
Potential of Bioactive Food Components against Gastric Cancer: Insights into Molecular Mechanism and Therapeutic Targets


 ,
,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
3. Apoptosis-Inducing Natural Bioactive Food Components in Gastric Cancer

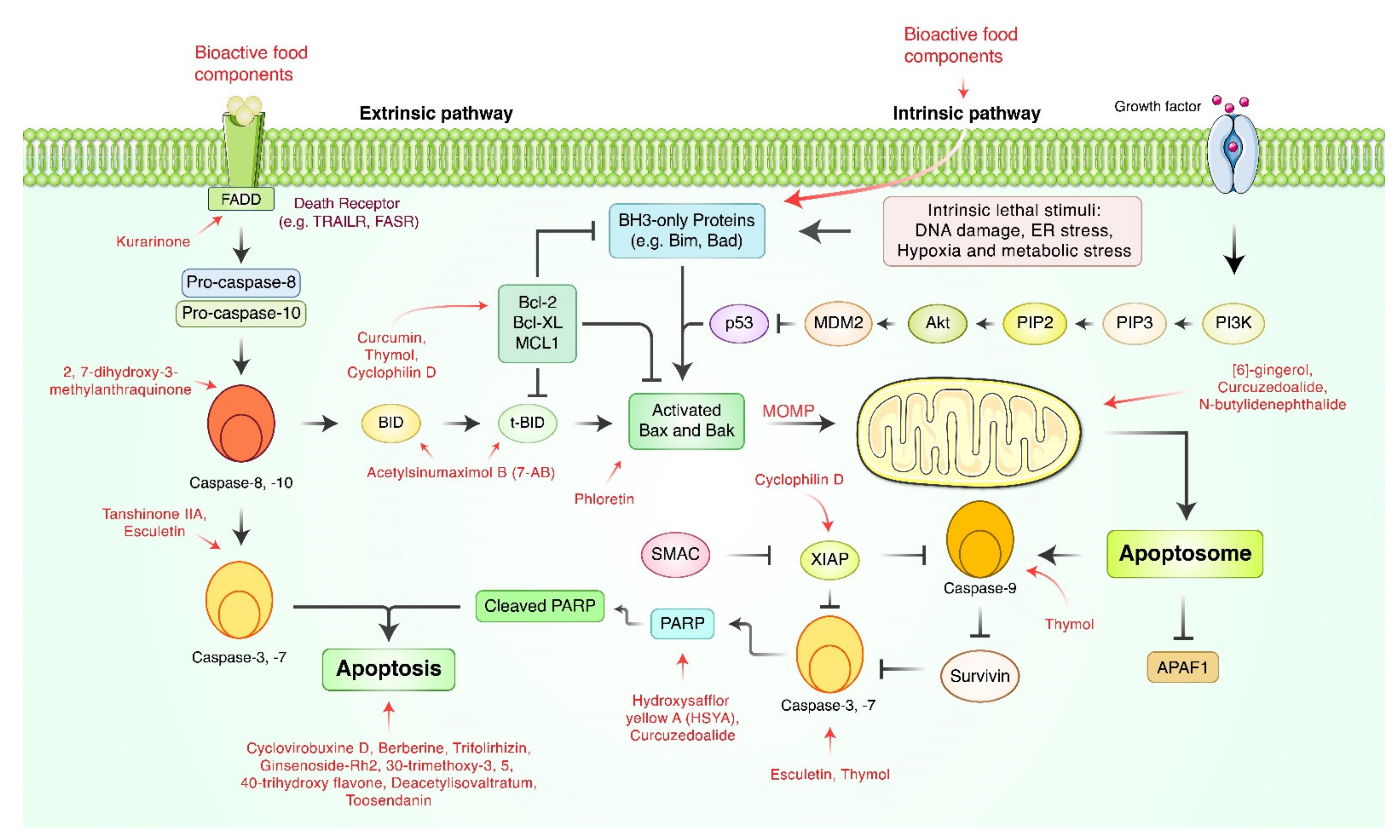{kind=link}
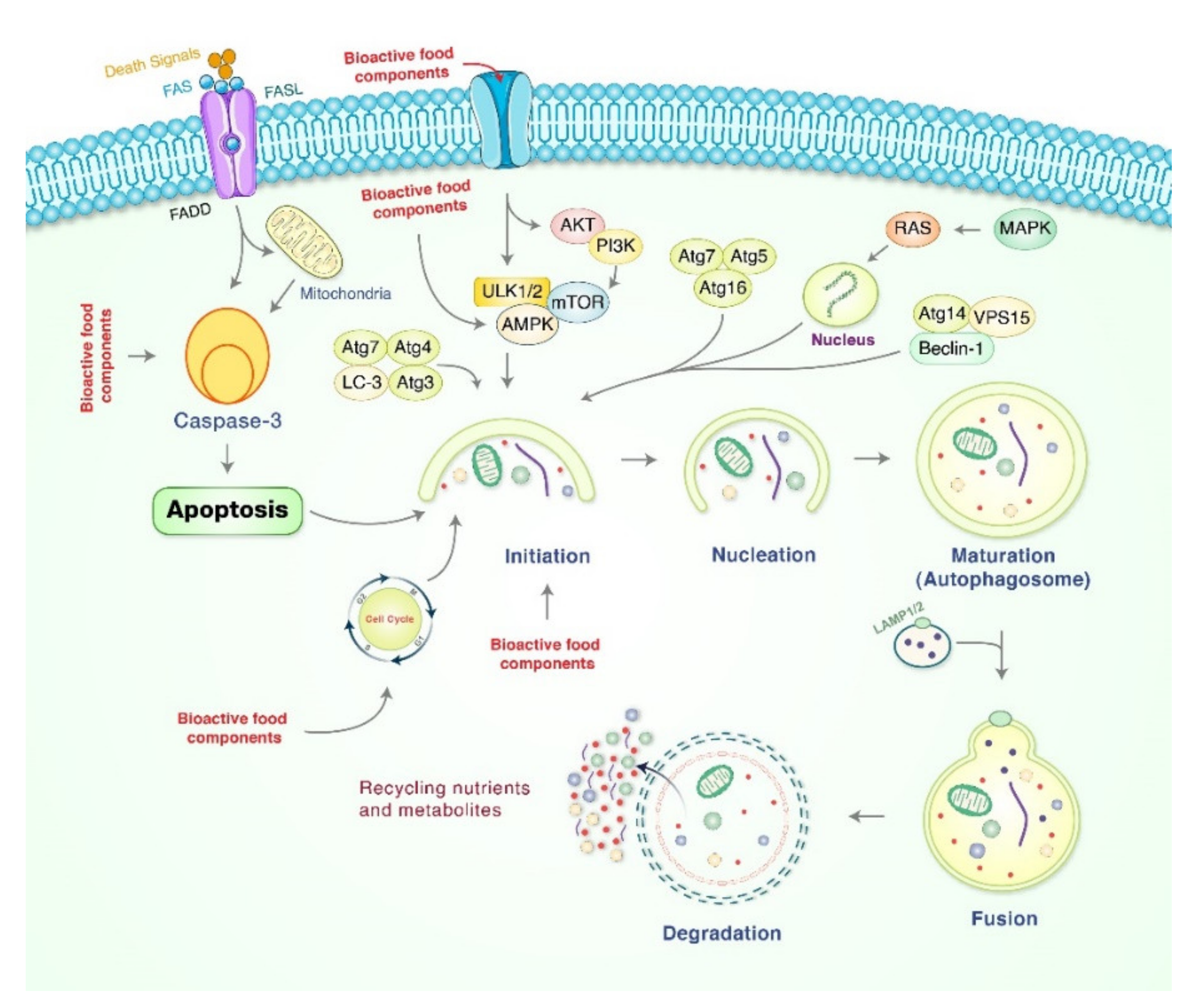{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classification | Compound | Source | Experimental Model | Dose; Duration | Efficacy | Mechanism | References |
|---|---|---|---|---|---|---|---|
| Alkaloids | Berberine | (family: Ranunculaceae) Coptidis japonica Makino Rhizoma | SGC-7901 | 5, 10, 20 µM; 24, 48 h | Induction of apoptosis | [35] | |
| Alkaloids | Cyclovirobuxine D | (family: Buxaceae) Buxus microphylla Richardii Radix | MGC-803, MKN-28 | 30, 60, 120 μM/L; 48 h | Induction of apoptosis | ↑c-caspase-3, Bax ↓Bcl-2 | [36] |
| Alkaloids | GFG-3a | (family: Meripilaceae) Grifola frondose (Diks.) Gray Mycelia | SGC-7901 | 100, 200 μg/mL; 24, 48 h | Induction of apoptosis | ↑RBBP4, caspase-3, -8, p53, Bax, Bad ↓RUVBL, NPM, Bcl-2, Bcl-xL, PI3K, Akt1 | [64] |
| Alkaloids | Melittin | (family: Apidae) Apis cerena Fabricius venom | SGC-7901 | 4 μg/mL; 1, 2, 4 h | Induction of apoptosis | ↑caspase-3, cyt c, Endo G, AIF, Smac/Diablo, ROS ↓ΔΨm | [65] |
| Alkaloids, Terpenoids | Berberine, d-Limonene | (1) (family: Ranunculaceae) Coptidis japonica Makino Rhizoma (2) (family: Rutaceae) Evodiae rutaecarpa Bentham. Fructus | MGC-803 | (1) 20 µM; 24, 36, 48 h (2) 80 µM; 24, 36, 48 h | Induction of apoptosis | ↑ROS, caspase-3 ↑ΔΨm, Bcl-2 | [66] |
| Flavonoids | Trifolirhizin | (family: Fabaceae) Sophora flavescens Aiton Radix | MKN-45 | 20, 30, 40 µg/mL; 48 h | Induction of apoptosis | ↑caspase-9, -3, c-PARP, p53, p38 ↓EGFR, CDC2, cyclin B, ΔΨm | [37] |
| Phytosterols | Ginsenoside-Rh2 | (family: Araliaceae) Panax ginseng C.A. Mey Radix | SGC-7901 | 5, 10, 20 μg/mL; 24, 48 h | Induction of apoptosis | ↑Bax ↓Bcl-2 | [38] |
| Phytosterols | Periplocin | (family: Apocynaceae) Periplocae sepium Bunge. | SGC-7901, MGC-803, BGC-823 | 50, 100, 200 ng/mL; 24, 48 h | Induction of apoptosis | ↑Mcl-1, c-caspase-3, EGR 1 ↓pro-Bid, p-ERK 1/2 | [67] |
| Phytosterols | Tanshinone IIA | (family: Lamiaceae) Salviae miltiorrhiza Bunge. Radix | AGS | 2.0, 3.7, 5.5 µg/mL; 24, 48 h | Induction of apoptosis | ↑TNF-α, Fas, p-p38, p-JNK, p53, p21, caspase-8, -3 ↓p-ERK, CDC2, cyclin A, cyclin B1 | [39] |
| Polyphenols | [6]-Gingerol | (family: Zingiberaceae) Zingiber officinale Roscoe Rhizoma | AGS | 100, 250 µM; 24 h | Induction of apoptosis | ↑cyt c, Bax ↓Bcl-2 | [68] |
| Polyphenols | 2,7-dihydroxy-3-methylanthraquinone (DDMN) | (family: Rubiaceae) Hedyotis diffusa Wild Herba | SGC-7901 | 10, 20, 40 µM; 48 h | Inhibition of proliferation | ↑Bax, Bad, caspase-3, -9, cyt c ↓Bcl-xL, Bcl-2 | [40] |
| Polyphenols | 6, 7, 30-trimethoxy-3, 5, 40 -trihydroxy flavone (TTF) | (family: Saxifragaceae) Chrysosplenium nudicaule Ledeb Herba | SGC-7901 | 2, 4, 8, 16, 32 µg/mL; 24, 48, 72 h | Induction of apoptosis | ↑endogenous Ca2+/Mg2+ dependent endonuclease | [41] |
| Polyphenols | Curcumin | (family: Zingiberaceae) Curcuma longa L. Rhizoma | SGC-7901, BGC-823 | 5, 10, 15, 20, 40 μM/L; 24 h | Induction of apoptosis | ↓XIAP ↑miR-33b | [42] |
| Polyphenols | Esculetin | (family: Asteraceae) Artemesia scoparia Waldst. et Kit, Artemesia capillaris Thunb.)) (family: Plumbaginaceae) | SGC-7901, MGC-803, BGC-823 | 12.5, 25, 50 μM; 24 h | Induction of apoptosis | ↑ROS, c-caspase-9, -3, c-PARP, cyt c, Bak, Bax, CypD ↓Bcl-2, Bcl-xL, XIAP | [43] |
| Polyphenols | Hydroxysafflor Yellow A | (family: Asteraceae) Carthamus tinctorius L. | BGC-823 | 100 µM; 48 h | Induction of apoptosis | ↑caspase-3, PPARγ | [44] |
| Polyphenols | Kurarinone | (family: Fabaceae) Sophora flavescens Aiton Radix | SGC-7901 | 5 μM; 24 h | Enhancement ofTRAIL-induced apoptosis | ↓Mcl-1, c-FLIP, p-STAT3 | [45] |
| Polyphenols | Licochalcone A | (family: Fabaceae) Glycyrrhiza glabra L. Root | MKN-45, SGC-7901 | 15, 30, 60 µM; 24 h | Inhibition of cell proliferation and tumor glycolysis | ↑c-caspase-3, c-PARP ↓Bcl-2, Mcl-1, HK2, p-Akt, p-ERK1/2, p-S6, p-GSK3β | [46] |
| Polyphenols | Ophiopogonin B | (family: Asparagaceae) Ophiopogon japonicus Thunb Root | SGC-7901 | 5, 10, 20 μM | Induction of apoptosis | ↑ROS, Bax, caspase-3 ↓p-ERK 1/2, p-JNK 1/2, ΔΨm, Bcl-2 | [49] |
| Polyphenols | Phloretin | AGS | 4, 8, 16µM; 24 h | Induction of apoptosis Inhibition of invasion | ↑Bax ↓Bcl-2 | [50] | |
| Polyphenols | Podophyllotoxin | (family: Linaceae) Linum album Kotschy | AGS | 200, 400, 600, 800, 1000 µg/mL; 24 h | Induction of apoptosis | ↓ZNF703 | [51] |
| Terpenoids | 7-Acetylsinumaximol B | (family: Alcyoniidae) Sinularia sandensis | NCI-N87 | 4, 8, 16 µM; 24 h | Induction of apoptosis | ↑Bad, Bim, Bax, cyt c ↓p-Bad, Mcl-1, Bcl-xL, Bcl-2 | [53] |
| Terpenoids | Crosolic Acid | (family: Actinidiaceae) Actinidia valvata Dunn Radix | BGC-823 | 20, 40, 80 μg/mL; 72 h | Induction of apoptosis | ↑Bax, smac, IκBα ↓Fas, Bcl-2, p65, p-IκBα, NF-κB | [54] |
| Terpenoids | Curcuzedoalide | (family: Zingiberaceae) Curcuma zedoaria Roscoe Rhizoma | AGS | 100, 200 µM; 24 h | Induction of apoptosis | ↑c-caspase-8, -9, -3, c-PARP | [47] |
| Terpenoids | Deacetylisovaltratum | (family: Caprifoliaceae) Patrinia heterophylla Bunge. | (1) AGS (2) HGC-27 | (1) 4, 8, 16 μM; 24 h (2) 10, 20, 30 μM; 24 h | Induction of apoptosis | ↑p21, caspase-3, c-PARP ↓p-STAT3, pro-caspase-9, ΔΨm | [55] |
| Terpenoids | Elemene | (family: Zingiberaceae) Curcuma zedoaria Roscoe Rhizoma | BGC-823 | 20, 40, 80, 160 μg/mL: 24 h | Induction of apoptosis | ↑Bax, p-ERK 1/2 ↓Bcl-2 | [56] |
| Terpenoids | Grifolin | (family: Albatrellaceae) Albatrellus confluens (Alb. and Schwein.) Kotl. and Pouzar | BGC-823, SGC-7901 | 10, 50 µM; 48 h | Induction of apoptosis | ↑caspase-9, -3, CDKN2 ↓MEK1, MEKK3 MEK5 | [52] |
| Terpenoids | N-butylidenephthalide | (family: Apiaceae) Angelica Sinensis Diels Radix | AGS | 25, 50, 75 µg/mL; 24 h | Induction of apoptosis | ↑REDD1 ↓mTOR | [57] |
| Terpenoids | Paeonol | (family: Paeoniaceae) Paeonia suffruticosa Andr Root bark, (family: Apocynaceae) Cynanchum paniculatum K. Schum Radix | BGC-823 | 0.1, 0.2, 0.4 mg/mL; 24, 48 h | Inhibition of proliferation, invasion, and migration Induction of apoptosis | ↓MMP-2, -9 | [58] |
| Terpenoids | Pseudolaric acid B | (family: Pinaceae) Pseudolarix kaempferi Gorden Root bark | SGC-7901/ADR | 5, 10, 20 μM/L; 24 h | Induction of apoptosis | ↑p53, Bax ↓P-gp, COX-2, Bcl-2, Bcl-xL | [59] |
| Terpenoids | Thymol | (family: Lamiaceae) Thymus quinquecostatus Celak Essential oil | AGS | 100, 200, 400 µM; 6, 12, 24 h | Induction of apoptosis | ↑Bax, c-PARP, caspase-8, caspase-7, caspase-9 ↓ΔΨm | [48] |
| Terpenoids | Toosendanin | (family: Meliaceae) Melia toosendan Sieb et zucc Cortex or Fructus | SGC-7901 | 0.5, 1 µM; 48 h | Inhibition of invasion, migration and EMT Induction of apoptosis | ↑E-cadherin ↓β-catenin | [60] |
| ↑miR-200a |
| Classification | Compound | Source | Experimental Model | Dose; Duration | Efficacy | Mechanism | References |
|---|---|---|---|---|---|---|---|
| Alkaloids | Phenolic alkaloids | (family: Menispermaceae) Menispermum dauricum DC. Rhizoma | Nude mice/SGC-7901 | 5, 10, 20 mg/kg/week; 3 weeks | Suppression of tumor growth | [62] | |
| Flavonoids | Trifolirhizin | (family: Fabaceae) Sophora flavescens Aiton. Radix | BALB/C nude mice/MKN-45 | 1–3 mg/kg; 3 weeks | Retardation of tumor growth | ↑c-caspase-3 ↓ΔΨm | [37] |
| Polyphenols | 2,7-dihydroxy-3-methylanthraquinone (DDMN) | (family: Rubiaceae) Hedyotis diffusa Wild. Herba | nude mice/SGC-7901 | 40 mg/kg; 5, 10, 15, 20 days | Inhibition of gastric cancer cell growth | ↑Bax, Bad, c-caspase-3, -9, cyt c ↓Bcl-xL, Bcl-2 | [40] |
| Polyphenols | Curcumin | (family: Zingiberaceae) Curcuma longa L. Rhizoma | BALB/C mice/MFC | 20, 40, 60 μM/L/day; 60 days | Inhibition of tumor growth Induction of apoptosis Activation of immune cells | ↑IFN-γ, TNF-α, granzyme B, perforin ↓DEC1, HIF-1α, STAT3, VEGF | [63] |
| Polyphenols | Licochalone A | (family: Fabaceae) Glycyrrhiza glabra L. Radix | BALB/ca nude mice/MKN-45 | 10 mg/kg/day; 33 days | Inhibition of tumor growth | [46] | |
| Terpenoids | Elemene | (family: Zingiberaceae) Curcuma longa L. Rhizoma | BALB/c athymic nude mice/BGC-823 | 200 mg/kg/day; 15 days | Retardation of tumor growth | [56] | |
| Terpenoids | Grifolin | (family: Albatrellaceae) Albatrellus confluens (Alb. and Schwein.) Kotl. and Pouzar | Balb/c nude mice/BGC-823, SGC-7901 | 15 mg/kg; 2 days | Improvement of survival time | [52] |
4. Role of Autophagy in Gastric Cancer Treatment Mediated by Natural Bioactive Food Products
5. Role of Bioactive Natural Compounds to Arrest Cell Cycle in Gastric Cancer
6. Anti-Angiogenesis Effects of Natural Bioactive Products in Gastric Cancer
| Phase of Cell Cycle Arrest | Classification | Compound | Source | Experimental Model | Dose; Duration | Mechanism | References |
|---|---|---|---|---|---|---|---|
| G0/G1 | Alkaloids | Berberine | (family: Ranunculaceae) Coptidis japonica Makino Rhizoma | SGC-7901 | 5, 10, 20 µM; 24, 48 h | [35] | |
| G0/G1 | Phytosterols | Ginsenoside-Rh2 | (family: Araliaceae) Panax ginseng C.A. Mey Radix | SGC-7901 | 5, 10, 20 μg/mL; 24, 48 h | ↑Bax ↓Bcl-2 | [38] |
| G0/G1 | Terpenoids | Crosolic acid | (family: Actinidiaceae) Actinidia valvata Dunn Radix | BGC-823 | 20, 40, 80 μg/mL; 72 h | ↑Bax, smac, IκBα ↓Fas, Bcl-2, p65, p-IκBα, NF-κB | [54] |
| G1 | Polyphenols | Rottlerin | (family: Euphorbiaceae) Mallotus philipensis Muell. | SGC-7901, MGC-803 | 2, 4, 8 µM; 24 h | ↑LC3-II ↓mTOR, Skp2 | [83] |
| G1/S | Terpenoids | Toosendanin | (family: Meliaceae) Melia toosendan Sieb et Zucc Cortex et Fructus | (1) AGS (2) HGC-27 | (1) 0.5, 1, 2 μM; 48 h (2) 0.5, 1, 2 μM; 36 h | ↑c-caspase-3, -8, -9, c-PARP, Bax, p-p38 ↓Bcl-2, Bcl-xL, Mcl-1, survivin, XIAP | [92] |
| S | Alkaloids | Cyclovirobuxine D | (family: Buxaceae) Buxus microphylla Richardii Radix | MGC-803, MKN-28 | 30, 60, 120 μM/L; 48 h | ↑c-caspase-3, Bax ↓Bcl-2 | [36] |
| S | Alkaloids | GFG-3a | (family: Meripilaceae) Grifola frondose (Diks.) Gray Mycelia | SGC-7901 | 100, 200 μg/mL; 24, 48 h | ↑RBBP4, caspase-3, -8, p53, Bax, Bad ↓RUVBL, NPM, Bcl-2, Bcl-xL, PI3K, Akt1 | [64] |
| G2/M | Flavonoids | Trifolirhizin | (family: Fabaceae) Sophora flavescens Aiton. Radix | MKN-45 | 20, 30, 40 µg/mL; 48 h | ↑caspase-9, -3, c-PARP, p53, p38 ↓EGFR, CDC2, cyclin B, ΔΨm | [37] |
| G2/M | Phytosterols | Tanshinone IIA | (family: Lamiaceae) Salviae miltiorrhiza Bunge. Radix | AGS | 2.0, 3.7, 5.5 µg/mL; 24, 48 h | ↑TNF-α, Fas, p-p38, p-JNK, p53, p21, caspase-8, -3 ↓p-ERK, CDC2, cyclin A, cyclin B1 | [39] |
| G2/M | Terpenoids | Deacetylisovaltratum | (family: Caprifoliaceae) Patrinia heterophylla Bunge. | (1) AGS (2) HGC-27 | (1) 4, 8, 16 μM; 24 h (2) 10, 20, 30 μM; 24 h | ↑p21, caspase-3, c-PARP ↓p-STAT3, pro-caspase-9, ΔΨm | [55] |
7. Anti-Metastasis Effects of Bioactive Compounds in Gastric Cancer
| Classification | Compound | Source | Experimental Model | Doses | Efficacy | Mechanisms | Reference |
|---|---|---|---|---|---|---|---|
| Alkaloids | Evodiamine | (family: Rutaceae) Tetradium ruticarpum | AGS, SGC-7901 | 2 µM; 48 h | Inhibition of EMT | ↓β-catenin, cyclin D1, c-Myc | [107] |
| Organosulfur compounds | Sulforaphane | (family: Brassicaceae) Brassica oleracea var. italica Plenk | AGS, MKN-45 | 31.25, 62.5, 125, 250 μg/mL; 48 h | Inhibition of metastasis | ↑CDX1, CDX2 | [102] |
| ↑miR-326, miR-9 | |||||||
| Polyphenols | Isoliquiritigenin | (family: Fabaceae) Glycyrrhiza glabra Radix | MKN-28 | 20 µM; 24, 48, 72 h | Inhibition of migration, invasion, Induction of apoptosis and autophagy | ↓Caspase-3, Bax, Bcl-2, PI3K, Akt, mTOR | [103] |
| Polyphenols | Dehydroeffusol | (family: Juncaceae) Juncus effusus L. Radix et Medulla | AGS, SGC-7901 | 12, 24, 48 µM; 24 h | Reduction of cell–cell adherent junction | ↓VE-cadherin, MMP-2 | [104] |
| Polyphenols | Paeonol | (family: Paeoniaceae) Paeonia suffruticosa Andr. Cortex, (family: Asclepiadaceae) Cynanchum paniculatum K. Schum Radix | BGC-823 | 0.1, 0.2, 0.4 mg/mL; 24, 48 h | Inhibition of proliferation, invasion, and migration, Induction of apoptosis | ↓MMP-2, MMP-9 | [58] |
| Polyphenols | Baicalein | (Lamiaceae) Scutellaria baicalensis Georgi Radix | AGS | 25, 50 µM; 24 h | Inhibition of motility, migration, invasion | ↓N-cadherin, vimentin, ZEB1, ZEB2, TGF-β/Smad4 | [105] |
| Terpenoids | Andrographolide | (family: Acanthaceae) Andrographis paniculata Nees Herba | SGC-7901 | 5, 20, 40 µg/mL; 24, 48, 72 h | Inhibition of proliferation, invasion, metastasis | ↑Bax, Bik, TIMP-1/2, ↓Bcl-2, CD147, MMP-2, MMP-9, survivin | [106] |
| Terpenoids | Toosendanin | (family: Meliaceae) Melia toosendan Sieb et Zucc Cortex et Fructus | SGC-7901 | 0.5, 1 µM; 48 h | Inhibition of invasion, migration, EMTInduction of apoptosis and cell cycle arrest | ↑E-cadherin ↓β-catenin | [60] |
| ↑miR-200a |
8. Chemotherapy Resistance and Natural Bioactive Products in Gastric Cancer
9. Limitation and Future Perspectives of Natural Bioactive Food Products in Gastric Cancer Treatments
10. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; El Hajj, N.; Sittler, S.; Lammert, N.; Barnes, R.; Meloni-Ehrig, A. Gastric cancer: Classification, histology and application of molecular pathology. J. Gastrointest. Oncol. 2012, 3, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Crew, K.D.; Neugut, A.I. Epidemiology of gastric cancer. World J. Gastroenterol. 2006, 12, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Rawla, P.; Barsouk, A. Epidemiology of gastric cancer: Global trends, risk factors and prevention. Gastroenterol. Rev. 2019, 14, 26–38. [Google Scholar] [CrossRef]
- A Ajani, J.; D’Amico, T.A.; Almhanna, K.; Bentrem, D.J.; Chao, J.; Das, P.; Denlinger, C.S.; Fanta, P.; Farjah, F.; Fuchs, C.S.; et al. Gastric Cancer, Version 3.2016, NCCN Clinical Practice Guidelines in Oncology. J. Natl. Compr. Cancer Netw. 2016, 14, 1286–1312. [Google Scholar] [CrossRef] [PubMed]
- Ronellenfitsch, U.; Schwarzbach, M.; Hofheinz, R.; Kienle, P.; Kieser, M.; E Slanger, T.; Jensen, K. GE adenocarcinoma meta-analysis group Perioperative chemo(radio)therapy versus primary surgery for resectable adenocarcinoma of the stomach, gastroesophageal junction, and lower esophagus. Cochrane Database Syst. Rev. 2013, 2013, 008107. [Google Scholar] [CrossRef] [Green Version]
- Diaz-Nieto, R.; Orti-Rodríguez, R.; Winslet, M. Post-surgical chemotherapy versus surgery alone for resectable gastric cancer. Cochrane Database Syst. Rev. 2013, 2013, CD008415. [Google Scholar] [CrossRef]
- Oba, K.; Paoletti, X.; Alberts, S.; Bang, Y.-J.; Benedetti, J.; Bleiberg, H.; Catalano, P.; Lordick, F.; Michiels, S.; Morita, S.; et al. Disease-Free Survival as a Surrogate for Overall Survival in Adjuvant Trials of Gastric Cancer: A Meta-Analysis. J. Natl. Cancer Inst. 2013, 105, 1600–1607. [Google Scholar] [CrossRef] [Green Version]
- Gibson, R.J.; Keefe, D.M.K. Cancer chemotherapy-induced diarrhoea and constipation: Mechanisms of damage and prevention strategies. Support. Care Cancer 2006, 14, 890–900. [Google Scholar] [CrossRef] [PubMed]
- Staff, N.P.; Grisold, A.; Grisold, W.; Windebank, A.J. Chemotherapy-induced peripheral neuropathy: A current review. Ann. Neurol. 2017, 81, 772–781. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Ma, J.; Zhou, W.; Cao, B.; Zhou, X.; Yang, Z.; Zhang, H.; Zhao, Q.; Fan, D.; Hong, L. Molecular mechanisms and theranostic potential of miRNAs in drug resistance of gastric cancer. Expert Opin. Ther. Targets 2017, 21, 1063–1075. [Google Scholar] [CrossRef]
- Singh, N.; Baby, D.; Rajguru, J.P.; Patil, P.B.; Thakkannavar, S.S.; Pujari, V.B. Inflammation and cancer. Ann. Afr. Med. 2019, 18, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Li, M. Helicobacter pylori eradication for preventing gastric cancer. World J. Gastroenterol. 2014, 20, 5660–5665. [Google Scholar] [CrossRef]
- Sokolova, O.; Naumann, M. NF-kappaB Signaling in Gastric Cancer. Toxins 2017, 9, 119. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Jeong, A.J.; Ye, S.-K. Highlighted STAT3 as a potential drug target for cancer therapy. BMB Rep. 2019, 52, 415–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naylor, M.S.; Stamp, G.W.; Foulkes, W.; Eccles, D.; Balkwill, F. Tumor necrosis factor and its receptors in human ovarian cancer. Potential role in disease progression. J. Clin. Investig. 1993, 91, 2194–2206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Lin, Y. Tumor necrosis factor and cancer, buddies or foes? Acta Pharmacol. Sin. 2008, 29, 1275–1288. [Google Scholar] [CrossRef] [Green Version]
- Rabelo, A.C.S.; Camini, F.C.; Bittencourt, M.M.; Lacerda, K.; De Lima, W.G.; Costa, D.C. Baccharis trimera (carqueja) promotes gastroprotection on ethanol-induced acute gastric ulcer. Adv. Tradit. Med. 2020, 20, 563–570. [Google Scholar] [CrossRef]
- Bastos, J.; Lunet, N.; Peleteiro, B.; Lopes, C.; Barros, H. Dietary patterns and gastric cancer in a Portuguese urban population. Int. J. Cancer 2010, 127, 433–441. [Google Scholar] [CrossRef]
- Nagata, C.; Takatsuka, N.; Kawakami, N.; Shimizu, H. A prospective cohort study of soy product intake and stomach cancer death. Br. J. Cancer 2002, 87, 31–36. [Google Scholar] [CrossRef] [Green Version]
- Steevens, J.; Schouten, L.J.; Goldbohm, R.A.; Brandt, P.V.D. Vegetables and fruits consumption and risk of esophageal and gastric cancer subtypes in the Netherlands Cohort Study. Int. J. Cancer 2011, 129, 2681–2693. [Google Scholar] [CrossRef] [PubMed]
- Mao, Q.-Q.; Xu, X.-Y.; Shang, A.; Gan, R.-Y.; Wu, D.-T.; Atanasov, A.G.; Li, H.-B. Phytochemicals for the Prevention and Treatment of Gastric Cancer: Effects and Mechanisms. Int. J. Mol. Sci. 2020, 21, 570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Long, Y.; Ni, L.; Yuan, X.; Yu, N.; Wu, R.; Tao, J.; Zhang, Y. Anticancer effect of berberine based on experimental animal models of various cancers: A systematic review and meta-analysis. BMC Cancer 2019, 19, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Hassanalilou, T.; Ghavamzadeh, S.; Khalili, L. Curcumin and Gastric Cancer: A Review on Mechanisms of Action. J. Gastrointest. Cancer 2019, 50, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Mahalanobish, S.; Saha, S.; Ghosh, S.; Sil, P.C. Natural products: An upcoming therapeutic approach to cancer. Food Chem. Toxicol. 2019, 128, 240–255. [Google Scholar] [CrossRef]
- Kim, H.-J.; Um, J.-Y.; Kim, Y.-K. Glutathione S-transferase gene polymorphism in Korean subjects with gastric and colorectal cancer. Orient. Pharm. Exp. Med. 2012, 12, 307–312. [Google Scholar] [CrossRef]
- Mann, J. Natural products in cancer chemotherapy: Past, present and future. Nat. Rev. Cancer 2002, 2, 143–148. [Google Scholar] [CrossRef]
- Wang, L.; Dong, C.; Li, X.; Han, W.; Su, X. Anticancer potential of bioactive peptides from animal sources (Review). Oncol. Rep. 2017, 38, 637–651. [Google Scholar] [CrossRef] [Green Version]
- Gras, M.; Vallard, A.; Brosse, C.; Beneton, A.; Sotton, S.; Guyotat, D.; Fournel, P.; Daguenet, E.; Magné, N.; Morisson, S. Use of Complementary and Alternative Medicines among Cancer Patients: A Single-Center Study. Oncology 2019, 97, 18–25. [Google Scholar] [CrossRef]
- Li, X.; Yang, G.; Li, X.; Zhang, Y.; Yang, J.; Chang, J.; Sun, X.; Zhou, X.; Guo, Y.; Xu, Y.; et al. Traditional Chinese medicine in cancer care: A review of controlled clinical studies published in chinese. PLoS ONE 2013, 8, e60338. [Google Scholar]
- Wode, K.; Henriksson, R.; Sharp, L.; Stoltenberg, A.; Nordberg, J.H. Cancer patients’ use of complementary and alternative medicine in Sweden: A cross-sectional study. BMC Complement. Altern. Med. 2019, 19, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kristoffersen, A.E.; Stub, T.; Broderstad, A.R.; Hansen, A.H. Use of traditional and complementary medicine among Norwegian cancer patients in the seventh survey of the Tromsø study. BMC Complement. Altern. Med. 2019, 19, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta Bioenerg. 2016, 1863, 2977–2992. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, N.; Li, K.; Chen, J.; Qiu, L.; Zhang, J. Integration of microRNA-mRNA profiles and pathway analysis of plant isoquinoline alkaloid berberine in SGC-7901 gastric cancers cells. Drug Des. Devel. Ther. 2018, 12, 393–408. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Tan, Z.; Chen, J.; Dong, C. Cyclovirobuxine D Inhibits Cell Proliferation and Induces Mitochondria-Mediated Apoptosis in Human Gastric Cancer Cells. Molecules 2015, 20, 20659–20668. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Ma, J.; Qiu, H.; Yang, L.; Cao, L.; Shen, J. Anti-proliferation effects of trifolirhizin on MKN45 cells and possible mechanism. Oncol. Rep. 2016, 36, 2785–2792. [Google Scholar] [CrossRef] [Green Version]
- Qian, J.; Li, J.; Jia, J.-G.; Jin, X.; Yu, D.-J.; Guo, C.-X.; Xie, B.; Qian, L.-Y. Ginsenoside-Rh2 Inhibits Proliferation and Induces Apoptosis of Human Gastric Cancer SGC-7901 Side Population Cells. Asian Pac. J. Cancer Prev. 2016, 17, 1817–1821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, C.-C. Tanshinone IIA inhibits gastric carcinoma AGS cells through increasing p-p38, p-JNK and p53 but reducing p-ERK, CDC2 and cyclin B1 expression. Anticancer. Res. 2014, 34, 7097–7110. [Google Scholar] [PubMed]
- Zhu, H.; Zheng, Z.; Zhang, J.; Liu, X.; Liu, Y.; Yang, W.; Liu, Y.; Zhang, T.; Zhao, Y.; Liu, Y.; et al. Anticancer effect of 2,7-dihydroxy-3-methylanthraquinone on human gastric cancer SGC-7901 cells in vitro and in vivo. Pharm. Biol. 2016, 54, 285–292. [Google Scholar] [CrossRef]
- Luo, Y.; Yu, H.; Yang, Y.; Tian, W.; Dong, K.; Shan, J.; Ma, X. A flavonoid compound from Chrysosplenium nudicaule inhibits growth and induces apoptosis of the human stomach cancer cell line SGC-7901. Pharm. Biol. 2016, 54, 1133–1139. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Zhang, W.; Guo, Y.; Li, Z.; Chen, X.; Wang, Y.; Du, Y.; Zang, W.; Zhao, G. Curcumin inhibits cell growth and induces cell apoptosis through upregulation of miR-33b in gastric cancer. Tumor Biol. 2016, 37, 13177–13184. [Google Scholar] [CrossRef]
- Pan, H.; Wang, B.-H.; Lv, W.; Jiang, Y.; He, L. Esculetin induces apoptosis in human gastric cancer cells through a cyclophilin D-mediated mitochondrial permeability transition pore associated with ROS. Chem. Interactions 2015, 242, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Si, N.; Ma, Y.; Ge, D.; Yu, X.; Fan, A.; Wang, X.; Hu, J.; Wei, P.; Chen, J.; et al. Hydroxysafflor-Yellow A Induces Human Gastric Carcinoma BGC-823 Cell Apoptosis by Activating Peroxisome Proliferator-Activated Receptor Gamma (PPARgamma). Med. Sci. Monit. 2018, 24, 803–811. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Cao, A.; Wang, L.; Wu, D. Kurarinone Synergizes TRAIL-Induced Apoptosis in Gastric Cancer Cells. Cell Biophys. 2015, 72, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhang, X.; Wang, Y.; Sun, Q.; Chen, M.; Liu, S.; Zou, X. Licochalcone A suppresses hexokinase 2-mediated tumor glycolysis in gastric cancer via downregulation of the Akt signaling pathway. Oncol. Rep. 2017, 39, 1181–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, E.B.; Trinh, T.A.; Lee, T.K.; Yamabe, N.; Kang, K.S.; Song, J.H.; Choi, S.; Lee, S.; Jang, T.S.; Kim, K.H.; et al. Curcuzedoalide contributes to the cytotoxicity of Curcuma zedoaria rhizomes against human gastric cancer AGS cells through induction of apoptosis. J. Ethnopharmacol. 2018, 213, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.-H.; Kim, Y.-S.; Kim, E.-K.; Hwang, J.-W.; Jeong, J.-H.; Dong, X.; Lee, J.-W.; Moon, S.-H.; Jeon, B.-T.; Park, P.-J. Anticancer Effect of Thymol on AGS Human Gastric Carcinoma Cells. J. Microbiol. Biotechnol. 2016, 26, 28–37. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, Q.; Jiang, Y.; Li, F.; Xin, H. Effects of ophiopogonin B on the proliferation and apoptosis of SGC-7901 human gastric cancer cells. Mol. Med. Rep. 2016, 13, 4981–4986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.; Gu, W.; Shen, Z.; Wang, F. Anticancer Activity of Phloretin Against Human Gastric Cancer Cell Lines Involves Apoptosis, Cell Cycle Arrest, and Inhibition of Cell Invasion and JNK Signalling Pathway. Med. Sci. Monit. 2018, 24, 6551–6558. [Google Scholar] [CrossRef]
- Asl, E.A.; Mehrabadi, J.F.; Afshar, D.; Noorbazargan, H.; Tahmasebi, H.; Rahimi, A. Apoptotic Effects of Linum album Extracts on AGS Human Gastric Adenocarcinoma Cells and ZNF703 Oncogene Expression. Asian Pac. J. Cancer Prev. 2018, 19, 2911–2916. [Google Scholar] [CrossRef]
- Wu, Z.; Li, Y. Grifolin exhibits anti-cancer activity by inhibiting the development and invasion of gastric tumor cells. Oncotarget 2017, 8, 21454–21460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, T.C.; Lai, K.-H.; Su, J.-H.; Wu, Y.-J.; Sheu, J.-H. 7-Acetylsinumaximol B Induces Apoptosis and Autophagy in Human Gastric Carcinoma Cells through Mitochondria Dysfunction and Activation of the PERK/eIF2alpha/ATF4/CHOP Signaling Pathway. Mar. Drugs 2018, 16, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Q.-L.; Li, H.-L.; Li, Y.-C.; Liu, Z.-W.; Guo, X.-H.; Cheng, Y.-J. CRA (Crosolic Acid) isolated from Actinidia valvata Dunn.Radix induces apoptosis of human gastric cancer cell line BGC823 in vitro via down-regulation of the NF-kappaB pathway. Food Chem. Toxicol. 2017, 105, 475–485. [Google Scholar] [CrossRef]
- Zhang, D.; Zhang, B.; Zhou, L.-X.; Zhao, J.; Yan, Y.-Y.; Li, Y.-L.; Zeng, J.-M.; Wang, L.-L.; Yang, B.; Lin, N.-M. Deacetylisovaltratum disrupts microtubule dynamics and causes G2/M-phase arrest in human gastric cancer cells in vitro. Acta Pharmacol. Sin. 2016, 37, 1597–1605. [Google Scholar] [CrossRef]
- Li, P.; Zhou, X.; Sun, W.; Sheng, W.; Tu, Y.; Yu, Y.; Dong, J.; Ye, B.; Zheng, Z.; Lu, M. Elemene Induces Apoptosis of Human Gastric Cancer Cell Line BGC-823 via Extracellular Signal-Regulated Kinase (ERK) 1/2 Signaling Pathway. Med. Sci. Monit. 2017, 23, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Liao, K.-F.; Chiu, T.-L.; Huang, S.-Y.; Hsieh, T.-F.; Chang, S.-F.; Ruan, J.-W.; Chen, S.-P.; Pang, C.-Y.; Chiu, S.-C. Anti-Cancer Effects of Radix Angelica Sinensis (Danggui) and N-Butylidenephthalide on Gastric Cancer: Implications for REDD1 Activation and mTOR Inhibition. Cell. Physiol. Biochem. 2018, 48, 2231–2246. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Z.-K.; Li, C.-L.; Jin, Y.; Liu, Y.-Z.; Zhang, X.; Zhang, F.; Ning, L.-N.; Liang, E.-S.; Ma, M.; Gao, W.; et al. Paeonol exerts potential activities to inhibit the growth, migration and invasion of human gastric cancer BGC823 cells via downregulating MMP-2 and MMP-9. Mol. Med. Rep. 2017, 16, 7513–7519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, F.; Li, K.; Chen, S.; Liu, Y.; Li, Y. Pseudolaric Acid B Circumvents Multidrug Resistance Phenotype in Human Gastric Cancer SGC7901/ADR Cells by Downregulating Cox-2 and P-gp Expression. Cell Biophys. 2014, 71, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Huang, Y.-X.; Zhang, R.; Hou, L.-D.; Liu, H.; Chen, X.-Y.; Zhu, J.-S.; Zhang, J. Toosendanin suppresses oncogenic phenotypes of human gastric carcinoma SGC7901 cells partly via miR200amediated downregulation of beta-catenin pathway. Int. J. Oncol. 2017, 51, 1563–1573. [Google Scholar] [CrossRef] [Green Version]
- Kapoor, S.; Dharmesh, S.M. Pectic Oligosaccharide from tomato exhibiting anticancer potential on a gastric cancer cell line: Structure-function relationship. Carbohydr. Polym. 2017, 160, 52–61. [Google Scholar] [CrossRef]
- Zhang, H.; Wu, D.; Du, J.; Zhang, Y.; Su, Y. Anti-tumor effects of phenolic alkaloids of menispermum dauricum on gastric cancer in vivo and in vitro. J. Cancer Res. Ther. 2018, 14, 505. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-P.; Wang, Q.-X.; Lin, H.-P.; Chang, N. Anti-tumor bioactivities of curcumin on mice loaded with gastric carcinoma. Food Funct. 2017, 8, 3319–3326. [Google Scholar] [CrossRef] [PubMed]
- Cui, F.; Zan, X.; Li, Y.; Sun, W.; Yang, Y.; Ping, L. Grifola frondosaGlycoprotein GFG-3a Arrests S phase, Alters Proteome, and Induces Apoptosis in Human Gastric Cancer Cells. Nutr. Cancer 2016, 68, 267–279. [Google Scholar] [CrossRef] [PubMed]
- Kong, G.-M.; Tao, W.-H.; Diao, Y.-L.; Fang, P.-H.; Wang, J.-J.; Bo, P.; Qian, F. Melittin induces human gastric cancer cell apoptosis via activation of mitochondrial pathway. World J. Gastroenterol. 2016, 22, 3186–3195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.-Z.; Wang, L.; Liu, D.-W.; Tang, G.-Y.; Zhang, H.-Y. Synergistic Inhibitory Effect of Berberine and d-Limonene on Human Gastric Carcinoma Cell Line MGC803. J. Med. Food 2014, 17, 955–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Zhao, L.-M.; Dai, S.-L.; Cui, W.-X.; Lv, H.-L.; Chen, L.; Shan, B.-E. Periplocin Extracted from Cortex Periplocae Induced Apoptosis of Gastric Cancer Cells via the ERK1/2-EGR1 Pathway. Cell Physiol. Biochem. 2016, 38, 1939–1951. [Google Scholar] [CrossRef]
- Mansingh, D.P.; Oj, S.; Sali, V.K.; Vasanthi, H.R. [6]-Gingerol-induced cell cycle arrest, reactive oxygen species generation, and disruption of mitochondrial membrane potential are associated with apoptosis in human gastric cancer (AGS) cells. J. Biochem. Mol. Toxicol. 2018, 32, e22206. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; He, D.; Yao, Z.; Klionsky, D.J. The machinery of macroautophagy. Cell Res. 2014, 24, 24–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.A.; Rhim, H. Therapeutic implication of autophagy in neurodegenerative diseases. BMB Rep. 2017, 50, 345–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, R.; Zeh, H.J.; Lotze, M.T.; Tang, D. The Beclin 1 network regulates autophagy and apoptosis. Cell Death Differ. 2011, 18, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Hannan, M.A.; Dash, R.; Rahman, M.H.; Islam, R.; Uddin, M.J.; Sohag, A.A.M.; Rahman, M.H.; Rhim, H. Phytochemicals as a Complement to Cancer Chemotherapy: Pharmacological Modulation of the Autophagy-Apoptosis Pathway. Front. Pharmacol. 2021, 12, 639628. [Google Scholar] [CrossRef]
- Rahman, M.A.; Rahman, M.S.; Rahman, M.H.; Rasheduzzaman, M.; Mamun-Or-Rashid, A.; Uddin, M.J.; Rahman, M.R.; Hwang, H.; Pang, M.G.; Rhim, H. Modulatory Effects of Autophagy on APP Processing as a Potential Treatment Target for Alzheimer’s Disease. Biomedicines 2021, 9, 5. [Google Scholar] [CrossRef]
- Onorati, A.V.; Dyczynski, M.; Ojha, R.; Amaravadi, R.K. Targeting autophagy in cancer. Cancer 2018, 124, 3307–3318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandhair, H.K.; Arambasic, M.; Novak, U.; Radpour, R. Molecular modulation of autophagy: New venture to target resistant cancer stem cells. World J. Stem Cells 2020, 12, 303–322. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.S.; Rahman, M.A.; Kabir, M.T.; Behl, T.; Mathew, B.; Perveen, A.; Barreto, G.E.; Bin-Jumah, M.N.; Abdel-Daim, M.M.; Ashraf, G.M. Multifarious roles of mTOR signaling in cognitive aging and cerebrovascular dysfunction of Alzheimer’s disease. Iubmb. Life 2020, 72, 1843–1855. [Google Scholar] [CrossRef]
- Rahman, M.A.; Cho, Y.; Nam, G.; Rhim, H. Antioxidant Compound, Oxyresveratrol, Inhibits APP Production through the AMPK/ULK1/mTOR-Mediated Autophagy Pathway in Mouse Cortical Astrocytes. Antioxidants 2021, 10, 408. [Google Scholar] [CrossRef]
- Tanida, I.; Ueno, T.; Kominami, E. LC3 conjugation system in mammalian autophagy. Int. J. Biochem. Cell Biol. 2004, 36, 2503–2518. [Google Scholar] [CrossRef]
- Dooley, H.C.; Razi, M.; Polson, H.E.J.; Girardin, S.E.; Wilson, M.I.; Tooze, S.A. WIPI2 Links LC3 Conjugation with PI3P, Autophagosome Formation, and Pathogen Clearance by Recruiting Atg12–5-16L1. Mol. Cell 2014, 55, 238–252. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.A.; Rahman, M.H.; Hossain, M.S.; Biswas, P.; Islam, R.; Uddin, M.J.; Rahman, M.H.; Rhim, H. Molecular Insights into the Multifunctional Role of Natural Compounds: Autophagy Modulation and Cancer Prevention. Biomedicines 2020, 8, 517. [Google Scholar] [CrossRef]
- Rahman, M.A.; Cho, Y.; Hwang, H.; Rhim, H. Pharmacological Inhibition of O-GlcNAc Transferase Promotes mTOR-Dependent Autophagy in Rat Cortical Neurons. Brain Sci. 2020, 10, 958. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Zhang, X.; Jiang, Y.; Su, Q.; Li, Q.; Li, Z. Autophagy: Mechanisms and Therapeutic Potential of Flavonoids in Cancer. Biomolecules 2021, 11, 135. [Google Scholar] [CrossRef]
- Song, J.; Zhou, Y.; Gong, Y.; Liu, H.; Tang, L. Rottlerin promotes autophagy and apoptosis in gastric cancer cell lines. Mol. Med. Rep. 2018, 18, 2905–2913. [Google Scholar] [CrossRef] [PubMed]
- Batool, S.; Joseph, T.P.; Hussain, M.; Vuai, M.S.; Khinsar, K.H.; Din, S.R.U.; Padhiar, A.A.; Zhong, M.; Ning, A.; Zhang, W.; et al. LP1 from Lentinula edodes C91-3 Induces Autophagy, Apoptosis and Reduces Metastasis in Human Gastric Cancer Cell Line SGC-7901. Int. J. Mol. Sci. 2018, 19, 2986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, Y.H.; Bishayee, K.; Rahman, A.; Hong, J.S.; Lim, S.-S.; Huh, S.-O. Morus alba Accumulates Reactive Oxygen Species to Initiate Apoptosis via FOXO-Caspase 3-Dependent Pathway in Neuroblastoma Cells. Mol. Cells 2015, 38, 630–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.A.; Bishayee, K.; Sadra, A.; Huh, S.-O. Oxyresveratrol activates parallel apoptotic and autophagic cell death pathways in neuroblastoma cells. Biochim. Biophys Acta Gen. Subj. 2017, 1861, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.W.; Lee, S.Y.; Kim, M.; Cheon, C.; Ko, S.-G. Kaempferol induces autophagic cell death via IRE1-JNK-CHOP pathway and inhibition of G9a in gastric cancer cells. Cell Death Dis. 2018, 9, 875. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Saralamma, V.V.G.; Kim, S.M.; Ha, S.E.; Raha, S.; Lee, W.S.; Kim, E.H.; Lee, S.J.; Heo, J.D.; Kim, G.S. Pectolinarigenin Induced Cell Cycle Arrest, Autophagy, and Apoptosis in Gastric Cancer Cell via PI3K/AKT/mTOR Signaling Pathway. Nutrients 2018, 10, 1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Liu, S.; Feng, Q.; Huang, X.; Wang, X.; Peng, Y.; Zhao, Z.; Liu, Z. Perilaldehyde activates AMP-activated protein kinase to suppress the growth of gastric cancer via induction of autophagy. J. Cell. Biochem. 2019, 120, 1716–1725. [Google Scholar] [CrossRef]
- Wang, K.; Liu, R.; Li, J.; Mao, J.; Lei, Y.; Wu, J.; Zeng, J.; Zhang, T.; Wu, H.; Chen, L.; et al. Quercetin induces protective autophagy in gastric cancer cells: Involvement of Akt-mTOR- and hypoxia-induced factor 1alpha-mediated signaling. Autophagy 2011, 7, 966–978. [Google Scholar] [CrossRef] [Green Version]
- Mrakovcic, M.; Fröhlich, L. p53-Mediated Molecular Control of Autophagy in Tumor Cells. Biomolecules 2018, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Wu, X.; Wen, C.; Wang, H.; Wang, H.; Liu, H.; Peng, J. Toosendanin induces caspase-dependent apoptosis through the p38 MAPK pathway in human gastric cancer cells. Biochem. Biophys. Res. Commun. 2018, 505, 261–266. [Google Scholar] [CrossRef]
- Cao, Y.; Arbiser, J.; D’Amato, R.J.; D’Amore, P.A.; Ingber, D.E.; Kerbel, R.; Klagsbrun, M.; Lim, S.; Moses, M.A.; Zetter, B.; et al. Forty-Year Journey of Angiogenesis Translational Research. Sci. Transl. Med. 2011, 3, 114rv3. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Dabrosin, C.; Yin, X.; Fuster, M.M.; Arreola, A.; Rathmell, W.K.; Generali, D.; Nagaraju, G.P.; El-Rayes, B.; Ribatti, D.; et al. Broad targeting of angiogenesis for cancer prevention and therapy. Semin. Cancer Biol. 2015, 35, S224–S243. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wang, Q.; Qiao, C.; Lin, Z.; Li, X.; Huang, Y.; Zhou, T.; Li, Y.; Shen, B.; Lv, M.; et al. Potent anti-angiogenesis and anti-tumor activity of a novel human anti-VEGF antibody, MIL60. Cell. Mol. Immunol. 2014, 11, 285–293. [Google Scholar] [CrossRef] [Green Version]
- Eklund, L.; Bry, M.; Alitalo, K. Mouse models for studying angiogenesis and lymphangiogenesis in cancer. Mol. Oncol. 2013, 7, 259–282. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Wang, J.; Liang, Y.; Ge, W.; Wang, G.; Li, Y.; Chung, H.Y. Potent anti-angiogenic component in Croton crassifolius and its mechanism of action. J. Ethnopharmacol. 2015, 175, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Tsuboi, K.; Matsuo, Y.; Shamoto, T.; Shibata, T.; Koide, S.; Morimoto, M.; Guha, S.; Sung, B.; Aggarwal, B.B.; Takahashi, H.; et al. Zerumbone inhibits tumor angiogenesis via NF-kappaB in gastric cancer. Oncol. Rep. 2014, 31, 57–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manu, K.A.; Shanmugam, M.K.; Rajendran, P.; Li, F.; Ramachandran, L.; Hay, H.S.; Kannaiyan, R.; Swamy, S.N.; Vali, S.; Kapoor, S.; et al. Plumbagin inhibits invasion and migration of breast and gastric cancer cells by downregulating the expression of chemokine receptor CXCR4. Mol. Cancer 2011, 10, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Wang, J.; Lin, L.; He, L.; Wu, Y.; Zhang, L.; Yi, Z.; Chen, Y.; Pang, X.; Liu, M. Inhibition of STAT3 Signaling Pathway by Nitidine Chloride Suppressed the Angiogenesis and Growth of Human Gastric Cancer. Mol. Cancer Ther. 2012, 11, 277–287. [Google Scholar] [CrossRef] [Green Version]
- Suhail, Y.; Cain, M.P.; Vanaja, K.; Kurywchak, P.A.; Levchenko, A.; Kalluri, R. Kshitiz Systems Biology of Cancer Metastasis. Cell Syst. 2019, 9, 109–127. [Google Scholar] [CrossRef] [Green Version]
- Kiani, S.; Akhavan-Niaki, H.; Fattahi, S.; Kavoosian, S.; Jelodar, N.B.; Bagheri, N.; Zarrini, H.N. Purified sulforaphane from broccoli (Brassica oleracea var. italica) leads to alterations of CDX1 and CDX2 expression and changes in miR-9 and miR-326 levels in human gastric cancer cells. Gene 2018, 678, 115–123. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, S.; Sun, W.; Wei, C. Isoliquiritigenin inhibits proliferation and metastasis of MKN28 gastric cancer cells by suppressing the PI3K/AKT/mTOR signaling pathway. Mol. Med. Rep. 2018, 18, 3429–3436. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Meng, M.; Zhang, B.; Du, L.; Pan, Y.; Yang, P.; Gu, Z.; Zhou, Q.; Cao, Z. Dehydroeffusol effectively inhibits human gastric cancer cell-mediated vasculogenic mimicry with low toxicity. Toxicol. Appl. Pharmacol. 2015, 287, 98–110. [Google Scholar] [CrossRef]
- Chen, F.; Zhuang, M.; Peng, J.; Wang, X.; Huang, T.; Li, S.; Lin, M.; Lin, H.; Xu, Y.; Li, J.; et al. Baicalein inhibits migration and invasion of gastric cancer cells through suppression of the TGF-beta signaling pathway. Mol. Med. Rep. 2014, 10, 1999–2003. [Google Scholar] [CrossRef] [Green Version]
- Dai, L.; Wang, G.; Pan, W. Andrographolide Inhibits Proliferation and Metastasis of SGC7901 Gastric Cancer Cells. BioMed Res. Int. 2017, 2017, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.; Feng, S.; Wei, L.; Wang, Z.; Hong, D.; Wang, Q. Evodiamine, a novel inhibitor of the Wnt pathway, inhibits the self-renewal of gastric cancer stem cells. Int. J. Mol. Med. 2015, 36, 1657–1663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Li, P.; Lu, S.-M.; Ling, Z.-Q. Chemoprevention of Low-Molecular-Weight Citrus Pectin (LCP) in Gastrointestinal Cancer Cells. Int. J. Biol. Sci. 2016, 12, 746–756. [Google Scholar] [CrossRef] [PubMed]
- Kirkin, V.; Joos, S.; Zornig, M. The role of Bcl-2 family members in tumorigenesis. Biochim. Biophys. Acta 2004, 1644, 229–249. [Google Scholar] [CrossRef]
- Vasan, N.; Baselga, J.; Hyman, D.M. A view on drug resistance in cancer. Nature 2019, 575, 299–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manu, K.A.; Shanmugam, M.K.; Ramachandran, L.; Li, F.; Siveen, K.S.; Chinnathambi, A.; Zayed, M.E.; Alharbi, S.A.; Arfuso, F.; Kumar, A.P.; et al. Isorhamnetin augments the anti-tumor effect of capecitabine through the negative regulation of NF-kappaB signaling cascade in gastric cancer. Cancer Lett. 2015, 363, 28–36. [Google Scholar] [CrossRef]
- Wei, F.; Jiang, X.; Gao, H.-Y.; Gao, S.-H. Liquiritin induces apoptosis and autophagy in cisplatin (DDP)-resistant gastric cancer cells in vitro and xenograft nude mice in vivo. Int. J. Oncol. 2017, 51, 1383–1394. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Yu, J.; Wang, J.; Zhang, C.; Shang, K.; Yao, X.; Cao, B. Astragalus polysaccharide enhanced antitumor effects of Apatinib in gastric cancer AGS cells by inhibiting AKT signalling pathway. Biomed. Pharmacother. 2018, 100, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Chen, L.; Xiao, Z.; Zhu, Y.; Jiang, H.; Jin, Y.; Gu, C.; Wu, Y.; Wang, L.; Zhang, W.; et al. Potentiation of the anticancer effect of doxorubicinin drug-resistant gastric cancer cells by tanshinone IIA. Phytomedicine 2018, 51, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Guerra, A.R.; Duarte, M.F.; Duarte, I.F. Targeting Tumor Metabolism with Plant-Derived Natural Products: Emerging Trends in Cancer Therapy. J. Agric. Food Chem. 2018, 66, 10663–10685. [Google Scholar] [CrossRef] [PubMed]
- Roe, A.L.; Paine, M.F.; Gurley, B.J.; Brouwer, K.R.; Jordan, S.; Griffiths, J.C. Assessing Natural Product–Drug Interactions: An End-to-End Safety Framework. Regul. Toxicol. Pharmacol. 2016, 76, 1–6. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, S.Y.; Hwang, D.; Shin, S.; Park, J.; Kim, M.; Rahman, M.H.; Rahman, M.A.; Ko, S.-G.; Kim, B. Potential of Bioactive Food Components against Gastric Cancer: Insights into Molecular Mechanism and Therapeutic Targets. Cancers 2021, 13, 4502. https://doi.org/10.3390/cancers13184502
Kang SY, Hwang D, Shin S, Park J, Kim M, Rahman MH, Rahman MA, Ko S-G, Kim B. Potential of Bioactive Food Components against Gastric Cancer: Insights into Molecular Mechanism and Therapeutic Targets. Cancers. 2021; 13(18):4502. https://doi.org/10.3390/cancers13184502
Chicago/Turabian StyleKang, Seog Young, Dongwon Hwang, Soyoung Shin, Jinju Park, Myoungchan Kim, MD. Hasanur Rahman, Md. Ataur Rahman, Seong-Gyu Ko, and Bonglee Kim. 2021. "Potential of Bioactive Food Components against Gastric Cancer: Insights into Molecular Mechanism and Therapeutic Targets" Cancers 13, no. 18: 4502. https://doi.org/10.3390/cancers13184502
APA StyleKang, S. Y., Hwang, D., Shin, S., Park, J., Kim, M., Rahman, M. H., Rahman, M. A., Ko, S. -G., & Kim, B. (2021). Potential of Bioactive Food Components against Gastric Cancer: Insights into Molecular Mechanism and Therapeutic Targets. Cancers, 13(18), 4502. https://doi.org/10.3390/cancers13184502