
Role of IQGAP1 in Carcinogenesis


Abstract
:Simple Summary
Abstract
1. Introduction
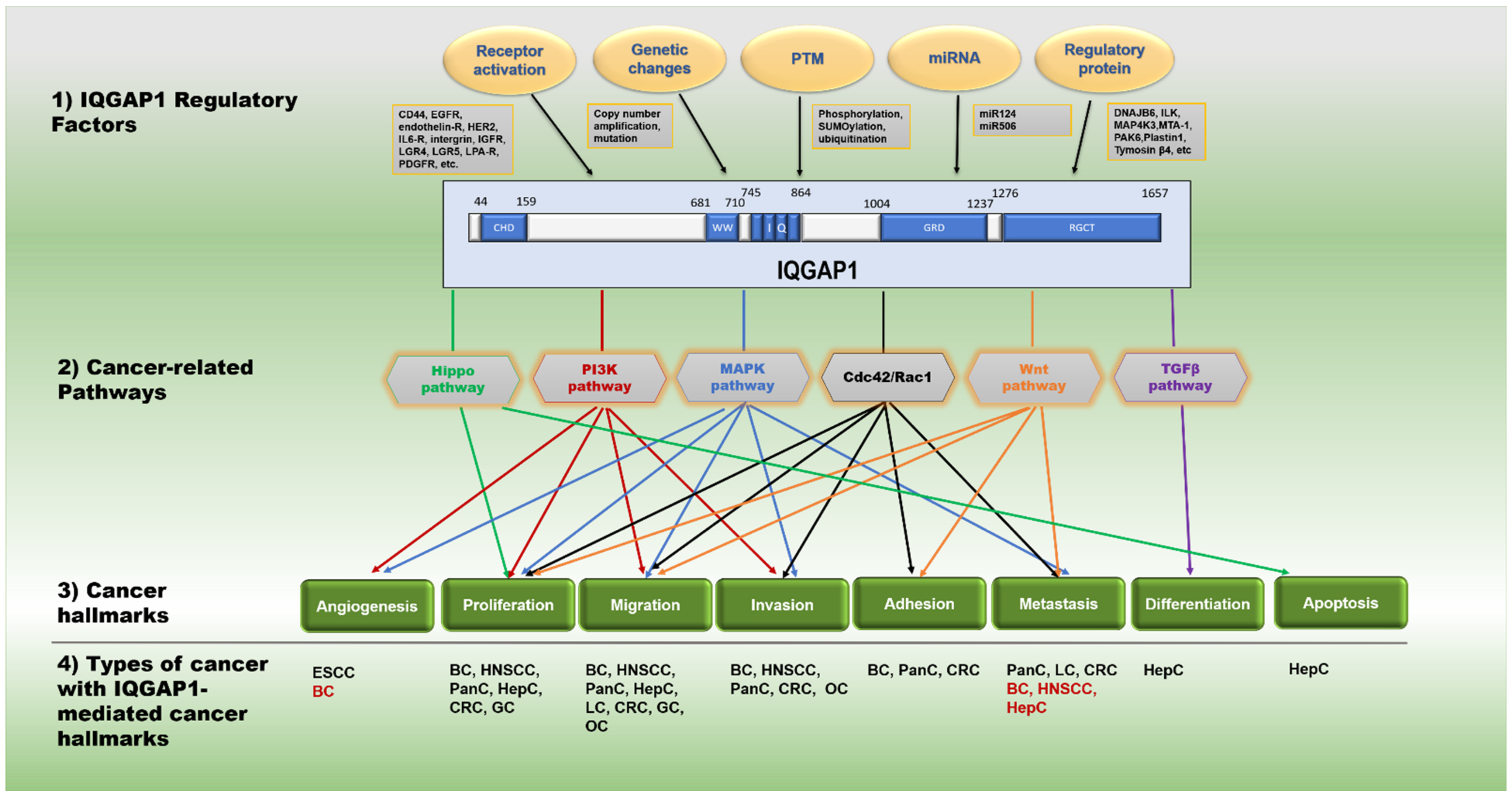
2. IQGAP1 Mediates Multiple Key Oncogenic Pathways
2.1. MAPK Signaling
2.2. RAC1/CDC42
2.3. Wnt Signaling
2.4. PI3K Signaling
2.5. Hippo Signaling
2.6. TGF-β Signaling
3. IQGAP1 Plays an Essential Role in Many Types of Cancer
3.1. IQGAP1 in Head and Neck Cancer
3.2. IQGAP1 in Breast Cancer
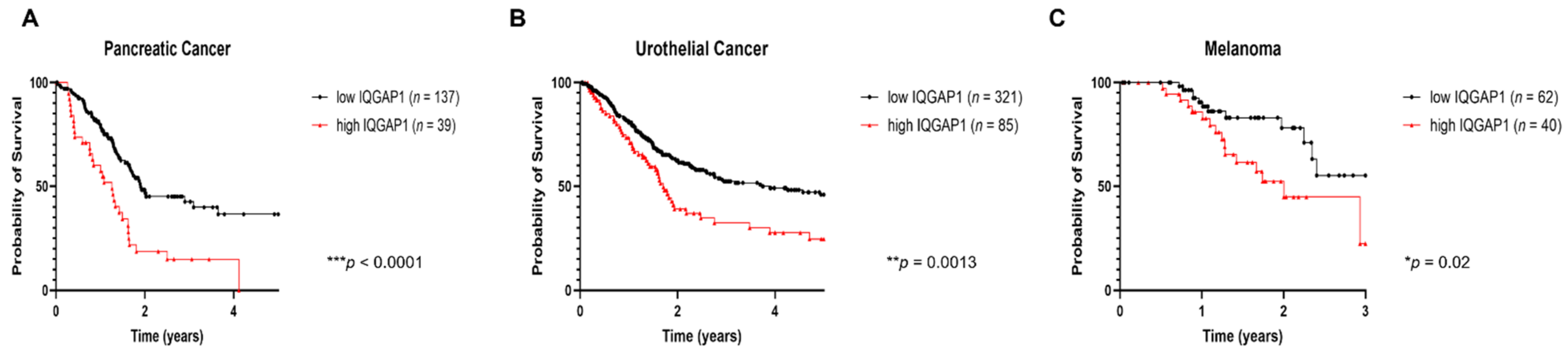
3.3. IQGAP1 in Pancreatic Cancer
3.4. IQGAP1 in Liver Cancer
3.5. IQGAP1 in Colorectal Cancer
3.6. IQGAP1 in Lung Cancer
3.7. IQGAP1 in Gastric Cancer
3.8. IQGAP1 in Ovarian Cancer
4. Links between IQGAP1 and Immune Cell Activities
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abel, A.M.; Schuldt, K.M.; Rajasekaran, K.; Hwang, D.; Riese, M.J.; Rao, S.; Thakar, M.S.; Malarkannan, S. IQGAP1: Insights into the function of a molecular puppeteer. Mol. Immunol. 2015, 65, 336–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedman, A.C.; Smith, J.M.; Sacks, D.B. The biology of IQGAP proteins: Beyond the cytoskeleton. EMBO Rep. 2015, 16, 427–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weissbach, L.; Settleman, J.; Kalady, M.F.; Snijders, A.J.; Murthy, A.E.; Yan, Y.X.; Bernards, A. Identification of a human RasGAP-related protein containing calmodulin- binding motifs. J. Biol. Chem. 1994, 269, 20517–20521. [Google Scholar] [CrossRef]
- Hart, M.J.; Callow, M.G.; Souza, B.; Polakis, P. IQGAP1, a calmodulin-binding protein with a rasGAP-related domain, is a potential effector for cdc42Hs. EMBO J. 1996, 15, 2997–3005. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.D.; Sacks, D.B. IQGAP1 in cellular signaling: Bridging the GAP. Trends Cell Biol. 2006, 16, 242–249. [Google Scholar] [CrossRef]
- Tanos, B.E.; Yeaman, C.; Rodriguez-Boulan, E. An emerging role for IQGAP1 in tight junction control. Small GTPases 2018, 9, 375–383. [Google Scholar] [CrossRef] [Green Version]
- White, C.D.; Erdemir, H.H.; Sacks, D.B. IQGAP1 and its binding proteins control diverse biological functions. Cell. Signal. 2012, 24, 826–834. [Google Scholar] [CrossRef] [Green Version]
- Mataraza, J.M.; Briggs, M.W.; Li, Z.; Entwistle, A.; Ridley, A.J.; Sacks, D.B. IQGAP1 Promotes Cell Motility and Invasion. J. Biol. Chem. 2003, 278, 41237–41245. [Google Scholar] [CrossRef] [Green Version]
- Noritake, J.; Watanabe, T.; Sato, K.; Wang, S.; Kaibuchi, K. IQGAP1: A key regulator of adhesion and migration. J. Cell Sci. 2005, 118, 2085–2092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.M.; Hedman, A.C.; Sacks, D.B. IQGAPs choreograph cellular signaling from the membrane to the nucleus. Trends Cell Biol. 2015, 25, 171–184. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.; Sharma, M.; Henderson, B.R. IQGAP1 regulation and roles in cancer. Cell. Signal. 2009, 21, 1471–1478. [Google Scholar] [CrossRef] [PubMed]
- White, C.D.; Brown, M.D.; Sacks, D.B. IQGAPs in cancer: A family of scaffold proteins underlying tumorigenesis. FEBS Lett. 2009, 583, 1817–1824. [Google Scholar] [CrossRef] [Green Version]
- Burotto, M.; Chiou, V.L.; Lee, J.-M.; Kohn, E.C. The MAPK pathway across different malignancies: A new perspective. Cancer 2014, 120, 3446–3456. [Google Scholar] [CrossRef] [Green Version]
- McNulty, D.E.; Li, Z.; White, C.D.; Sacks, D.B.; Annan, R.S. MAPK scaffold IQGAP1 binds the EGF receptor and modulates its activation. J. Biol. Chem. 2011, 286, 15010–15021. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.G.; Li, Z.; Sacks, D.B. IQGAP1 modulates activation of B-Raf. Proc. Natl. Acad. Sci. USA 2007, 104, 10465–10469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, M.; Li, Z.; Sacks, D.B. IQGAP1 Binds ERK2 and Modulates Its Activity. J. Biol. Chem. 2004, 279, 17329–17337. [Google Scholar] [CrossRef] [Green Version]
- Roy, M.; Li, Z.; Sacks, D.B. IQGAP1 Is a Scaffold for Mitogen-Activated Protein Kinase Signaling. Mol. Cell. Biol. 2005, 25, 7940–7952. [Google Scholar] [CrossRef] [Green Version]
- Bardwell, A.J.; Lagunes, L.; Zebarjedi, R.; Bardwell, L. The WW domain of the scaffolding protein IQGAP1 is neither necessary nor sufficient for binding to the MAPKs ERK1 and ERK2. J. Biol. Chem. 2017, 292, 8750–8761. [Google Scholar] [CrossRef] [Green Version]
- Morgan, C.J.; Hedman, A.C.; Li, Z.; Sacks, D.B. Endogenous IQGAP1 and IQGAP3 do not functionally interact with Ras. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, H.; Kubota, K.; Inoue, T.; Isono, F.; Ando, O. IQGAP1 selectively interacts with K-Ras but not with H-Ras and modulates K-Ras function. Biochem. Biophys. Res. Commun. 2014, 444, 360–364. [Google Scholar] [CrossRef]
- White, C.D.; Li, Z.; Dillon, D.A.; Sacks, D.B. IQGAP1 binds human epidermal growth factor receptor 2 (HER2) and modulates trastuzumab resistance in HER2-positive (HER2(+)) human breast cancer. J. Biol. Chem. 2011. [Google Scholar] [CrossRef] [Green Version]
- Jameson, K.L.; Mazur, P.K.; Zehnder, A.M.; Zhang, J.; Zarnegar, B.; Sage, J.; Khavari, P.A. IQGAP1 scaffold-kinase interaction blockade selectively targets RAS-MAP kinase—Driven tumors. Nat. Med. 2013, 19. [Google Scholar] [CrossRef] [Green Version]
- Tanos, B.; Perez Bay, A.; Salvarezza, S.; Vivanco, I.; Mellinghoff, I.; Osman, M.; Sacks, D.; Rodriguez-Boulan, E. IQGAP1 controls tight junction formation through differential regulation of claudin recruitment. J. Cell Sci. 2015, 128, 853–862. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, S.; Fukata, M.; Kobayashi, K.; Nakafuku, M.; Nomura, N.; Iwamatsu, A.; Kaibuchi, K. Identification of IQGAP as a putative target for the small GTPases, Cdc42 and Rac1. J. Biol. Chem. 1996, 271, 23363–23367. [Google Scholar] [CrossRef] [Green Version]
- Fukataf, M.; Kuroda, S.; Fujii, K.; Nakamura, T.; Shoji, I.; Matsuura, Y.; Okawa, K.; Iwamatsu, A.; Kikuchi, A.; Kaibuchi, K. Regulation of cross-linking of actin filament by IQGAP1, a target for Cdc42. J. Biol. Chem. 1997, 272, 29579–29583. [Google Scholar] [CrossRef] [Green Version]
- Mateer, S.C.; McDaniel, A.E.; Nicolas, V.; Habermacher, G.M.; Lin, M.J.S.; Cromer, D.A.; King, M.E.; Bloom, G.S. The mechanism for regulation of the F-actin binding activity of IQGAP1 by calcium/calmodulin. J. Biol. Chem. 2002, 277, 12324–12333. [Google Scholar] [CrossRef] [Green Version]
- Briggs, M.W.; Sacks, D.B. IQGAP1 as signal integrator: Ca2+, calmodulin, Cdc42 and the cytoskeleton. FEBS Lett. 2003, 542, 7–11. [Google Scholar] [CrossRef] [Green Version]
- Grohmanova, K.; Schlaepfer, D.; Hess, D.; Gutierrez, P.; Beck, M.; Kroschewski, R. Phosphorylation of IQGAP1 modulates its binding to Cdc42, revealing a new type of Rho-GTPase regulator. J. Biol. Chem. 2004, 279, 48495–48504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorisse, L.; Li, Z.; Wagner, C.D.; Worthylake, D.K.; Zappacosta, F.; Hedman, A.C.; Annan, R.S.; Sacks, D.B. Ubiquitination of the scaffold protein IQGAP1 diminishes its interaction with and activation of the Rho GTPase CDC42. J. Biol. Chem. 2020, 295, 4822–4835. [Google Scholar] [CrossRef] [PubMed]
- Fukata, M.; Watanabe, T.; Noritake, J.; Nakagawa, M.; Yamaga, M.; Kuroda, S.; Matsuura, Y.; Iwamatsu, A.; Perez, F.; Kaibuchi, K. Rac1 and Cdc42 capture microtubules through IQGAP1 and CLIP-170. Cell 2002, 109, 873–885. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, T.; Noritake, J.; Kakeno, M.; Matsui, T.; Harada, T.; Wang, S.; Itoh, N.; Sato, K.; Matsuzawa, K.; Iwamatsu, A.; et al. Phosphorylation of CLASP2 by GSK-3β regulates its interaction with IQGAP1, EB1 and microtubules. J. Cell Sci. 2009, 122, 2969–2979. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, T.; Wang, S.; Noritake, J.; Sato, K.; Fukata, M.; Takefuji, M.; Nakagawa, M.; Izumi, N.; Akiyama, T.; Kaibuchi, K. Interaction with IQGAP1 links APC to Rac1, Cdc42, and actin filaments during cell polarization and migration. Dev. Cell 2004, 7, 871–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razidlo, G.L.; Burton, K.M.; McNiven, M.A. Interleukin-6 promotes pancreatic cancer cell migration by rapidly activating the small GTPase CDC42. J. Biol. Chem. 2018, 293, 11143–11153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, T.; Wang, S.; Kaibuchi, K. IQGAPS as key regulators of actin-cytoskeleton dynamics. Cell Struct. Funct. 2015, 40, 69–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmon, K.S.; Gong, X.; Yi, J.; Wu, L.; Thomas, A.; Moore, C.M.; Masuho, I.; Timson, D.J.; Martemyanov, K.A.; Liu, Q.J. LGR5 receptor promotes cell-cell adhesion in stem cells and colon cancer cells via the IQGAP1-Rac1 pathway. J. Biol. Chem. 2017, 292, 14989–15001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hage, B.; Meinel, K.; Baum, I.; Giehl, K.; Menke, A. Rac1 activation inhibits E-cadherin-mediated adherens junctions via binding to IQGAP1 in pancreatic carcinoma cells. Cell Commun. Signal. 2009, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Sakurai-Yageta, M.; Recchi, C.; Le Dez, G.; Sibarita, J.B.; Daviet, L.; Camonis, J.; D’Souza-Schorey, C.; Chavrier, P. The interaction of IQGAP1 with the exocyst complex is required for tumor cell invasion downstream of Cdc42 and RhoA. J. Cell Biol. 2008, 181, 985–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jadeski, L.; Mataraza, J.M.; Jeong, H.W.; Li, Z.; Sacks, D.B. IQGAP1 stimulates proliferation and enhances tumorigenesis of human breast epithelial cells. J. Biol. Chem. 2008, 283, 1008–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cvetković, D.; Dragan, M.; Leith, S.J.; Mir, Z.M.; Leong, H.S.; Pampillo, M.; Lewis, J.D.; Babwah, A.V.; Bhattacharya, M. KISS1R induces invasiveness of estrogen receptor-negative human mammary epithelial and breast cancer cells. Endocrinology 2013, 154, 1999–2014. [Google Scholar] [CrossRef] [Green Version]
- Alemayehu, M.; Dragan, M.; Pape, C.; Siddiqui, I.; Sacks, D.B.; Di Guglielmo, G.M.; Babwah, A.V.; Bhattacharya, M. β-Arrestin2 Regulates Lysophosphatidic Acid-Induced Human Breast Tumor Cell Migration and Invasion via Rap1 and IQGAP1. PLoS ONE 2013, 8, e56174. [Google Scholar] [CrossRef]
- Schatoff, E.M.; Leach, B.I.; Dow, L.E. WNT Signaling and Colorectal Cancer. Curr. Colorectal Cancer Rep. 2017, 13, 101–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmon, K.S.; Gong, X.; Yi, J.; Thomas, A.; Liu, Q. RSPO-LGR4 functions via IQGAP1 to potentiate Wnt signaling. Proc. Natl. Acad. Sci. USA 2014, 111, E1221–E1229. [Google Scholar] [CrossRef] [Green Version]
- Goto, T.; Sato, A.; Shimizu, M.; Adachi, S.; Satoh, K.; Iemura, S.I.; Natsume, T.; Shibuya, H. IQGAP1 Functions as a Modulator of Dishevelled Nuclear Localization in Wnt Signaling. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Goto, T.; Sato, A.; Adachi, S.; Iemura, S.I.; Natsume, T.; Shibuya, H. IQGAP1 protein regulates nuclear localization of β-Catenin via importin-β5 protein in wnt signaling. J. Biol. Chem. 2013, 288, 36351–36360. [Google Scholar] [CrossRef] [Green Version]
- Simpson, D.R.; Mell, L.K.; Cohen, E.E.W. Targeting the PI3K/AKT/mTOR pathway in squamous cell carcinoma of the head and neck. Oral Oncol. 2015, 51, 291–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Zhu, H.H.; Zhou, L.F.; Wu, S.S.; Wang, J.; Chen, Z. IQGAP1 is overexpressed in hepatocellular carcinoma and promotes cell proliferation by Akt activation. Exp. Mol. Med. 2010, 42, 477–483. [Google Scholar] [CrossRef]
- Choi, S.; Anderson, R.A. IQGAP1 is a phosphoinositide effector and kinase scaffold. Adv. Biol. Regul. 2016, 60, 29–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.; Hedman, A.C.; Sayedyahossein, S.; Thapa, N.; Sacks, D.B.; Anderson, R.A. Agonist-stimulated phosphatidylinositol-3,4,5-trisphosphate generation by scaffolded phosphoinositide kinases. Nat. Cell Biol. 2016, 18, 1324–1335. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.; Thapa, N.; Hedman, A.C.; Li, Z.; Sacks, D.B.; Anderson, R.A. IQGAP1 is a novel phosphatidylinositol 4,5 bisphosphate effector in regulation of directional cell migration. EMBO J. 2013. [Google Scholar] [CrossRef]
- Chen, M.; Choi, S.; Jung, O.; Wen, T.; Baum, C.; Thapa, N.; Lambert, P.F.; Rapraeger, A.C.; Anderson, R.A. The Specificity of EGF-Stimulated IQGAP1 Scaffold Towards the PI3K-Akt Pathway is Defined by the IQ3 motif. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef]
- Wei, T.; Choi, S.; Buehler, D.; Anderson, R.A.; Lambert, P.F. A PI3K/AKT scaffolding protein, IQ motif–containing GTPase associating protein 1 (IQGAP1), promotes head and neck carcinogenesis. Clin. Cancer Res. 2020, 26, 301–311. [Google Scholar] [CrossRef] [Green Version]
- Harvey, K.F.; Zhang, X.; Thomas, D.M. The Hippo pathway and human cancer. Nat. Rev. Cancer 2013, 13, 246–257. [Google Scholar] [CrossRef]
- Quinn, N.P.; García-Gutiérrez, L.; Doherty, C.; von Kriegsheim, A.; Fallahi, E.; Sacks, D.B.; Matallanas, D. IQGAP1 Is a Scaffold of the Core Proteins of the Hippo Pathway and Negatively Regulates the Pro-Apoptotic Signal Mediated by This Pathway. Cells 2021, 10, 478. [Google Scholar] [CrossRef] [PubMed]
- Sayedyahossein, S.; Li, Z.; Hedman, A.C.; Morgan, C.J.; Sacks, D.B. IQGAP1 Binds to Yes-associated Protein (YAP) and Modulates Its Transcriptional Activity. J. Biol. Chem. 2016, 291, 19261–19273. [Google Scholar] [CrossRef] [Green Version]
- Delgado, E.R.; Erickson, H.L.; Tao, J.; Monga, S.P.; Duncan, A.W.; Anakk, S. Scaffolding Protein IQGAP1 Is Dispensable, but Its Overexpression Promotes Hepatocellular Carcinoma via YAP1 Signaling. Mol. Cell. Biol. 2021, 41. [Google Scholar] [CrossRef]
- Massagué, J. TGFβ in Cancer. Cell 2008, 134, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Shah, V.H.; Kang, N.; Billadeau, D.D.; Abdelhakim, H.; Leof, E.; Kaibuchi, K.; Bernabeu, C.; Bloom, G.S.; Yang, L.; et al. IQGAP1 suppresses Tb bRII-mediated myofibroblastic activation and metastatic growth in liver IQGAP1 suppresses TβRII-mediated myofibroblastic activation and metastatic growth in liver. J. Clin. Investig. 2013, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, L.; Heller, M.; Meng, Z.; Yu, L.; Tang, Y.; Zhou, M.; Zhang, Y. Transforming Growth Factor-β (TGF-β) Directly Activates the JAK1-STAT3 Axis to Induce Hepatic Fibrosis in Coordination with the SMAD Pathway. J. Biol. Chem. 2017, 292, 4302–4312. [Google Scholar] [CrossRef] [Green Version]
- Zong, C.; Zhang, X.; Xie, Y.; Cheng, J. Transforming growth factor-β inhibits IQ motif containing guanosine triphosphatase activating protein 1 expression in lung fibroblasts via the nuclear factor-κB signaling pathway. Mol. Med. Rep. 2015, 12, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Law, B.; Aakre, M.; Edgerton, M.; Shyr, Y.; Bhowmick, N.; Moses, H. Transforming growth factor beta-regulated gene expression in a mouse mammary gland epithelial cell line. Breast Cancer Res. 2003, 5. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Zhao, J.; Mao, G.; Lee, G.S.; Zhang, J.; Bi, L.; Gu, L.; Chang, Z.; Valentino, J.; Li, G.-M. Identification of novel genetic variants predisposing to familial oral squamous cell carcinomas. Cell Discov. 2019, 5, 57. [Google Scholar] [CrossRef] [Green Version]
- Morris, L.E.; Bloom, G.S.; Frierson, H.F.; Powell, S.M. Nucleotide variants within the IQGAP1 gene in diffuse-type gastric cancers. Genes Chromosom. Cancer 2005, 42, 280–286. [Google Scholar] [CrossRef]
- Nakamura, H.; Fujita, K.; Nakagawa, H.; Kishi, F.; Takeuchi, A.; Aute, I.; Kato, H. Expression pattern of the scaffold protein IQGAP1 in lung cancer. Oncol. Rep. 2005, 13, 427–431. [Google Scholar] [CrossRef]
- Dong, P.; Nabeshima, K.; Nishimura, N.; Kawakami, T.; Hachisuga, T.; Kawarabayashi, T.; Iwasaki, H. Overexpression and diffuse expression pattern of IQGAP1 at invasion fronts are independent prognostic parameters in ovarian carcinomas. Cancer Lett. 2006, 243, 120–127. [Google Scholar] [CrossRef]
- Zhao, H.Y.; Han, Y.; Wang, J.; Yang, L.H.; Zheng, X.Y.; Du, J.; Wu, G.P.; Wang, E.H. IQ-domain GTPase-activating protein 1 promotes the malignant phenotype of invasive ductal breast carcinoma via canonical Wnt pathway. Tumor Biol. 2017, 39, 1010428317705769. [Google Scholar] [CrossRef] [Green Version]
- Zeng, F.; Jiang, W.; Zhao, W.; Fan, Y.; Zhu, Y.; Zhang, H. Ras GTPase-activating-like protein IQGAP1 (IQGAP1) promotes breast cancer proliferation and invasion and correlates with poor clinical outcomes. Med. Sci. Monit. 2018, 24, 3315–3323. [Google Scholar] [CrossRef]
- Wu, C.-C.; Li, H.; Xiao, Y.; Yang, L.-L.; Chen, L.; Deng, W.-W.; Wu, L.; Zhang, W.-F.; Sun, Z.-J. Over-expression of IQGAP1 indicates poor prognosis in head and neck squamous cell carcinoma. J. Mol. Histol. 2018, 49, 389–398. [Google Scholar] [CrossRef]
- Patel, V.; Hood, B.L.; Molinolo, A.A.; Lee, N.H.; Conrads, T.P.; Braisted, J.C.; Krizman, D.B.; Veenstra, T.D.; Gutkind, J.S. Proteomic analysis of laser-captured paraffin-embedded tissues: A molecular portrait of head and neck cancer progression. Clin. Cancer Res. 2008, 14, 1002–1014. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.-X.; Li, X.-Z.; Zhai, L.-Q.; Liu, Z.-R.; Chen, X.-J.; Pei, Y. Overexpression of IQGAP1 in human pancreatic cancer. Hepatobiliary Pancreat. Dis. Int 2013, 12, 540–545. [Google Scholar] [CrossRef]
- Hu, W.; Wang, Z.; Zhang, S.; Lu, X.; Wu, J.; Yu, K.; Ji, A.; Lu, W.; Wang, Z.; Wu, J.; et al. IQGAP1 promotes pancreatic cancer progression and epithelial-mesenchymal transition (EMT) through Wnt/β-catenin signaling. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Wen, T.-F.; Zhang, X.-Y.; Chen, X.; Shen, J.-Y. IQGAP1 expression in hepatocellular carcinoma predicts poor prognosis by inducing epithelial-mesenchymal transition. Transl. Cancer Res. 2017, 6, 530–540. [Google Scholar] [CrossRef]
- Xia, F.D.; Wang, Z.L.; Chen, H.X.; Huang, Y.; Li, J.D.; Wang, Z.M.; Li, X.Y. Differential expression of IQGAP1/2 in hepatocellular carcinoma and its relationship with clinical outcomes. Asian Pac. J. Cancer Prev. 2014, 15, 4951–4956. [Google Scholar] [CrossRef] [Green Version]
- Bertucci, F.; Salas, S.; Eysteries, S.; Nasser, V.; Finetti, P.; Ginestier, C.; Charafe-Jauffret, E.; Loriod, B.; Bachelart, L.; Montfort, J.; et al. Gene expression profiling of colon cancer by DNA microarrays and correlation with histoclinical parameters. Oncogene 2004, 23, 1377–1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, H.; Nabeshima, K.; Aoki, M.; Hamasaki, M.; Enatsu, S.; Yamauchi, Y.; Yamashita, Y.; Iwasaki, H. Overexpression of IQGAP1 in advanced colorectal cancer correlates with poor prognosis-critical role in tumor invasion. Int. J. Cancer 2010, 126, NA-NA. [Google Scholar] [CrossRef]
- Nabeshima, K.; Shimao, Y.; Inoue, T.; Koono, M. Immunohistochemical analysis of IQGAP1 expression in human colorectal carcinomas: Its overexpression in carcinomas and association with invasion fronts. Cancer Lett. 2002, 176, 101–109. [Google Scholar] [CrossRef]
- Takemoto, H.; Doki, Y.; Shiozaki, H.; Imamura, H.; Utsunomiya, T.; Miyata, H.; Inoue, M.Y.M.; Fujiwara, Y.; Monden, M. Localization of IQGAP1 is inversely correlated with intercellular adhesion mediated by E-cadherin in gastric cancers. Int. J. Cancer 2001, 91, 783–788. [Google Scholar] [CrossRef]
- Walch, A.; Seidl, S.; Hermannstädter, C.; Rauser, S.; Deplazes, J.; Langer, R.; Von Weyhern, C.H.; Sarbia, M.; Busch, R.; Feith, M.; et al. Combined analysis of Rac1, IQGAP1, Tiam1 and E-cadherin expression in gastric cancer. Mod. Pathol. 2008, 21, 544–552. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Gao, Y.; Cui, Y.; Zhang, T.; Cui, R.; Jiang, Y.; Shi, J. Overexpression of CD44 is associated with the occurrence and migration of non-small cell lung cancer. Mol. Med. Rep. 2016, 14, 3159–3167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyoshi, T.; Shirakusa, T.; Ishikawa, Y.; Iwasaki, A.; Shiraishi, T.; Makimoto, Y.; Iwasaki, H.; Nabeshima, K. Possible mechanism of metastasis in lung adenocarcinomas with a micropapillary pattern. Pathol. Int. 2005, 55, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Zhang, K.; Zhang, X.; Lei, W.; Xiao, T.; Ma, J.; Guo, S.; Shao, S.; Zhang, H.; Liu, Y.; et al. Identification of differentially expressed genes in human lung squamous cell carcinoma using suppression subtractive hybridization. Cancer Lett. 2004, 212, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Shao, Y.; Ren, L.; Liu, X.; Li, Y.; Xu, J.; Ye, Y. IQGAP2 inhibits migration and invasion of gastric cancer cells via elevating SHIP2 phosphatase activity. Int. J. Mol. Sci. 2020, 21, 1968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.; Zheng, L.; Tao, L. Downregulation of IQGAP2 Correlates with Prostate Cancer Recurrence and Metastasis. Transl. Oncol. 2019, 12, 236–244. [Google Scholar] [CrossRef]
- Kumar, D.; Hassan, M.K.; Pattnaik, N.; Mohapatra, N.; Dixit, M. Reduced expression of IQGAP2 and higher expression of IQGAP3 correlates with poor prognosis in cancers. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhao, W.; Xu, Q.W.; Wang, X.S.; Zhang, Y.; Zhang, J. IQGAP3 promotes EGFR-ERK signaling and the growth and metastasis of lung cancer cells. PLoS ONE 2014, 9, e97578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteleon, C.L.; McNeal, A.; Duperret, E.K.; Oh, S.J.; Schapira, E.; Ridky, T.W. IQGAP1 and IQGAP3 Serve Individually Essential Roles in Normal Epidermal Homeostasis and Tumor Progression. J. Investig. Dermatol. 2015, 135, 2258–2265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- American Cancer Society. Cancer Facts& Figures 2020; American Cancer Society: Atlanta, GA, USA, 2020. [Google Scholar]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA. Cancer J. Clin. 2021, caac.21660. [Google Scholar] [CrossRef]
- Li, Z.X.; Zheng, Z.Q.; Wei, Z.H.; Zhang, L.L.; Li, F.; Lin, L.; Liu, R.Q.; Huang, X.D.; Lv, J.W.; Chen, F.P.; et al. Comprehensive characterization of the alternative splicing landscape in head and neck squamous cell carcinoma reveals novel events associated with tumorigenesis and the immune microenvironment. Theranostics 2019, 9, 7648–7665. [Google Scholar] [CrossRef]
- Lawrence, M.S.; Sougnez, C.; Lichtenstein, L.; Cibulskis, K.; Lander, E.; Gabriel, S.B.; Getz, G.; Ally, A.; Balasundaram, M.; Birol, I.; et al. Comprehensive genomic characterization of head and neck squamous cell carcinomas. Nature 2015, 517, 576–582. [Google Scholar] [CrossRef] [Green Version]
- Mountzios, G.; Rampias, T.; Psyrri, A. The mutational spectrum of squamous-cell carcinoma of the head and neck: Targetable genetic events and clinical impact. Ann. Oncol. 2014, 25, 1889–1900. [Google Scholar] [CrossRef] [PubMed]
- Lui, V.W.Y.; Hedberg, M.L.; Li, H.; Vangara, B.S.; Pendleton, K.; Zeng, Y.; Lu, Y.; Zhang, Q.; Du, Y.; Gilbert, B.R.; et al. Frequent mutation of the PI3K pathway in head and neck cancer defines predictive biomarkers. Cancer Discov. 2013, 3, 761–769. [Google Scholar] [CrossRef] [Green Version]
- Lu, B.; Lian, R.; Wu, Z.; Miao, W.; Li, X.; Li, J.; Shi, Y.; Yu, W. MTA1 promotes viability and motility in nasopharyngeal carcinoma by modulating IQGAP1 expression. J. Cell. Biochem. 2018, 119, 3864–3872. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.X.; Wang, K.; Li, X.Z.; Zhai, L.Q.; Qu, C.X.; Zhao, Y.; Liu, Z.R.; Wang, H.Z.; An, Q.J.; Jing, L.W.; et al. Targeted knockdown of IQGAP1 inhibits the progression of esophageal squamous cell carcinoma in vitro and in vivo. PLoS ONE 2014, 9, e96501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; He, C.; Li, C.; Ren, B.; Deng, Q.; Gao, W.; Wang, B. IQGAP1 silencing suppresses the malignant characteristics of laryngeal squamous cell carcinoma cells. Int. J. Biol. Markers 2018, 33, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Li, C.H.; Sun, X.J.; Niu, S.S.; Yang, C.Y.; Hao, Y.P.; Kou, J.T.; Li, X.Z.; Wang, X.X. Overexpression of IQGAP1 promotes the angiogenesis of esophageal squamous cell carcinoma through the AKT and ERK-mediated VEGF-VEGFR2 signaling pathway. Oncol. Rep. 2018, 40, 1795–1802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Wang, Q.; Chakladar, A.; Bronson, R.T.; Bernards, A. Gastric Hyperplasia in Mice Lacking the Putative Cdc42 Effector IQGAP1. Mol. Cell. Biol. 2000, 20, 697–701. [Google Scholar] [CrossRef] [Green Version]
- CDC. Cancers Associated with Human Papillomavirus, United States—2013–2017; CDC: Atlanta, GA, USA, 2020. [Google Scholar]
- National Center for Health Statistics. Health, United States 2018 Chartbook; National Center for Health Statistics: Hyattsville, MD, USA, 2019.
- Gillison, M.L.; Koch, W.M.; Capone, R.B.; Spafford, M.; Westra, W.H.; Wu, L.; Zahurak, M.L.; Daniel, R.W.; Viglione, M.; Symer, D.E.; et al. Evidence for a Causal Association Between Human Papillomavirus and a Subset of Head and Neck Cancers. J. Natl. Cancer Inst. 2000, 92, 709–720. [Google Scholar] [CrossRef]
- Mahal, B.A.; Catalano, P.J.; Haddad, R.I.; Hanna, G.J.; Kass, J.I.; Schoenfeld, J.D.; Tishler, R.B.; Margalit, D.N. Incidence and demographic burden of HPV-associated oropharyngeal head and neck cancers in the United States. Cancer Epidemiol. Biomarkers Prev. 2019, 28, 1660–1667. [Google Scholar] [CrossRef] [Green Version]
- Marur, S.; D’Souza, G.; Westra, W.H.; Forastiere, A.A. HPV-associated head and neck cancer: A virus-related cancer epidemic. Lancet Oncol. 2010, 11, 781–789. [Google Scholar] [CrossRef] [Green Version]
- Cancer Genome Atlas Network Comprehensive genomic characterization of head and neck squamous cell carcinomas. Nature 2015, 517, 576–582. [CrossRef] [PubMed] [Green Version]
- Wei, T.; Choi, S.; Buehler, D.; Lee, D.; Ward-Shaw, E.; Anderson, R.A.; Lambert, P.F. Role of IQGAP1 in Papillomavirus-Associated Head and Neck Tumorigenesis. Cancers 2021, 13, 2276. [Google Scholar] [CrossRef]
- Wei, T.; Buehler, D.; Ward-Shaw, E.; Lambert, P.F. An infection-based murine model for papillomavirus-associated head and neck cancer. MBio 2020, 11. [Google Scholar] [CrossRef]
- Spurgeon, M.E.; Uberoi, A.; McGregor, S.M.; Wei, T.; Ward-Shaw, E.; Lambert, P.F. A Novel In Vivo Infection Model To Study Papillomavirus-Mediated Disease of the Female Reproductive Tract. MBio 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Uberoi, A.; Yoshida, S.; Frazer, I.H.; Pitot, H.C.; Lambert, P.F. Role of Ultraviolet Radiation in Papillomavirus-Induced Disease. PLoS Pathog. 2016, 12. [Google Scholar] [CrossRef] [PubMed]
- Spurgeon, M.E.; Lambert, P.F. MmuPV1: A New Frontier in Animal Models of Papillomavirus Pathogenesis. J. Virol. 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surviladze, Z.; Sterk, R.T.; DeHaro, S.A.; Ozbun, M.A. Cellular Entry of Human Papillomavirus Type 16 Involves Activation of the Phosphatidylinositol 3-Kinase/Akt/mTOR Pathway and Inhibition of Autophagy. J. Virol. 2013, 87, 2508–2517. [Google Scholar] [CrossRef] [Green Version]
- Herber, R.; Liem, A.; Pitot, H.; Lambert, P.F. Squamous epithelial hyperplasia and carcinoma in mice transgenic for the human papillomavirus type 16 E7 oncogene. J. Virol. 1996, 70, 1873–1881. [Google Scholar] [CrossRef] [Green Version]
- Song, S.; Pitot, H.C.; Lambert, P.F. The Human Papillomavirus Type 16 E6 Gene Alone Is Sufficient To Induce Carcinomas in Transgenic Animals. J. Virol. 1999, 73, 5887–5893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stelzer, M.K.; Pitot, H.C.; Liem, A.; Lee, D.; Kennedy, G.D.; Lambert, P.F. Rapamycin Inhibits Anal Carcinogenesis in Two Preclinical Animal Models. Cancer Prev. Res. 2010, 3, 1542–1551. [Google Scholar] [CrossRef] [Green Version]
- Branca, M.; Ciotti, M.; Santini, D.; Bonito, L. Di Benedetto, A.; Giorgi, C.; Paba, P.; Favalli, C.; Costa, S.; Agarossi, A.; et al. Activation of the ERK/MAP Kinase Pathway in Cervical Intraepithelial Neoplasia Is Related to Grade of the Lesion but Not to High-Risk Human Papillomavirus, Virus Clearance, or Prognosis in Cervical Cancer on behalf of the HPV-Pathogen ISS Study Group. Am J Clin Pathol 2004, 122, 902–911. [Google Scholar] [CrossRef] [PubMed]
- Rong, C.; Muller, M.; Flechtenmacher, C.; Holzinger, D.; Dyckhoff, G.; Bulut, O.C.; Horn, D.; Plinkert, P.; Hess, J.; Affolter, A. Differential activation of erk signaling in hpv-related oropharyngeal squamous cell carcinoma. Cancers 2019, 11, 584. [Google Scholar] [CrossRef] [Green Version]
- Shin, M.-K.; Payne, S.; Bilger, A.; Matkowskyj, K.A.; Carchman, E.; Meyer, D.S.; Bentires-Alj, M.; Deming, D.A.; Lambert, P.F. Activating Mutations in Pik3ca Contribute to Anal Carcinogenesis in the Presence or Absence of HPV-16 Oncogenes. Clin. Cancer Res. 2019, 25, 1889–1900. [Google Scholar] [CrossRef]
- Deshmukh, J.; Pofahl, R.; Pfister, H.; Haase, I. Deletion of epidermal Rac1 inhibits HPV-8 induced skin papilloma formation and facilitates HPV-8- and UV-light induced skin carcinogenesis. Oncotarget 2016, 7, 57841–57850. [Google Scholar] [CrossRef] [Green Version]
- Oliver, A.W.; He, X.; Borthwick, K.; Donne, A.J.; Hampson, L.; Hampson, I.N. The HPV16 E6 binding protein Tip-1 interacts with ARHGEF16, which activates Cdc42. Br. J. Cancer 2011, 104, 324–331. [Google Scholar] [CrossRef] [Green Version]
- Morgan, E.L.; Macdonald, A. Autocrine STAT3 activation in hpv positive cervical cancer through a virus-driven Rac1—NFκB—IL-6 signalling axis. PLoS Pathog. 2019, 15, e1007835. [Google Scholar] [CrossRef] [Green Version]
- Patel, V.; Rosenfeldt, H.M.; Lyons, R.; Servitja, J.M.; Bustelo, X.R.; Siroff, M.; Gutkind, J.S. Persistent activation of Rac1 in squamous carcinomas of the head and neck: Evidence for an EGFR/Vav2 signaling axis involved in cell invasion. Carcinogenesis 2007, 28, 1145–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, E.L.; Scarth, J.A.; Patterson, M.R.; Wasson, C.W.; Hemingway, G.C.; Barba-Moreno, D.; Macdonald, A. E6-mediated activation of JNK drives EGFR signalling to promote proliferation and viral oncoprotein expression in cervical cancer. Cell Death Differ. 2021, 28, 1669–1687. [Google Scholar] [CrossRef]
- Gkouveris, I.; Nikitakis, N.G. Role of JNK signaling in oral cancer: A mini review. Tumour Biol. 2017, 39. [Google Scholar] [CrossRef] [Green Version]
- Erdemir, H.H.; Li, Z.; Sacks, D.B. IQGAP1 binds to estrogen receptor-α and modulates its function. J. Biol. Chem. 2014, 289, 9100–9112. [Google Scholar] [CrossRef] [Green Version]
- Hebert, J.D.; Tian, C.; Lamar, J.M.; Rickelt, S.; Abbruzzese, G.; Liu, X.; Hynes, R.O. The scaffold protein IQGAP1 is crucial for extravasation and metastasis. Sci. Rep. 2020, 10, 2439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, G.; Liu, Y.; Wang, K.; Xu, Z. MIR-506 regulates breast cancer cell metastasis by targeting IQGAP1. Int. J. Oncol. 2015, 47, 1963–1970. [Google Scholar] [CrossRef] [PubMed]
- Bryant, K.L.; Mancias, J.D.; Kimmelman, A.C.; Der, C.J. KRAS: Feeding pancreatic cancer proliferation. Trends Biochem. Sci. 2014, 39. [Google Scholar] [CrossRef] [Green Version]
- Li, J.H.; McMillan, R.H.; Begum, A.; Gocke, C.B.; Matsui, W. IQGAP1 Maintains Pancreatic Ductal Adenocarcinoma Clonogenic Growth and Metastasis. Pancreas 2019, 48, 94–98. [Google Scholar] [CrossRef]
- Jin, Y.; Lv, X.; Zhou, J.; Chen, J. Potential involvement of IQGAP1 in proliferation and metastasis of human pancreatic cancer. Front. Biosci. 2016, 21, 1076–1083. [Google Scholar]
- Jin, X.; Pan, Y.; Wang, L.; Ma, T.; Zhang, L.; Tang, A.H.; Billadeau, D.D.; Wu, H.; Huang, H. Fructose-1,6-bisphosphatase inhibits ERK activation and bypasses gemcitabine resistance in pancreatic cancer by blocking IQGAP1–MAPK interaction. Cancer Res. 2017, 77, 4328–4341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Sousa Cavalcante, L.; Monteiro, G. Gemcitabine: Metabolism and molecular mechanisms of action, sensitivity and chemoresistance in pancreatic cancer. Eur. J. Pharmacol. 2014, 741, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Fryer, R.A.; Barlett, B.; Galustian, C.; Dalgleish, A.G. Mechanisms Underlying Gemcitabine Resistance in Pancreatic Cancer and Sensitisation by the iMiDTM Lenalidomide. Anticancer Res. 2011, 31, 3747–3756. [Google Scholar] [PubMed]
- Schmidt, V.A. Watch the GAP: Emerging Roles for IQ Motif-Containing GTPase-Activating Proteins IQGAPs in Hepatocellular Carcinoma. Int. J. Hepatol. 2012, 2012, 958673. [Google Scholar] [CrossRef] [Green Version]
- White, C.D.; Khurana, H.; Gnatenko, D.V.; Li, Z.; Odze, R.D.; Sacks, D.B.; Schmidt, V.A. IQGAP1 and IQGAP2 are Reciprocally Altered in Hepatocellular Carcinoma. BMC Gastroenterol. 2010, 10, 125. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, V.A.; Chiariello, C.S.; Capilla, E.; Miller, F.; Bahou, W.F. Development of hepatocellular carcinoma in Iqgap2-deficient mice is IQGAP1 dependent. Mol. Cell. Biol. 2008, 28, 1489–1502. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Liang, S.; Fang, L.; Chen, L.; Tang, M.; Xu, Y.; Fu, A.; Yang, J.; Wei, Y. Quantitative Proteomic Analysis of HepG2 Cells Treated with Quercetin Suggests IQGAP1 Involved in Quercetin-Induced Regulation of Cell Proliferation and Migration. Omi. A J. Integr. Biol. 2009, 13, 93–103. [Google Scholar] [CrossRef]
- Zoheir, K.M.; Abd-Rabou, A.A.; Harisa, G.I.; Kumar, A.; Ahmad, S.F.; Ansari, M.A.; Abd-Allah, A.R. IQGAP1 gene silencing induces apoptosis and decreases the invasive capacity of human hepatocellular carcinoma cells. Tumor Biol. 2016, 37, 13927–13939. [Google Scholar] [CrossRef]
- Furuta, M.; Kozaki, K.; Tanaka, S.; Arii, S.; Imoto, I.; Inazawa, J. miR-124 and miR-203 are epigenetically silenced tumor-suppressive microRNAs in hepatocellular carcinoma. Carcinogenesis 2010, 31, 766–776. [Google Scholar] [CrossRef] [Green Version]
- Jin, X.; Liu, Y.; Liu, J.; Lu, W.; Liang, Z.; Zhang, D.; Liu, G.; Zhu, H.; Xu, N.; Liang, S. The Overexpression of IQGAP1 and β-Catenin Is Associated with Tumor Progression in Hepatocellular Carcinoma In Vitro and In Vivo. PLoS ONE 2015, 10, e0133770. [Google Scholar] [CrossRef]
- Manmadhan, S.; Ehmer, U. Hippo signaling in the liver—A long and ever-expanding story. Front. Cell Dev. Biol. 2019, 7, 33. [Google Scholar] [CrossRef] [PubMed]
- Anakk, S.; Bhosale, M.; Schmidt, V.A.; Johnson, R.L.; Finegold, M.J.; Moore, D.D. Bile Acids Activate YAP to Promote Liver Carcinogenesis. Cell Rep. 2013, 5, 1060–1069. [Google Scholar] [CrossRef] [Green Version]
- Fram, S.; King, H.; Sacks, D.B.; Wells, C.M. A PAK6-IQGAP1 complex promotes disassembly of cell-cell adhesions. Cell. Mol. Life Sci. 2014, 71, 2759–2773. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.T.; Jiang, Y.Y.; Shang, L.; Shi, Z.Z.; Liang, J.W.; Wang, Z.; Zhang, Y.; Hao, J.J.; Jia, X.M.; Xu, X.; et al. Overexpression of DNAJB6 promotes colorectal cancer cell invasion through an IQGAP1/ERK-dependent signaling pathway. Mol. Carcinog. 2015, 54, 1205–1213. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Yang, Y.; He, Y.; Yang, P.; Wang, X.; He, G.; Zhang, P.; Zhu, H.; Xu, N.; Zhao, X.; et al. SUMOylation of IQGAP1 promotes the development of colorectal cancer. Cancer Lett. 2017, 411, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Zhang, W.; Wu, Y.; Wan, P.; Guo, Q.; Zhang, Y. MiR-124 inhibits cell growth through targeting IQGAP1 in colorectal cancer. Mol. Med. Rep. 2018, 18, 5270–5278. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Xie, C.; Lin, X.; Zhao, Y.; Han, Y.; Fan, C.; Zhang, X.; Du, J.; Han, Y.; Han, Q.; et al. Coexpression of IQ-domain GTPase-activating protein 1 (IQGAP1) and Dishevelled (Dvl) is correlated with poor prognosis in non-small cell lung cancer. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, A.; Wang, F.; Wang, M.; Zhu, M.; Ma, Y.; Wu, R. IQGAP1 activates Tcf signal independent of Rac1 and Cdc42 in injury and repair of bronchial epithelial cells. Exp. Mol. Pathol. 2008, 85, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Yi, J.; Carmon, K.S.; Crumbley, C.A.; Xiong, W.; Thomas, A.; Fan, X.; Guo, S.; An, Z.; Chang, J.T.; et al. Aberrant RSPO3-LGR4 signaling in Keap1-deficient lung adenocarcinomas promotes tumor aggressiveness. Oncogene 2015, 34, 4692–4701. [Google Scholar] [CrossRef] [Green Version]
- Chuang, H.C.; Chang, C.C.; Teng, C.F.; Hsueh, C.H.; Chiu, L.L.; Hsu, P.M.; Lee, M.C.; Hsu, C.P.; Chen, Y.R.; Liu, Y.C.; et al. MAP4K3/GLk promotes lung cancer metastasis by phosphorylating and activating IQGAP1. Cancer Res. 2019, 79, 4978–4993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, S.; Xu, A.; Chen, Y.; Li, W.; Zha, X.; Wang, Y.; Sun, G. Transcriptomic Analysis of Pulmonary Microvascular Endothelial Cells with IQGAP1 Knockdown. DNA Cell Biol. 2020, 39, 1127–1140. [Google Scholar] [CrossRef]
- Sheen, Y.; Lin, M.; Tzeng, W.; Chu, C. Purpuric drug eruptions induced by EGFR tyrosine kinase inhibitors are associated with IQGAP1-mediated increase in vascular permeability. J. Pathol. 2020, 250, 452–463. [Google Scholar] [CrossRef]
- Sugimoto, N.; Imoto, I.; Fukuda, Y.; Kurihara, N.; Kuroda, S.; Tanigami, A.; Kaibuchi, K.; Kamiyama, R.; Inazawa, J.; Sugimoto, N.; et al. IQGAP1, a Negative Regulator of Cell-Cell Adhesion, is Upregulated by Gene Amplification at 15q26 in Gastric Cancer Cell Lines HSC39 and 40A. J. Hum. Genet. 2001, 46, 21–25. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Chen, Y.C.; Sang, J.R.; Xu, W.R. RhoC protein stimulates migration of gastric cancer cells through interaction with scaffold protein IQGAP1. Mol. Med. Rep. 2011, 4, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Tao, Y.; Chen, Y.; Xu, W. RhoC Regulates the Proliferation of Gastric Cancer Cells through Interaction with IQGAP1. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Tseng, P.C.; Chen, C.L.; Shan, Y.S.; Chang, W.T.; Liu, H.S.; Hong, T.M.; Hsieh, C.Y.; Lin, S.H.; Lin, C.F. An increase in integrin-linked kinase noncanonically confers NF-κB-mediated growth advantages to gastric cancer cells by activating ERK1/2. Cell Commun. Signal. 2014, 12. [Google Scholar] [CrossRef] [Green Version]
- Bessède, E.; Molina, S.; Amador, L.A.; Dubus, P.; Staedel, C.; Chambonnier, L.; Buissonnière, A.; Sifré, E.; Giese, A.; Bénéjat, L.; et al. Deletion of IQGAP1 promotes Helicobacter pylori-induced gastric dysplasia in mice and acquisition of cancer stem cell properties in vitro. Oncotarget 2016, 7, 80688–80699. [Google Scholar] [CrossRef] [Green Version]
- Pan, C.; Jin, X.; Zhao, Y.; Pan, Y.; Yang, J.; Karnes, R.J.; Zhang, J.; Wang, L.; Huang, H. AKT -phosphorylated FOXO 1 suppresses ERK activation and chemoresistance by disrupting IQGAP 1- MAPK interaction. EMBO J. 2017, 36, 995–1010. [Google Scholar] [CrossRef] [PubMed]
- Arienti, C.; Zanoni, M.; Pignatta, S.; Del Rio, A.; Carloni, S.; Tebaldi, M.; Tedaldi, G.; Tesei, A. Preclinical evidence of multiple mechanisms underlying trastuzumab resistance in gastric cancer. Oncotarget 2016, 7, 18424–18439. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Li, Q.; Li, H.; Wang, Y.; Wang, H.; Chen, W.; Zhang, S.; Cao, J.; Liu, T. Asparaginyl endopeptidase improves the resistance of microtubule-targeting drugs in gastric cancer through IQGAP1 modulating the EGFR/JNK/ERERK signaling pathway. Onco. Targets Ther. 2017, 10, 627–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, S.M. Estrogen, progesterone and epithelial ovarian cancer. Reprod. Biol. Endocrinol. 2003, 1, 73. [Google Scholar] [CrossRef] [Green Version]
- Jacquemet, G.; Green, D.M.; Bridgewater, R.E.; von Kriegsheim, A.; Humphries, M.J.; Norman, J.C.; Caswell, P.T. Rcp-driven α5β1 recycling suppresses rac and promotes rhoa activity via the racgap1-iqgap1 complex. J. Cell Biol. 2013, 202, 917–935. [Google Scholar] [CrossRef] [Green Version]
- Dong, P.X.; Jia, N.; Xu, Z.J.; Liu, Y.T.; Li, D.J.; Feng, Y.J. Silencing of IQGAP1 by shRNA inhibits the invasion of ovarian carcinoma HO-8910PM cells in vitro. J. Exp. Clin. Cancer Res. 2008, 27, 77. [Google Scholar] [CrossRef] [Green Version]
- Bourguignon, L.Y.W.; Gilad, E.; Rothman, K.; Peyrollier, K. Hyaluronan-CD44 interaction with IQGAP1 promotes Cdc42 and ERK signaling, leading to actin binding, Elk-1/estrogen receptor transcriptional activation, and ovarian cancer progression. J. Biol. Chem. 2005, 280, 11961–11972. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Xu, S.; Hu, D.; Lu, W.; Xie, X.; Cheng, X. IQGAP1 is involved in enhanced aggressive behavior of epithelial ovarian cancer stem cell-like cells during differentiation. Int. J. Gynecol. Cancer 2015, 25, 559–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chellini, L.; Caprara, V.; Spadaro, F.; Sestito, R.; Bagnato, A.; Rosanò, L. Regulation of extracellular matrix degradation and metastatic spread by IQGAP1 through endothelin-1 receptor signalling in ovarian cancer. Matrix Biol. 2019, 81, 17–33. [Google Scholar] [CrossRef] [PubMed]
- Angus, K.L.; Griffiths, G.M. Cell polarisation and the immunological synapse. Curr. Opin. Cell Biol. 2013, 25, 85. [Google Scholar] [CrossRef] [Green Version]
- PV, P.; DD, B. Molecular regulation of the plasma membrane-proximal cellular steps involved in NK cell cytolytic function. J. Cell Sci. 2020, 133. [Google Scholar] [CrossRef] [Green Version]
- Ritter, A.T.; Angus, K.L.; Griffiths, G.M. The role of the cytoskeleton at the immunological synapse. Immunol. Rev. 2013, 256, 107–117. [Google Scholar] [CrossRef] [Green Version]
- TS, G.; DD, B. T cell activation and the cytoskeleton: You can’t have one without the other. Adv. Immunol. 2008, 97, 1–64. [Google Scholar] [CrossRef]
- Wang, J.C.; Lee, J.Y.-J.; Christian, S.; Dang-Lawson, M.; Pritchard, C.; Freeman, S.A.; Gold, M.R. The Rap1–cofilin-1 pathway coordinates actin reorganization and MTOC polarization at the B cell immune synapse. J. Cell Sci. 2017, 130, 1094–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorman, J.A.; Babich, A.; Dick, C.J.; Schoon, R.A.; Koenig, A.; Gomez, T.S.; Burkhardt, J.K.; Billadeau, D.D. The Cytoskeletal Adaptor Protein IQGAP1 Regulates TCR-Mediated Signaling and Filamentous Actin Dynamics. J. Immunol. 2012. [Google Scholar] [CrossRef] [Green Version]
- Awasthi, A.; Samarakoon, A.; Chu, H.; Kamalakannan, R.; Quilliam, L.A.; Chrzanowska-Wodnicka, M.; White, G.C.; Malarkannan, S. Rap1b facilitates NK cell functions via IQGAP1-mediated signalosomes. J. Exp. Med. 2010, 207, 1923–1938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, L.K.; Philip, N.H.; Schmidt, V.A.; Koller, A.; Strowig, T.; Flavell, R.A.; Brodsky, I.E.; Bliska, J.B. IQGAP1 is important for activation of caspase-1 in macrophages and is targeted by Yersinia pestis type III effector YopM. MBio 2014, 5, e01402-14. [Google Scholar] [CrossRef] [Green Version]
- Neel, N.F.; Sai, J.; Ham, A.-J.L.; Sobolik-Delmaire, T.; Mernaugh, R.L.; Richmond, A. IQGAP1 is a novel CXCR2-interacting protein and essential component of the “chemosynapse”. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [Green Version]
- Abel, A.M.; Tiwari, A.A.; Gerbec, Z.J.; Siebert, J.R.; Yang, C.; Schloemer, N.J.; Dixon, K.J.; Thakar, M.S.; Malarkannan, S. IQ domain-containing GTPase-activating protein 1 regulates cytoskeletal reorganization and facilitates NKG2D-mediated mechanistic target of rapamycin complex 1 activation and cytokine gene translation in natural killer cells. Front. Immunol. 2018. [Google Scholar] [CrossRef]
- Brandt, D.T.; Marion, S.; Griffiths, G.; Watanabe, T.; Kaibuchi, K.; Grosse, R. Dia1 and IQGAP1 interact in cell migration and phagocytic cup formation. J. Cell Biol. 2007, 178, 193–200. [Google Scholar] [CrossRef]
- Okada, M.; Hozumi, Y.; Iwazaki, K.; Misaki, K.; Yanagida, M.; Araki, Y.; Watanabe, T.; Yagisawa, H.; Topham, M.K.; Kaibuchi, K.; et al. DGKζ is involved in LPS-activated phagocytosis through IQGAP1/Rac1 pathway. Biochem. Biophys. Res. Commun. 2012, 420, 479–484. [Google Scholar] [CrossRef]
- Kanwar, N.; Wilkins, J.A. IQGAP1 involvement in MTOC and granule polarization in NK-cell cytotoxicity. Eur. J. Immunol. 2011. [Google Scholar] [CrossRef]
- Urao, N.; Razvi, M.; Oshikawa, J.; McKinney, R.D.; Chavda, R.; Bahou, W.F.; Fukai, T.; Ushio-Fukai, M. IQGAP1 is involved in post-ischemic neovascularization by regulating angiogenesis and macrophage infiltration. PLoS ONE 2010, 5. [Google Scholar] [CrossRef] [PubMed]
- Beemiller, P.; Krummel, M.F. Mediation of T-Cell Activation by Actin Meshworks. Cold Spring Harb. Perspect. Biol. 2010, 2. [Google Scholar] [CrossRef] [Green Version]
- Ostuni, R.; Kratochvill, F.; Murray, P.J.; Natoli, G. Macrophages and cancer: From mechanisms to therapeutic implications. Trends Immunol. 2015, 36, 229–239. [Google Scholar] [CrossRef]
- Mordoh, J.; Levy, E.M.; Roberti, M.P. Natural killer cells in human cancer: From biological functions to clinical applications. J. Biomed. Biotechnol. 2011, 2011, 1–11. [Google Scholar]
- Sharma, S.; Findlay, G.M.; Bandukwala, H.S.; Oberdoerffer, S.; Baust, B.; Li, Z.; Schmidt, V.; Hogan, P.G.; Sacks, D.B.; Rao, A. Dephosphorylation of the nuclear factor of activated T cells (NFAT) transcription factor is regulated by an RNA-protein scaffold complex. Proc. Natl. Acad. Sci. USA 2011. [Google Scholar] [CrossRef] [Green Version]
- Okuyama, Y.; Nagashima, H.; Ushio-Fukai, M.; Croft, M.; Ishii, N.; So, T. IQGAP1 restrains T-cell cosignaling mediated by OX40. FASEB J. 2020. [Google Scholar] [CrossRef] [Green Version]
- Aspeslagh, S.; Postel-Vinay, S.; Rusakiewicz, S.; Soria, J.C.; Zitvogel, L.; Marabelle, A. Rationale for anti-OX40 cancer immunotherapy. Eur. J. Cancer 2016, 52, 50–66. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Cancer Type | Method | Target | Comparison | Prognostic * | References |
|---|---|---|---|---|---|
| Breast | IB | protein | cancer vs. normal | yes | [65,66] |
| IHC | protein | cancer vs. normal | |||
| RT-PCR | mRNA | cancer vs. normal | |||
| IHC | protein | high-grade vs. low-grade cancer | |||
| Head & neck | MS | protein | cancer vs. normal | yes | [51,67,68] |
| IHC | protein | cancer vs. normal | |||
| Pancreas | IHC | protein | cancer vs. normal | yes | [69,70] |
| IB | protein | cancer vs. normal | |||
| IHC | protein | high-grade vs. low-grade cancer | |||
| Liver | IHC | protein | cancer vs. normal | yes | [46,71,72] |
| IB | protein | cancer vs. normal | |||
| Colorectal | Array | gene | cancer vs. normal | yes | [73,74,75] |
| IHC | protein | cancer vs. normal | |||
| Gastric | IB | protein | cancer vs. normal | no | [76,77] |
| IHC | protein | high-grade vs. low-grade cancer | |||
| Lung | RT-PCR | mRNA | cancer vs. normal | yes | [63,78,79,80] |
| IHC | protein | high-grade vs. low-grade cancer | |||
| Ovary | IHC | protein | adenocarcinoma vs. adenoma | yes | [64] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, T.; Lambert, P.F. Role of IQGAP1 in Carcinogenesis. Cancers 2021, 13, 3940. https://doi.org/10.3390/cancers13163940
Wei T, Lambert PF. Role of IQGAP1 in Carcinogenesis. Cancers. 2021; 13(16):3940. https://doi.org/10.3390/cancers13163940
Chicago/Turabian StyleWei, Tao, and Paul F. Lambert. 2021. "Role of IQGAP1 in Carcinogenesis" Cancers 13, no. 16: 3940. https://doi.org/10.3390/cancers13163940
APA StyleWei, T., & Lambert, P. F. (2021). Role of IQGAP1 in Carcinogenesis. Cancers, 13(16), 3940. https://doi.org/10.3390/cancers13163940