
Molecular Pathogenesis and Regulation of the miR-29-3p-Family: Involvement of ITGA6 and ITGB1 in Intra-Hepatic Cholangiocarcinoma

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Results
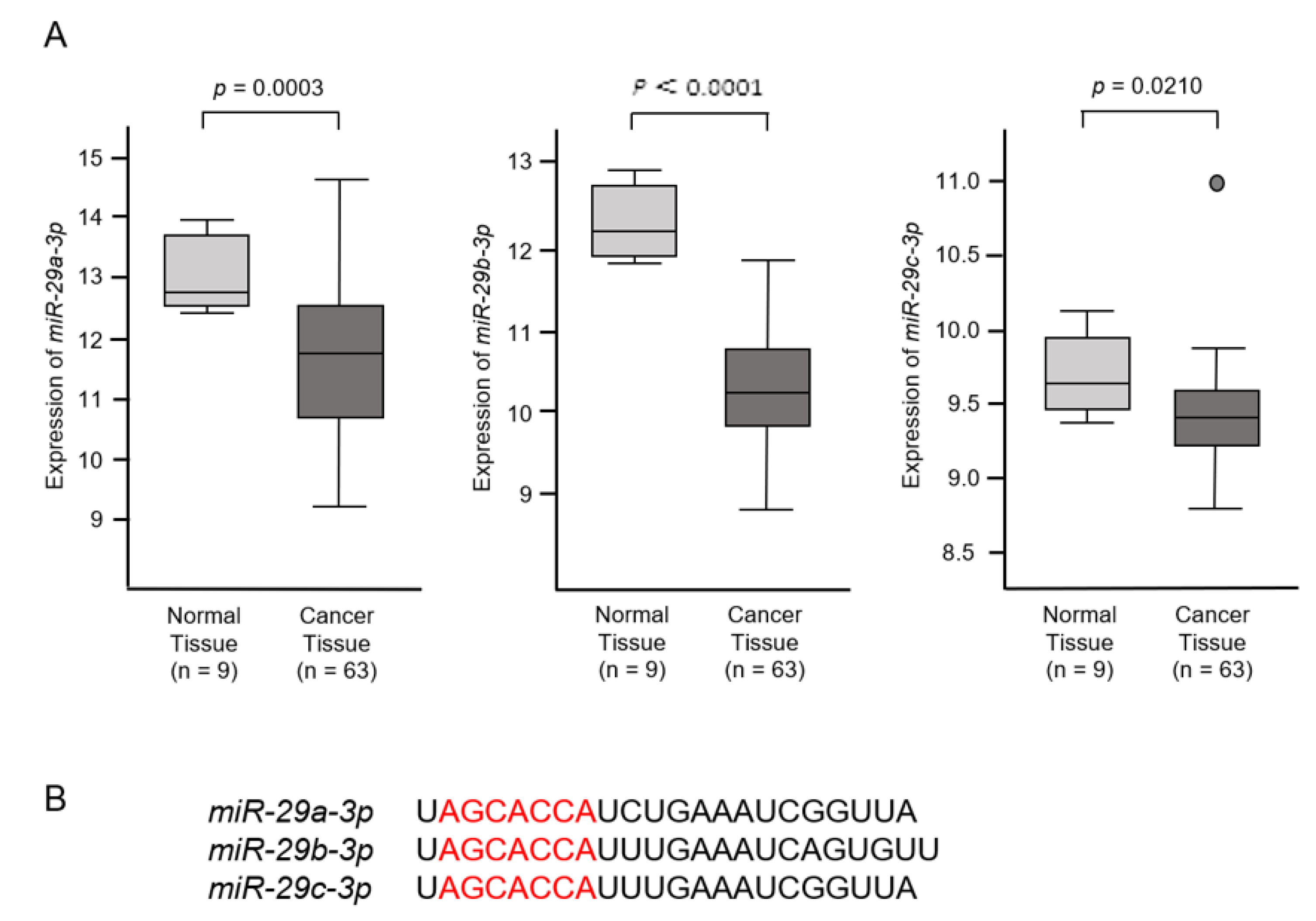
2.1. Downregulation of miR-29a-3p, miR-29b-3p and miR-29c-3p in ICC Clinical Specimens
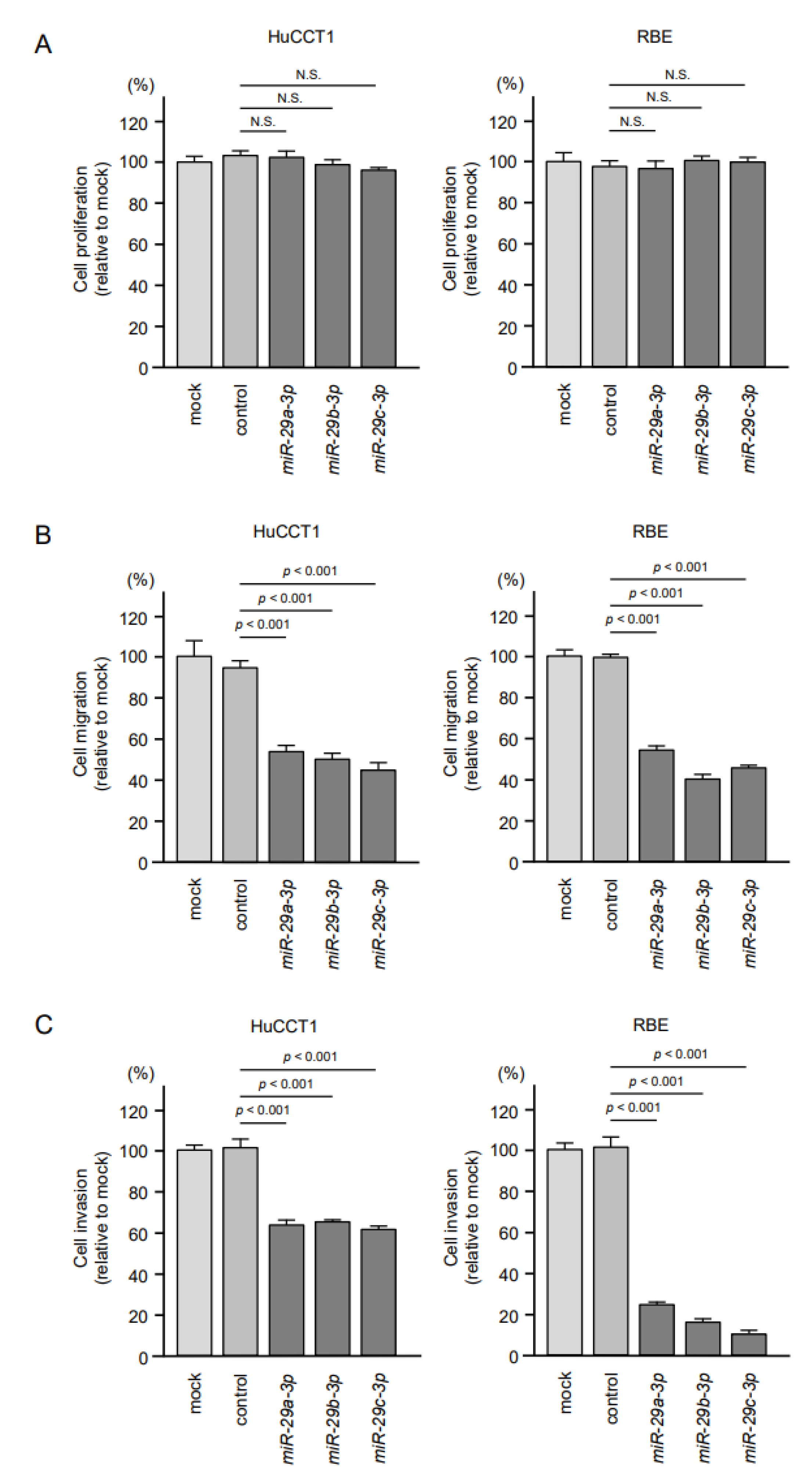
2.2. Effects of Ectopic Expression of miR-29a-3p, miR-29b-3p and miR-29c-3p on ICC Cell Proliferation, Migration and Invasion
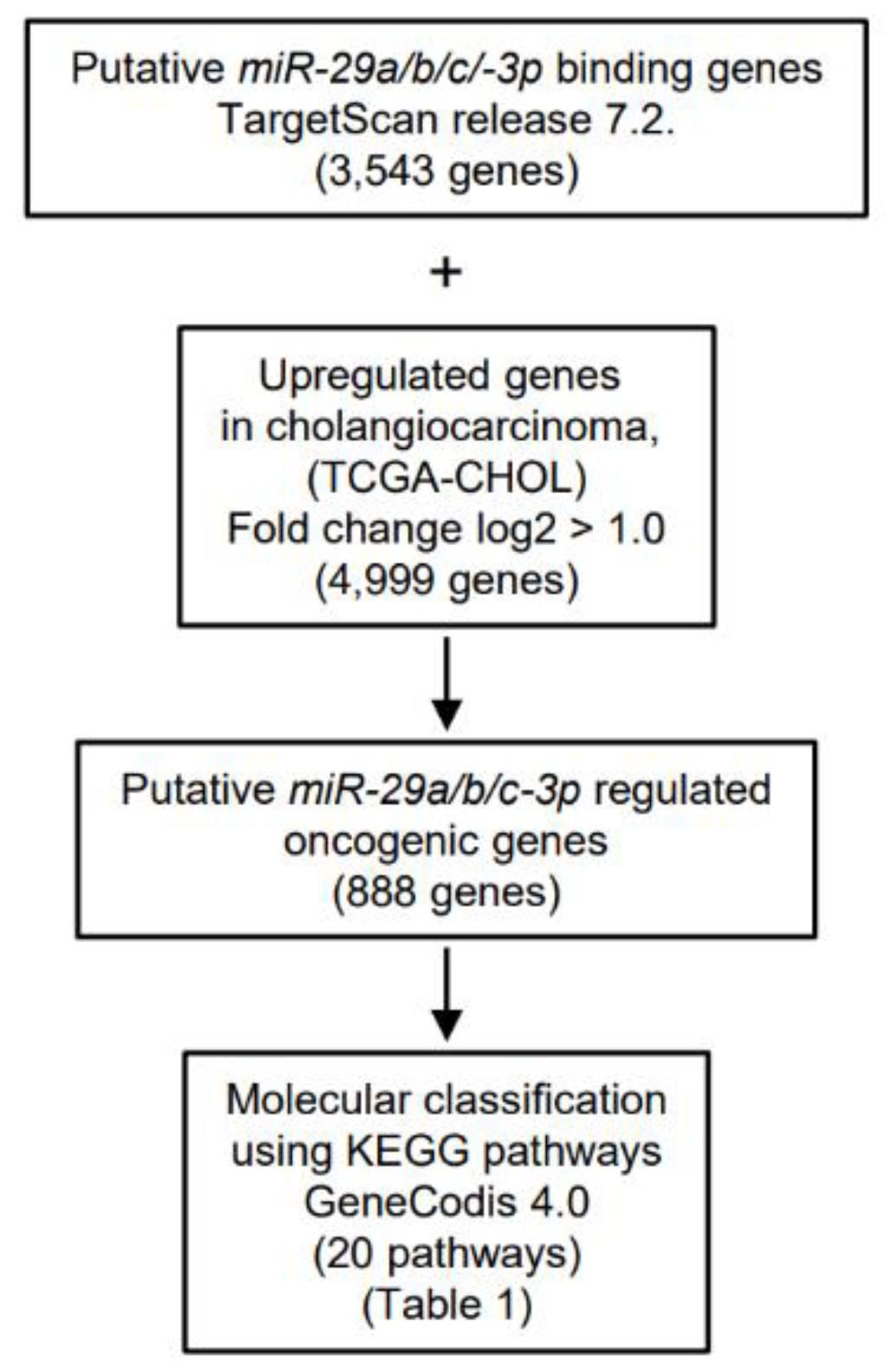
2.3. Identification of Putative Pathways and Targets of the miR-29-3p-Family (miR-29a/b/c-3p) Regulation in ICC Cells
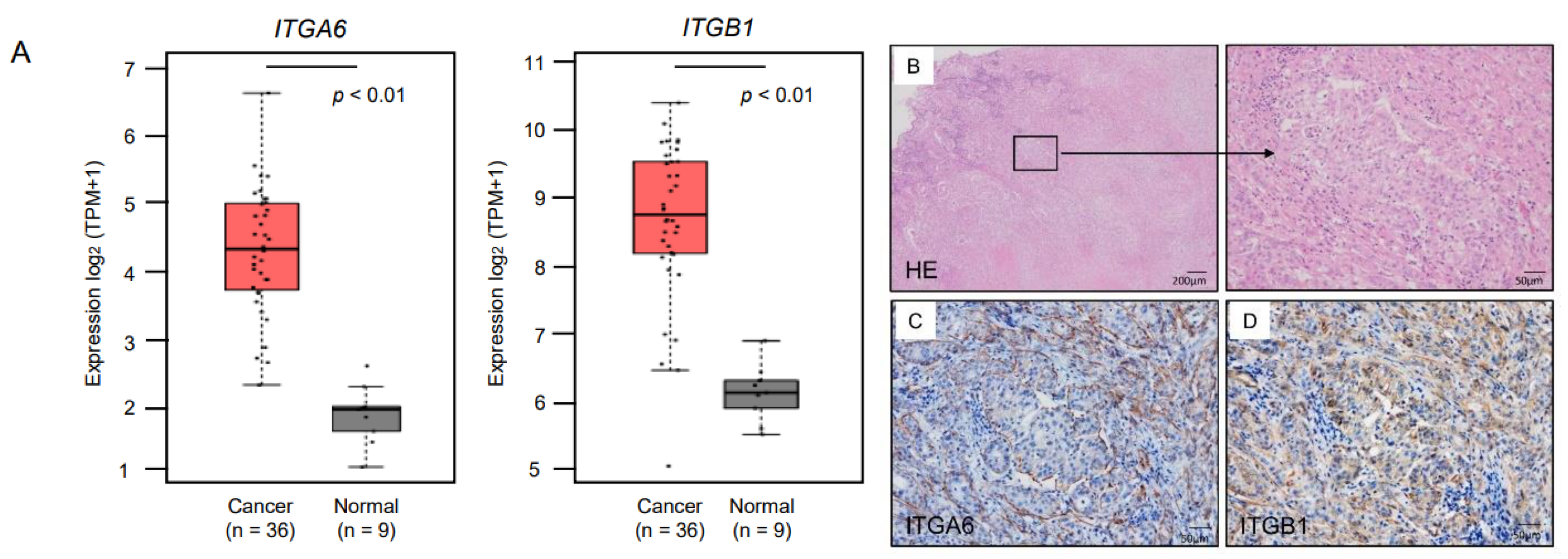
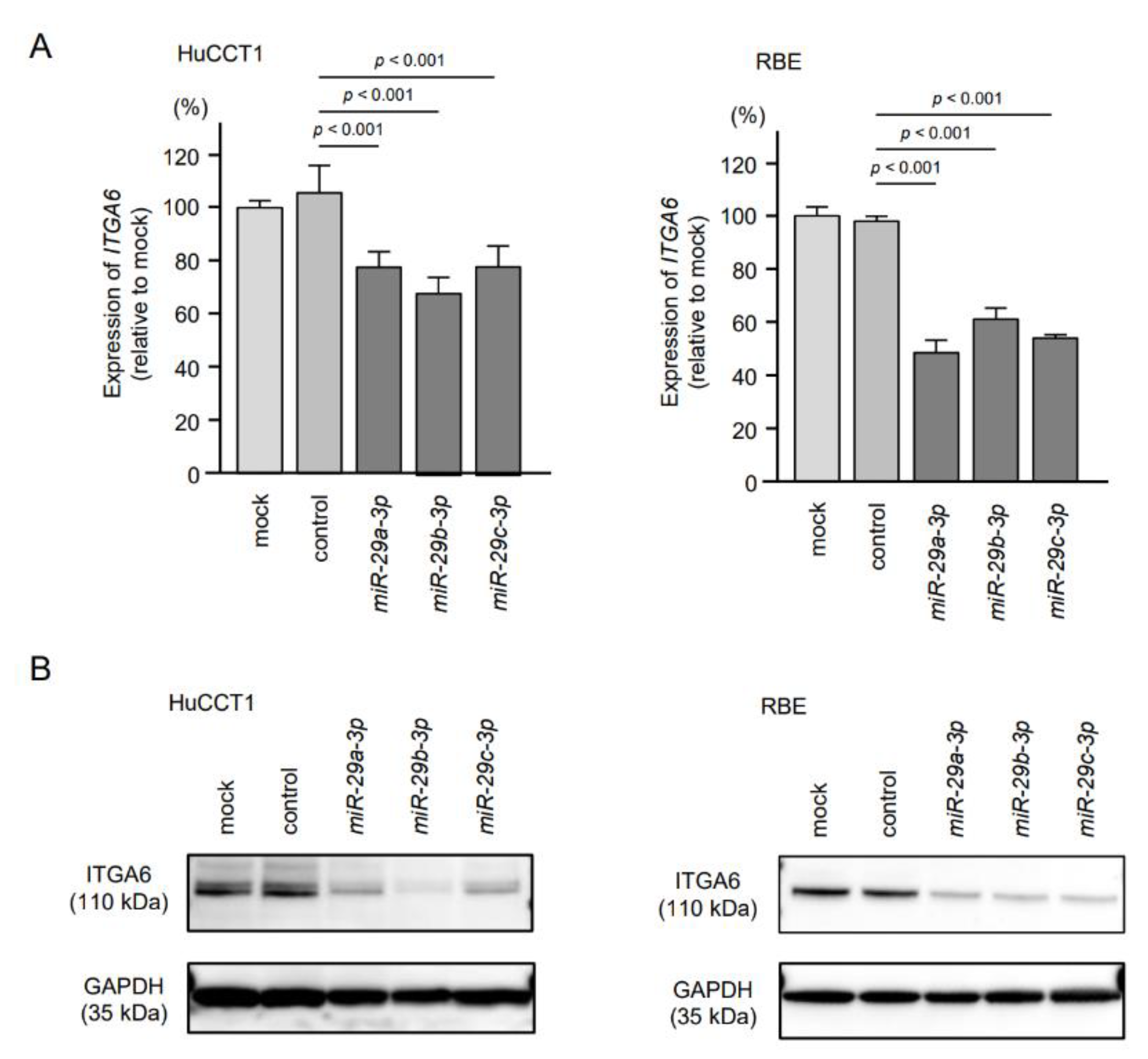
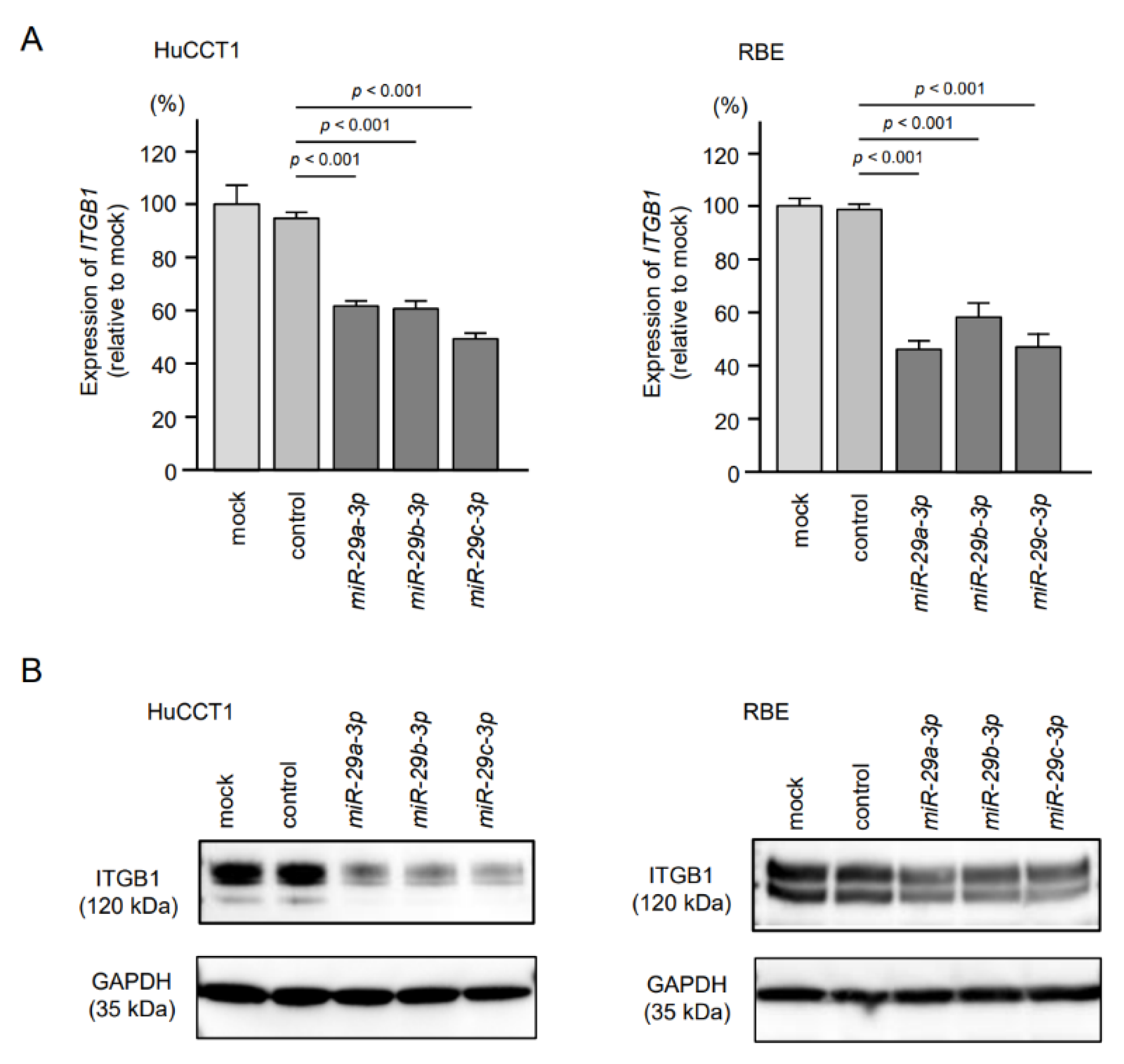
2.4. Expression of ITGA6/ITGA6 and ITGB1/ITGB1 Genes and Proteins in ICC Clinical Specimens
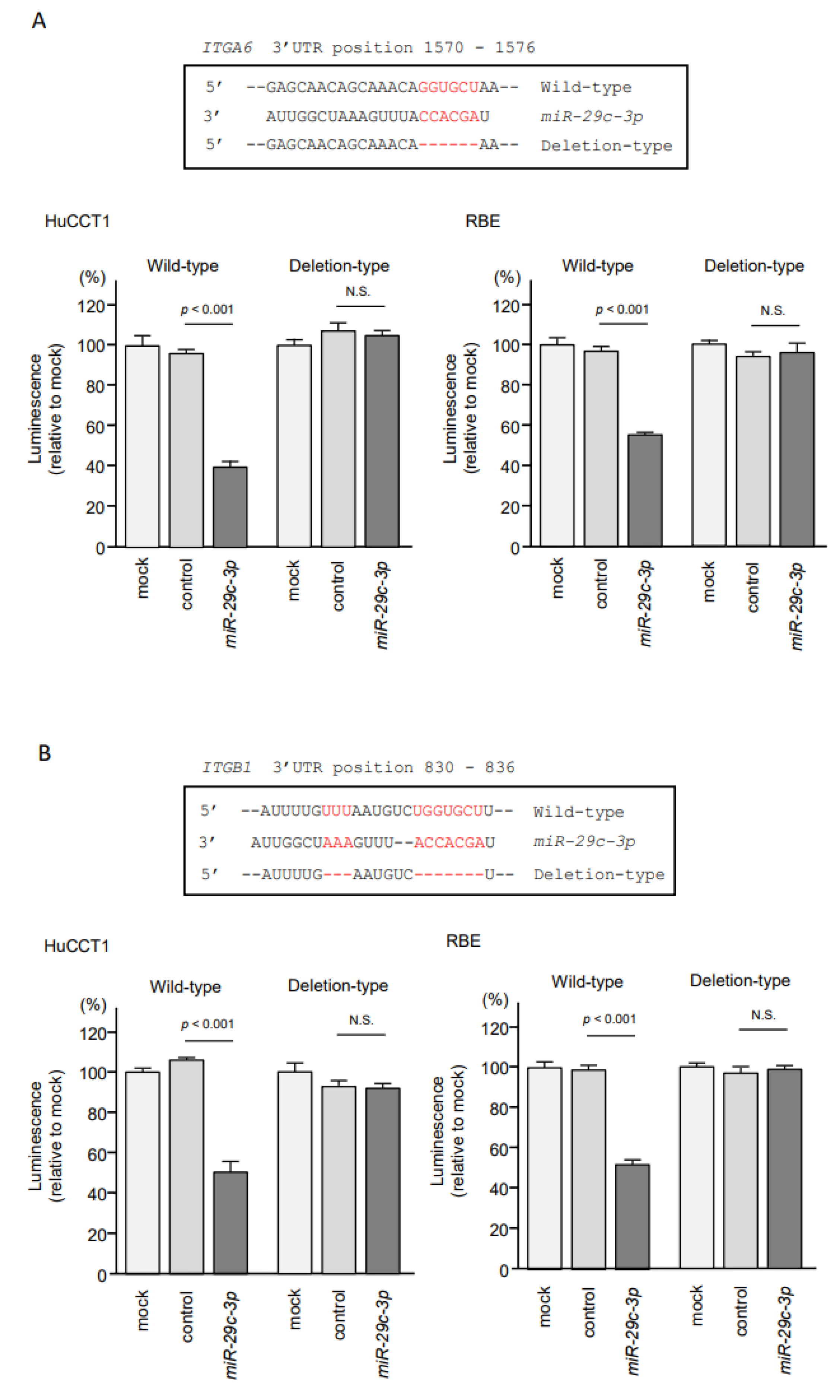
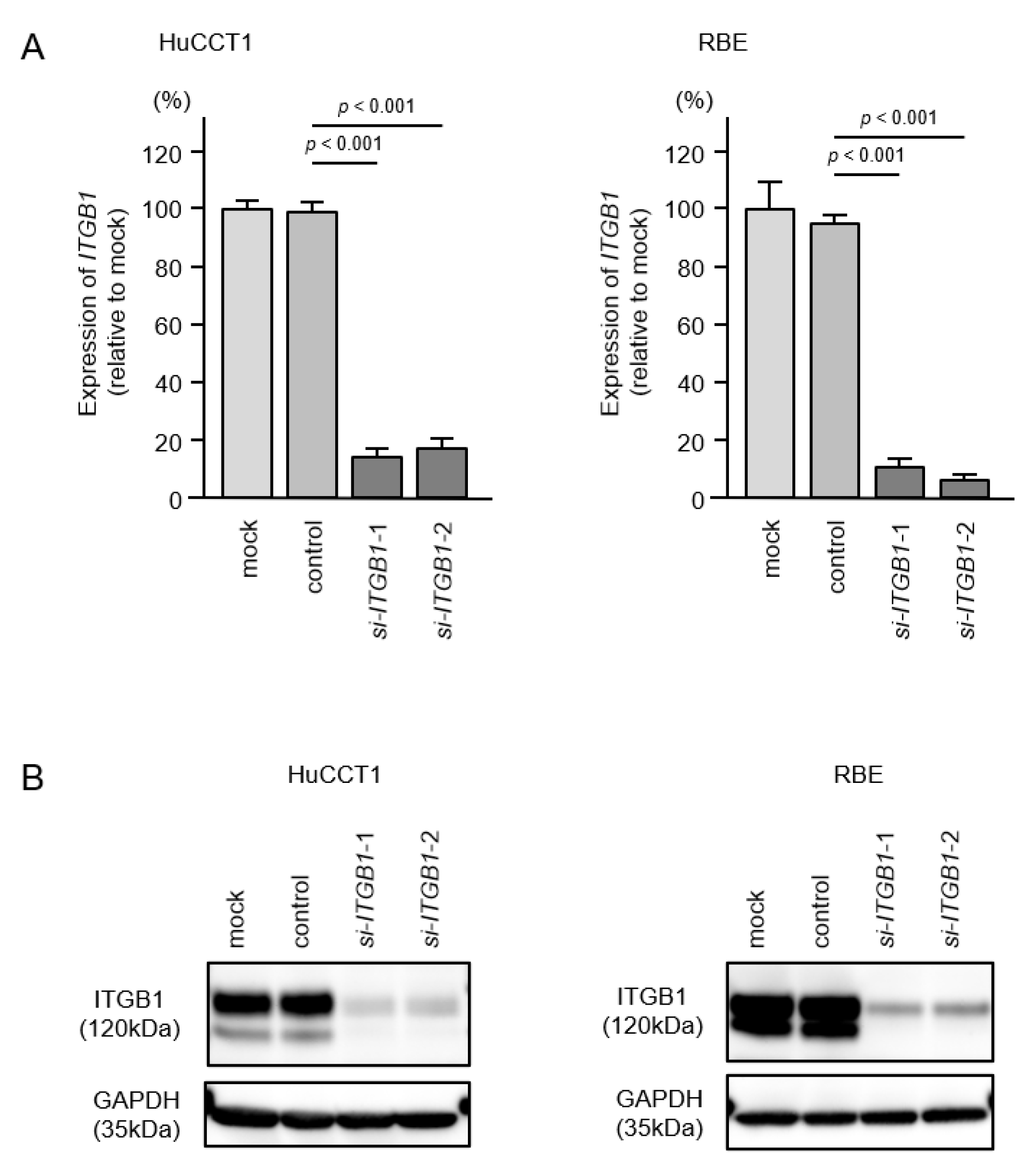
2.5. Direct Regulation of ITGA6 and ITGB1 by miR-29a-3p, miR-29b-3p and miR-29c-3p in ICC Cells
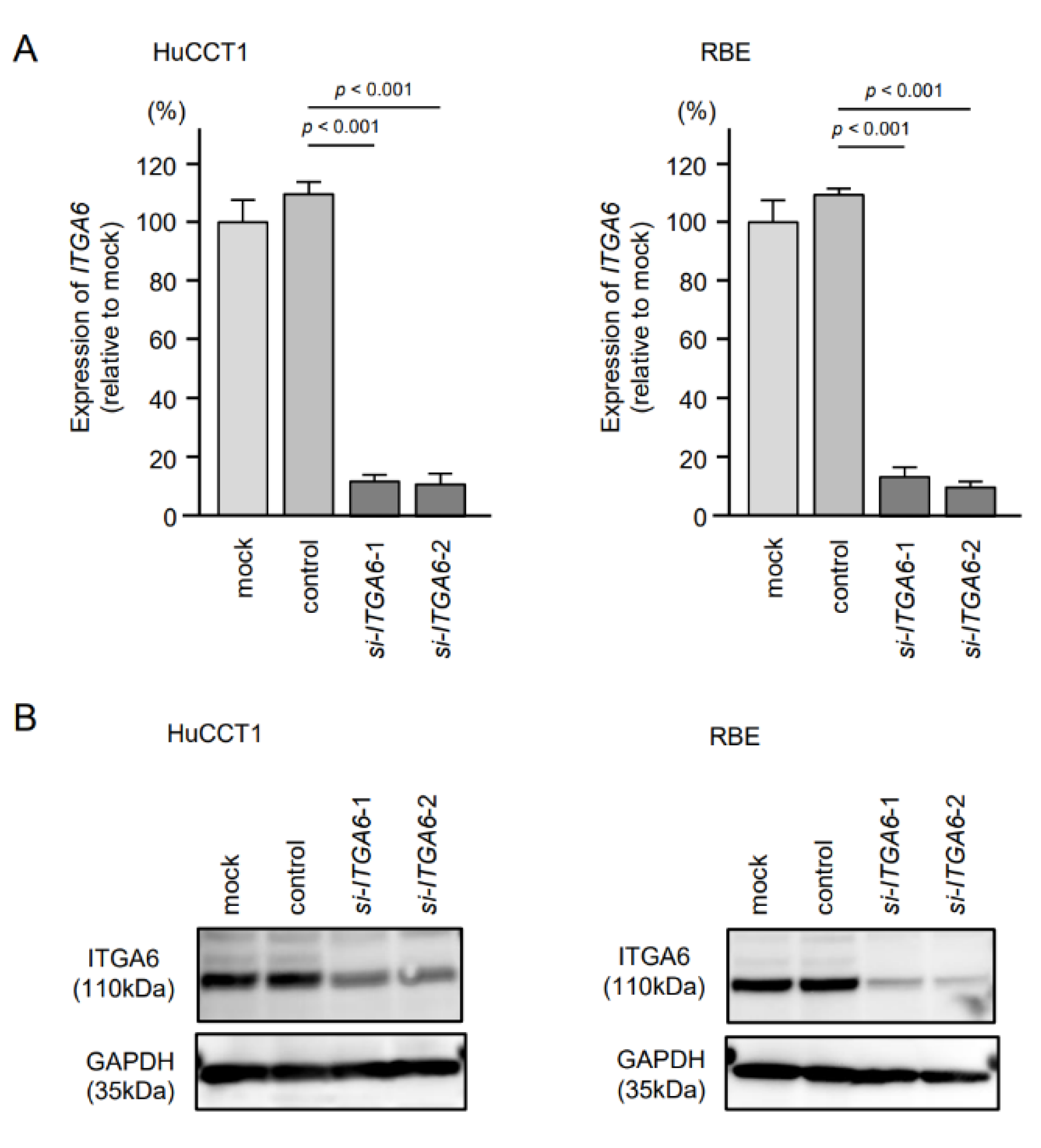
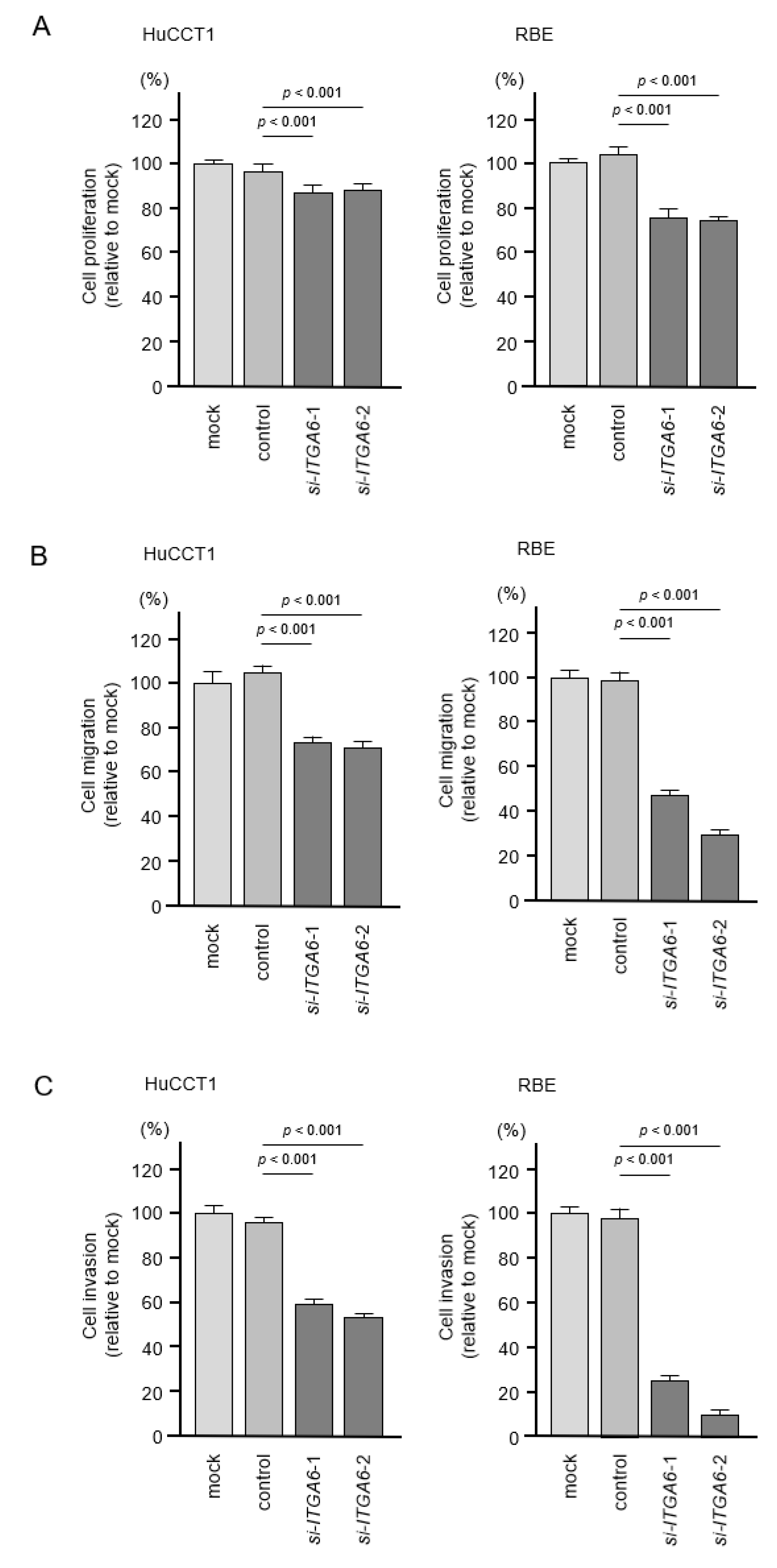
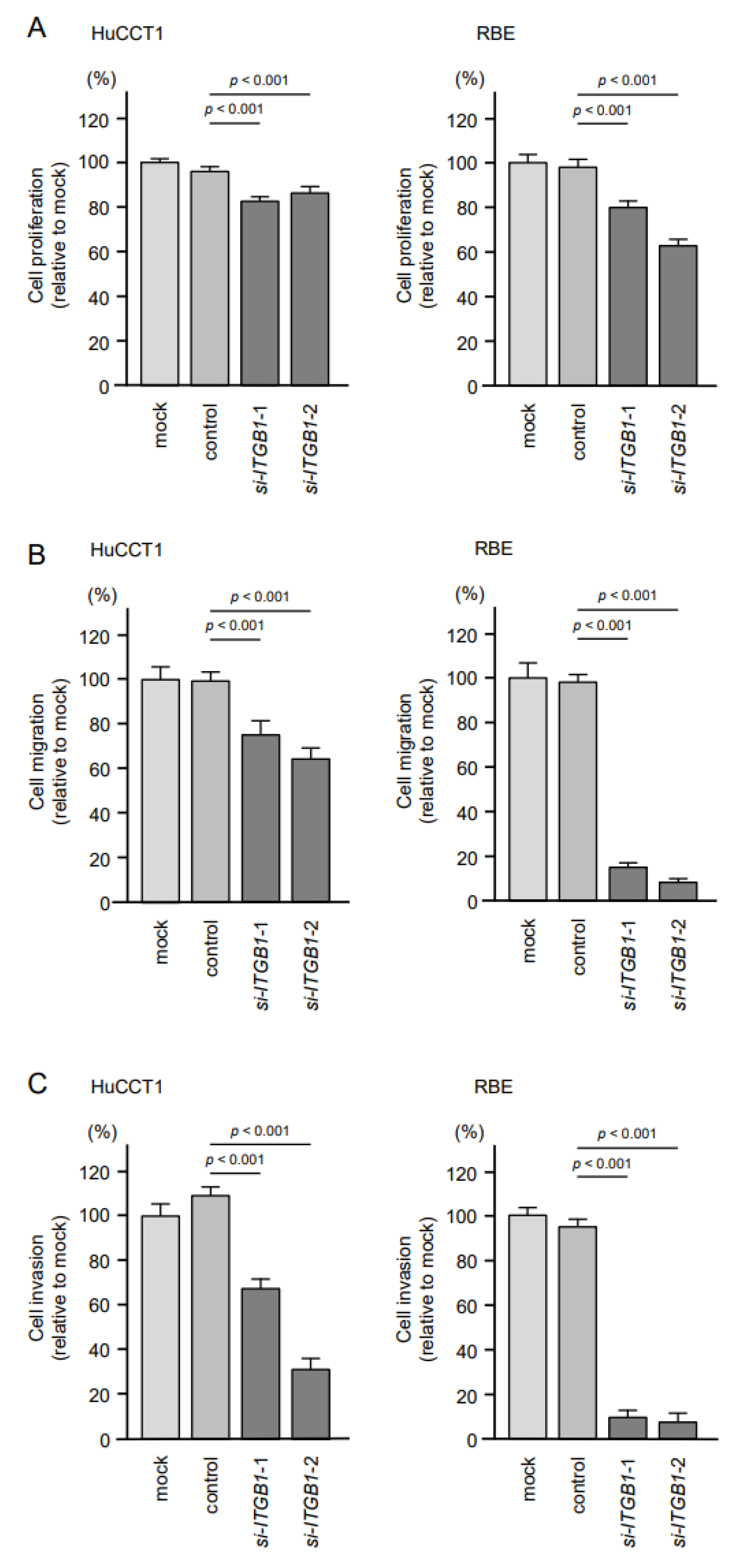
2.6. Effects of Knockdown of ITGA6 and ITGB1 on Cell Proliferation, Migration, and Invasion in ICC Cells
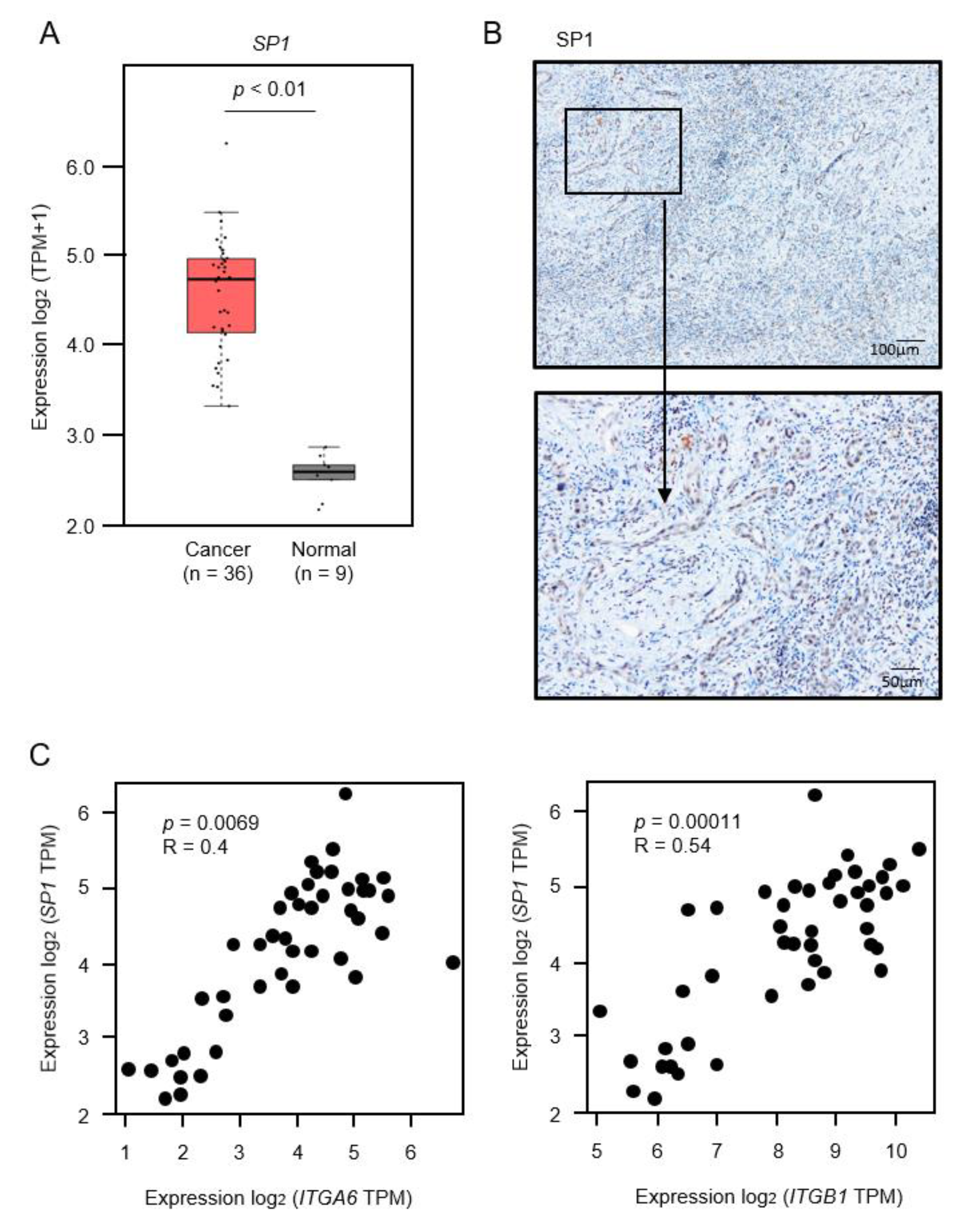
2.7. Identification of Transcriptional Regulators of ITGA6 and ITGB1 in ICC Cells
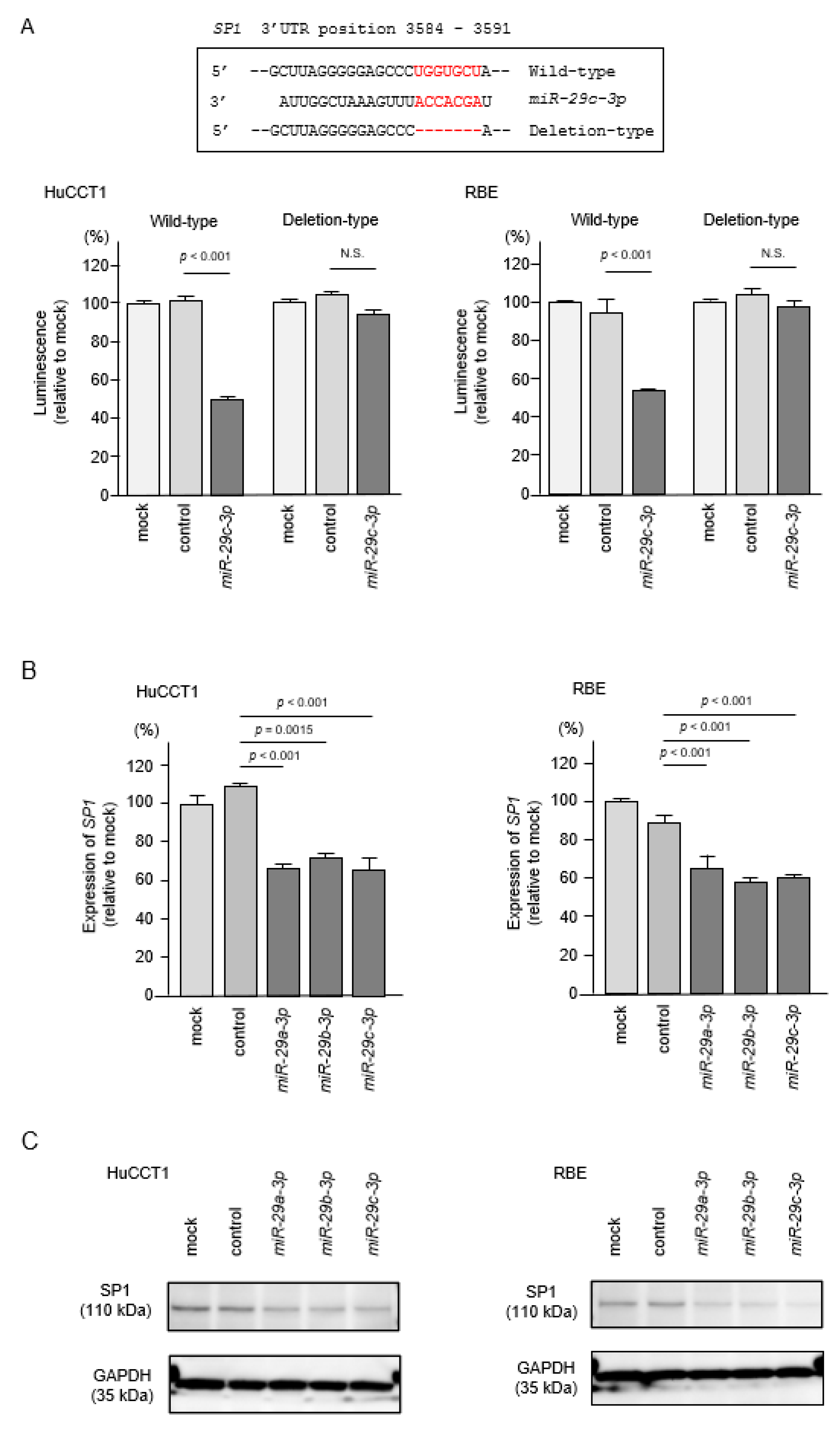
2.8. Regulation of SP1 by miR-29a-3p, miR-29b-3p and miR-29c-3p in ICC Cells
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. ICC Cell Lines and Cell Culture
5.2. RNA Extraction and Quantitative Real-Time Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
5.3. Transfection of miRNAs, siRNAs, and Plasmid Vectors into ICC Cells
5.4. Functional Assays in ICC Cells (Cell Proliferation, Migration and Invasion)
5.5. Expression Analysis of miRNA, Target Genes and Transcriptional Factors Using Public Databases
5.6. Plasmid Construction and Dual-Luciferase Reporter Assays
5.7. Western Blotting and Immunohistochemistry
5.8. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Banales, J.M.; Marin, J.J.G.; Lamarca, A.; Rodrigues, P.M.; Khan, S.A.; Roberts, L.R.; Cardinale, V.; Carpino, G.; Andersen, J.B.; Braconi, C.; et al. Cholangiocarcinoma 2020: The next horizon in mechanisms and management. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 577–588. [Google Scholar] [CrossRef]
- Fostea, R.M.; Fontana, E.; Torga, G.; Arkenau, H.-T. Recent Progress in the Systemic Treatment of Advanced/Metastatic Cholangiocarcinoma. Cancers 2020, 12, 2599. [Google Scholar] [CrossRef]
- Ghidini, M.; Pizzo, C.; Botticelli, A.; Hahne, J.C.; Passalacqua, R.; Tomasello, G.; Petrelli, F. Biliary tract cancer: Current challenges and future prospects. Cancer Manag. Res. 2018, 11, 379–388. [Google Scholar] [CrossRef] [Green Version]
- McGlynn, K.A.; Tarone, R.E.; El-Serag, H.B. A Comparison of Trends in the Incidence of Hepatocellular Carcinoma and Intrahepatic Cholangiocarcinoma in the United States. Cancer Epidemiol. Biomark. Prev. 2006, 15, 1198–1203. [Google Scholar] [CrossRef] [Green Version]
- Bertuccio, P.; Bosetti, C.; Levi, F.; Decarli, A.; Negri, E.; La Vecchia, C. A comparison of trends in mortality from primary liver cancer and intrahepatic cholangiocarcinoma in Europe. Ann. Oncol. 2013, 24, 1667–1674. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.A.; Taylor-Robinson, S.D.; Toledano, M.B.; Beck, A.; Elliott, P.; Thomas, H.C. Changing international trends in mortality rates for liver, biliary and pancreatic tumours. J. Hepatol. 2002, 37, 806–813. [Google Scholar] [CrossRef]
- Patel, T. Worldwide trends in mortality from biliary tract malignancies. BMC Cancer 2002, 2, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaib, Y.H.; Davila, J.A.; McGlynn, K.; El-Serag, H.B. Rising incidence of intrahepatic cholangiocarcinoma in the United States: A true increase? J. Hepatol. 2004, 40, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhong, M.; Feng, Y.; Zeng, S.; Wang, Y.; Xu, H.; Zhou, H. Prognostic Factors and Treatment Strategies for Intrahepatic Cholangiocarcinoma from 2004 to 2013: Population-Based SEER Analysis. Transl. Oncol. 2019, 12, 1496–1503. [Google Scholar] [CrossRef]
- Doherty, B.; Nambudiri, V.E.; Palmer, W.C. Update on the Diagnosis and Treatment of Cholangiocarcinoma. Curr. Gastroenterol. Rep. 2017, 19, 2. [Google Scholar] [CrossRef] [PubMed]
- Macias, R.I.R.; Kornek, M.; Rodrigues, P.M.; Paiva, N.A.; Castro, R.E.; Urban, S.; Pereira, S.P.; Cadamuro, M.; Rupp, C.; Loosen, S.H.; et al. Diagnostic and prognostic biomarkers in cholangiocarcinoma. Liver Int. 2019, 39, 108–122. [Google Scholar] [CrossRef] [Green Version]
- Anfossi, S.; Babayan, A.; Pantel, K.; Calin, G.A. Clinical utility of circulating non-coding RNAs—An update. Nat. Rev. Clin. Oncol. 2018, 15, 541–563. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Gregory, R.I. MicroRNA biogenesis pathways in cancer. Nat. Rev. Cancer 2015, 15, 321–333. [Google Scholar] [CrossRef]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Wada, M.; Goto, Y.; Tanaka, T.; Okada, R.; Moriya, S.; Idichi, T.; Noda, M.; Sasaki, K.; Kita, Y.; Kurahara, H.; et al. RNA sequencing-based microRNA expression signature in esophageal squamous cell carcinoma: Oncogenic targets by antitumor miR-143-5p and miR-143-3p regulation. J. Hum. Genet. 2020, 65, 1019–1034. [Google Scholar] [CrossRef]
- Idichi, T.; Seki, N.; Kurahara, H.; Fukuhisa, H.; Toda, H.; Shimonosono, M.; Yamada, Y.; Arai, T.; Kita, Y.; Kijima, Y.; et al. Involvement of anti-tumor miR-124-3p and its targets in the pathogenesis of pancreatic ductal adenocarcinoma: Direct regulation of ITGA3 and ITGB1 by miR-124-3p. Oncotarget 2018, 9, 28849–28865. [Google Scholar] [CrossRef]
- Kawagoe, K.; Wada, M.; Idichi, T.; Okada, R.; Yamada, Y.; Moriya, S.; Okubo, K.; Matsushita, D.; Arigami, T.; Kurahara, H.; et al. Regulation of aberrantly expressed SERPINH1 by antitumor miR-148a-5p inhibits cancer cell aggressiveness in gastric cancer. J. Hum. Genet. 2020, 65, 647–656. [Google Scholar] [CrossRef]
- Zhang, M.-Y.; Li, S.-H.; Huang, G.-L.; Lin, G.-H.; Shuang, Z.-Y.; Lao, X.-M.; Xu, L.; Lin, X.-J.; Wang, H.-Y.; Li, S.-P. Identification of a novel microRNA signature associated with intrahepatic cholangiocarcinoma (ICC) patient prognosis. BMC Cancer 2015, 15, 64. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Yan, H.-X.; Yang, W.; Hu, L.; Yu, L.-X.; Liu, Q.; Li, L.; Huang, D.-D.; Ding, J.; Shen, F.; et al. The role of microRNA expression pattern in human intrahepatic cholangiocarcinoma. J. Hepatol. 2009, 50, 358–369. [Google Scholar] [CrossRef]
- Li, H.; Zhou, Z.-Q.; Yang, Z.-R.; Tong, D.-N.; Guan, J.; Shi, B.-J.; Nie, J.; Ding, X.-T.; Li, B.; Zhou, G.-W.; et al. MicroRNA-191 acts as a tumor promoter by modulating the TET1-p53 pathway in intrahepatic cholangiocarcinoma. Hepatology 2017, 66, 136–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koshizuka, K.; Kikkawa, N.; Hanazawa, T.; Yamada, Y.; Okato, A.; Arai, T.; Katada, K.; Okamoto, Y.; Seki, N. Inhibition of integrin β1-mediated oncogenic signalling by the antitumor microRNA-29 family in head and neck squamous cell carcinoma. Oncotarget 2017, 9, 3663–3676. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, K.; Seki, N.; Mataki, H.; Matsushita, R.; Kamikawaji, K.; Kumamoto, T.; Takagi, K.; Goto, Y.; Nishikawa, R.; Kato, M.; et al. Tumor-suppressive microRNA-29 family inhibits cancer cell migration and invasion directly targeting LOXL2 in lung squamous cell carcinoma. Int. J. Oncol. 2016, 48, 450–460. [Google Scholar] [CrossRef]
- Qin, X.; Song, Y. Bioinformatics Analysis Identifies the Estrogen Receptor 1 (ESR1) Gene and hsa-miR-26a-5p as Potential Prognostic Biomarkers in Patients with Intrahepatic Cholangiocarcinoma. Med. Sci. Monit. 2020, 26, 921815. [Google Scholar] [CrossRef]
- Poos, K.; Smida, J.; Nathrath, M.; Maugg, D.; Baumhoer, D.; Korsching, E. How MicroRNA and Transcription Factor Co-regulatory Networks Affect Osteosarcoma Cell Proliferation. PLoS Comput. Biol. 2013, 9, e1003210. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, H.; Zhou, Z.; Ma, Z.; Li, Z.; Liu, C.; Huang, S.; Zhang, C.; Hou, B. Characterization of the prognostic and oncologic values of ITGB superfamily members in pancreatic cancer. J. Cell. Mol. Med. 2020, 24, 13481–13493. [Google Scholar] [CrossRef]
- Hamurcu, Z.; Kahraman, N.; Ashour, A.; Ozpolat, B. FOXM1 transcriptionally regulates expression of integrin β1 in triple-negative breast cancer. Breast Cancer Res. Treat. 2017, 163, 485–493. [Google Scholar] [CrossRef]
- Nakamura, H.; Arai, Y.; Totoki, Y.; Shirota, T.; ElZawahry, A.; Kato, M.; Hama, N.; Hosoda, F.; Urushidate, T.; Ohashi, S.; et al. Genomic spectra of biliary tract cancer. Nat. Genet. 2015, 47, 1003–1010. [Google Scholar] [CrossRef]
- Fujimoto, A.; Furuta, M.; Totoki, Y.; Tsunoda, T.; Kato, M.; Shiraishi, Y.; Tanaka, H.; Taniguchi, H.; Kawakami, Y.; Ueno, M.; et al. Whole-genome mutational landscape and characterization of noncoding and structural mutations in liver cancer. Nat. Genet. 2016, 48, 500–509. [Google Scholar] [CrossRef]
- Xia, L.; Chen, X.; Yang, J.; Zhu, S.; Zhang, L.; Yin, Q.; Hong, Y.; Chen, H.; Chen, G.; Li, H. Long Non-Coding RNA-PAICC Promotes the Tumorigenesis of Human Intrahepatic Cholangiocarcinoma by Increasing YAP1 Transcription. Front. Oncol. 2021, 10, 595533. [Google Scholar] [CrossRef]
- Li, O.; Yi, W.; Yang, P.; Guo, C.; Peng, C. Long non-coding RNA UCA1 promotes proliferation and invasion of intrahepatic cholangiocarcinoma cells through targeting microRNA-122. Exp. Ther. Med. 2019, 18, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, T.; Nohata, N.; Hanazawa, T.; Kikkawa, N.; Yamamoto, N.; Yoshino, H.; Itesako, T.; Enokida, H.; Nakagawa, M.; Okamoto, Y.; et al. Tumour-suppressive microRNA-29s inhibit cancer cell migration and invasion by targeting laminin–integrin signalling in head and neck squamous cell carcinoma. Br. J. Cancer 2013, 109, 2636–2645. [Google Scholar] [CrossRef] [Green Version]
- Leng, Y.; Chen, Z.; Ding, H.; Zhao, X.; Qin, L.; Pan, Y. Overexpression of microRNA-29b inhibits epithelial-mesenchymal transition and angiogenesis of colorectal cancer through the ETV4/ERK/EGFR axis. Cancer Cell Int. 2021, 21, 17. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Luo, S.; Ji, C.; Shi, J. miR-29c-3p regulates proliferation and migration in ovarian cancer by targeting KIF4A. World J. Surg. Oncol. 2020, 18, 315. [Google Scholar] [CrossRef]
- Yang, J.; Gong, X.; Yang, J.; Ouyang, L.; Xiao, R.; You, X.; Ouyang, Y. Suppressive role of microRNA-29 in hepatocellular carcinoma via targeting IGF2BP1. Int. J. Clin. Exp. Pathol. 2018, 11, 1175–1185. [Google Scholar] [PubMed]
- Zhang, N.-S.; Dai, G.-L.; Liu, S.-J. MicroRNA-29 family functions as a tumor suppressor by targeting RPS15A and regulating cell cycle in hepatocellular carcinoma. Int. J. Clin. Exp. Pathol. 2017, 10, 8031–8042. [Google Scholar]
- Wong, C.C.-L.; Tse, A.P.-W.; Huang, Y.-P.; Zhu, Y.-T.; Chiu, D.K.-C.; Lai, R.K.-H.; Au, S.L.-K.; Kai, A.K.-L.; Lee, J.M.-F.; Wei, L.L.; et al. Lysyl oxidase-like 2 is critical to tumor microenvironment and metastatic niche formation in hepatocellular carcinoma. Hepatology 2014, 60, 1645–1658. [Google Scholar] [CrossRef]
- Loosen, S.H.; Lurje, G.; Wiltberger, G.; Vucur, M.; Koch, A.; Kather, J.N.; Paffenholz, P.; Tacke, F.; Ulmer, F.T.; Trautwein, C.; et al. Serum levels of miR-29, miR-122, miR-155 and miR-192 are elevated in patients with cholangiocarcinoma. PLoS ONE 2019, 14, e0210944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takada, Y.; Ye, X.; Simon, S. The integrins. Genome Biol. 2007, 8, 215. [Google Scholar] [CrossRef] [Green Version]
- Sekiya, Y.; Ogawa, T.; Yoshizato, K.; Ikeda, K.; Kawada, N. Suppression of hepatic stellate cell activation by microRNA-29b. Biochem. Biophys. Res. Commun. 2011, 412, 74–79. [Google Scholar] [CrossRef]
- Volpes, R.; Oord, J.J.V.D.; Desmet, V.J. Integrins as differential cell lineage markers of primary liver tumors. Am. J. Pathol. 1993, 142, 1483–1492. [Google Scholar]
- Koshizuka, K.; Nohata, N.; Hanazawa, T.; Kikkawa, N.; Arai, T.; Okato, A.; Fukumoto, I.; Katada, K.; Okamoto, Y.; Seki, N. Deep sequencing-based microRNA expression signatures in head and neck squamous cell carcinoma: Dual strands of pre-miR-150 as antitumor miRNAs. Oncotarget 2017, 8, 30288–30304. [Google Scholar] [CrossRef] [Green Version]
- Koshizuka, K.; Hanazawa, T.; Kikkawa, N.; Arai, T.; Okato, A.; Kurozumi, A.; Kato, M.; Katada, K.; Okamoto, Y.; Seki, N. Regulation of ITGA3 by the anti-tumor miR-199 family inhibits cancer cell migration and invasion in head and neck cancer. Cancer Sci. 2017, 108, 1681–1692. [Google Scholar] [CrossRef] [Green Version]
- Sandborn, W.J.; Feagan, B.G.; Rutgeerts, P.; Hanauer, S.; Colombel, J.-F.; Sands, B.E.; Lukas, M.; Fedorak, R.; Lee, S.; Bressler, B.; et al. Vedolizumab as Induction and Maintenance Therapy for Crohn’s Disease. N. Engl. J. Med. 2013, 369, 711–721. [Google Scholar] [CrossRef] [Green Version]
- Mas-Moruno, C.; Rechenmacher, F.; Kessler, H. Cilengitide: The First Anti-Angiogenic Small Molecule Drug Candidate. Design, Synthesis and Clinical Evaluation. Anti-Cancer Agents Med. Chem. 2010, 10, 753–768. [Google Scholar] [CrossRef] [Green Version]
- Kurozumi, K.; Ichikawa, T.; Onishi, M.; Fujii, K.; Date, I. Cilengitide Treatment for Malignant Glioma: Current Status and Future Direction. Neurol. Med. Chir. 2012, 52, 539–547. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, P.C.; Peter, K. GPIIb/IIIa inhibitors: From bench to bedside and back to bench again. Thromb. Haemost. 2012, 107, 808–814. [Google Scholar] [CrossRef]
- Li, J.; Du, S.; Sheng, X.; Liu, J.; Cen, B.; Huang, F.; He, Y. MicroRNA-29b Inhibits Endometrial Fibrosis by Regulating the Sp1-TGF-β1/Smad-CTGF Axis in a Rat Model. Reprod. Sci. 2015, 23, 386–394. [Google Scholar] [CrossRef]
- Zhang, G.; Shi, H.; Wang, L.; Zhou, M.; Wang, Z.; Liu, X.; Cheng, L.; Li, W.; Li, X. MicroRNA and Transcription Factor Mediated Regulatory Network Analysis Reveals Critical Regulators and Regulatory Modules in Myocardial Infarction. PLoS ONE 2015, 10, e0135339. [Google Scholar] [CrossRef]
- Beishline, K.; Azizkhan-Clifford, J. Sp1 and the ‘hallmarks of cancer’. FEBS J. 2015, 282, 224–258. [Google Scholar] [CrossRef]
- Safe, S.; Abbruzzese, J.L.; Abdelrahim, M.; Hedrick, E. Specificity Protein Transcription Factors and Cancer: Opportunities for Drug Development. Cancer Prev. Res. 2018, 11, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Vizcaíno, C.; Mansilla, S.; Portugal, J. Sp1 transcription factor: A long-standing target in cancer chemotherapy. Pharmacol. Ther. 2015, 152, 111–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Genes | p-Value | Annotations |
|---|---|---|
| 30 | 1.07 × 10−6 | (KEGG) 04510: Focal adhesion |
| 19 | 2.04 × 10−6 | (KEGG) 05222: Small cell lung cancer |
| 18 | 4.72 × 10−5 | (KEGG) 04974: Protein digestion and absorption |
| 48 | 6.98 × 10−5 | (KEGG) 05200: Pathways in cancer |
| 35 | 7.63 × 10−5 | (KEGG) 05165: Human papillomavirus infection |
| 16 | 8.83 × 10−5 | (KEGG) 04512: ECM-receptor interaction |
| 28 | 0.000184427 | (KEGG) 04144: Endocytosis |
| 34 | 0.000560778 | (KEGG) 04151: PI3K-Akt signaling pathway |
| 20 | 0.000585526 | (KEGG) 04390: Hippo signaling pathway |
| 13 | 0.00911125 | (KEGG) 05215: Prostate cancer |
| 13 | 0.0112006 | (KEGG) 04933: AGE-RAGE signaling pathway in diabetic complications |
| 12 | 0.0247066 | (KEGG) 05231: Choline metabolism in cancer |
| 18 | 0.0252777 | (KEGG) 04360: Axon guidance |
| 10 | 0.0253268 | (KEGG) 05218: Melanoma |
| 8 | 0.026522 | (KEGG) 04340: Hedgehog signaling pathway |
| 15 | 0.0283549 | (KEGG) 04540: Signaling pathways regulating pluripotency of stem cells |
| 19 | 0.0302379 | (KEGG) 05205: Proteoglycans in cancer |
| 15 | 0.0354345 | (KEGG) 04072: Phospholipase D signaling pathway |
| 11 | 0.0412279 | (KEGG) 04350: TGF-beta signaling pathway |
| 20 | 0.0463112 | (KEGG) 04014: Ras signaling pathway |
| Gene Symbol | Gene Name | Expression in ICC 1 Cancer Tissues (p-Value) | Correlation with ITGA6 (p-Value) | Correlation with ITGB1 (p-Value) | miR-29-3p Binding Site | Reference |
|---|---|---|---|---|---|---|
| SP1 | Sp1 transcription factor | <0.01 | <0.01 | <0.01 | + | [24,25] |
| CREB1 | cAMP responsive element binding protein 1 | <0.01 | <0.01 | <0.01 | − | [25] |
| MAX | MYC associated factor X | <0.01 | <0.01 | 0.011 | − | [25] |
| FHL2 | four and a half LIM domains 2 | <0.01 | <0.01 | <0.01 | − | [25] |
| RFX1 | regulatory factor X, 1 (influences HLA class II expression) | <0.01 | <0.01 | <0.01 | − | [25] |
| HIF1A | hypoxia inducible factor 1, alpha subunit (basic helix-loop-helix transcription factor) | <0.01 | <0.01 | 0.019 | − | [25] |
| TFAP2A | transcription factor AP-2 alpha (activating enhancer binding protein 2 alpha) | <0.01 | N.S | <0.01 | − | [25] |
| CUX1 | cut-like homeobox 1 | <0.01 | <0.01 | N.S | − | [25] |
| USF1 | upstream transcription factor 1 | <0.01 | N.S | <0.01 | − | [25] |
| TFAP2C | transcription factor AP-2 gamma (activating enhancer binding protein 2 gamma) | <0.01 | N.S | N.S | + | [25] |
| FOXM1 | forkhead box M1 | <0.01 | N.S | N.S | − | [26] |
| SPI1 | Spi-1 proto-oncogene | <0.05 | N.S | N.S | − | [25] |
| MXI1 | MAX interactor 1, dimerization protein | N.S 2 | 0.011 | N.S | + | [25] |
| PAX6 | paired box 6 | N.S | <0.01 | <0.01 | − | [25] |
| ESR1 | estrogen receptor 1 | N.S | 0.015 | <0.01 | − | [25] |
| MEF2A | myocyte enhancer factor 2A | N.S | <0.01 | 0.038 | − | [25] |
| HOXD3 | homeobox D3 | N.S | 0.012 | N.S | − | [25] |
| MYC | MYC proto-oncogene, bHLH transcription factor | N.S | N.S | N.S | − | [25] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hozaka, Y.; Seki, N.; Tanaka, T.; Asai, S.; Moriya, S.; Idichi, T.; Wada, M.; Tanoue, K.; Kawasaki, Y.; Mataki, Y.; et al. Molecular Pathogenesis and Regulation of the miR-29-3p-Family: Involvement of ITGA6 and ITGB1 in Intra-Hepatic Cholangiocarcinoma. Cancers 2021, 13, 2804. https://doi.org/10.3390/cancers13112804
Hozaka Y, Seki N, Tanaka T, Asai S, Moriya S, Idichi T, Wada M, Tanoue K, Kawasaki Y, Mataki Y, et al. Molecular Pathogenesis and Regulation of the miR-29-3p-Family: Involvement of ITGA6 and ITGB1 in Intra-Hepatic Cholangiocarcinoma. Cancers. 2021; 13(11):2804. https://doi.org/10.3390/cancers13112804
Chicago/Turabian StyleHozaka, Yuto, Naohiko Seki, Takako Tanaka, Shunichi Asai, Shogo Moriya, Tetsuya Idichi, Masumi Wada, Kiyonori Tanoue, Yota Kawasaki, Yuko Mataki, and et al. 2021. "Molecular Pathogenesis and Regulation of the miR-29-3p-Family: Involvement of ITGA6 and ITGB1 in Intra-Hepatic Cholangiocarcinoma" Cancers 13, no. 11: 2804. https://doi.org/10.3390/cancers13112804
APA StyleHozaka, Y., Seki, N., Tanaka, T., Asai, S., Moriya, S., Idichi, T., Wada, M., Tanoue, K., Kawasaki, Y., Mataki, Y., Kurahara, H., & Ohtsuka, T. (2021). Molecular Pathogenesis and Regulation of the miR-29-3p-Family: Involvement of ITGA6 and ITGB1 in Intra-Hepatic Cholangiocarcinoma. Cancers, 13(11), 2804. https://doi.org/10.3390/cancers13112804