83008. [Google Scholar] [CrossRef] [Green Version]
- Su, C.; Huang, D.P.; Liu, J.W.; Liu, W.Y.; Cao, Y.O. miR-27a-3p regulates proliferation and apoptosis of colon cancer cells by potentially targeting BTG1. Oncol. Lett. 2019, 18, 2825–2834. [Google Scholar] [CrossRef] [Green Version]
- Rao, X.; Wan, L.; Jie, Z.; Zhu, X.; Yin, J.; Cao, H. Upregulated miR-27a-3p indicates a poor prognosis in pancreatic carcinoma patients and promotes the angiogenesis and migration by epigenetic silencing of GATA6 and activating VEGFA/VEGFR2 signaling pathway. Onco. Targets. Ther. 2019, 12, 11241–11254. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Liang, X.; Zhang, L.; Yang, L.; Nagao, N.; Wu, H.; Liu, C.; Lin, S.; Cai, G.; Liu, J. MiR-27a-3p functions as an oncogene in gastric cancer by targeting BTG2. Oncotarget 2016, 7, 51943–51954. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.; Yu, H.; Liu, Y.; Hou, J.; Yang, Q.; Zhao, Y. miR-27a-3p Functions as a Tumor Suppressor and Regulates Non-Small Cell Lung Cancer Cell Proliferation via Targeting HOXB8. Technol. Cancer Res. Treat. 2019, 18. [Google Scholar] [CrossRef] [Green Version]
- Zhao, N.; Sun, H.; Sun, B.; Zhu, D.; Zhao, X.; Wang, Y.; Gu, Q.; Dong, X.; Liu, F.; Zhang, Y.; et al. miR-27a-3p suppresses tumor metastasis and VM by down-regulating VE-cadherin expression and inhibiting EMT: An essential role for Twist-1 in HCC. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Liu, X.; Pan, B.; Sun, L.; Chen, X.; Zeng, K.; Hu, X.; Xu, T.; Xu, M.; Wang, S. Circulating exosomal miR-27a and miR-130a act as novel diagnostic and prognostic biomarkers of colorectal cancer. Cancer Epidemiol. Biomarkers Prev. 2018, 27, 746–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.S.; Liu, L.X.; Li, G.P.; Chen, Y.; Li, C.Y.; Jin, D.Y.; Wang, X.L. Combined serum CA19-9 and miR-27a-3p in peripheral blood mononuclear cells to diagnose pancreatic cancer. Cancer Prev. Res. 2013, 6, 331–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, W.; Hong, Z.; Huang, H.; Zhu, A.; Lin, S.; Cheng, C.; Zhang, X.; Zou, G.; Shi, Z. miR-27a in serum acts as biomarker for prostate cancer detection and promotes cell proliferation by targeting Sprouty2. Oncol. Lett. 2018, 16, 5291–5298. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Li, D.; Zhu, Z.; Li, L.; Jin, Y.; Ma, C.; Zhang, W. miR-27a-3p negatively regulates osteogenic differentiation of MC3T3-E1 preosteoblasts by targeting osterix. Mol. Med. Rep. 2020, 22, 1717–1726. [Google Scholar] [CrossRef]
- Li, X.; Wu, X. MiR-21-5p promotes the progression of non-small-cell lung cancer by regulating the expression of SMAD7. Onco. Targets. Ther. 2018, 11, 8445–8454. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Li, B.; Li, Q.; Wei, S.; He, Z.; Huang, X.; Wang, L.; Xia, Y.; Xu, Z.; Li, Z.; et al. Exosomal miR-21-5p derived from gastric cancer promotes peritoneal metastasis via mesothelial-to-mesenchymal transition. Cell Death Dis. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Liu, H.; Zou, H.; Chen, R.; Dou, Y.; Sheng, S.; Dai, S.; Ai, J.; Melson, J.; Kittles, R.A.; et al. Evaluation of plasma miR-21 and miR-152 as diagnostic biomarkers for common types of human cancers. J. Cancer 2016, 7, 490–499. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, T.; Chen, C.; Wu, Z.; Bai, P.; Li, S.; Chen, B.; Liu, R.; Zhang, K.; Li, W.; et al. Serum exosomal miR-210 as a potential biomarker for clear cell renal cell carcinoma. J. Cell. Biochem. 2019, 120, 1492–1502. [Google Scholar] [CrossRef] [PubMed]
- Faragalla, H.; Youssef, Y.M.; Scorilas, A.; Khalil, B.; White, N.M.A.; Mejia-Guerrero, S.; Khella, H.; Jewett, M.A.S.; Evans, A.; Lichner, Z.; et al. The clinical utility of miR-21 as a diagnostic and prognostic marker for renal cell carcinoma. J. Mol. Diagn. 2012, 14, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Pitari, M.R.; Rossi, M.; Amodio, N.; Botta, C.; Morelli, E.; Federico, C.; Gullà, A.; Caracciolo, D.; Di Martino, M.T.; Arbitrio, M.; et al. Inhibition of miR-21 restores RANKL/OPG ratio in multiple myeloma-derived bone marrow stromal cells and impairs the resorbing activity of mature osteoclasts. Oncotarget 2015, 6, 27343–27358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Huang, B.; Zhang, Q.; He, X.; Wei, H.; Zhang, D. NOTCH1 regulates the proliferation and migration of bladder cancer cells by cooperating with long non-coding RNA HCG18 and microRNA-34c-5p. J. Cell. Biochem. 2019, 120, 6596–6604. [Google Scholar] [CrossRef] [PubMed]
- Daugaard, I.; Knudsen, A.; Kjeldsen, T.E.; Hager, H.; Hansen, L.L. The association between miR-34 dysregulation and distant metastases formation in lung adenocarcinoma. Exp. Mol. Pathol. 2017, 102, 484–491. [Google Scholar] [CrossRef]
- Re, M.; Magliulo, G.; Gioacchini, F.M.; Bajraktari, A.; Bertini, A.; Çeka, A.; Rubini, C.; Ferrante, L.; Procopio, A.D.; Olivieri, F. Expression Levels and Clinical Significance of miR-21-5p, miR-let-7a, and miR-34c-5p in Laryngeal Squamous Cell Carcinoma. Biomed Res. Int. 2017, 2017. [Google Scholar] [CrossRef] [Green Version]
- White, N.M.A.; Bao, T.T.; Grigull, J.; Youssef, Y.M.; Girgis, A.; Diamandis, M.; Fatoohi, E.; Metias, M.; Honey, R.J.; Stewart, R.; et al. miRNA Profiling for Clear Cell Renal Cell Carcinoma: Biomarker Discovery and Identification of Potential Controls and Consequences of miRNA Dysregulation. J. Urol. 2011, 186, 1077–1083. [Google Scholar] [CrossRef]
- Wang, K.; Chen, X.; Zhan, Y.; Jiang, W.; Liu, X.; Wang, X.; Wu, B. miR-335 inhibits the proliferation and invasion of clear cell renal cell carcinoma cells through direct suppression of BCL-W. Tumor Biol. 2015, 36, 6875–6882. [Google Scholar] [CrossRef]
- Qin, S.; Shi, X.; Wang, C.; Jin, P.; Ma, F. Transcription factor and miRNA interplays can manifest the survival of ccRCC patients. Cancers 2019, 11. [Google Scholar] [CrossRef] [Green Version]
- Shu, M.; Zheng, X.; Wu, S.; Lu, H.; Leng, T.; Zhu, W.; Zhou, Y.; Ou, Y.; Lin, X.; Lin, Y.; et al. Targeting oncogenic miR-335 inhibits growth and invasion of malignant astrocytoma cells. Mol. Cancer 2011, 10, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavazoie, S.F.; Alarcón, C.; Oskarsson, T.; Padua, D.; Wang, Q.; Bos, P.D.; Gerald, W.L.; Massagué, J. Endogenous human microRNAs that suppress breast cancer metastasis. Nature 2008, 451, 147–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, M.; Ma, J.; Guillemette, R.; Zhou, M.; Yang, Y.; Yang, Y.; Hock, J.M.; Yu, X. MiR-335 inhibits small cell lung cancer bone metastases via IGF-IR and RANKL pathways. Mol. Cancer Res. 2014, 12, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Wu, J.; Li, S.; Hu, Z.; Xu, X.; Zhu, Y.; Liang, Z.; Wang, X.; Lin, Y.; Mao, Y.; et al. Downregulation of microRNA-182-5p contributes to renal cell carcinoma proliferation via activating the AKT/FOXO3a signaling pathway. Mol. Cancer 2014, 13, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Li, H.; Cui, L.; Feng, J.; Fan, Q. MicroRNA-182 suppresses clear cell renal cell carcinoma migration and invasion by targeting IGF1R. Neoplasma 2016, 63, 717–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, J.H.; Yang, S.; Nan, C.J.; Zhou, C.C.; Lu, D.Q.; Li, S.; Mu, H.Q. miR-182 affects renal cancer cell proliferation, apoptosis, and invasion by regulating PI3K/ AKT/mTOR signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Deng, Z.; Chen, Y.; Giannopoulou, E.; Xu, R.; Gong, S.; Greenblatt, M.B.; Mangala, L.S.; Lopez-Berestein, G.; Kirsch, D.G.; et al. Bone protection by inhibition of microRNA-182. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef]
- Sliwinska-Jewsiewicka, A.; Kowalczyk, A.E.; Krazinski, B.E.; Godlewski, J.; Kwiatkowski, P.; Kiewisz, J.; Grzegrzolka, J.; Dziegiel, P.; Kmiec, Z. Decreased expression of SATB2 associates with tumor growth and predicts worse outcome in patients with clear cell renal cell carcinoma. Anticancer Res. 2018, 38, 839–846. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Selected miRNAs | ||
|---|---|---|
| let-7a-3p [18,19], let-7a-5p [19,20], let-7g-3p [19], let-7g-5p [19], miR-10b-3p [21,22], miR-10b-5p [21,22], miR-15b-3p [23], miR-15b-5p [23], miR-16-1-3p [24], miR-16-2-3p [24], miR-16-5p [24], miR-17-3p [25], miR-17-5p [26], miR-19a-3p [27], miR-20a-5p [28,29], miR-21-3p [30,31], miR-21-5p [26,30], miR-23a-3p [32,33,34], miR-27a-3p [26,34], miR-28-3p [35], miR-30a-3p [36], miR-30a-5p [36], miR-30b-3p [36], miR-30c-5p [36], miR-30d-3p [36], miR-30d-5p [36], miR-30e-3p [36], miR-30e-5p [36], miR-31-5p [26], miR-33a-5p [37], miR-34a-5p [38], miR-34c-5p [39], miR-125a-3p [26], miR-125a-5p [26], miR-126-5p [26,40], miR-133a-3p [41], miR-135a-5p [42], miR-138-5p [43], miR-139-3p [44,45], miR-139-5p [46], miR-141-3p [47], miR-142-3p [48,49], miR-142-5p [48], miR-143-3p [50,51], miR-143-5p [50], miR-145-3p [50,52], miR-145-5p [50,52], miR-148a-3p [27,53], miR-155-5p [54,55], miR-182-5p [56,57], miR-190a-5p [24], miR-190b [24], miR-200a-3p [58], miR-200a-5p [58], miR-200b-3p [58], miR-200b-5p [58], miR-200c-3p [58], miR-203a-3p [42], miR-204-5p [59], miR-210-3p [55,60], miR-211-5p [59], miR-214-3p [61], miR-214-5p [61], miR-218-5p [62], miR-223-3p [47], miR-223-5p [47], miR-296-5p [27], miR-326 [63], miR-335-3p [18], miR-335-5p [64], miR-378a-3p [24], miR-379-5p [65], miR-409-3p [65], miR-409-5p [65], miR-503-5p [24], miR-542-3p [66], miR-542-5p [67], miR-543 [68] | ||
| Selected genes | ||
| Protective for BM | ||
| BMP7 | Bone Morphogenetic Protein 7 | Binds BMPR2 receptor on tumor cells and enhances tumor cell dormancy through ERK/MAPK signaling [69]. |
| BMPR2 | Bone Morphogenetic Protein Receptor Type 2 | Promotes tumor cell dormancy in bone [40]. Inversely correlated with prostate BM occurrence [70]. |
| FOS | Fos Proto-Oncogene | Transcription factor for osteoclastogenesis [71]. |
| NOS3 | Nitric Oxide Synthase 3 | Decreases invasiveness [68]. |
| NOTCH1 | Notch Receptor 1 | Involved in oncogenesis [40]. |
| PDCD4 | Programmed Cell Death 4 | Represses osteoclastogenesis [71]. Downregulation in NSCLC potentially enhances BM [30]. |
| SATB2 | Special AT-Rich Sequence-Binding Protein 2 | Promotes osteoblastogenesis and bone regeneration [72]. Downregulation is associated with metastasis in ccRCC [72]. |
| SRCIN1 | SRC Kinase Signaling Inhibitor 1 | Inhibits osteoclast differentiation [28]. |
| SMAD2 | SMAD Family Member 2 | Decreases invasiveness. Component of the TGFβ pathway. In BC cells, loss of SMAD2 increases BM potential [73]. |
| SMAD4 | SMAD Family Member 4 | Decreases invasiveness [40,74]. |
| OPG | Osteoprotegerin | Decoy receptor preventing RANKL-RANK signaling [9]. |
| Associated with BM | ||
| CD44 | Cluster of differentiator 44 | Promotes invasiveness [40,75]. |
| CDH11 | Cadherin 11 | Promotes osteomimicry [36]. |
| CTGF | Connective Tissue Growth Factor | Stimulates osteoclasts [40]. |
| CXCL8 | C-X-C Motif Chemokine Ligand 8 | Promotes osteoclastogenesis [76]. |
| CXCR4 | C-X-C Motif Chemokine Receptor 4 | Promotes metastasis to osteogenic niches [40,77]. |
| DKK1 | Dickkopf WNT Signaling Pathway Inhibitor 1 | Osteoclast inhibitor [40]. |
| ELK1 | ETS Transcription Factor ELK1 | Promotes osteoblast differentiation [44]. Inducer of c-FOS proto-oncogene and part of ERK/MSK1/Elk-1/Snail signaling pathway (enhances cancer proliferation) [78]. |
| IL11 | Interleukin 11 | Stimulates osteoclasts [40]. |
| ITGA3 | Integrin Subunit Alpha 3 | Promotes invasiveness [40,75]. |
| ITGA5 | Integrin Subunit Alpha 5 | Involved in invasiveness [36], anchors cancer cells to bone [79]. |
| ITGB3 | Integrin Subunit Beta 3 | Integrin promoting osteomimicry and osteolytic metastases [80]. |
| RUNX2 | Runt-Related Transcription Factor 2 | TF essential in osteoblastogenesis and osteomimicry in PC and BC [42]. |
| SMAD1 | SMAD Family Member 1 | Osteoblast differentiation [81]. |
| TCF7 | Transcription Factor 7 | Involved in BM, part of the Wnt/beta-catenin signaling pathway [38]. |
| TGIF2 | TGF-Beta-induced transcription factor 2 | Pro-osteoclastic factor [82]. |
| TGFB1 | Transforming Growth Factor Beta 1 | Stimulates pro-osteoclastic factor production in cancer cells [40]. |
| RANKL | Receptor Activator Of Nuclear Factor Kappa B Ligand | Promotes osteoclastogenesis [10]. |
| RANK | Receptor Activator Of Nuclear Factor Kappa B | Osteoclast activation [83]. |
| All Patients | n = 128 | |
|---|---|---|
| Gender: male | 86 | 67% |
| Median age at diagnosis (years) | 62 | IQR: 55–69 |
| Median OS after diagnosis (months) | 49 | IQR: 21–103.25 |
| Median OS after stage IV (months) | 34 | IQR: 16.5–62.25 |
| Bone metastasis | ||
| BM at time of nephrectomy (n) | 22 | 17.2% |
| Metachronous BM (n) | 106 | 82.8% |
| Median time to metachronous BM (months) | 34 | IQR: 24.25–100.5 |
| IMDC risk group at start of first-line therapy | ||
| Favorable (n) | 14 | 11% |
| Intermediate (n) | 81 | 63% |
| Poor (n) | 33 | 26% |
| First-line targeted therapy | ||
| Sunitinib (n) | 68 | 53% |
| Pazopanib (n) | 31 | 24% |
| Sorafenib (n) | 11 | 9% |
| Temsirolimus (n) | 9 | 7% |
| Nivolumab-Ipilimumab (n) | 6 | 5% |
| Other (n) | 3 | 2% |
| miRNAs Associated with Longer TTBM | |||
|---|---|---|---|
| miR-30b-3p | BM promoting genes: inverse correlation | ||
| RANKL | rho = −0.33; p = 0.002 | miR-30 family inhibits BM in BC by targeting CXCL8 [36] | |
| ITGA3 | rho = −0.31; p = 0.003 | ||
| TCF7 | rho = −0.25; p = 0.018 | ||
| CD44 | rho = −0.24; p = 0.021 | ||
| CXCL8 | rho = −0.21; p = 0.045 | ||
| miR-139-3p | BM protective genes: positive correlation | ||
| NOTCH1 | rho = 0.29; p = 0.005 | ||
| BMPR2 | rho = 0.27; p = 0.011 | ||
| BM promoting genes: inverse correlation | |||
| TCF7 | rho = −0.23; p = 0.027 | ||
| ITGA3 | rho = −0.22; p = 0.038 | ||
| miR-204-5p | BM protective genes: positive correlation | ||
| FOS | rho = 0.28; p = 0.007 | ||
| BMPR2 | rho = 0.32; p = 0.003 | ||
| SATB2 | rho = 0.34; p = 0.001 | ||
| SMAD4 | rho = 0.35; p = 0.001 | ||
| BM promoting genes: inverse correlation | |||
| CD44 | rho = −0.55; p < 0.001 | miR-204 regulates RUNX2 expression and MSC differentiation [85]. | |
| RANKL | rho = −0.42; p < 0.001 | Sponging of miR-204-5p by lncRNA SNHG4 upregulates RUNX2 and promotes tumor progression in RCC [86]. | |
| ITGA3 | rho = −0.42; p < 0.001 | miR-204-5p inhibits TGF-β-induced IL11 production in BM cells of BC [59]. | |
| RUNX2 ° | rho = −0.41; p < 0.001 | miR-204 inhibits growth and motility of CRC cells by CXCL8 downregulation [87]. | |
| ITGA5 | rho = −0.36; p = 0.001 | ||
| CDH11 ° | rho = −0.39; p < 0.001 | ||
| TGFB1 | rho = −0.33; p = 0.002 | ||
| CXCL8 | rho = −0.3; p = 0.005 | ||
| TCF7 | rho = −0.36; p < 0.001 | ||
| CTGF | rho = −0.26; p = 0.014 | ||
| miR-542-5p | BM protective genes: positive correlation | ||
| SATB2 | rho = 0.34; p = 0.001 | miR-542-5p increases SMAD2 expression levels in ICU weakness [88]. | |
| SMAD2 | rho = 0.29; p = 0.006 | ||
| BM promoting genes: inverse correlation | |||
| TCF7 | rho = −0.37; p < 0.001 | ||
| RUNX2 | rho = −0.31; p = 0.003 | ||
| CD44 | rho = −0.32; p = 0.002 | ||
| CXCR4 | rho = −0.21; p = 0.044 | ||
| miRNAs Associated With Shorter TTBM | |||
| miR-21-5p | BM protective genes: inverse correlation | ||
| SATB2 | rho = −0.22; p = 0.04 | ||
| SMAD4 | rho = −0.24; p = 0.026 | ||
| BM promoting genes: positive correlation | |||
| TCF7 | rho = 0.27; p = 0.011 | ||
| CD44 * | rho = 0.25; p = 0.018 | ||
| ITGA3 | rho = 0.29; p = 0.005 | ||
| miR-21-3p | BM protective genes: inverse correlation | ||
| FOS | rho = −0.45; p < 0.001 | miR-21 deficiency results in cFOS upregulation in periodontal tissues [89]. | |
| BMPR2 | rho = −0.42; p < 0.001 | BMPR2 is directly targeted by miR-21 in PC cells [90]. | |
| SATB2 | rho = −0.31; p = 0.003 | miR-21-3p is involved in proliferation and invasion through SMAD4/Erk signaling in CRC [31]. | |
| SMAD4 | rho = 0.22; p = 0.043 | ||
| BM promoting genes: positive correlation | |||
| ITGA3 | rho = 0.42; p < 0.001 | TGFB1 signaling increases miR-21 expression in renal fibrosis [91]. | |
| RANKL | rho = 0.38; p < 0.001 | ||
| CD44 * | rho = 0.37; p < 0.001 | ||
| TGFB1 | rho = 0.25; p = 0.018 | ||
| CXCL8 * | rho = 0.22; p = 0.043 | ||
| miR-28-3p | BM protective genes: inverse correlation | ||
| SATB2 | rho = −0.24; p = 0.022 | ||
| SRCIN1 | rho = −0.22; p = 0.038 | ||
| BM promoting genes: positive correlation | |||
| CXCR4 | rho = 0.24; p = 0.025 | ||
| miR-34c-5p | BM protective genes: inverse correlation | ||
| SATB2 ° | rho = −0.23; p = 0.033 | miR-34s inhibit osteoblast proliferation and differentiation in mouse by targeting SATB2 [92]. | |
| FOS | rho = −0.26; p = 0.015 | SATB2 targeted by miR-34c-5p suppresses proliferation and metastasis attenuating EMT in CRC [39]. | |
| BM promoting genes: positive correlation | |||
| CD44 | rho = 0.3; p = 0.004 | ||
| ITGA3 | rho = 0.38; p < 0.001 | ||
| RANK | rho = 0.21; p = 0.047 | ||
| TGFB1 | rho = 0.26; p = 0.015 | ||
| miR-23a-3p | BM protective genes: positive correlation | ||
| BMPR2 * | rho = 0.23; p = 0.027 | ||
| BM promoting genes: inverse correlation | |||
| CXCL8 * | rho = −0.25; p = 0.017 | ||
| ELK1 | rho = −0.24; p = 0.021 | ||
| miR-20a-5p | BM protective genes: inverse correlation | ||
| OPG * | rho = −0.21; p = 0.044 | ||
| BM protective genes: positive correlation | |||
| PDCD4 ° | rho = 0.28; p = 0.009 | ||
| BM promoting genes: inverse correlation | |||
| ITGB3 | rho = −0.23; p = 0.03 | ||
| BM promoting genes: positive correlation | |||
| IL11 | rho = 0.27; p = 0.011 | RUNX2 is positively correlated with miR-20a-5p in adipose SC during osteogenic differentiation [93]. | |
| TCF7 | rho = 0.26; p = 0.012 | ||
| RUNX2 | rho = 0.22; p = 0.035 | ||
| miR-335-3p | BM protective genes: inverse correlation | ||
| OPG | rho = −0.23; p = 0.03 | ||
| BM promoting genes: positive correlation | |||
| CD44 | rho = 0.31; p = 0.003 | ||
| ITGA5 | rho = 0.29; p = 0.007 | ||
| RANKL | rho = 0.22; p = 0.04 | ||
| CDH11 | rho = 0.36; p = 0.001 | ||
| ITGA3 | rho = 0.27; p = 0.011 | ||
| TGFB1 | rho = 0.29; p = 0.006 | ||
| TCF7 | rho = 0.35; p = 0.001 | ||
| RUNX2 | rho = 0.3; p = 0.004 | ||
| miR-182-5p | BM protective genes: inverse correlation | ||
| SATB2 *,° | rho = −0.29; p = 0.006 | microRNA-182 targets SATB2 to promote CRC proliferation and metastasis [94]. | |
| BMPR2 * | rho = −0.28; p = 0.009 | ||
| OPG | rho = −0.28; p = 0.007 | ||
| BM promoting genes: positive correlation | |||
| CD44 | rho = 0.37; p < 0.001 | ||
| CDH11 * | rho = 0.25; p = 0.02 | ||
| ITGA3 | rho = 0.37; p < 0.001 | ||
| IL11 * | rho = 0.21; p = 0.046 | ||
| CXCL8 | rho = 0.32; p = 0.002 | ||
| TCF7 | rho = 0.34; p = 0.001 | ||
| RUNX2 | rho = 0.27; p = 0.011 | ||
| miR-27a-3p | BM protective genes: inverse correlation | ||
| BMP7 * | rho = −0.29; p = 0.007 | ||
| BM protective genes: positive correlation | |||
| PDCD4 | rho = 0.23; p = 0.031 | ||
| BM promoting genes: inverse correlation | |||
| CXCL8 * | rho = −0.26; p = 0.013 | ||
| miRNA | OS Since Diagnosis | PFS on VEGFR-TKIs | OS on VEGFR-TKIs | |||
| HR (95% CI) | p-value | HR (95% CI) | p-value | HR (95% CI) | p-value | |
| Longer time to bone metastasis | ||||||
| miR-204-5p | 0.73 (0.65–0.82) | <0.001 | 0.84 (0.74–0.94) | 0.003 | 0.81 (0.72–0.92) | 0.001 |
| miR-30b-3p | 0.93 (0.78–1.10) | 0.386 | 1.07 (0.88–1.31) | 0.496 | 0.99 (0.81–1.21) | 0.921 |
| miR-542-5p | 0.85 (0.69–1.05) | 0.133 | 0.96 (0.77–1.20) | 0.728 | 0.89 (0.71–1.11) | 0.296 |
| miR-139-3p | 0.86 (0.73–1.00) | 0.057 | 0.91 (0.76–1.09) | 0.321 | 0.87 (0.73–1.03) | 0.105 |
| Shorter time to bone metastasis | ||||||
| miR-21-5p | 1.69 (1.31–2.18) | <0.001 | 1.14 (0.88–1.47) | 0.327 | 1.29 (1.01–1.65) | 0.043 |
| miR-21-3p | 1.50 (1.31–2.18) | <0.001 | 1.24 (1.04–1.48) | 0.016 | 1.35 (1.13–1.61) | 0.001 |
| miR-28-3p | 1.38 (0.98–1.93) | 0.064 | 1.01 (0.70–1.47) | 0.959 | 1.04 (0.72–1.50) | 0.823 |
| miR-34c-5p | 1.31 (1.16–1.48) | <0.001 | 1.19 (1.05–1.35) | 0.007 | 1.27 (1.11–1.44) | <0.001 |
| miR-23a-3p | 1.05 (0.83–1.33) | 0.697 | 0.98 (0.75–1.30) | 0.903 | 1.01 (0.76–1.34) | 0.953 |
| miR-20a-5p | 1.12 (0.91–1.39) | 0.279 | 0.95 (0.77–1.19) | 0.680 | 0.94 (0.75–1.18) | 0.598 |
| miR-335-3p | 1.22 (1.06–1.41) | 0.005 | 1.10 (0.95–1.26) | 0.193 | 1.11 (0.96–1.28) | 0.155 |
| miR-182-5p | 1.25 (1.09–1.43) | 0.002 | 1.11 (0.96–1.29) | 0.145 | 1.16 (1.01–1.34) | 0.040 |
| miR-27a-3p | 1.13 (0.87–1.47) | 0.351 | 0.87 (0.65–1.17) | 0.365 | 1.03 (0.77–1.39) | 0.831 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kinget, L.; Roussel, E.; Lambrechts, D.; Boeckx, B.; Vanginderhuysen, L.; Albersen, M.; Rodríguez-Antona, C.; Graña-Castro, O.; Inglada-Pérez, L.; Verbiest, A.; et al. MicroRNAs Possibly Involved in the Development of Bone Metastasis in Clear-Cell Renal Cell Carcinoma. Cancers 2021, 13, 1554. https://doi.org/10.3390/cancers13071554
Kinget L, Roussel E, Lambrechts D, Boeckx B, Vanginderhuysen L, Albersen M, Rodríguez-Antona C, Graña-Castro O, Inglada-Pérez L, Verbiest A, et al. MicroRNAs Possibly Involved in the Development of Bone Metastasis in Clear-Cell Renal Cell Carcinoma. Cancers. 2021; 13(7):1554. https://doi.org/10.3390/cancers13071554
Chicago/Turabian StyleKinget, Lisa, Eduard Roussel, Diether Lambrechts, Bram Boeckx, Loïc Vanginderhuysen, Maarten Albersen, Cristina Rodríguez-Antona, Osvaldo Graña-Castro, Lucía Inglada-Pérez, Annelies Verbiest, and et al. 2021. "MicroRNAs Possibly Involved in the Development of Bone Metastasis in Clear-Cell Renal Cell Carcinoma" Cancers 13, no. 7: 1554. https://doi.org/10.3390/cancers13071554
APA StyleKinget, L., Roussel, E., Lambrechts, D., Boeckx, B., Vanginderhuysen, L., Albersen, M., Rodríguez-Antona, C., Graña-Castro, O., Inglada-Pérez, L., Verbiest, A., Zucman-Rossi, J., Couchy, G., Caruso, S., Laenen, A., Baldewijns, M., & Beuselinck, B. (2021). MicroRNAs Possibly Involved in the Development of Bone Metastasis in Clear-Cell Renal Cell Carcinoma. Cancers, 13(7), 1554. https://doi.org/10.3390/cancers13071554