
Mechanisms Regulating Abnormal Circular RNA Biogenesis in Cancer



Abstract
:Simple Summary
Abstract
1. Introduction
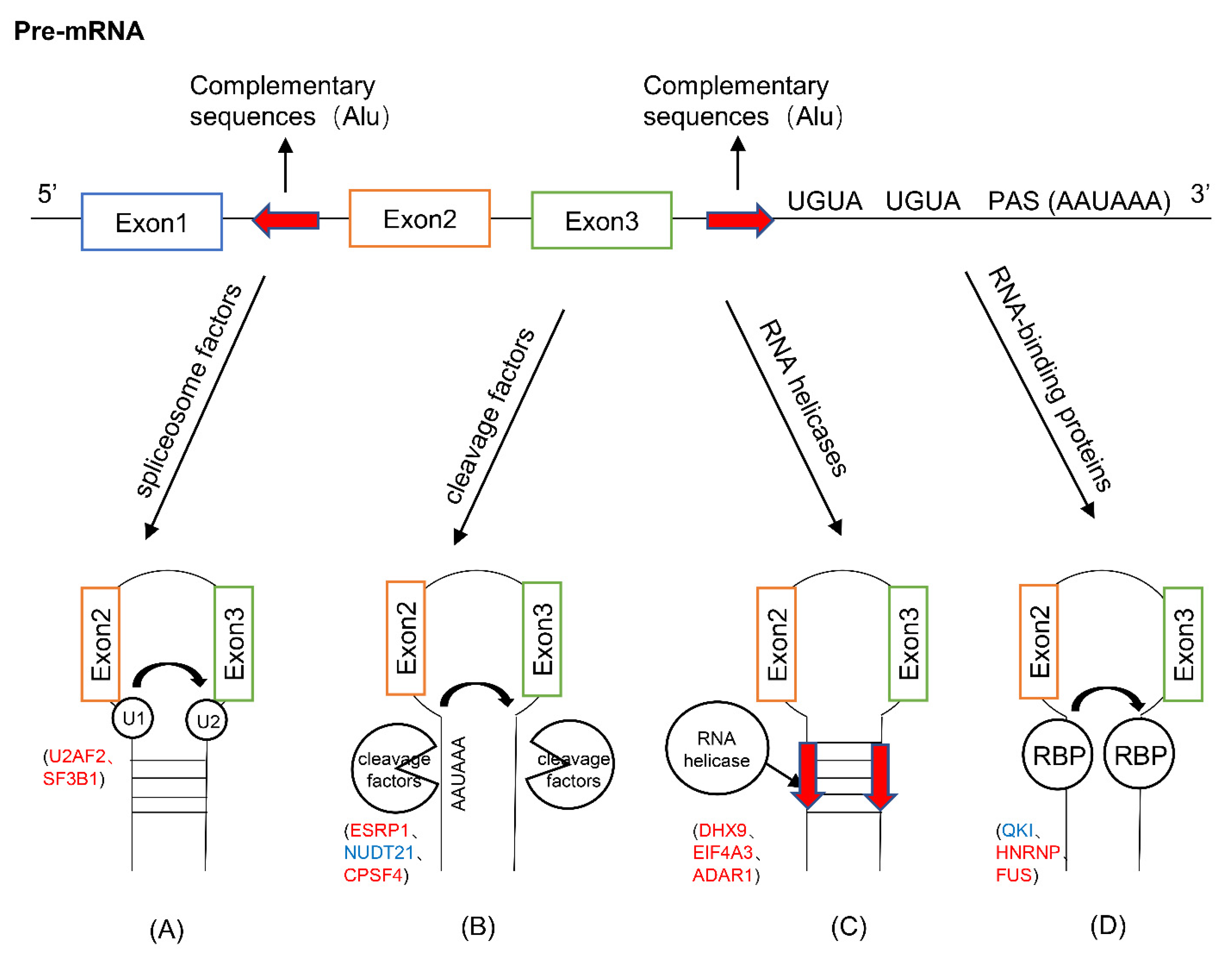
2. The Biogenesis of circRNAs
3. Trans-Acting Proteins and RNAs
3.1. Spliceosome Factors
3.2. Cleavage Factors
3.3. RNA Helicases
3.4. RNA-Binding Proteins
4. Cis-Regulatory Elements
4.1. Intronic Complementary Sequences
4.2. Splice Sites
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Meng, S.; Zhou, H.; Feng, Z.; Xu, Z.; Tang, Y.; Li, P.; Wu, M. CircRNA: Functions and properties of a novel potential biomarker for cancer. Mol. Cancer 2017, 16, 94. [Google Scholar] [CrossRef]
- Qu, S.; Zhong, Y.; Shang, R.; Zhang, X.; Song, W.; Kjems, J.; Li, H. The emerging landscape of circular RNA in life processes. RNA Biol. 2017, 14, 992–999. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Zhou, W.; Li, X.; Sun, M.; Ding, J.; Zhu, Q. Tandem DNAzyme for double digestion: A new tool for circRNA suppression. Biol. Chem. 2019, 400, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Wilusz, J.E.; Sharp, P.A. Molecular biology. A circuitous route to noncoding RNA. Science 2013, 340, 440–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhang, X.O.; Chen, T.; Xiang, J.F.; Yin, Q.F.; Xing, Y.H.; Zhu, S.; Yang, L.; Chen, L.L. Circular intronic long noncoding RNAs. Mol. Cell 2013, 51, 792–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Shan, G. CircRNA in cancer: Fundamental mechanism and clinical potential. Cancer Lett. 2021, 505, 49–57. [Google Scholar] [CrossRef]
- Meng, X.; Chen, Q.; Zhang, P.; Chen, M. CircPro: An integrated tool for the identification of circRNAs with protein-coding potential. Bioinformatics 2017, 33, 3314–3316. [Google Scholar] [CrossRef]
- Wang, X.; Li, H.J.; Lu, Y.J.; Cheng, L.M. Circular RNAs in Human Cancer. Front. Oncol. 2021, 10, 3102. [Google Scholar] [CrossRef]
- Zhang, H.D.; Jiang, L.H.; Sun, D.W.; Hou, J.C.; Ji, Z.L. CircRNA: A novel type of biomarker for cancer. Breast Cancer 2018, 25, 1–7. [Google Scholar] [CrossRef]
- Wei, G.; Zhu, J.; Hu, H.B.; Liu, J.Q. Circular RNAs: Promising biomarkers for cancer diagnosis and prognosis. Gene 2021, 771, 145365. [Google Scholar] [CrossRef] [PubMed]
- Qu, S.; Yang, X.; Li, X.; Wang, J.; Gao, Y.; Shang, R.; Sun, W.; Dou, K.; Li, H. Circular RNA: A new star of noncoding RNAs. Cancer Lett. 2015, 365, 141–148. [Google Scholar] [CrossRef]
- Chen, L.L.; Yang, L. Regulation of circRNA biogenesis. RNA Biol. 2015, 12, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, L.; Chen, L.L. The Biogenesis, Functions, and Challenges of Circular RNAs. Mol. Cell 2018, 71, 428–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pervouchine, D.D. Circular exonic RNAs: When RNA structure meets topology. BBA-Gene Regul. Mech. 2019, 1862, 194384. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Tatomer, D.C.; Luo, Z.; Wu, H.; Yang, L.; Chen, L.L.; Cherry, S.; Wilusz, J.E. The Output of Protein-Coding Genes Shifts to Circular RNAs When the Pre-mRNA Processing Machinery Is Limiting. Mol. Cell 2017, 68, 940–954.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzo, E.; Stefano, D.M.; Pietro, L.; Alessio, C.; Ivano, L.; Davide, C.; Alessandro, R.; Riccardo, D.S.; Rebecca, S.; Giovanna, P.; et al. FUS affects circular RNA expression in murine embryonic stem cell-derived motor neurons. Nat. Commun. 2017, 8, 14741. [Google Scholar]
- Gruber, A.J.; Zavolan, M. Alternative cleavage and polyadenylation in health and disease. Nat. Rev. Genet. 2019, 20, 599–614. [Google Scholar] [CrossRef]
- Li, J.; Sun, D.; Pu, W.; Wang, J.; Peng, Y. Circular RNAs in Cancer: Biogenesis, Function, and Clinical Significance. Trends Cancer 2020, 6, 319–336. [Google Scholar] [CrossRef]
- Zhang, X.O.; Wang, H.B.; Zhang, Y.; Lu, X.; Chen, L.L.; Yang, L. Complementary sequence-mediated exon circularization. Cell 2014, 159, 134–147. [Google Scholar] [CrossRef] [Green Version]
- Lasda, E.; Parker, R. Circular RNAs: Diversity of form and function. RNA 2014, 20, 1829–1842. [Google Scholar] [CrossRef] [Green Version]
- Tang, T.H.; Rozhdestvensky, T.S.; d’Orval, B.C.; Bortolin, M.L.; Huber, H.; Charpentier, B.; Branlant, C.; Bachellerie, J.P.; Brosius, J.; Huttenhofer, A. RNomics in Archaea reveals a further link between splicing of archaeal introns and rRNA processing. Nucleic Acids Res. 2002, 30, 921–930. [Google Scholar] [CrossRef]
- Flores, R.; Navarro, J.A.; de la Pena, M.; Navarro, B.; Ambros, S.; Vera, A. Viroids with hammerhead ribozymes: Some unique structural and functional aspects with respect to other members of the group. Biol. Chem. 1999, 380, 849–854. [Google Scholar] [CrossRef] [Green Version]
- Karousi, P.; Artemaki, P.I.; Sotiropoulou, C.D.; Christodoulou, S.; Scorilas, A.; Kontos, C.K. Identification of Two Novel Circular RNAs Deriving from BCL2L12 and Investigation of Their Potential Value as a Molecular Signature in Colorectal Cancer. Int. J. Mol. Sci. 2020, 21, 8867. [Google Scholar] [CrossRef]
- Jeck, W.R.; Sharpless, N.E. Detecting and characterizing circular RNAs. Nat. Biotechnol. 2014, 32, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Ashwal-Fluss, R.; Meyer, M.; Pamudurti, N.R.; Ivanov, A.; Bartok, O.; Hanan, M.; Evantal, N.; Memczak, S.; Rajewsky, N.; Kadener, S. circRNA biogenesis competes with pre-mRNA splicing. Mol. Cell 2014, 56, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA 2013, 19, 141–157. [Google Scholar] [CrossRef] [Green Version]
- Conn, S.J.; Pillman, K.A.; Toubia, J.; Conn, V.M.; Salmanidis, M.; Phillips, C.A.; Roslan, S.; Schreiber, A.W.; Gregory, P.A.; Goodall, G.J. The RNA binding protein quaking regulates formation of circRNAs. Cell 2015, 160, 1125–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahebi, M.; Hanafi, M.M.; van Wijnen, A.J.; Azizi, P.; Abiri, R.; Ashkani, S.; Taheri, S. Towards understanding pre-mRNA splicing mechanisms and the role of SR proteins. Gene 2016, 587, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.C.; Cheng, S.C. Functional roles of protein splicing factors. Biosci. Rep. 2012, 32, 345–359. [Google Scholar] [CrossRef] [Green Version]
- Kramer, M.C.; Liang, D.; Tatomer, D.C.; Gold, B.; March, Z.M.; Cherry, S.; Wilusz, J.E. Combinatorial control of Drosophila circular RNA expression by intronic repeats, hnRNPs, and SR proteins. Genes Dev. 2015, 29, 2168–2182. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, H.; Nobe, Y.; Izumikawa, K.; Taoka, M.; Yamauchi, Y.; Nakayama, H.; Simpson, R.J.; Isobe, T.; Takahash, N. Truncated forms of U2 snRNA (U2-tfs) are shunted toward a novel uridylylation pathway that differs from the degradation pathway for U1-tfs. RNA Biol. 2018, 15, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Feng, S.; Liu, G.; Wang, H.; Fu, A.; Zhu, H.; Ren, Q.; Wang, B.; Xu, X.; Bai, H.; et al. CD82 suppresses CD44 alternative splicing-dependent melanoma metastasis by mediating U2AF2 ubiquitination and degradation. Oncogene 2016, 35, 5056–5069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Cheng, D.; Zhu, M.; Yu, H.; Pan, Z.; Liu, L.; Geng, Q.; Pan, H.; Yan, M.; Yao, M. OTUB2 stabilizes U2AF2 to promote the Warburg effect and tumorigenesis via the AKT/mTOR signaling pathway in non-small cell lung cancer. Theranostics 2019, 9, 179–195. [Google Scholar] [CrossRef] [PubMed]
- Chul-Kee, P.; Inho, P.; Seungmook, L.; Choong-Hyun, S.; Youngil, K.; Sung-Hye, P.; Eun, K.J.; Hongseok, Y.; Se-Hoon, L. Genomic dynamics associated with malignant transformation in IDH1 mutated gliomas. Oncotarget 2015, 6, 43653–43666. [Google Scholar]
- Marzese, D.M.; Liu, M.; Huynh, J.L.; Hirose, H.; Donovan, N.C.; Huynh, K.T.; Kiyohara, E.; Chong, K.; Cheng, D.; Tanaka, R.; et al. Brain metastasis is predetermined in early stages of cutaneous melanoma by CD44v6 expression through epigenetic regulation of the spliceosome. Pigment. Cell Melanoma Res. 2015, 28, 82–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Jinpeng, Z.; Junshuang, Z.; Haiying, Z.; Long, L.; Hao, L.; Lian, C.; Jiangfeng, H.; Wei, Z.; Zhitao, J. The U2AF2 /circRNA ARF1/miR-342-3p/ISL2 feedback loop regulates angiogenesis in glioma stem cells. J. Exp. Clin. Cancer Res. 2020, 39, 182. [Google Scholar]
- Zhou, Z.; Gong, Q.; Wang, Y.; Li, M.; Wang, L.; Ding, H.; Li, P. The biological function and clinical significance of SF3B1 mutations in cancer. Biomark. Res. 2020, 8, 38. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Hussaini, M.; Qin, D.; Zhang, X.; Shao, H.; Zhang, L.; Gajzer, D.; Basra, P.; Moscinski, L.; Zhang, H. Comparison of SF3B1/DNMT3A Comutations with DNMT3A or SF3B1 Mutation Alone in Myelodysplastic Syndrome and Clonal Cytopenia of Undetermined Significance. Am. J. Clin. Pathol. 2020, 154, 48–56. [Google Scholar] [CrossRef]
- Bergot, T.; Lippert, E.; Douet-Guilbert, N.; Commet, S.; Corcos, L.; Bernard, D.G. Human Cancer-Associated Mutations of SF3B1 Lead to a Splicing Modification of Its Own RNA. Cancers 2020, 12, 652. [Google Scholar] [CrossRef] [Green Version]
- Hou, C.; Xiao, L.; Ren, X.; Tang, F.; Guo, B.; Zeng, W.; Liang, C.; Yan, N. Mutations of GNAQ, GNA11, SF3B1, EIF1AX, PLCB4 and CYSLTR in Uveal Melanoma in Chinese Patients. Ophthalmic Res. 2020, 63, 358–368. [Google Scholar] [CrossRef]
- Küsters-Vandevelde, H.V.; Creytens, D.; van Engen-van Grunsven, A.C.; Jeunink, M.; Winnepenninckx, V.; Groenen, P.J.; Küsters, B.; Wesseling, P.; Blokx, W.A.; Prinsen, C.F. SF3B1 and EIF1AX mutations occur in primary leptomeningeal melanocytic neoplasms; yet another similarity to uveal melanomas. Acta Neuropathol. Commun. 2016, 4, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, Y.; Krauthammer, M.; Halaban, R. Rare SF3B1 R625 mutations in cutaneous melanoma. Melanoma Res. 2014, 24, 332–334. [Google Scholar] [CrossRef] [Green Version]
- Kaixuan, Z.; Bangshun, H.; Yang, B.B.; Tao, X.; Xiaoxiang, C.; Mu, X.; Xiangxiang, L.; Huiling, S.; Yuqin, P.; Shukui, W. The pro-metastasis effect of circANKS1B in breast cancer. Mol. Cancer 2018, 17, 160. [Google Scholar]
- Zhao, W.; Cui, Y.; Liu, L.; Qi, X.; Liu, J.; Ma, S.; Hu, X.; Zhang, Z.; Wang, Y.; Li, H.; et al. Splicing factor derived circular RNA circUHRF1 accelerates oral squamous cell carcinoma tumorigenesis via feedback loop. Cell Death Differ. 2020, 27, 919–933. [Google Scholar] [CrossRef]
- Yang, Q.; Coseno, M.; Gilmartin, G.M.; Doublié, S. Crystal structure of a human cleavage factor CFI(m)25/CFI(m)68/RNA complex provides an insight into poly(A) site recognition and RNA looping. Structure 2011, 19, 368–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, S.; Li, H.; Zhang, W.; Shao, Y.; Liu, Y.; Guan, H.; Wu, J.; Kang, Y.; Zhao, J.; Yu, Q.; et al. NUDT21 negatively regulates PSMB2 and CXXC5 by alternative polyadenylation and contributes to hepatocellular carcinoma suppression. Oncogene 2018, 37, 4887–4900. [Google Scholar] [CrossRef]
- Li, X.; Ding, J.; Wang, X.; Cheng, Z.; Zhu, Q. NUDT21 regulates circRNA cyclization and ceRNA crosstalk in hepatocellular carcinoma. Oncogene 2020, 39, 891–904. [Google Scholar] [CrossRef]
- Wang, X.; Dong, J.; Li, X.; Cheng, Z.; Zhu, Q. CPSF4 regulates circRNA formation and microRNA mediated gene silencing in hepatocellular carcinoma. Oncogene 2021, 40, 4338–4351. [Google Scholar] [CrossRef]
- Aktaş, T.; Ilık, İ.A.; Maticzka, D.; Bhardwaj, V.; Rodrigues, C.P.; Mittler, G.; Manke, T.; Backofen, R.; Akhtar, A. DHX9 suppresses RNA processing defects originating from the Alu invasion of the human genome. Nature 2017, 544, 115–119. [Google Scholar] [CrossRef]
- Yingkuan, L.; Hui, W.; Bing, C.; Qixing, M.; Wenjie, X.; Te, Z.; Xuming, S.; Zeyu, Z.; Lin, X.; Gaochao, D.; et al. circDCUN1D4 suppresses tumor metastasis and glycolysis in lung adenocarcinoma by stabilizing TXNIP expression. Mol. Ther.-Nucleic Acids 2021, 23, 355–368. [Google Scholar]
- Yan, D.; Dong, W.; He, Q.; Yang, M.; Huang, L.; Kong, J.; Qin, H.; Lin, T.; Huang, J. Circular RNA circPICALM sponges miR-1265 to inhibit bladder cancer metastasis and influence FAK phosphorylation. EBioMedicine 2019, 48, 316–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jian, Y.; Qing-Guo, X.; Zhen-Guang, W.; Yuan, Y.; Ling, Z.; Jin-Zhao, M.; Shu-Han, S.; Fu, Y.; Wei-Ping, Z. Circular RNA cSMARCA5 inhibits growth and metastasis in hepatocellular carcinoma. J. Hepatol. 2018, 68, 1214–1227. [Google Scholar]
- Linder, P.; Jankowsky, E. From unwinding to clamping—The DEAD box RNA helicase family. Nat. Rev. Mol. Cell Biol. 2011, 12, 505–516. [Google Scholar] [CrossRef] [Green Version]
- Noble, C.G.; Song, H. MLN51 Stimulates the RNA-Helicase Activity of eIF4AIII. PLoS ONE 2007, 2, e303. [Google Scholar] [CrossRef] [Green Version]
- Ballut, L.; Marchadier, B.; Baguet, A.; Tomasetto, C.; Séraphin, B.; Hir, H.L. The exon junction core complex is locked onto RNA by inhibition of eIF4AIII ATPase activity. Nat. Struct. Mol. Biol. 2005, 12, 861–869. [Google Scholar] [CrossRef]
- Wang, R.; Zhang, S.; Chen, X.; Li, N.; Li, J.; Jia, R.; Pan, Y.; Liang, H. EIF4A3-induced circular RNA MMP9 (circMMP9) acts as a sponge of miR-124 and promotes glioblastoma multiforme cell tumorigenesis. Mol. Cancer 2018, 17, 166. [Google Scholar] [CrossRef]
- Yutian, W.; Chenfei, L.; Peng, Z.; Lin, Z.; Xiao, L.; Jianxing, Y.; ZhuMei, S.; Yongping, Y. EIF4A3-induced circular RNA ASAP1 promotes tumorigenesis and temozolomide resistance of glioblastoma via NRAS/MEK1/ERK1–2 signaling. Neuro-Oncol. 2021, 23, 611–624. [Google Scholar]
- Zheng, X.; Huang, M.; Xing, L.; Yang, R.; Wang, X.; Jiang, R.; Zhang, L.; Chen, J. The circRNA circSEPT9 mediated by E2F1 and EIF4A3 facilitates the carcinogenesis and development of triple-negative breast cancer. Mol. Cancer 2020, 19, 73. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, A.; Memczak, S.; Wyler, E.; Torti, F.; Porath, H.T.; Orejuela, M.R.; Piechotta, M.; Levanon, E.Y.; Landthaler, M.; Dieterich, C.; et al. Analysis of intron sequences reveals hallmarks of circular RNA biogenesis in animals. Cell Rep. 2015, 10, 170–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, L.; Yan, P.; Liang, Y.; Sun, Y.; Shen, J.; Zhou, S.; Lin, H.; Liang, X.; Cai, X. Circular RNA expression is suppressed by androgen receptor (AR)-regulated adenosine deaminase that acts on RNA (ADAR1) in human hepatocellular carcinoma. Cell Death Dis. 2017, 8, e3171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, C.; Wang, X.; Yang, F.; Zang, Y.; Liu, J.; Wang, X.; Xu, X.; Li, W.; Jia, J.; Liu, Z. Circular RNA hsa_circ_0004872 inhibits gastric cancer progression via the miR-224/Smad4/ADAR1 successive regulatory circuit. Mol. Cancer 2020, 19, 157. [Google Scholar] [CrossRef] [PubMed]
- Shen, P.; Yang, T.; Chen, Q.; Yuan, H.; Wu, P.; Cai, B.; Meng, L.; Huang, X.; Liu, J.; Zhang, Y.; et al. CircNEIL3 regulatory loop promotes pancreatic ductal adenocarcinoma progression via miRNA sponging and A-to-I RNA-editing. Mol. Cancer 2021, 20, 51. [Google Scholar] [CrossRef]
- Zhao, X.; Cai, Y.; Xu, J. Circular RNAs: Biogenesis, Mechanism, and Function in Human Cancers. Int. J. Mol. Sci. 2019, 20, 3926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fei, T.; Chen, Y.; Xiao, T.; Li, W.; Cato, L.; Zhang, P.; Cotter, M.B.; Bowden, M.; Lis, R.T.; Zhao, S.G.; et al. Genome-wide CRISPR screen identifies HNRNPL as a prostate cancer dependency regulating RNA splicing. Proc. Natl. Acad. Sci. USA 2017, 114, E5207–E5215. [Google Scholar] [CrossRef] [Green Version]
- Jiacheng, C.; Xing, Z.; Penghui, X.; Haixiao, W.; Sen, W.; Lu, Z.; Zheng, L.; Li, X.; Guangli, S.; Yiwen, X.; et al. Circular RNA circLMO7 acts as a microRNA-30a-3p sponge to promote gastric cancer progression via the WNT2/β-catenin pathway. J. Exp. Clin. Cancer Res. 2021, 40, 6. [Google Scholar]
- Stark, H.; Dube, P.; Lührmann, R.; Kastner, B. Arrangement of RNA and proteins in the spliceosomal U1 small nuclear ribonucleoprotein particle. Nature 2001, 409, 539–542. [Google Scholar] [CrossRef]
- Wu, J.I.; Reed, R.B.; Grabowski, P.J.; Artzt, K. Function of quaking in Myelination: Regulation of Alternative Splicing. Proc. Natl. Acad. Sci. USA 2002, 99, 4233–4238. [Google Scholar] [CrossRef] [Green Version]
- Teplova, M.; Hafner, M.; Teplov, D.; Essig, K.; Tuschl, T.; Patel, D.J. Structure–function studies of STAR family Quaking proteins bound to their in vivo RNA target sites. Genes Dev. 2013, 27, 928–940. [Google Scholar] [CrossRef] [Green Version]
- Yan-Jing, Z.; Bo, Z.; Gui-Juan, L.; Xu-Kai, M.; Xin-Yuan, L.; Xi-Meng, L.; Shuai, Y.; Qing, Z.; Tong, W.; Zhi-Xuan, L.; et al. Circular RNAs negatively regulate cancer stem cells by physically binding FMRP against CCAR1 complex in hepatocellular carcinoma. Theranostics 2019, 9, 3526–3540. [Google Scholar]
- Artemaki, P.I.; Scorilas, A.; Kontos, C.K. Circular RNAs: A New Piece in the Colorectal Cancer Puzzle. Cancers 2020, 12, 2464. [Google Scholar] [CrossRef]
- Han, K.; Wang, F.-W.; Cao, C.-H.; Ling, H.; Chen, J.-W.; Chen, R.-X.; Feng, Z.-H.; Luo, J.; Jin, X.-H.; Duan, J.-L.; et al. CircLONP2 enhances colorectal carcinoma invasion and metastasis through modulating the maturation and exosomal dissemination of microRNA-17. Mol. Cancer 2020, 19, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaoxia, L.; Ningning, Y.; Xu, J.; Jian, W.; Junqiang, D.; Yonghua, G. FUS-induced circular RNA ZNF609 promotes tumorigenesis and progression via sponging miR-142-3p in lung cancer. J. Cell. Physiol. 2020, 236, 79–92. [Google Scholar]
- Tong, C.; Xiaolong, W.; Chen, L.; Hanwen, Z.; Ying, L.; Dianwen, H.; Yaming, L.; Zheng, L.; Dan, L.; Ning, Z.; et al. CircHIF1A regulated by FUS accelerates triple-negative breast cancer progression by modulating NFIB expression and translocation. Oncogene 2021, 40, 2756–2771. [Google Scholar]
- Zhang, X.O.; Dong, R.; Zhang, Y.; Zhang, J.L.; Luo, Z.; Zhang, J.; Chen, L.L.; Yang, L. Diverse alternative back-splicing and alternative splicing landscape of circular RNAs. Genome Res. 2016, 26, 1277–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, D.; Wilusz, J.E. Short intronic repeat sequences facilitate circular RNA production. Genes Dev. 2014, 28, 2233–2247. [Google Scholar] [CrossRef] [Green Version]
- Jin, C.; Zhao, J.; Zhang, Z.P.; Wu, M.; Li, J.; Liu, B.; Bin Liao, X.; Liao, Y.X.; Liu, J.P. CircRNA EPHB4 modulates stem properties and proliferation of gliomas via sponging miR-637 and up-regulating SOX10. Mol. Oncol. 2021, 15, 596–622. [Google Scholar] [CrossRef]
- Proudfoot, N.J. Ending the message: Poly(A) signals then and now. Genes Dev. 2011, 25, 1770–1782. [Google Scholar] [CrossRef] [Green Version]
- Millevoi, S.; Vagner, S. Molecular mechanisms of eukaryotic pre-mRNA 3′ end processing regulation. Nucleic Acids Res. 2010, 38, 2757–2774. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Ding, J.; Li, D.; Yang, G.; Cheng, Z.; Zhu, Q. NUDT21 regulates 3′-UTR length and microRNA-mediated gene silencing in hepatocellular carcinoma. Cancer Lett. 2017, 410, 158–168. [Google Scholar] [CrossRef]
- Papatsirou, M.; Artemaki, P.I.; Scorilas, A.; Kontos, C.K. The role of circular RNAs in therapy resistance of patients with solid tumors. Pers. Med. 2020, 17, 469–490. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Trans-Acting Proteins | Proteins/RNA | Target circRNAs | Correlation | Cancer | Ref. |
|---|---|---|---|---|---|
| spliceosome factors | U2AF | cARF1 | Positive | Glioma | [37] |
| cleavage factors | ESRP1 | circANKS1B | Positive | Breast cancer | [44] |
| circUHRF1 | Positive | Oral squamous cell carcinoma | [45] | ||
| NUDT21 | circRNAs with UGUA | Positive | Hepatocellular carcinoma | [48] | |
| CPSF4 | circRNAs with PAS | Negative | Hepatocellular carcinoma | [49] | |
| RNA helicases | DHX9 | circDCUN1D4 | Negative | Lung adenocarcinoma | [51] |
| circPICALM | Negative | Bladder cancer | [52] | ||
| cSMARCA5 | Negative | Hepatocellular carcinoma | [53] | ||
| EIF4A3 | circMMP9 | Positive | Glioblastoma multiforme | [57] | |
| circASAP1 | Positive | Glioblastoma multiforme | [58] | ||
| circSEPT9 | Positive | Triple-negative breast cancer | [59] | ||
| ADAR1 | CircARSP91 | Negative | Hepatocellular carcinoma | [61] | |
| hsa_circ_0004872 | Negative | gastric cancer | [62] | ||
| circNEIL3 | Negative | pancreatic ductal adenocarcinoma | [63] | ||
| RNA-binding proteins | HNRNPL | circLMO7 | Positive | Gastric cancer | [66] |
| QKI | circZKSCAN | Positive | Hepatocellular carcinoma | [70] | |
| FUS | CircLONP2 | Positive | Primary colorectal cancer | [72] | |
| CircZNF609 | Positive | Lung cancer | [73] | ||
| circHIF1A | Positive | Triple-negative breast cancer | [74] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.; Zhu, Q. Mechanisms Regulating Abnormal Circular RNA Biogenesis in Cancer. Cancers 2021, 13, 4185. https://doi.org/10.3390/cancers13164185
Huang Y, Zhu Q. Mechanisms Regulating Abnormal Circular RNA Biogenesis in Cancer. Cancers. 2021; 13(16):4185. https://doi.org/10.3390/cancers13164185
Chicago/Turabian StyleHuang, Ying, and Qubo Zhu. 2021. "Mechanisms Regulating Abnormal Circular RNA Biogenesis in Cancer" Cancers 13, no. 16: 4185. https://doi.org/10.3390/cancers13164185
APA StyleHuang, Y., & Zhu, Q. (2021). Mechanisms Regulating Abnormal Circular RNA Biogenesis in Cancer. Cancers, 13(16), 4185. https://doi.org/10.3390/cancers13164185