
The Expression Profiles and Deregulation of UDP-Glycosyltransferase (UGT) Genes in Human Cancers and Their Association with Clinical Outcomes

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Assessment of the Expression Profiles of UGT Genes in Human Cancers
2.2. Assessment of the Deregulation of UGT Genes in Human Cancers
2.3. Assessment of Associations between the Intratumoral Expression Levels of UGT Genes and Overall Survival of Cancer Patients Using Kaplan–Meier Survival Analysis
3. Results
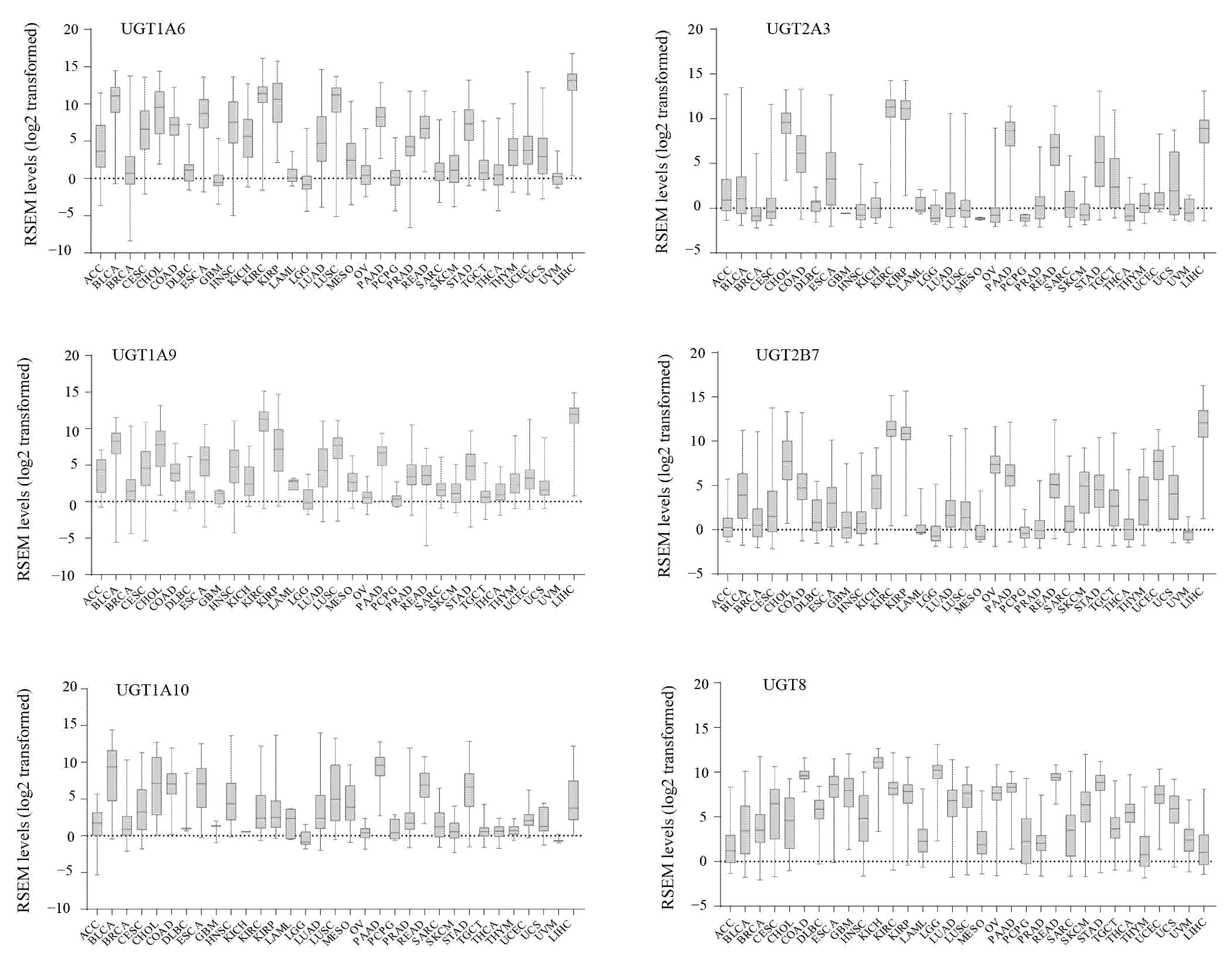
3.1. The Expression Profiles of UGT Genes in Human Cancers
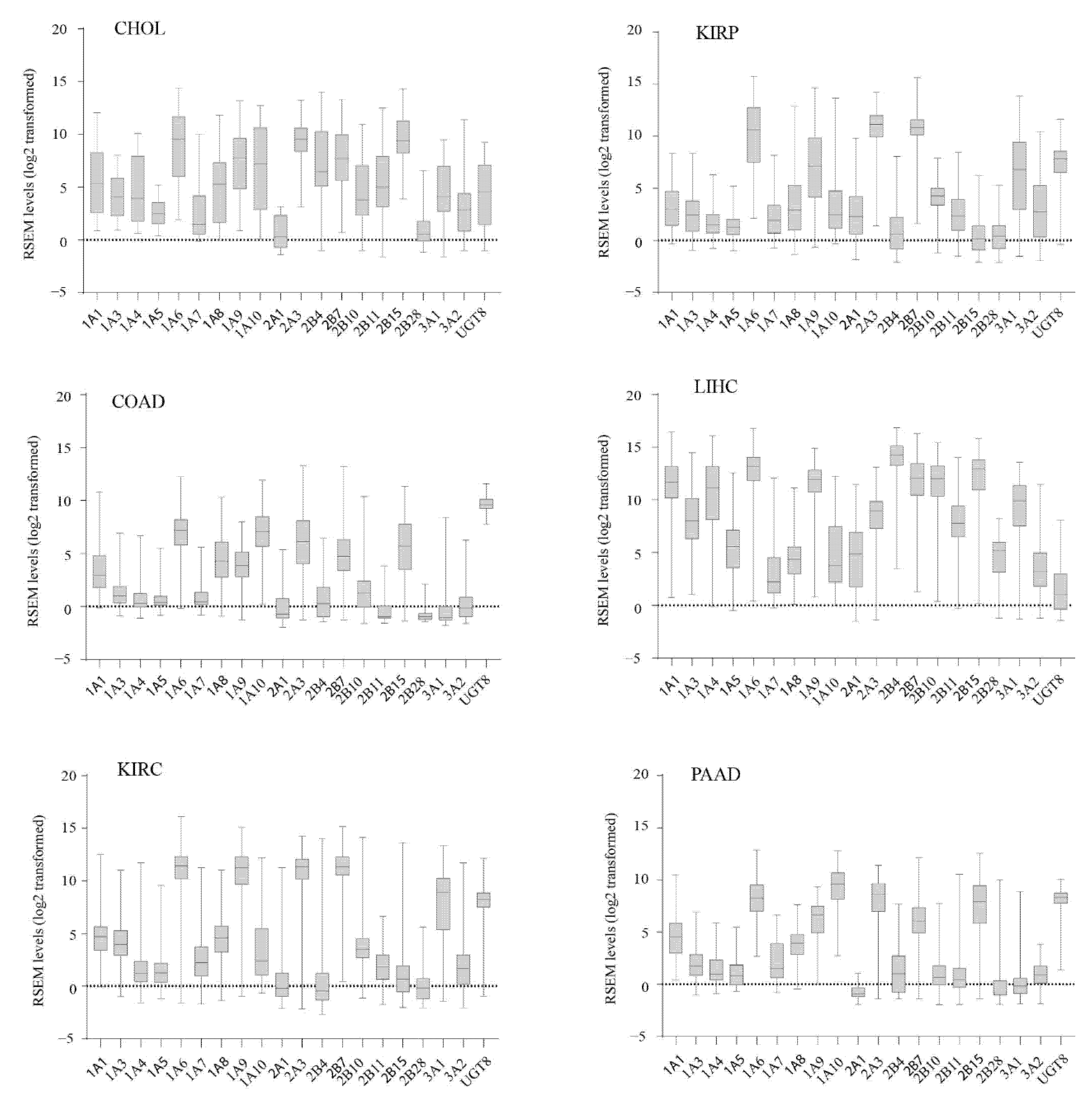
3.2. Inter-Individual Variation in UGT Expression within Human Cancers
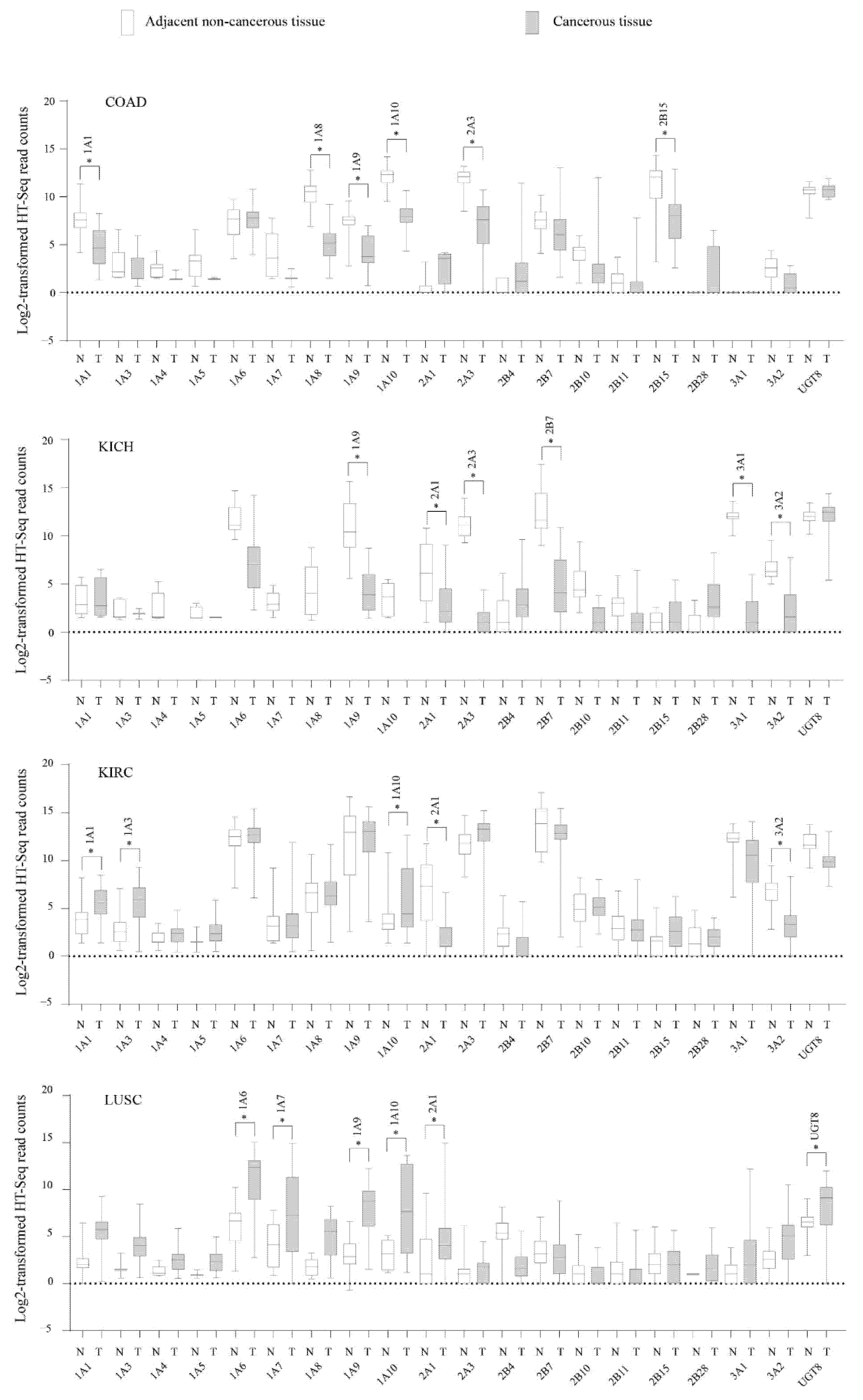
3.3. Deregulation of UGT Genes in Human Cancers
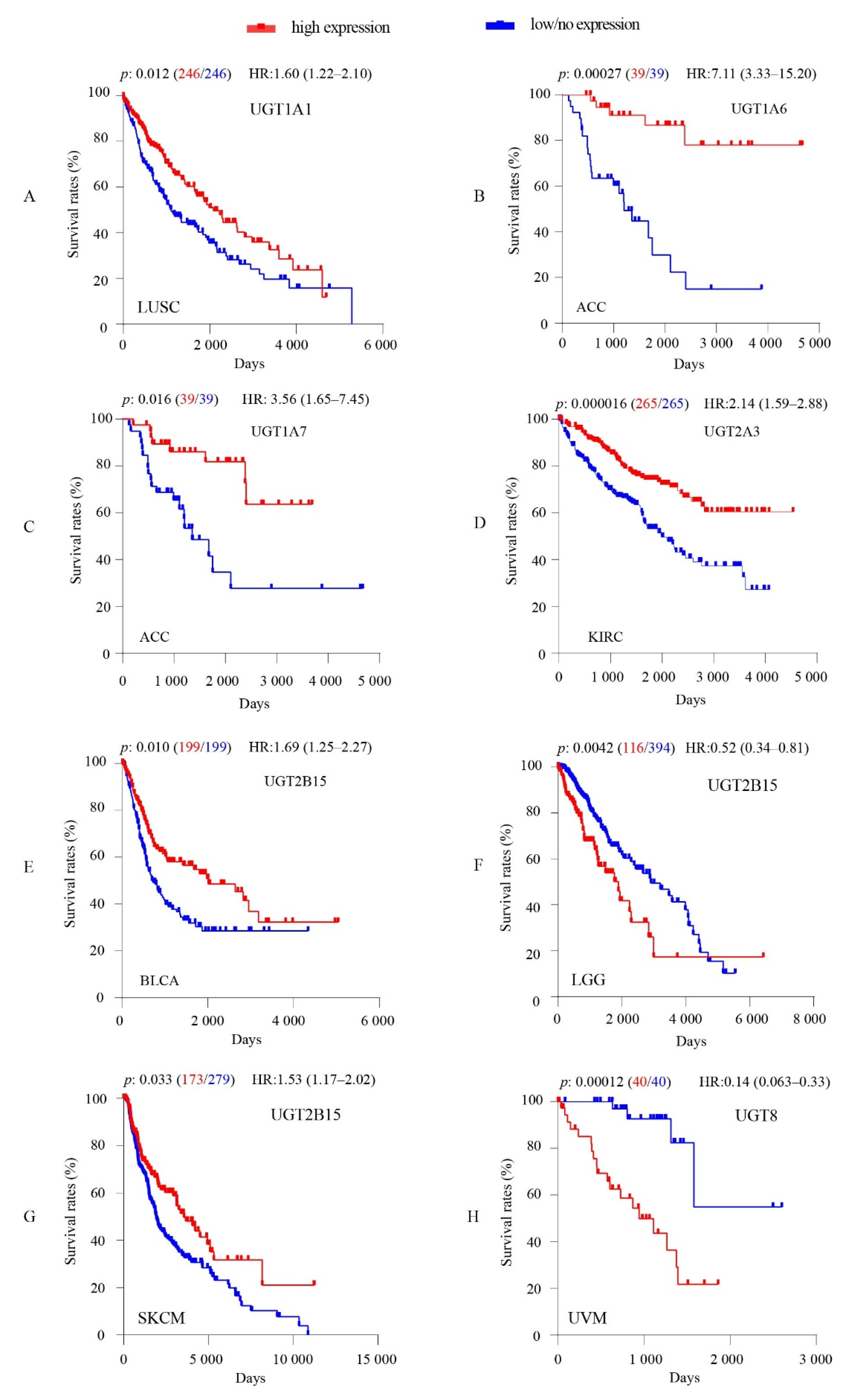
3.4. Associations between Intratumoral UGT Expression Levels and Overall Survival of Cancer Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mackenzie, P.I.; Bock, K.W.; Burchell, B.; Guillemette, C.; Ikushiro, S.-I.; Iyanagi, T.; Miners, J.O.; Owens, I.S.; Nebert, D.W. Nomenclature update for the mammalian UDP glycosyltransferase (UGT) gene superfamily. Pharmacogenet. Genom. 2005, 15, 677–685. [Google Scholar] [CrossRef]
- Guillemette, C.; Lévesque, É.; Harvey, M.; Bellemare, J.; Ménard, V. UGT genomic diversity: Beyond gene duplication. Drug Metab. Rev. 2009, 42, 24–44. [Google Scholar] [CrossRef] [PubMed]
- Meech, R.; Hu, D.G.; McKinnon, R.; Mubarokah, S.N.; Haines, A.Z.; Nair, P.; Rowland, A.; Mackenzie, P. The UDP-Glycosyltransferase (UGT) Superfamily: New Members, New Functions, and Novel Paradigms. Physiol. Rev. 2019, 99, 1153–1222. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, P.I.; Owens, I.S.; Burchell, B.; Bock, K.W.; Bairoch, A.; Belanger, A.; Fournel-Gigleux, S.; Green, M.; Hum, D.W.; Iyanagi, T.; et al. The UDP glycosyltransferase gene superfamily: Recommended nomenclature update based on evolutionary divergence. Pharmacogenetics 1997, 7, 255–269. [Google Scholar] [CrossRef] [PubMed]
- Tukey, R.H.; Strassburg, C.P. Human UDP-Glucuronosyltransferases: Metabolism, Expression, and Disease. Annu. Rev. Pharmacol. Toxicol. 2000, 40, 581–616. [Google Scholar] [CrossRef] [PubMed]
- Belledant, A.; Hovington, H.; Garcia, L.; Caron, P.; Brisson, H.; Villeneuve, L.; Simonyan, D.; Têtu, B.; Fradet, Y.; Lacombe, L.; et al. The UGT2B28 Sex-steroid Inactivation Pathway Is a Regulator of Steroidogenesis and Modifies the Risk of Prostate Cancer Progression. Eur. Urol. 2016, 69, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Émond, J.-P.; Labriet, A.; Desjardins, S.; Rouleau, M.; Villeneuve, L.; Hovington, H.; Brisson, H.; Lacombe, L.; Simonyan, D.; Caron, P.; et al. Factors Affecting Interindividual Variability of Hepatic UGT2B17 Protein Expression Examined Using a Novel Specific Monoclonal Antibody. Drug Metab. Dispos. 2019, 47, 444–452. [Google Scholar] [CrossRef]
- Milne, A.M.; Burchell, B.; Coughtrie, M.W.H. A Novel Method for the Immunoquantification of UDP-Glucuronosyltransferases in Human Tissue. Drug Metab. Dispos. 2011, 39, 2258–2263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerdpin, O.; Mackenzie, P.I.; Bowalgaha, K.; Finel, M.; Miners, J.O. Influence of N-Terminal Domain Histidine and Proline Residues on the Substrate Selectivities of Human UDP-Glucuronosyltransferase 1A1, 1A6, 1A9, 2B7, and 2B10. Drug Metab. Dispos. 2009, 37, 1948–1955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbier, O.; Lapointe, H.; El Alfy, M.; Hum, D.W.; Bélanger, A. Cellular Localization of Uridine Diphosphoglucuronosyltransferase 2B Enzymes in the Human Prostate by in Situ Hybridization and Immunohistochemistry 1. J. Clin. Endocrinol. Metab. 2000, 85, 4819–4826. [Google Scholar] [CrossRef] [PubMed]
- Lévesque, E.; Labriet, A.; Hovington, H.; Allain, E.; Melo-Garcia, L.; Rouleau, M.; Brisson, H.; Turcotte, V.; Caron, P.; Villeneuve, L.; et al. Alternative promoters control UGT2B17-dependent androgen catabolism in prostate cancer and its influence on progression. Br. J. Cancer 2020, 122, 1068–1076. [Google Scholar] [CrossRef] [PubMed]
- Paquet, S.; Fazli, L.; Grosse, L.; Verreault, M.; Têtu, B.; Rennie, P.S.; Bélanger, A.; Barbier, O. Differential Expression of the Androgen-Conjugating UGT2B15 and UGT2B17 Enzymes in Prostate Tumor Cells during Cancer Progression. J. Clin. Endocrinol. Metab. 2012, 97, E428–E432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, D.J.; Chen, Z.; Howard, L.E.; Wiggins, E.; De Hoedt, A.; Vidal, A.C.; Carney, S.T.; Squires, J.; Magyar, C.E.; Huang, J.; et al. UDP-glucuronosyltransferases and biochemical recurrence in prostate cancer progression. BMC Cancer 2017, 17, 463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achour, B.; Russell, M.R.; Barber, J.; Rostami-Hodjegan, A. Simultaneous Quantification of the Abundance of Several Cytochrome P450 and Uridine 5′-Diphospho-Glucuronosyltransferase Enzymes in Human Liver Microsomes Using Multiplexed Targeted Proteomics. Drug Metab. Dispos. 2014, 42, 500–510. [Google Scholar] [CrossRef]
- Fallon, J.K.; Neubert, H.; Goosen, T.; Smith, P.C. Targeted Precise Quantification of 12 Human Recombinant Uridine-Diphosphate Glucuronosyl Transferase 1A and 2B Isoforms Using Nano-Ultra-High-Performance Liquid Chromatography/Tandem Mass Spectrometry with Selected Reaction Monitoring. Drug Metab. Dispos. 2013, 41, 2076–2080. [Google Scholar] [CrossRef] [Green Version]
- Fallon, J.K.; Neubert, H.; Hyland, R.; Goosen, T.C.; Smith, P.C. Targeted quantitative proteomics for the analysis of 14 UGT1As and -2Bs in human liver using NanoUPLC-MS/MS with selected reaction monitoring. J. Proteome Res. 2013, 12, 4402–4413. [Google Scholar] [CrossRef]
- Sato, Y.; Nagata, M.; Kawamura, A.; Miyashita, A.; Usui, T. Protein quantification of UDP-glucuronosyltransferases 1A1 and 2B7 in human liver microsomes by LC-MS/MS and correlation with glucuronidation activities. Xenobiotica 2012, 42, 823–829. [Google Scholar] [CrossRef]
- Sato, Y.; Nagata, M.; Tetsuka, K.; Tamura, K.; Miyashita, A.; Kawamura, A.; Usui, T. Optimized Methods for Targeted Peptide-Based Quantification of Human Uridine 5′-Diphosphate-Glucuronosyltransferases in Biological Specimens Using Liquid Chromatography–Tandem Mass Spectrometry. Drug Metab. Dispos. 2014, 42, 885–889. [Google Scholar] [CrossRef] [Green Version]
- Sridar, C.; Hanna, I.; Hollenberg, P.F. Quantitation of UGT1A1 in human liver microsomes using stable isotope-labelled peptides and mass spectrometry based proteomic approaches. Xenobiotica 2012, 43, 336–345. [Google Scholar] [CrossRef]
- Margaillan, G.; Rouleau, M.; Fallon, J.K.; Caron, P.; Villeneuve, L.; Turcotte, V.; Smith, P.C.; Joy, M.S.; Guillemette, C. Quantitative Profiling of Human Renal UDP-glucuronosyltransferases and Glucuronidation Activity: A Comparison of Normal and Tumoral Kidney Tissues. Drug Metab. Dispos. 2015, 43, 611–619. [Google Scholar] [CrossRef] [Green Version]
- Court, M.H.; Duan, S.X.; Von Moltke, L.L.; Greenblatt, D.J.; Patten, C.J.; Miners, J.O.; Mackenzie, P. Interindividual variability in acetaminophen glucuronidation by human liver microsomes: Identification of relevant acetaminophen UDP-glucuronosyltransferase isoforms. J. Pharmacol. Exp. Ther. 2001, 299, 998–1006. [Google Scholar] [PubMed]
- Court, M.H.; Zhang, X.; Ding, X.; Yee, K.K.; Hesse, L.M.; Finel, M. Quantitative distribution of mRNAs encoding the 19 human UDP-glucuronosyltransferase enzymes in 26 adult and 3 fetal tissues. Xenobiotica 2011, 42, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Izukawa, T.; Nakajima, M.; Fujiwara, R.; Yamanaka, H.; Fukami, T.; Takamiya, M.; Aoki, Y.; Ikushiro, S.-I.; Sakaki, T.; Yokoi, T. Quantitative Analysis of UDP-Glucuronosyltransferase (UGT) 1A and UGT2B Expression Levels in Human Livers. Drug Metab. Dispos. 2009, 37, 1759–1768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, A.; Nakajima, M.; Yamanaka, H.; Fujiwara, R.; Yokoi, T. Expression of UGT1A and UGT2B mRNA in Human Normal Tissues and Various Cell Lines. Drug Metab. Dispos. 2008, 36, 1461–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, M.; Naito, S. Tissue-Specific mRNA Expression Profiles of Human Phase I Metabolizing Enzymes Except for Cytochrome P450 and Phase II Metabolizing Enzymes. Drug Metab. Pharmacokinet. 2006, 21, 357–374. [Google Scholar] [CrossRef]
- Ohno, S.; Nakajin, S. Determination of mRNA Expression of Human UDP-Glucuronosyltransferases and Application for Localization in Various Human Tissues by Real-Time Reverse Transcriptase-Polymerase Chain Reaction. Drug Metab. Dispos. 2009, 37, 32–40. [Google Scholar] [CrossRef] [Green Version]
- Schaefer, O.; Ohtsuki, S.; Kawakami, H.; Inoue, T.; Liehner, S.; Saito, A.; Sakamoto, A.; Ishiguro, N.; Matsumaru, T.; Terasaki, T.; et al. Absolute Quantification and Differential Expression of Drug Transporters, Cytochrome P450 Enzymes, and UDP-Glucuronosyltransferases in Cultured Primary Human Hepatocytes. Drug Metab. Dispos. 2011, 40, 93–103. [Google Scholar] [CrossRef] [Green Version]
- Rouleau, M.; Tourancheau, A.; Girard-Bock, C.; Villeneuve, L.; Vaucher, J.; Duperré, A.-M.; Audet-Delage, Y.; Gilbert, I.; Popa, I.; Droit, A.; et al. Divergent Expression and Metabolic Functions of Human Glucuronosyltransferases through Alternative Splicing. Cell Rep. 2016, 17, 114–124. [Google Scholar] [CrossRef] [Green Version]
- Tourancheau, A.; Margaillan, G.; Rouleau, M.; Gilbert, I.; Villeneuve, L.; Lévesque, E.; Droit, A.; Guillemette, C. Unravelling the transcriptomic landscape of the major phase II UDP-glucuronosyltransferase drug metabolizing pathway using targeted RNA sequencing. Pharm. J. 2015, 16, 60–70. [Google Scholar] [CrossRef] [Green Version]
- Tourancheau, A.; Rouleau, M.; Guauque-Olarte, S.; Villeneuve, L.; Gilbert, I.; Droit, A.; Guillemette, C. Quantitative profiling of the UGT transcriptome in human drug-metabolizing tissues. Pharm. J. 2017, 18, 251–261. [Google Scholar] [CrossRef] [Green Version]
- Hu, D.G.; Marri, S.; McKinnon, R.A.; MacKenzie, P.I.; Meech, R. Deregulation of the Genes that Are Involved in Drug Absorption, Distribution, Metabolism, and Excretion in Hepatocellular Carcinoma. J. Pharmacol. Exp. Ther. 2019, 368, 363–381. [Google Scholar] [CrossRef] [Green Version]
- Strassburg, C.P.; Manns, M.P.; Tukey, R.H. Differential down-regulation of the UDP-glucuronosyltransferase 1A locus is an early event in human liver and biliary cancer. Cancer Res. 1997, 57, 2979–2985. [Google Scholar]
- Beyerle, J.; Holowatyj, A.N.; Haffa, M.; Frei, E.; Gigic, B.; Schrotz-King, P.; Boehm, J.; Habermann, N.; Stiborova, M.; Scherer, D.; et al. Expression Patterns of Xenobiotic-Metabolizing Enzymes in Tumor and Adjacent Normal Mucosa Tissues among Patients with Colorectal Cancer: The ColoCare Study. Cancer Epidemiol. Biomarkers Prev. 2020, 29, 460–469. [Google Scholar] [CrossRef]
- Yang, W.; Ma, J.; Zhou, W.; Li, Z.; Zhou, X.; Cao, B.; Zhang, Y.; Liu, J.; Yang, Z.; Zhang, H.; et al. Identification of hub genes and outcome in colon cancer based on bioinformatics analysis. Cancer Manag. Res. 2019, 11, 323–338. [Google Scholar] [CrossRef] [Green Version]
- Cengiz, B.; Yumrutas, O.; Bozgeyik, E.; Borazan, E.; Igci, Y.Z.; Bozgeyik, I.; Oztuzcu, S. Differential expression of the UGT1A family of genes in stomach cancer tissues. Tumor Biol. 2015, 36, 5831–5837. [Google Scholar] [CrossRef]
- Strassburg, C.P.; Nguyen, N.; Manns, M.P.; Tukey, R.H. Polymorphic expression of the UDP-glucuronosyltransferase UGT1A gene locus in human gastric epithelium. Mol. Pharmacol. 1998, 54, 647–654. [Google Scholar]
- Hu, D.G.; Mackenzie, P.; McKinnon, R.; Meech, R. Genetic polymorphisms of human UDP-glucuronosyltransferase (UGT) genes and cancer risk. Drug Metab. Rev. 2016, 48, 47–69. [Google Scholar] [CrossRef]
- Allain, E.; Rouleau, M.; Vanura, K.; Tremblay, S.; Vaillancourt, J.; Bat, V.; Caron, P.; Villeneuve, L.; Labriet, A.; Turcotte, V.; et al. UGT2B17 modifies drug response in chronic lymphocytic leukaemia. Br. J. Cancer 2020, 123, 240–251. [Google Scholar] [CrossRef]
- Gruber, M.; Bellemare, J.; Hoermann, G.; Gleiss, A.; Porpaczy, E.; Bilban, M.; Le, T.; Zehetmayer, S.; Mannhalter, C.; Gaiger, A.; et al. Overexpression of uridine diphospho glucuronosyltransferase 2B17 in high-risk chronic lymphocytic leukemia. Blood 2013, 121, 1175–1183. [Google Scholar] [CrossRef] [Green Version]
- Weinstein, J.N.; Collisson, E.A.; Mills, G.B.; Shaw, K.R.M.; Ozenberger, B.A.; Ellrott, K.; Shmulevich, I.; Sander, C.; Stuart, J.M.; The Cancer Genome Atlas Research Network. The Cancer Genome Atlas Pan-Cancer analysis project. Nat. Genet. 2013, 45, 1113–1120. [Google Scholar] [CrossRef]
- Hu, D.G.; MacKenzie, P.I.; Nair, P.C.; McKinnon, R.A.; Meech, R. The Expression Profiles of ADME Genes in Human Cancers and Their Associations with Clinical Outcomes. Cancers 2020, 12, 3369. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Lichtenberg, T.; Hoadley, K.A.; Poisson, L.M.; Lazar, A.J.; Cherniack, A.D.; Kovatich, A.J.; Benz, C.C.; Levine, D.A.; Lee, A.V.; et al. An Integrated TCGA Pan-Cancer Clinical Data Resource to Drive High-Quality Survival Outcome Analytics. Cell 2018, 173, 400–416. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323-323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, D.G.; Hulin, J.-A.; Nair, P.C.; Haines, A.Z.; McKinnon, R.; Mackenzie, P.I.; Meech, R. The UGTome: The expanding diversity of UDP glycosyltransferases and its impact on small molecule metabolism. Pharmacol. Ther. 2019, 204, 107414. [Google Scholar] [CrossRef] [PubMed]
- Risso, D.; Schwartz, K.; Sherlock, G.; Dudoit, S. GC-Content Normalization for RNA-Seq Data. BMC Bioinform. 2011, 12, 480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhodes, D.R.; Kalyana-Sundaram, S.; Mahavisno, V.; Varambally, R.; Yu, J.; Briggs, B.B.; Barrette, T.R.; Anstet, M.J.; Kincead-Beal, C.; Kulkarni, P.; et al. Oncomine 3.0: Genes, Pathways, and Networks in a Collection of 18,000 Cancer Gene Expression Profiles. Neoplasia 2007, 9, 166–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welsh, J.B.; Sapinoso, L.M.; Su, A.I.; Kern, S.G.; Wang-Rodriguez, J.; Moskaluk, C.A.; Frierson, H.F., Jr.; Hampton, G.M. Analysis of gene expression identifies candidate markers and pharmacological targets in prostate cancer. Cancer Res. 2001, 61, 5974–5978. [Google Scholar]
- Okayama, H.; Kohno, T.; Ishii, Y.; Shimada, Y.; Shiraishi, K.; Iwakawa, R.; Furuta, K.; Tsuta, K.; Shibata, T.; Yamamoto, S.; et al. Identification of Genes Upregulated in ALK-Positive and EGFR/KRAS/ALK-Negative Lung Adenocarcinomas. Cancer Res. 2012, 72, 100–111. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Aerts, J.; den Hamer, B.; van Ijcken, W.; den Bakker, M.; Riegman, P.; Van Der Leest, C.; Van Der Spek, P.; Foekens, J.A.; Hoogsteden, H.C.; et al. Gene Expression-Based Classification of Non-Small Cell Lung Carcinomas and Survival Prediction. PLoS ONE 2010, 5, e10312. [Google Scholar] [CrossRef]
- Kaiser, S.; Park, Y.-K.; Franklin, J.L.; Halberg, R.B.; Yu, M.; Jessen, W.J.; Freudenberg, J.; Chen, X.; Haigis, K.; Jegga, A.G.; et al. Transcriptional recapitulation and subversion of embryonic colon development by mouse colon tumor models and human colon cancer. Genome Biol. 2007, 8, R131. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Leung, S.Y.; Yuen, S.T.; Chu, K.-M.; Ji, J.; Li, R.; Chan, A.S.Y.; Law, S.; Troyanskaya, O.G.; Wong, J.; et al. Variation in Gene Expression Patterns in Human Gastric Cancers. Mol. Biol. Cell 2003, 14, 3208–3215. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.; Otu, H.; Spentzos, D.; Kolia, S.; Inan, M.; Beecken, W.D.; Fellbaum, C.; Gu, X.; Joseph, M.; Pantuck, A.J.; et al. Gene Signatures of Progression and Metastasis in Renal Cell Cancer. Clin. Cancer Res. 2005, 11, 5730–5739. [Google Scholar] [CrossRef] [Green Version]
- Dinse, G.E.; Lagakos, S.W. Nonparametric Estimation of Lifetime and Disease Onset Distributions from Incomplete Observations. Biometrics 1982, 38, 921. [Google Scholar] [CrossRef]
- Uhlén, M.; Zhang, C.; Lee, S.; Sjöstedt, E.; Fagerberg, L.; Bidkhori, G.; Benfeitas, R.; Arif, M.; Liu, Z.; Edfors, F.; et al. A pathology atlas of the human cancer transcriptome. Science 2017, 357, eaan2507. [Google Scholar] [CrossRef] [Green Version]
- Lonsdale, J.; Thomas, J.; Slavatore, M.; Phillips, R.; Lo, E.; Shad, S.; Hasz, R.; Walters, G.; Garcia, F.; Young, N.; et al. The Genotype-Tissue Expression (GTEx) project. Nat. Genet. 2013, 45, 580–585. [Google Scholar] [CrossRef]
- Allain, E.; Rouleau, M.; Lévesque, E.; Guillemette, C. Emerging roles for UDP-glucuronosyltransferases in drug resistance and cancer progression. Br. J. Cancer 2020, 122, 1277–1287. [Google Scholar] [CrossRef] [Green Version]
- De Almagro, M.C.; Selga, E.; Thibaut, R.; Porte, C.; Noé, V.; Ciudad, C. UDP-glucuronosyltransferase 1A6 overexpression in breast cancer cells resistant to methotrexate. Biochem. Pharmacol. 2011, 81, 60–70. [Google Scholar] [CrossRef] [Green Version]
- Hu, D.G.; MacKenzie, P.I.; Lu, L.; Meech, R.; McKinnon, R.A. Induction of Human UDP-Glucuronosyltransferase 2B7 Gene Expression by Cytotoxic Anticancer Drugs in Liver Cancer HepG2 Cells. Drug Metab. Dispos. 2015, 43, 660–668. [Google Scholar] [CrossRef] [Green Version]
- Hu, D.G.; Rogers, A.; Mackenzie, P. Epirubicin Upregulates UDP Glucuronosyltransferase 2B7 Expression in Liver Cancer Cells via the p53 Pathway. Mol. Pharmacol. 2014, 85, 887–897. [Google Scholar] [CrossRef] [Green Version]
- Acquaviva, J.; He, S.; Zhang, C.; Jimenez, J.-P.; Nagai, M.; Sang, J.; Sequeira, M.; Smith, N.L.; Ogawa, L.S.; Inoue, T.; et al. FGFR3 Translocations in Bladder Cancer: Differential Sensitivity to HSP90 Inhibition Based on Drug Metabolism. Mol. Cancer Res. 2014, 12, 1042–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landmann, H.; Proia, D.A.; He, S.; Ogawa, L.S.; Kramer, F.; Beißbarth, T.; Grade, M.; Gaedcke, J.; Ghadimi, M.; Moll, U.; et al. UDP glucuronosyltransferase 1A expression levels determine the response of colorectal cancer cells to the heat shock protein 90 inhibitor ganetespib. Cell Death Dis. 2014, 5, e1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oguri, T.; Takahashi, T.; Miyazaki, M.; Isobe, T.; Kohno, N.; Mackenzie, P.; Fujiwara, Y. UGT1A10 is responsible for SN-38 glucuronidation and its expression in human lung cancers. Anticancer. Res. 2004, 24, 2893–2896. [Google Scholar] [PubMed]
- Li, P.; Chen, J.; Miyamoto, H. Androgen Receptor Signaling in Bladder Cancer. Cancers 2017, 9, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombard, A.P.; Mudryj, M. The emerging role of the androgen receptor in bladder cancer. Endocrine-Related Cancer 2015, 22, R265–R277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizushima, T.; Tirador, K.A.; Miyamoto, H. Androgen receptor activation: A prospective therapeutic target for bladder cancer? Expert Opin. Ther. Targets 2017, 21, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Bélanger, A.; Pelletier, G.; Labrie, F.; Barbier, O.; Chouinard, S. Inactivation of androgens by UDP-glucuronosyltransferase enzymes in humans. Trends Endocrinol. Metab. 2003, 14, 473–479. [Google Scholar] [CrossRef]
- Crona, J.; Beuschlein, F. Adrenocortical carcinoma—towards genomics guided clinical care. Nat. Rev. Endocrinol. 2019, 15, 548–560. [Google Scholar] [CrossRef]
- Waszut, U.; Szyszka, P.; Dworakowska, D. Understanding mitotane mode of action. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2017, 68, 13–26. [Google Scholar]
- Cai, W.; Benitez, R.; Counsell, R.E.; Djanegara, T.; Schteingart, D.E.; Sinsheimer, J.E.; Wotring, L.L. Bovine adrenal cortex transformations of mitotane [1-(2-chlorophenyl)-1-(4-chlorophenyl)-2,2-dichloroethane; o,p′-DDD] and its p,p′- and m,p′-isomers. Biochem. Pharmacol. 1995, 49, 1483–1489. [Google Scholar] [CrossRef]
- Robertson, A.G.; Shih, J.; Yau, C.; Gibb, E.A.; Oba, J.; Mungall, K.L.; Hess, J.M.; Uzunangelov, V.; Walter, V.; Danilova, L.; et al. Integrative Analysis Identifies Four Molecular and Clinical Subsets in Uveal Melanoma. Cancer Cell 2017, 32, 204–220. [Google Scholar] [CrossRef] [Green Version]
- Kaliki, S.; Shields, C.L. Uveal melanoma: Relatively rare but deadly cancer. Eye 2017, 31, 241–257. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.; Ma, C. Identification of prognostic genes in uveal melanoma microenvironment. PLoS ONE 2020, 15, e0242263. [Google Scholar] [CrossRef]
- Meech, R.; Mubarokah, N.; Shivasami, A.; Rogers, A.; Nair, P.C.; Hu, D.G.; McKinnon, R.A.; MacKenzie, P.I. A Novel Function for UDP Glycosyltransferase 8: Galactosidation of Bile Acids. Mol. Pharmacol. 2015, 87, 442–450. [Google Scholar] [CrossRef] [Green Version]
- Bosioa, A.; Binczeka, E.; Le Beau, M.M.; Fernald, A.A.; Stoffel, W. The Human Gene CGT Encoding the UDP-Galactose Ceramide Galactosyl Transferase (Cerebroside Synthase): Cloning, Characterization, and Assignment to Human Chromosome 4, Band q26. Genomics 1996, 34, 69–75. [Google Scholar] [CrossRef]
- Honke, K.; Tsuda, M.; Koyota, S.; Wada, Y.; Iida-Tanaka, N.; Ishizuka, I.; Nakayama, J.; Taniguchi, N. Molecular cloning and characterization of a human beta-Gal-3’-sulfotransferase that acts on both type 1 and type 2 (Gal beta 1-3/1-4GlcNAc-R) oligosaccharides. J. Biol. Chem. 2001, 276, 267–274. [Google Scholar] [CrossRef] [Green Version]
- Lai, M.; La Rocca, V.; Amato, R.; Freer, G.; Pistello, M. Sphingolipid/Ceramide Pathways and Autophagy in the Onset and Progression of Melanoma: Novel Therapeutic Targets and Opportunities. Int. J. Mol. Sci. 2019, 20, 3436. [Google Scholar] [CrossRef] [Green Version]
- Owczarek, T.B.; Suchanski, J.; Pula, B.; Kmiecik, A.M.; Chadalski, M.; Jethon, A.; Dziegiel, P.; Ugorski, M. Galactosylceramide Affects Tumorigenic and Metastatic Properties of Breast Cancer Cells as an Anti-Apoptotic Molecule. PLoS ONE 2013, 8, e84191. [Google Scholar] [CrossRef] [Green Version]
- Cao, Q.; Chen, X.; Wu, X.; Liao, R.; Huang, P.; Tan, Y.; Wang, L.; Ren, G.; Huang, J.; Dong, C. Inhibition of UGT8 suppresses basal-like breast cancer progression by attenuating sulfatide-alphaVbeta5 axis. J. Exp. Med. 2018, 215, 1679–1692. [Google Scholar] [CrossRef] [Green Version]
- Izumi, K.; Li, Y.; Ishiguro, H.; Zheng, Y.; Yao, J.L.; Netto, G.J.; Miyamoto, H. Expression of UDP-glucuronosyltransferase 1A in bladder cancer: Association with prognosis and regulation by estrogen. Mol. Carcinog. 2012, 53, 314–324. [Google Scholar] [CrossRef]
- Kaushik, A.K.; Vareed, S.K.; Basu, S.; Putluri, V.; Putluri, N.; Panzitt, K.; Brennan, C.A.; Chinnaiyan, A.M.; Vergara, I.A.; Erho, N.; et al. Metabolomic Profiling Identifies Biochemical Pathways Associated with Castration-Resistant Prostate Cancer. J. Proteome Res. 2014, 13, 1088–1100. [Google Scholar] [CrossRef] [Green Version]
- Olivier, B.; Xie, N.; Chen, R.; Verreault, M.; Fazli, L.; Gleave, M.; Barbier, O.; Dong, X. UGT2B17 Expedites Progression of Castration-Resistant Prostate Cancers by Promoting Ligand-Independent AR Signaling. Cancer Res. 2016, 76, 6701–6711. [Google Scholar] [CrossRef] [Green Version]
- Stanbrough, M.; Bubley, G.J.; Ross, K.; Golub, T.R.; Rubin, M.; Penning, T.; Febbo, P.G.; Balk, S.P. Increased Expression of Genes Converting Adrenal Androgens to Testosterone in Androgen-Independent Prostate Cancer. Cancer Res. 2006, 66, 2815–2825. [Google Scholar] [CrossRef] [Green Version]
- Hu, D.G.; Selth, L.A.; Tarulli, G.A.; Meech, R.; Wijayakumara, D.; Chanawong, A.; Russell, R.; Caldas, C.; Robinson, J.L.L.; Carroll, J.S.; et al. Androgen and estrogen receptors in breast cancer coregulate human UDP-glucuronosyltransferases 2B15 and 2B17. Cancer Res. 2016, 76, 5881–5893. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer Type | Description | No. of Patients | No. of Paired Cancerous and Non-Cancerous Tissues |
|---|---|---|---|
| ACC | Adrenocortical carcinoma | 78 | |
| BLCA | Bladder Urothelial Carcinoma | 398 | |
| BRCA | Breast Invasive Carcinoma | 1080 | 113 |
| CESC | Cervical Squamous Cell Carcinoma and | ||
| endocervical Adenocarcinoma | 304 | ||
| CHOL | Cholangiocarcinoma | 35 | |
| COAD | Colon Adenocarcinoma | 235 | 24 |
| DLBC | Lymphoid Neoplasm Diffuse Large B-cell | ||
| lymphoma | 47 | ||
| ESCA | Esophageal Carcinoma | 182 | |
| GBM | Glioblastoma Multiforme | 148 | |
| HNSC | Head and Neck Squamous Cell Carcinoma | 519 | 43 |
| KICH | Kidney Chromophobe | 65 | 25 |
| KIRC | Kidney Renal Clear Cell Carcinoma | 531 | 72 |
| KIRP | Kidney Renal Papillary Cell Carcinoma | 287 | 32 |
| LAML | Acute Myeloid Leukemia | 149 | |
| LGG | Brain Lower Grade Glioma | 510 | |
| LIHC | Liver Hepatocellular Carcinoma | 365 | 50 |
| LUAD | Lung Adenocarcinoma | 502 | 58 |
| LUSC | Lung Squamous Cell Carcinoma | 492 | 51 |
| MESO | Mesothelioma | 85 | |
| OV | Ovary Serous Cystadenocarcinoma | 294 | |
| PCPG | Pheochromocytoma and Paraganglioma | 179 | |
| PAAD | Pancreatic Adenocarcinoma | 144 | |
| PRAD | Prostate adenocarcinoma | 462 | 52 |
| READ | Rectum Adenocarcinoma | 93 | |
| SARC | Sarcoma | 258 | |
| SKCM | Skin Cutaneous Melanoma | 452 | |
| STAD | Stomach Adenocarcinoma | 357 | 32 |
| TGCT | Testicular Germ Cell Tumors | 134 | |
| THCA | Thyroid Carcinoma | 504 | 59 |
| THYM | Thymoma | 119 | |
| UCEC | Uterine Corpus Endometrial Carcinoma | 370 | |
| UCS | Uterine Carcinosarcoma | 56 | |
| UVM | Uveal Melanoma | 80 |
| TCGA Datasets | Non-TCGA Datasets | |||||
|---|---|---|---|---|---|---|
| UGT Genes | Cancer Types | Fold Change | p-Values | Fold Change | p-Values | Independent Studies |
| 1A1 | COAD | −7.0 | 2.74 × 10−6 | −10.5 | 6.66 × 10−5 | Kaiser S et al., 2007 [51] |
| 1A1 | KIRC | 4.8 | 9.69 × 10−11 | |||
| 1A1 | STAD | −10.1 | 1.90 × 10−5 | |||
| 1A3 | KIRC | 13.4 | 3.35 × 10−25 | |||
| 1A3 | PRAD | 6.8 | 1.49 × 107 | |||
| 1A6 | LUAD | 6.5 | 4.82 × 10−10 | |||
| 1A6 | LUSC | 48.5 | 1.63 × 10−45 | 11.9 | 2.04 × 10−11 | Hou J et al., 2010 [50] |
| 1A7 | LUSC | 1074.9 | 2.96 × 10−28 | |||
| 1A7 | STAD | −8.8 | 0.0051 | |||
| 1A8 | COAD | −27.0 | 7.70 × 10−34 | −12.5 | 2.01 × 10−5 | Kaiser S et al., 2007 [51] |
| 1A8 | HNSC | −6.3 | 0.00011 | |||
| 1A9 | COAD | −10.8 | 2.33 × 10−11 | −11.7 | 2.66 × 10−5 | Kaiser S et al., 2007 [51] |
| 1A9 | KICH | −229.1 | 3.36 × 10−22 | |||
| 1A9 | LUSC | 65.3 | 3.00 × 10−33 | 14.9 | 1.19 × 10−11 | Hou J et al., 2010 [50] |
| 1A10 | COAD | −16.5 | 2.18 × 10−39 | |||
| 1A10 | KIRC | 39.1 | 7.84 × 10−19 | |||
| 1A10 | LIHC | 54.5 | 4.81 × 10−12 | |||
| 2A1 | HNSC | −6.5 | 0.0033 | |||
| 2A1 | KICH | −8.3 | 2.24 × 10−5 | −2.5 | 1.38 × 10−4 | Jones J et al., 2005 [53] |
| 2A1 | KIRC | −90.5 | 4.34 × 10−36 | −4.0 | 3.63 × 10−18 | Jones J et al., 2005 [53] |
| 2A1 | KIRP | −6.5 | 0.000673 | |||
| 2A1 | LUSC | 16.4 | 5.19 × 10−11 | |||
| 2A3 | COAD | −15.8 | 9.33 × 10−22 | −7.6 | 6.38 × 10−10 | Kaiser S et al., 2007 [51] |
| 2A3 | KICH | −1112.8 | 2.90 × 10−101 | −3.1 | 1.62 × 10−6 | Jones J et al., 2005 [53] |
| 2B4 | PRAD | 29.4 | 2.62 × 10−25 | 4.3 | 6.18 × 10−5 | Welsh JB et al., 2001 [48] |
| 2B7 | KICH | −110.6 | 7.06 × 10−23 | |||
| 2B7 | KIRP | −6.0 | 3.99 × 10−9 | |||
| 2B7 | STAD | −4.1 | 0.0093 | −2.5 | 1.15 × 10−4 | Chen X et al., 2003 [52] |
| 2B7 | PRAD | −56.8 | 4.32 × 10−18 | |||
| 2B11 | LIHC | 5.3 | 1.30 × 10−12 | |||
| 2B15 | COAD | −10.5 | 7.09 × 10−10 | −12.0 | 3.42 × 10−6 | Kaiser S et al., 2007 [51] |
| 2B15 | LUAD | 6 | 4.56 × 10−10 | 3.3 | 3.82 × 10−16 | Okayama H et al., 2003 [49] |
| 3A1 | KICH | −661.6 | 3.10 × 10−99 | |||
| 3A2 | KICH | −11.7 | 2.90 × 10−8 | |||
| 3A2 | KIRC | −6.6 | 3.04 × 10−22 | |||
| UGT8 | LUAD | 5.3 | 1.63 × 10−26 | 3 | 4.33 × 10−8 | Hou J et al., 2010 [50] |
| UGT8 | LUSC | 6.8 | 2.45 × 10−22 | 5.8 | 4.11 × 10−9 | Hou J et al., 2010 [50] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, D.G.; Marri, S.; Mackenzie, P.I.; Hulin, J.-A.; McKinnon, R.A.; Meech, R. The Expression Profiles and Deregulation of UDP-Glycosyltransferase (UGT) Genes in Human Cancers and Their Association with Clinical Outcomes. Cancers 2021, 13, 4491. https://doi.org/10.3390/cancers13174491
Hu DG, Marri S, Mackenzie PI, Hulin J-A, McKinnon RA, Meech R. The Expression Profiles and Deregulation of UDP-Glycosyltransferase (UGT) Genes in Human Cancers and Their Association with Clinical Outcomes. Cancers. 2021; 13(17):4491. https://doi.org/10.3390/cancers13174491
Chicago/Turabian StyleHu, Dong Gui, Shashikanth Marri, Peter I. Mackenzie, Julie-Ann Hulin, Ross A. McKinnon, and Robyn Meech. 2021. "The Expression Profiles and Deregulation of UDP-Glycosyltransferase (UGT) Genes in Human Cancers and Their Association with Clinical Outcomes" Cancers 13, no. 17: 4491. https://doi.org/10.3390/cancers13174491
APA StyleHu, D. G., Marri, S., Mackenzie, P. I., Hulin, J. -A., McKinnon, R. A., & Meech, R. (2021). The Expression Profiles and Deregulation of UDP-Glycosyltransferase (UGT) Genes in Human Cancers and Their Association with Clinical Outcomes. Cancers, 13(17), 4491. https://doi.org/10.3390/cancers13174491