
Anti-Angiogenic Treatments Interact with Steroid Secretion in Inflammatory Breast Cancer Triple Negative Cell Lines



Abstract
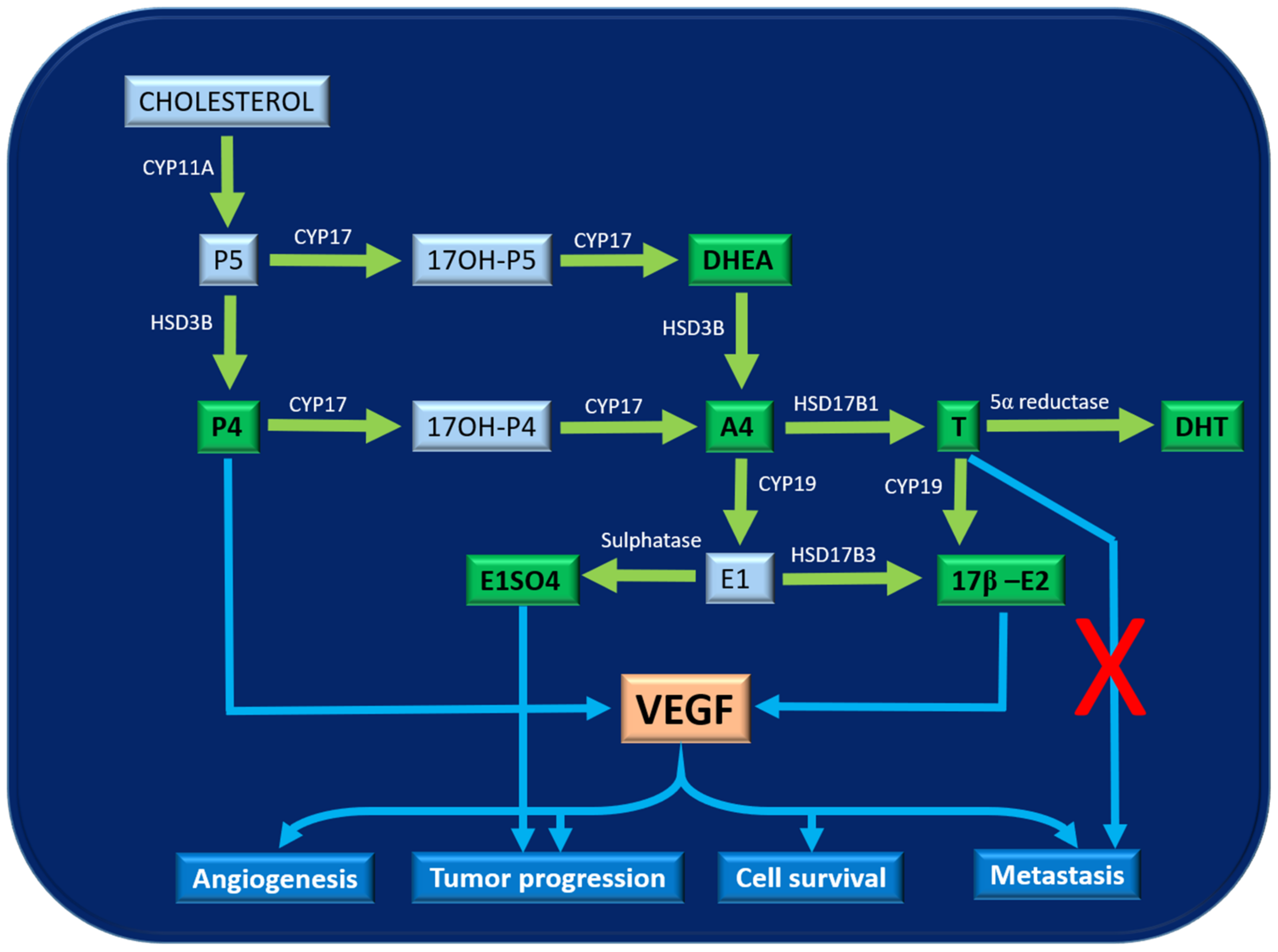
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Line Culture
2.2. In Vitro Treatments
2.3. Cell Viability Assay (MTS Assay)
2.4. Tube Formation Assay
2.5. Experimental Animals and Treatments
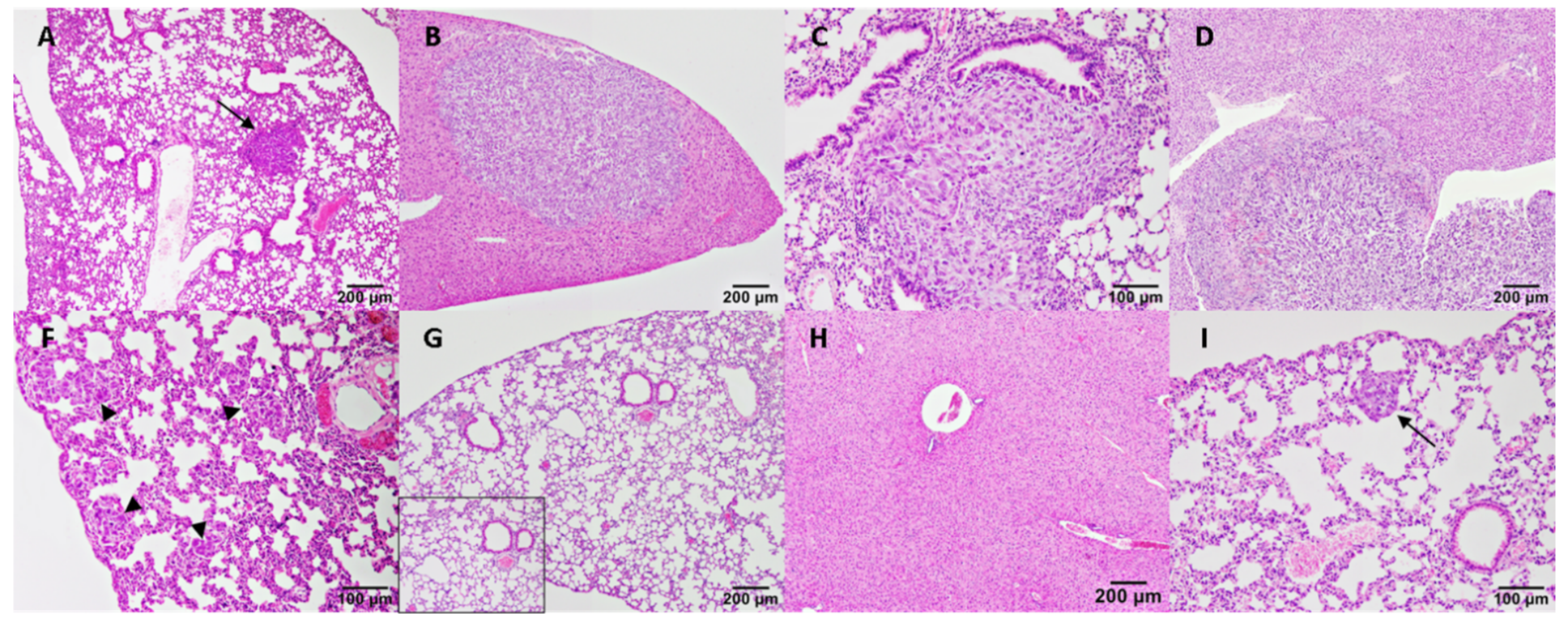
2.6. Histological Examination
2.7. Steroid Determinations in Culture Media, Serum and Tumour Homogenates
2.8. Statistics
3. Results
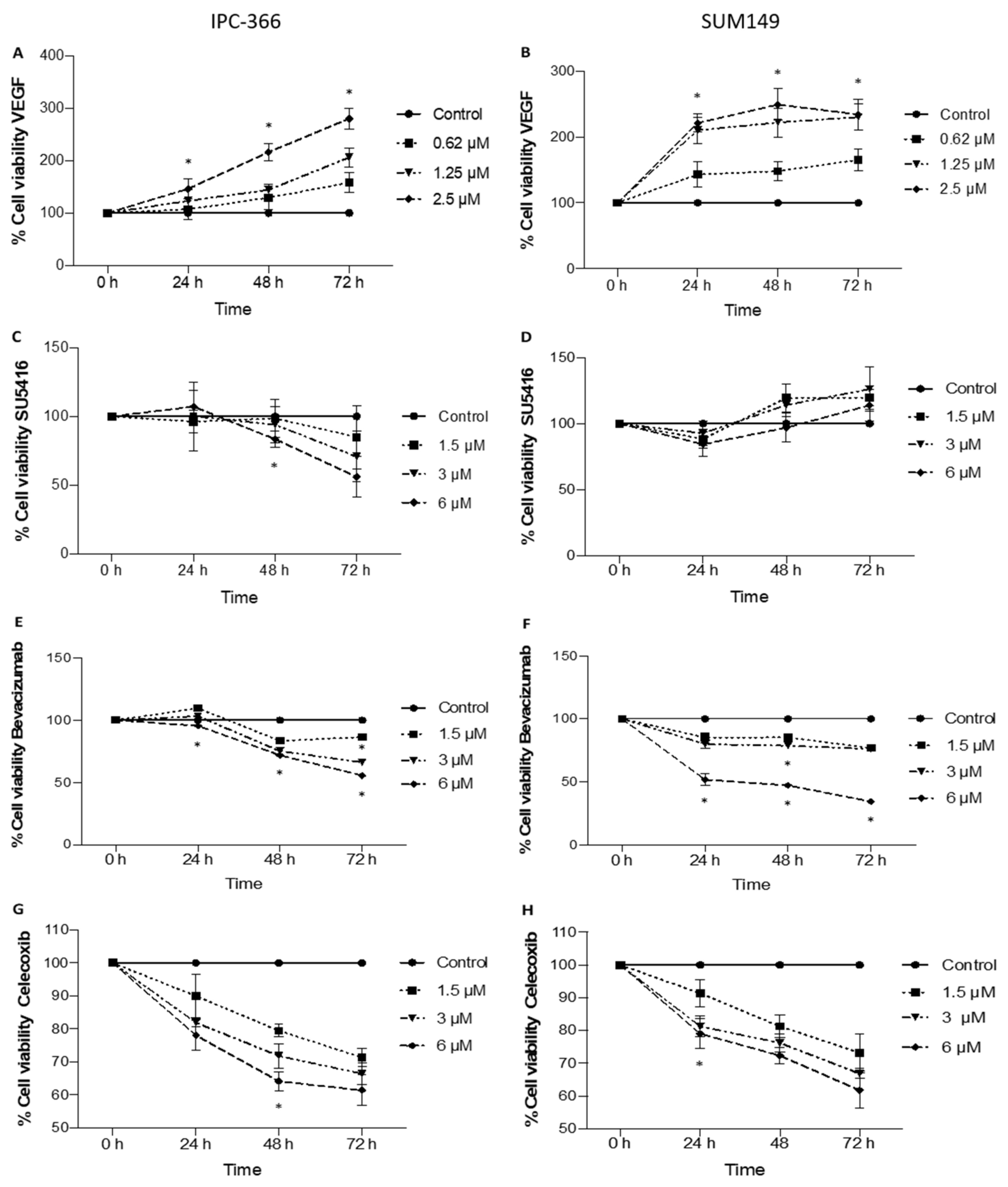
3.1. Anti-Proliferative Effect on In Vitro Cell Viability with the Addition of Anti-Angiogenic Treatments
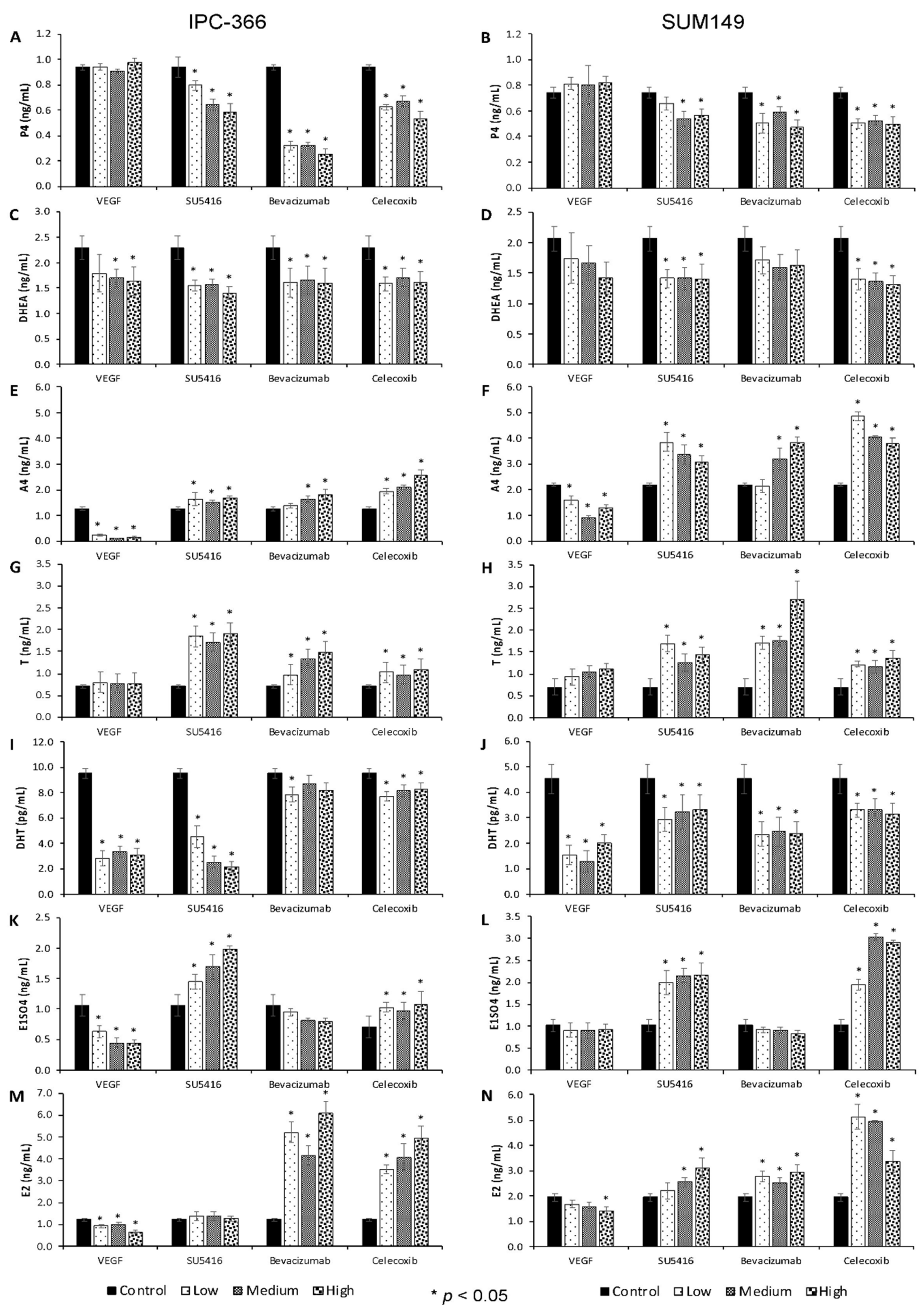
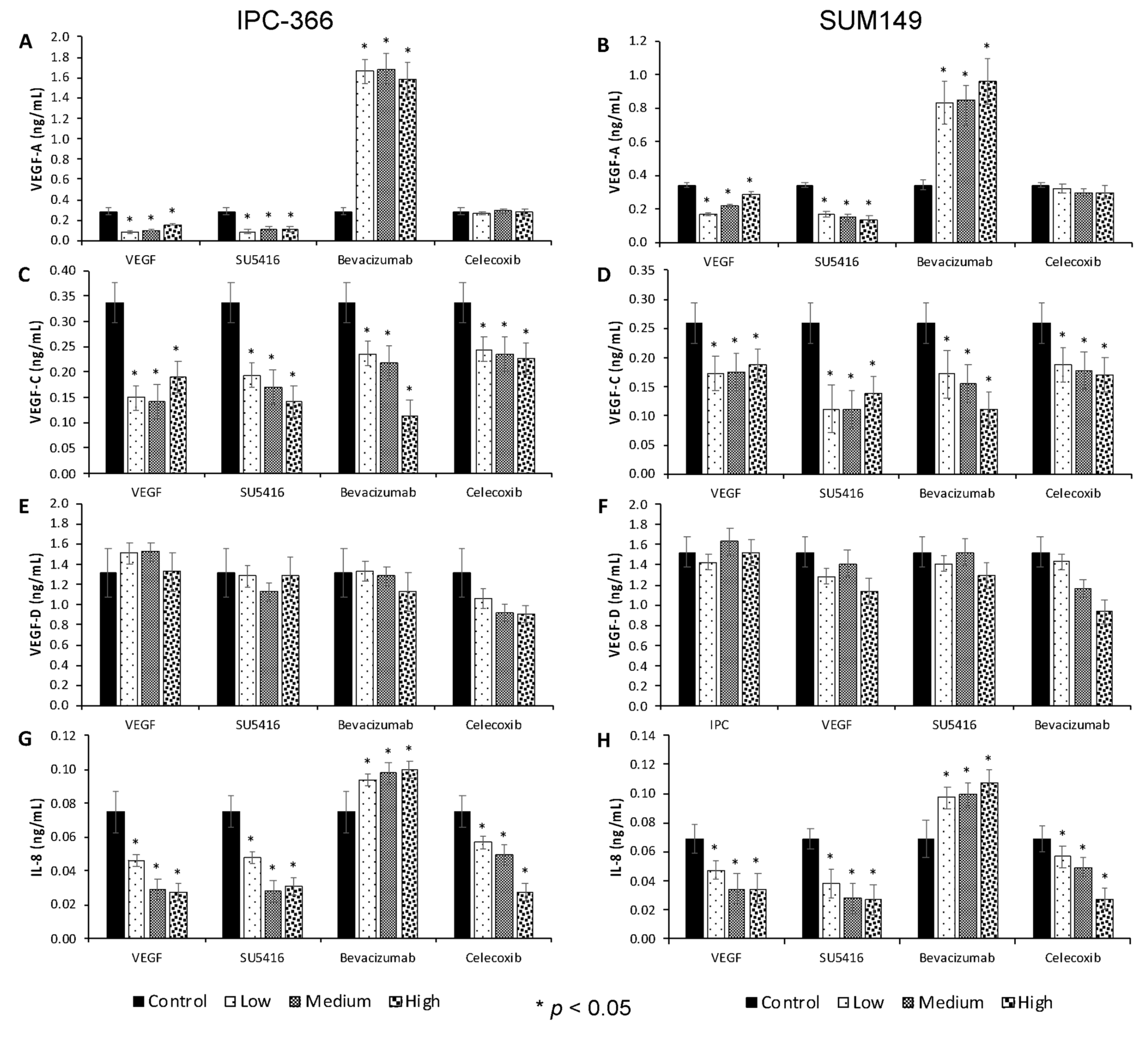
3.2. Anti-Angiogenic Treatments Alter In Vitro Steroid Hormone Secretion
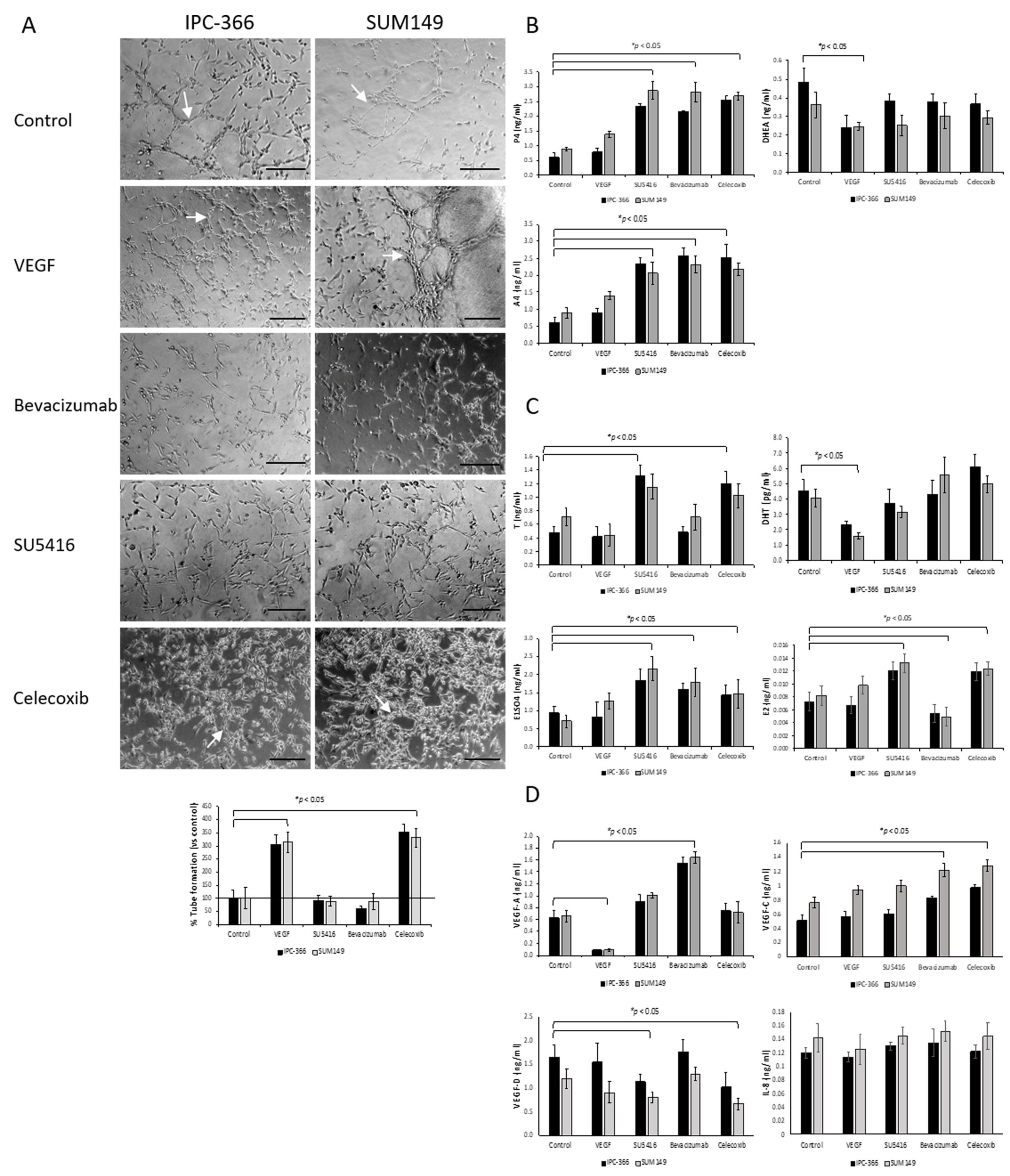
3.3. Effect of Anti-Angiogenic Compounds and VEGF in Tube Formation
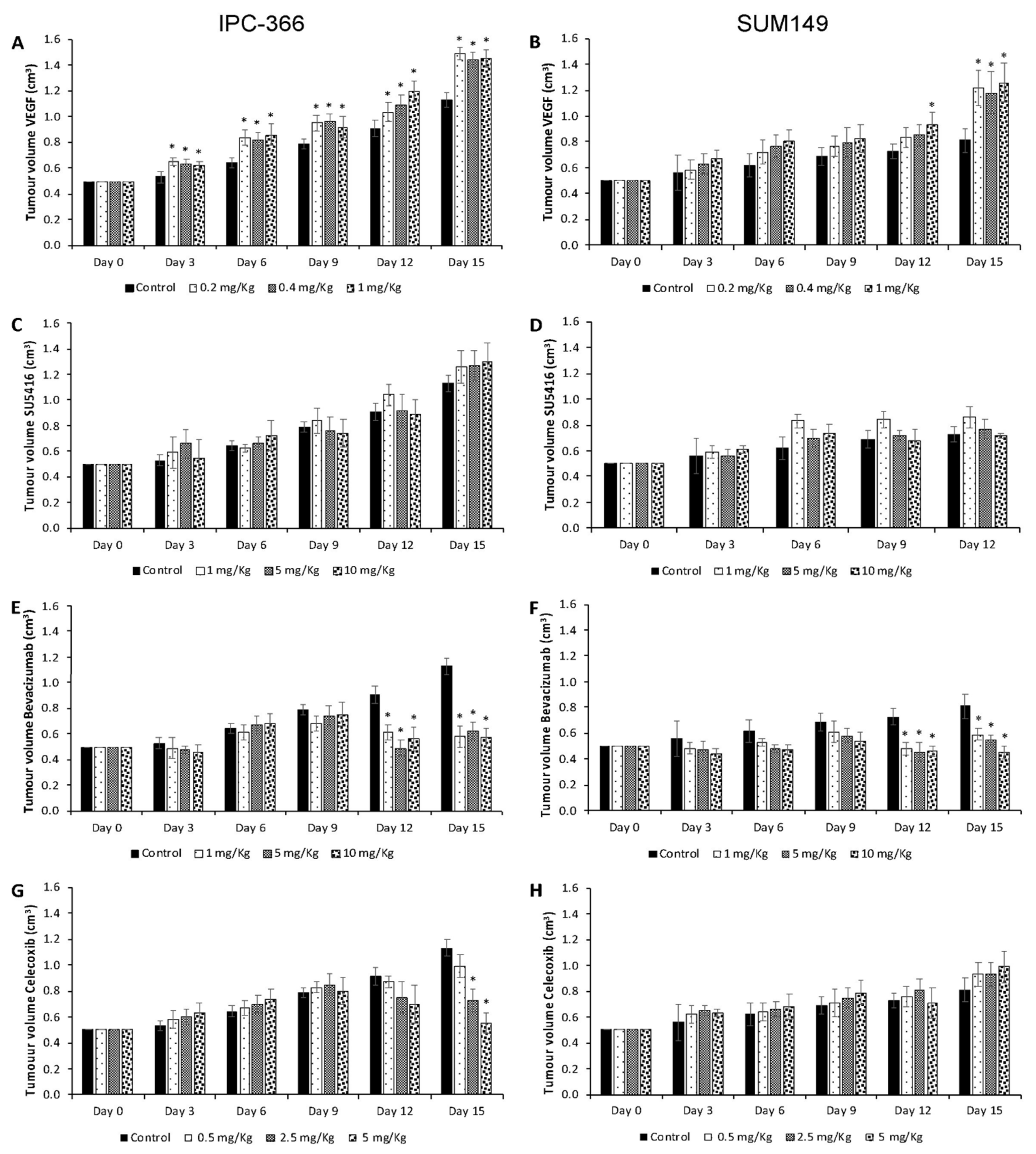
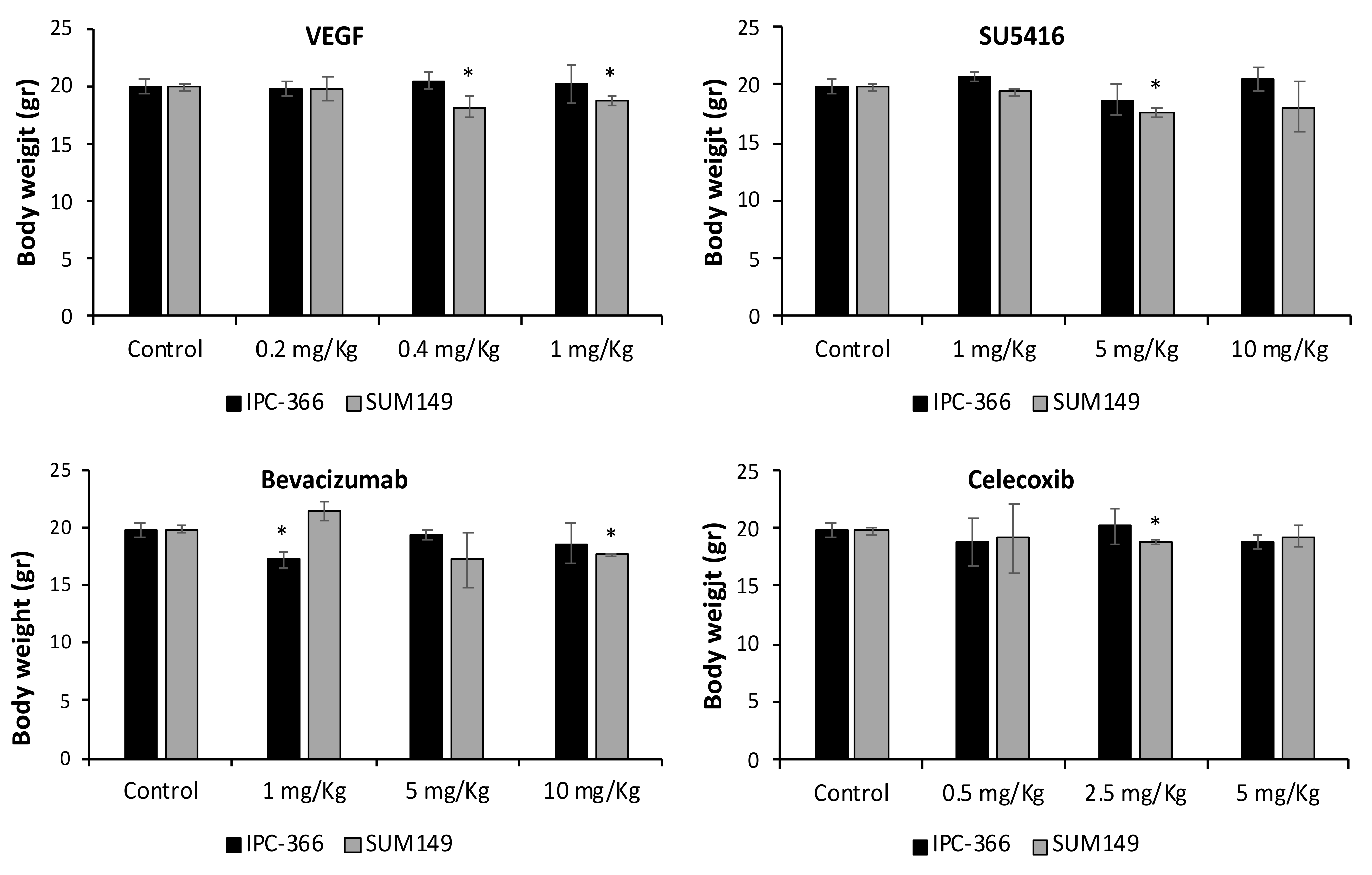
3.4. VEGF, SU5416, Bevacizumab and Celecoxib Effects on Tumour Progression and Metastasis
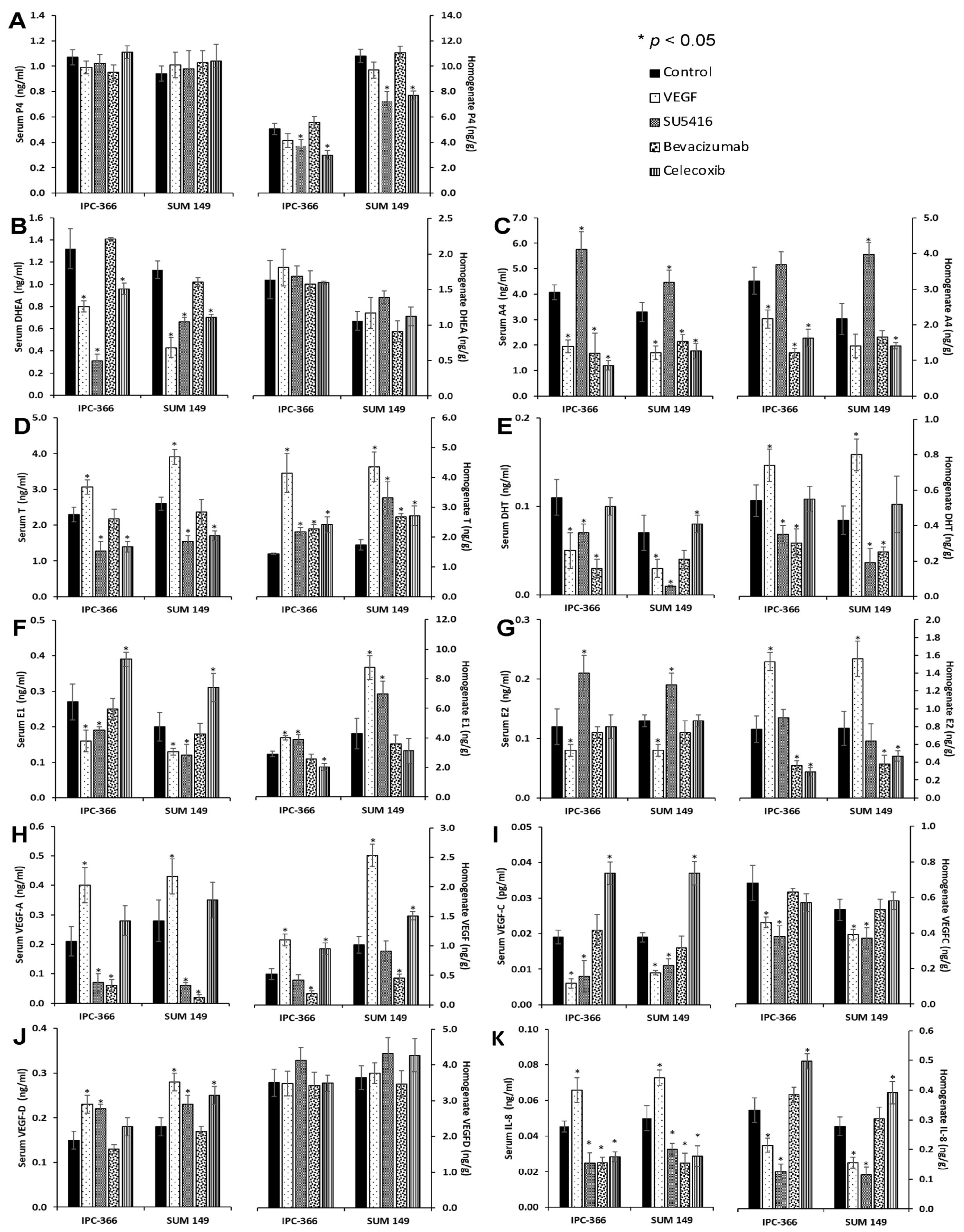
3.5. Steroid Determinations in Tumour Homogenates and Serum Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abul-Hajj, Y. Aromatization of androgens by human breast cancer. Steroids 1979, 33, 205–222. [Google Scholar] [CrossRef]
- Blankenstein, M.; Van De Ven, J.; Maitimu-Smeele, I.; Donker, G.; De Jong, P.; Daroszewski, J.; Szymczak, J.; Milewicz, A.; Thijssen, J. Intratumoral levels of estrogens in breast cancer. J. Steroid Biochem. Mol. Biol. 1999, 69, 293–297. [Google Scholar] [CrossRef]
- Peña, L.; Silván, G.; Pérez-Alenza, M.D.; Nieto, A.; Illera, J.C.; Peña, L.; Silván, G.; Pérez-Alenza, M.D.; Nieto, A.; Illera, J.C. Steroid hormone profile of canine inflammatory mammary carcinoma: A preliminary study. J. Steroid Biochem. Mol. Biol. 2003, 84, 211–216. [Google Scholar] [CrossRef]
- McNamara, K.M.; Sasano, H. The intracrinology of breast cancer. J. Steroid Biochem. Mol. Biol. 2015, 145, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Capper, C.P.; Rae, J.M.; Auchus, R.J.; Capper, C.P.; Rae, J.M.; Auchus, R.J. The Metabolism, Analysis, and Targeting of Steroid Hormones in Breast and Prostate Cancer. Horm. Cancer 2016, 7, 149–164. [Google Scholar] [CrossRef] [Green Version]
- Yager, J.D.; Davidson, N.E. Estrogen Carcinogenesis in Breast Cancer. N. Engl. J. Med. 2006, 354, 270–282. [Google Scholar] [CrossRef] [Green Version]
- Chang, M.-S. Dual roles of estrogen metabolism in mammary carcinogenesis. BMB Rep. 2011, 44, 423–434. [Google Scholar] [CrossRef]
- Fuhrman, B.J.; Schairer, C.; Gail, M.H.; Boyd-Morin, J.; Xu, X.; Sue, L.Y.; Buys, S.S.; Isaacs, C.; Keefer, L.K.; Veenstra, T.D.; et al. Estrogen Metabolism and Risk of Breast Cancer in Postmenopausal Women. J. Natl. Cancer Inst. 2012, 104, 326–339. [Google Scholar] [CrossRef]
- Falk, R.T.; A Brinton, L.; Dorgan, J.F.; Fuhrman, B.J.; Veenstra, T.D.; Xu, X.; Gierach, G.L. Relationship of serum estrogens and estrogen metabolites to postmenopausal breast cancer risk: A nested case-control study. Breast Cancer Res. 2013, 15, R34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jerry, D.J.; Shull, J.D.; Hadsell, D.L.; Rijnkels, M.; Dunphy, K.A.; Schneider, S.S.; Vandenberg, L.N.; Majhi, P.D.; Byrne, C.; Trentham-Dietz, A. Genetic variation in sensitivity to estrogens and breast cancer risk. Mamm. Genome 2018, 29, 24–37. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.B.; E Hankinson, S. Endogenous estrogens and the risk of breast, endometrial, and ovarian cancers. Steroids 2015, 99, 8–10. [Google Scholar] [CrossRef]
- Hilborn, E.; Stål, O.; Jansson, A. Estrogen and androgen-converting enzymes 17β-hydroxysteroid dehydrogenase and their involvement in cancer: With a special focus on 17β-hydroxysteroid dehydrogenase type 1, 2, and breast cancer. Oncotarget 2017, 8, 30552–30562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baglietto, L.; Severi, G.; English, D.; Krishnan, K.; Hopper, J.L.; McLean, C.; Morris, H.A.; Tilley, W.; Giles, G. Circulating Steroid Hormone Levels and Risk of Breast Cancer for Postmenopausal Women. Cancer Epidemiol. Biomarkers Prev. 2010, 19, 492–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeleniuch-Jacquotte, A.; Afanasyeva, Y.; Kaaks, R.; Rinaldi, S.; Scarmo, S.; Liu, M.; A Arslan, A.; Toniolo, P.; E Shore, R.; Koenig, K.L. Premenopausal serum androgens and breast cancer risk: A nested case-control study. Breast Cancer Res. 2012, 14, R32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caceres, S.; Peña, L.; Silvan, G.; Illera, M.J.; Woodward, W.; Reuben, J.M.; Illera, J.C. Steroid Tumor Environment in Male and Female Mice Model of Canine and Human Inflammatory Breast Cancer. BioMed Res. Int. 2016, 2016, 8909878. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madu, C.O.; Wang, S.; Madu, C.O.; Lu, Y. Angiogenesis in Breast Cancer Progression, Diagnosis, and Treatment. J. Cancer 2020, 11, 4474–4494. [Google Scholar] [CrossRef]
- Goussia, A.; Simou, N.; Zagouri, F.; Manousou, K.; Lazaridis, G.; Gogas, H.; Koutras, A.; Sotiropoulou, M.; Pentheroudakis, G.; Bafaloukos, D.; et al. Associations of angiogenesis-related proteins with specific prognostic factors, breast cancer subtypes and survival outcome in early-stage breast cancer patients. A Hellenic Cooperative Oncology Group (HeCOG) trial. PLoS ONE 2018, 13, e0200302. [Google Scholar] [CrossRef]
- Ramjiawan, R.R.; Griffioen, A.W.; Duda, D.G. Anti-angiogenesis for cancer revisited: Is there a role for combinations with immunotherapy? Angiogenesis 2017, 20, 185–204. [Google Scholar] [CrossRef]
- Hyder, S.M. Sex-steroid regulation of vascular endothelial growth factor in breast cancer. Endocrine-Related Cancer 2006, 13, 667–687. [Google Scholar] [CrossRef]
- Botelho, M.C.; Soares, R.; Alves, H. Progesterone in Breast Cancer Angiogenesis. SM J. Reprod. Heal. Infertil. 2015, 1, 1001. [Google Scholar]
- Petrovic, N. Targeting Angiogenesis in Cancer Treatments: Where do we Stand? J. Pharm. Pharm. Sci. 2016, 19, 226–238. [Google Scholar] [CrossRef] [PubMed]
- Ranieri, G.; Patruno, R.; Ruggieri, E.; Montemurro, S.; Valerio, P.; Ribatti, D. Vascular Endothelial Growth Factor (VEGF) as a Target of Bevacizumab in Cancer: From the Biology to the Clinic. Curr. Med. Chem. 2006, 13, 1845–1857. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Yan, H.; Zhao, P.; Yang, Y.; Cao, B. Efficacy and Safety of Bevacizumab Combined with Chemotherapy for Managing Metastatic Breast Cancer: A Meta-Analysis of Randomized Controlled Trials. Sci. Rep. 2015, 5, 15746. [Google Scholar] [CrossRef] [Green Version]
- Grailer, J.J.; Steeber, U.A. Vascular Endothelial Growth Factor Receptor Inhibitor SU5416 Suppresses Lymphocyte Generation and Immune Responses in Mice by Increasing Plasma Corticosterone. PLoS ONE 2013, 8, e75390. [Google Scholar] [CrossRef] [Green Version]
- Vajkoczy, P.; Menger, M.D.; Vollmar, B.; Schilling, L.; Schmiedek, P.; Hirth, K.P.; Ullrich, A.; Fong, T.T. Inhibition of Tumor Growth, Angiogenesis, and Microcirculation by the Novel Flk-1 Inhibitor SU5416 as Assessed by Intravital Multi-fluorescence Videomicroscopy. Neoplasia 1999, 1, 31–41. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, A.; Padhani, A.; Hayes, C.; Kakkar, A.J.; Leach, M.; Trigo, J.M.; Scurr, M.; Raynaud, F.; Phillips, S.; Aherne, W.; et al. A Phase I study of the angiogenesis inhibitor SU5416 (semaxanib) in solid tumours, incorporating dynamic contrast MR pharmacodynamic end points. Br. J. Cancer 2005, 93, 876–883. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, K.; Bustin, S.; Ogunkolade, B.; Khalaf, S.; Laban, C.; McVittie, C.; Carpenter, R.; Jenkins, P. Cyclo-oxygenase-2 (COX-2) mRNA expression and hormone receptor status in breast cancer. Eur. J. Surg. Oncol. (EJSO) 2006, 32, 707–709. [Google Scholar] [CrossRef]
- Hamy, A.-S.; Tury, S.; Wang, X.; Gao, J.; Pierga, J.-Y.; Giacchetti, S.; Brain, E.; Pistilli, B.; Marty, M.; Espié, M.; et al. Celecoxib With Neoadjuvant Chemotherapy for Breast Cancer Might Worsen Outcomes Differentially by COX-2 Expression and ER Status: Exploratory Analysis of the REMAGUS02 Trial. J. Clin. Oncol. 2019, 37, 624–635. [Google Scholar] [CrossRef]
- Harris, R.E. Cyclooxygenase-2 (cox-2) blockade in the chemoprevention of cancers of the colon, breast, prostate, and lung. Inflammopharmacology 2009, 17, 55–67. [Google Scholar] [CrossRef]
- Stasinopoulos, I.; Shah, T.; Penet, M.-F.; Krishnamachary, B.; Bhujwalla, Z.M. COX-2 in cancer: Gordian knot or Achilles heel? Front. Pharmacol. 2013, 4, 34. [Google Scholar] [CrossRef] [Green Version]
- Vosooghi, M.; Amini, M. The discovery and development of cyclooxygenase-2 inhibitors as potential anticancer therapies. Expert Opin. Drug Discov. 2014, 9, 255–267. [Google Scholar] [CrossRef]
- Suri, A.; Sheng, X.; Schuler, K.M.; Zhong, Y.; Han, X.; Jones, H.M.; Gehrig, P.A.; Zhou, C.; Bae-Jump, V.L. The effect of celecoxib on tumor growth in ovarian cancer cells and a genetically engineered mouse model of serous ovarian cancer. Oncotarget 2016, 7, 39582–39594. [Google Scholar] [CrossRef]
- Wang, Q.; Wu, X. Primary and acquired resistance to PD-1/PD-L1 blockade in cancer treatment. Int. Immunopharmacol. 2017, 46, 210–219. [Google Scholar] [CrossRef]
- Wang, X.; Semba, T.; Phi, L.T.H.; Chainitikun, S.; Iwase, T.; Lim, B.; Ueno, N.T. Targeting Signaling Pathways in Inflammatory Breast Cancer. Cancers 2020, 12, 2479. [Google Scholar] [CrossRef]
- Li, J.; Hao, Q.; Cao, W.; Vadgama, J.V.; Wu, Y. Celecoxib in breast cancer prevention and therapy. Cancer Manag. Res. 2018, 10, 4653–4667. [Google Scholar] [CrossRef] [Green Version]
- Yamauchi, H.; Ueno, N.T. Targeted therapy in inflammatory breast cancer. Cancer 2010, 116, 2758–2759. [Google Scholar] [CrossRef]
- Van Der Auwera, I.; Van Laere, S.; Eynden, G.G.V.D.; Benoy, I.; Van Dam, P.; Colpaert, C.G.; Fox, S.; Turley, H.; Harris, A.; Van Marck, E.A.; et al. Increased Angiogenesis and Lymphangiogenesis in Inflammatory versus Noninflammatory Breast Cancer by Real-Time Reverse Transcriptase-PCR Gene Expression Quantification. Clin. Cancer Res. 2004, 10, 7965–7971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arias-Pulido, H.; Chaher, N.; Gong, Y.; Qualls, C.; Vargas, J.; Royce, M. Tumor stromal vascular endothelial growth factor A is predictive of poor outcome in inflammatory breast cancer. BMC Cancer 2012, 12, 298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clemente, M.; Pérez-Alenza, M.D.; Illera, J.C.; Peña, L. Histological, Immunohistological, and Ultrastructural Description of Vasculogenic Mimicry in Canine Mammary Cancer. Veter. Pathol. 2009, 47, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Clemente, M.; Pérez-Alenza, M.; Pena, L. Metastasis of Canine Inflammatory versus Non-Inflammatory Mammary Tumours. J. Comp. Pathol. 2010, 143, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Caceres, S.; Peña, L.; De Andres, P.J.; Illera, M.J.; Lopez, M.S.; Woodward, W.; Reuben, J.M.; Illera, J.C. Establishment and Characterization of a New Cell Line of Canine Inflammatory Mammary Cancer: IPC-366. PLoS ONE 2015, 10, e0122277. [Google Scholar] [CrossRef] [Green Version]
- Illera, J.C.; Caceres, S.; Peña, L.; De Andres, P.J.; Monsalve, B.; Illera, M.J.; Woodward, W.; Reuben, J.M.; Silvan, G. Steroid hormone secretion in inflammatory breast cancer cell lines. Horm. Mol. Biol. Clin. Investig. 2015, 24, 137–145. [Google Scholar] [CrossRef]
- Millanta, F.; Caneschi, V.; Ressel, L.; Citi, S.; Poli, A. Expression of Vascular Endothelial Growth Factor in Canine Inflammatory and Non-inflammatory Mammary Carcinoma. J. Comp. Pathol. 2010, 142, 36–42. [Google Scholar] [CrossRef]
- Guo, S.; Lok, J.; Liu, Y.; Hayakawa, K.; Leung, W.; Xing, C.; Ji, X.; Lo, E.H. Assays to Examine Endothelial Cell Migration, Tube Formation, and Gene Expression Profiles. Methods Mol. Biol. 2014, 1135, 393–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneko, M.; Kaneko, S.; Suzuki, K. Prolonged low-dose administration of the cyclooxygenase-2 inhibitor celecoxib enhances the antitumor activity of irinotecan against neuroblastoma xenografts. Cancer Sci. 2009, 100, 2193–2201. [Google Scholar] [CrossRef] [PubMed]
- Adamcic, U.; Skowronski, K.; Peters, C.; Morrison, J.; Coomber, B.L. The Effect of Bevacizumab on Human Malignant Melanoma Cells with Functional VEGF/VEGFR2 Autocrine and Intracrine Signaling Loops. Neoplasia 2012, 14, 612–623. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Fu, X.; Jiang, C.; Yu, L.; Wang, M.; Han, W.; Liu, L.; Wang, J. Bone marrow mononuclear cell transplantation promotes therapeutic angiogenesis via upregulation of the VEGF–VEGFR2 signaling pathway in a rat model of vascular dementia. Behav. Brain Res. 2014, 265, 171–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.-X.; Li, X.; Liu, W.-D.; Liu, X.-Z.; Wu, S.-J.; Hu, X.-H. Inhibition of Growth and Metastasis of Tumor in Nude Mice after Intraperitoneal Injection of Bevacizumab. Orthop. Surg. 2016, 8, 234–240. [Google Scholar] [CrossRef] [Green Version]
- Hather, G.; Liu, R.; Bandi, S.; Mettetal, J.; Manfredi, M.; Shyu, W.-C.; Donelan, J.; Chakravarty, A. Growth Rate Analysis and Efficient Experimental Design for Tumor Xenograft Studies. Cancer Inform. 2014, 13s4, 65–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Queiroga, F.; Pérez-Alenza, M.; Silvan, G.; Peña, L.; Lopes, C.; Illera, J. Role of steroid hormones and prolactin in canine mammary cancer. J. Steroid Biochem. Mol. Biol. 2005, 94, 181–187. [Google Scholar] [CrossRef]
- Ebos, J.M.L.; Kerbel, R.S. Antiangiogenic therapy: Impact on invasion, disease progression, and metastasis. Nat. Rev. Clin. Oncol. 2011, 8, 210–221. [Google Scholar] [CrossRef] [Green Version]
- Palazzo, A.; Dellapasqua, S.; Munzone, E.; Bagnardi, V.; Mazza, M.; Cancello, G.; Ghisini, R.; Iorfida, M.; Montagna, E.; Goldhirsch, A.; et al. Phase II Trial of Bevacizumab Plus Weekly Paclitaxel, Carboplatin, and Metronomic Cyclophosphamide With or Without Trastuzumab and Endocrine Therapy as Preoperative Treatment of Inflammatory Breast Cancer. Clin. Breast Cancer 2018, 18, 328–335. [Google Scholar] [CrossRef]
- Basu, G.D.; Pathangey, L.B.; Tinder, T.L.; Gendler, S.J.; Mukherjee, P.; Basu, G.D.; Pathangey, L.B.; Tinder, T.L.; Gendler, S.J.; Mukherjee, P. Mechanisms underlying the growth inhibitory effects of the cyclo-oxygenase-2 inhibitor celecoxib in human breast cancer cells. Breast Cancer Res. 2005, 7, R422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emlet, D.R.; Brown, K.A.; Kociban, D.L.; Pollice, A.A.; Smith, C.A.; Ong, B.B.L.; Shackney, S.E. Response to trastuzumab, erlotinib, and bevacizumab, alone and in combination, is correlated with the level of human epidermal growth factor receptor-2 expression in human breast cancer cell lines. Mol. Cancer Ther. 2007, 6, 2664–2674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higgins, B.; Kolinsky, K.; Linn, M.; Adames, V.; Zhang, Y.E.; Moisa, C.; Dugan, U.; Heimbrook, D.; Packman, K. Antitumor activity of capecitabine and bevacizumab combination in a human estrogen receptor-negative breast adenocarcinoma xeno-graft model. Anticancer Res 2007, 27, 2279–2287. [Google Scholar] [PubMed]
- Gui, X.; Li, H.; Song, G.; Shao, B.; Jiang, H. Long term use of bevacizumab in the treatment of triple negative breast cancer with giant tumor in chest wall. Medicine 2018, 97, e13410. [Google Scholar] [CrossRef]
- Duggan, C.; Stanczyk, F.; Campbell, K.; Neuhouser, M.L.; Baumgartner, R.N.; Baumgartner, K.B.; Bernstein, L.; Ballard, R.; McTiernan, A. Associations of sex steroid hormones with mortality in women with breast cancer. Breast Cancer Res. Treat. 2016, 155, 559–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kensler, K.H.; Eliassen, A.H.; Rosner, B.A.; Hankinson, S.E.; Brown, M.; Tamimi, R.M. Pre-diagnostic sex hormone levels and survival among breast cancer patients. Breast Cancer Res. Treat. 2019, 174, 749–758. [Google Scholar] [CrossRef]
- Wu, J.; Richer, J.; Horwitz, K.B.; Hyder, S.M. Progestin-Dependent Induction of Vascular Endothelial Growth Factor in Human Breast Cancer Cells. Cancer Res. 2004, 64, 2238–2244. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Zvelebil, M.; Furet, P.; Mueller-Vieira, U.; Evans, D.B.; Dowsett, M.; Martin, L.-A. The Vascular Endothelial Growth Factor Receptor Inhibitor PTK787/ZK222584 Inhibits Aromatase. Cancer Res. 2009, 69, 4716–4723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- E Bachelder, R.; Crago, A.; Chung, J.; A Wendt, M.; Shaw, L.M.; Robinson, G.; Mercurio, A.M. Vascular endothelial growth factor is an autocrine survival factor for neuropilin-expressing breast carcinoma cells. Cancer Res. 2001, 61, 5736–5740. [Google Scholar]
- Harmey, J.H.; Bouchier-Hayes, D. Vascular endothelial growth factor (VEGF), a survival factor for tumour cells: Implications for anti-angiogenic therapy. BioEssays 2002, 24, 280–283. [Google Scholar] [CrossRef]
- Bates, R.C.; Goldsmith, J.D.; Bachelder, R.E.; Brown, C.; Shibuya, M.; Oettgen, P.; Mercurio, A.M. Flt-1-Dependent Survival Characterizes the Epithelial-Mesenchymal Transition of Colonic Organoids. Curr. Biol. 2003, 13, 1721–1727. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Xiong, W.; Xiong, Y.; Liu, H.; Liu, Y. 17 β-Estradiol promotes vascular endothelial growth factor expression via the Wnt/β-catenin pathway during the pathogenesis of endometriosis. Mol. Hum. Reprod. 2016, 22, 526–535. [Google Scholar] [CrossRef] [Green Version]
- Sasich, L.D.; Sukkari, S.R. The US FDAs withdrawal of the breast cancer indication for Avastin (bevacizumab). Saudi Pharm. J. 2012, 20, 381–385. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.J.; Yao, P.-L.; Yuan, A.; Hong, T.-M.; Shun, C.-T.; Kuo, M.-L.; Lee, Y.-C.; Yang, P.-C. Up-regulation of tumor interleukin-8 expression by infiltrating macrophages: Its correlation with tumor angiogenesis and patient survival in non-small cell lung cancer. Clin. Cancer Res. 2003, 9, 729–737. [Google Scholar]
- Al-Khalaf, H.H.; Al-Harbi, B.; Al-Sayed, A.; Arafah, M.; Tulbah, A.; Jarman, A.; Al-Mohanna, F.; Aboussekhra, A. Interleukin-8 Activates Breast Cancer-Associated Adipocytes and Promotes Their Angiogenesis- and Tumorigenesis-Promoting Effects. Mol. Cell. Biol. 2019, 39, e00332-18. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.; Galisteo, R.; Gutkind, J.S. CXCL8/IL8 Stimulates Vascular Endothelial Growth Factor (VEGF) Expression and the Autocrine Activation of VEGFR2 in Endothelial Cells by Activating NFκB through the CBM (Carma3/Bcl10/Malt1) Complex. J. Biol. Chem. 2009, 284, 6038–6042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Kang, K.-T. Differential Angiogenic Responses of Human Endothelial Colony-Forming Cells to Different Molecular Subtypes of Breast Cancer Cells. J. Lipid Atheroscler. 2021, 10, 111–122. [Google Scholar] [CrossRef]
- Iii, R.A.F.; Faibish, M.; Shao, R. A Matrigel-Based Tube Formation Assay to Assess the Vasculogenic Activity of Tumor Cells. J. Vis. Exp. 2011, 55, e3040. [Google Scholar] [CrossRef]
- Lansink, M.; Koolwijk, P.; van Hinsbergh, V.; Kooistra, T. Effect of steroid hormones and retinoids on the formation of capil-lary-like tubular structures of human microvascular endothelial cells in fibrin matrices is related to urokinase expression. Blood 1998, 92, 927–938. [Google Scholar] [CrossRef]
- Liu, P.; Li, X.; Song, F.; Li, P.; Wei, J.; Yan, Q.; Xu, X.; Yang, J.; Li, C.; Fu, X. Testosterone promotes tube formation of endothelial cells isolated from veins via activation of Smad1 protein. Mol. Cell. Endocrinol. 2017, 446, 21–31. [Google Scholar] [CrossRef]
- Stacker, S.A.; Achen, M.G. Emerging Roles for VEGF-D in Human Disease. Biomolecules 2018, 8, 1. [Google Scholar] [CrossRef] [Green Version]
- Haddad, J.J. The immunopharmacologic potential of Semaxanib and new generation directed therapeutic drugs: Receptor tyrosine kinase regulation with anti-tumorigenensis/angiogenesis properties. Saudi Pharm. J. 2012, 20, 103–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, L.A.; Brekken, R.A. The VEGF family in cancer and antibody-based strategies for their inhibition. mAbs 2010, 2, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Venturelli, E.; Orenti, A.; Fabricio, A.S.C.; Garrone, G.; Agresti, R.; Paolini, B.; Bonini, C.; Gion, M.; Berrino, F.; Desmedt, C.; et al. Observational study on the prognostic value of testosterone and adiposity in postmenopausal estrogen receptor positive breast cancer patients. BMC Cancer 2018, 18, 651. [Google Scholar] [CrossRef] [Green Version]
- Somboonporn, W.; Davis, S.R. Testosterone Effects on the Breast: Implications for Testosterone Therapy for Women. Endocr. Rev. 2004, 25, 374–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Schoultz, B. Androgens and the breast. Matur. 2007, 57, 47–49. [Google Scholar] [CrossRef] [PubMed]
- Caceres, S.; Monsalve, B.; Peña, L.; De Andres, P.J.; Alonso-Diez, A.; Illera, M.J.; Woodward, W.; Reuben, J.M.; Silvan, G.; Illera, J.C. In vitro and in vivo effect of flutamide on steroid hormone secretion in canine and human inflammatory breast cancer cell lines. Veter. Comp. Oncol. 2017, 16, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, V.N.; Børresen-Dale, A.L. Molecular epidemiology of breast cancer: Genetic variation in steroid hormone metabolism. Mutat. Res. Mutat. Res. 2000, 462, 323–333. [Google Scholar] [CrossRef]
- Eibl, G.; Takata, Y.; Boros, L.G.; Liu, J.; Okada, Y.; A Reber, H.; Hines, O.J. Growth stimulation of COX-2-negative pancreatic cancer by a selective COX-2 inhibitor. Cancer Res. 2005, 65, 982–990. [Google Scholar] [PubMed]
- Xu, K.; Gao, H.; Shu, H.-K.G. Celecoxib Can Induce Vascular Endothelial Growth Factor Expression and Tumor Angiogenesis. Mol. Cancer Ther. 2011, 10, 138–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueno, T.; Chow, L.; Toi, M. Increases in circulating VEGF levels during COX-2 inhibitor treatment in breast cancer patients. Biomed. Pharmacother. 2006, 60, 277–279. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhang, Y.; Feurino, L.W.; Wang, H.; Fisher, W.E.; Brunicardi, F.C.; Chen, C.; Yao, Q. Interleukin-8 increases vascular endothelial growth factor and neuropilin expression and stimulates ERK activation in human pancreatic cancer. Cancer Sci. 2008, 99, 733–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dore-Savard, L.; Lee, E.; Kakkad, S.; Popel, A.S.; Bhujwalla, Z.M. The Angiogenic Secretome in VEGF overexpressing Breast Cancer Xenografts. Sci. Rep. 2016, 6, 39460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, P.B.; Kuperwasser, C. Contributions of estrogen to ER-negative breast tumor growth. J. Steroid Biochem. Mol. Biol. 2006, 102, 71–78. [Google Scholar] [CrossRef]
- Zhang, H.-T.; A E Scott, P.; Morbidelli, L.; Peak, S.; Moore, J.; Turley, H.; Harris, A.; Ziche, M.; Bicknell, R. The 121 amino acid isoform of vascular endothelial growth factor is more strongly tumorigenic than other splice variants in vivo. Br. J. Cancer 2000, 83, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Garvin, S.; Nilsson, U.W.; Dabrosin, C. Effects of oestradiol and tamoxifen on VEGF, soluble VEGFR-1, and VEGFR-2 in breast cancer and endothelial cells. Br. J. Cancer 2005, 93, 1005–1010. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibody | Abbreviation | Reference |
|---|---|---|
| Progesterone | P4 | C6E91 |
| Androstenedione | A4 | C9111 |
| Oestrone sulphate | E1SO4 | R522-2 |
| Testosterone | T | R156 |
| 17β-oestradiol | E2 | C6E91 |
| Dehydroepiandrostenedione | DHEA | DEH3344 |
| Dihydrotestosterone | DHT | DE2330 |
| Vascular endothelial growth factor A | VEGF-A | RAB0107-1KT |
| Vascular endothelial growth factor C | VEGF-C | RAB0313-1KT |
| Vascular endothelial growth factor D | VEGF-D | RAB0390-1KT |
| Interleukin-8 | IL-8/CXCL8 | RAB0319-1KT |
| IPC-366 | SUM149 | ||||
|---|---|---|---|---|---|
| Treatments | Dosage a | Foci of Pulmonary Metastasis (Mean ± SD) | Foci of Hepatic Metastasis (Mean ± SD) | Foci of Pulmonary Metastasis (Mean ± SD) | Foci of Hepatic Metastasis (Mean ± SD) |
| Control | 13.20 ± 2.17 | 0.90 ± 0.60 | 10.40 ± 1,77 | 0.40 ± 0.19 | |
| VEGF | 0.2 mg/Kg | 13.40 ± 3.05 | 0.40 ± 0.15 | 16.40 ± 2.07 | 0.60 ± 0.29 |
| 0.4 mg/kg | 14.20 ± 1.92 | 0.60 ± 0.89 | 16.60 ± 2.30 | 2.20 ± 1.10 * | |
| 1 mg/Kg | 25.00 ± 4.85 * | 3.40 ± 1.19 | 50.60 ± 6.95 * | 4.33 ± 1.06 * | |
| SU5416 | 1 mg/Kg | 11.80 ± 1.48 | 0.40 ± 0.15 | 13.00 ± 5.00 | 1.00 ± 0.35 |
| 5 mg/Kg | 10.20 ± 3.96 | 0.40 ± 0.15 | 13.00 ± 2.83 | 0.40 ± 0.15 | |
| 10 mg/Kg | 10.40 ± 2.30 | 0.20 ± 0.10 | 12.40 ± 2.70 | 0.00 ± 0.00 * | |
| Bevacizumab | 1 mg/Kg | 6.80 ± 3.79 * | 0.40 ± 0.15 | 7.40 ± 2.58 | 0.00 ± 0.00 * |
| 5 mg/Kg | 1.20 ± 2.17 * | 1.00 ± 0.41 | 0.00 ± 0.00 * | 0.00 ± 0.00 * | |
| 10 mg/Kg | 0.00 ± 0.00 * | 0.00 ± 0.00 * | 0.00 ± 0.00 * | 0.00 ± 0.00 * | |
| Celecoxib | 0.5 mg/kg | 10.40 ± 2.88 | 0.40 ± 0.15 | 6.80 ± 1.82 | 0.00 ± 0.00 * |
| 2.5 mg/kg | 7.00 ± 2.21 | 0.00 ± 0.00 * | 7.40 ± 2.84 | 0.00 ± 0.00 * | |
| 5 mg/kg | 5.80 ± 3.10 * | 0.00 ± 0.00 * | 3.20 ± 1.44 | 0.00 ± 0.00 * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alonso-Diez, Á.; Cáceres, S.; Peña, L.; Crespo, B.; Illera, J.C. Anti-Angiogenic Treatments Interact with Steroid Secretion in Inflammatory Breast Cancer Triple Negative Cell Lines. Cancers 2021, 13, 3668. https://doi.org/10.3390/cancers13153668
Alonso-Diez Á, Cáceres S, Peña L, Crespo B, Illera JC. Anti-Angiogenic Treatments Interact with Steroid Secretion in Inflammatory Breast Cancer Triple Negative Cell Lines. Cancers. 2021; 13(15):3668. https://doi.org/10.3390/cancers13153668
Chicago/Turabian StyleAlonso-Diez, Ángela, Sara Cáceres, Laura Peña, Belén Crespo, and Juan Carlos Illera. 2021. "Anti-Angiogenic Treatments Interact with Steroid Secretion in Inflammatory Breast Cancer Triple Negative Cell Lines" Cancers 13, no. 15: 3668. https://doi.org/10.3390/cancers13153668
APA StyleAlonso-Diez, Á., Cáceres, S., Peña, L., Crespo, B., & Illera, J. C. (2021). Anti-Angiogenic Treatments Interact with Steroid Secretion in Inflammatory Breast Cancer Triple Negative Cell Lines. Cancers, 13(15), 3668. https://doi.org/10.3390/cancers13153668