
A Recurrent STAT5BN642H Driver Mutation in Feline Alimentary T Cell Lymphoma

 , , , , and
, , , , and 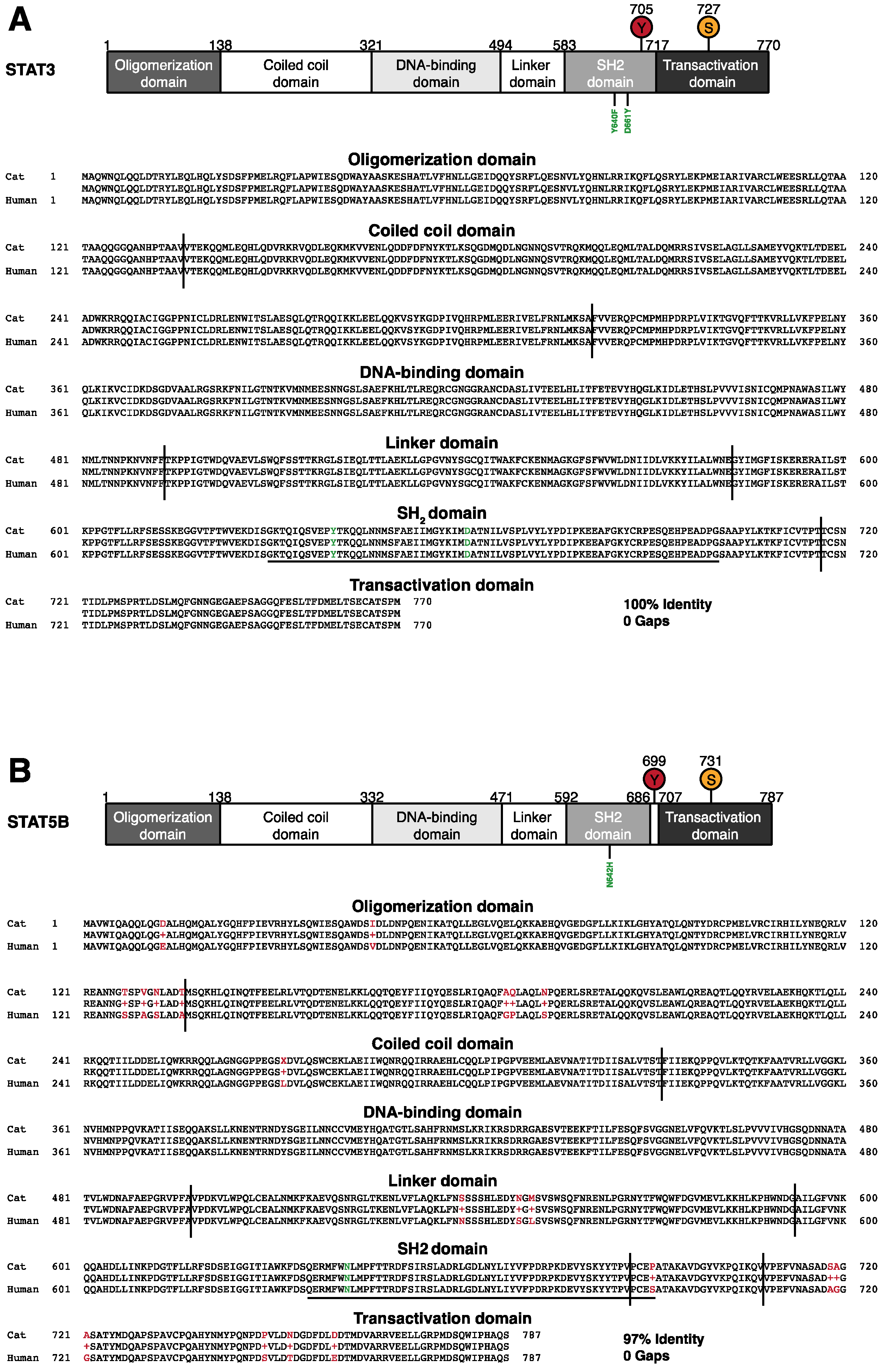{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Biological Samples
2.2. Immunohistochemistry
2.3. DNA Isolation
2.4. PCR Screening
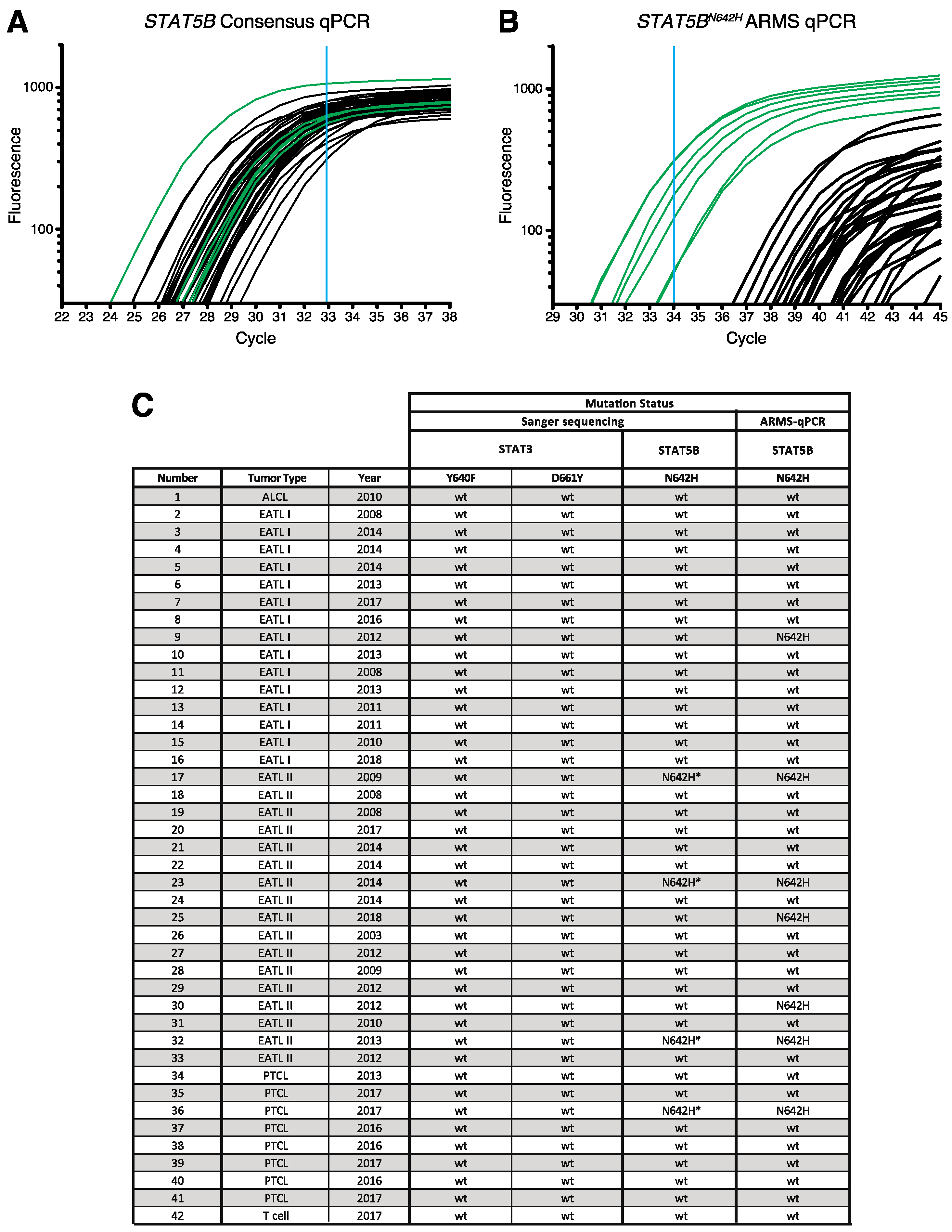
2.5. Amplification-Refractory Mutation System Assay (ARMS-qPCR)
2.6. Statistics and Bioinformatics
3. Results
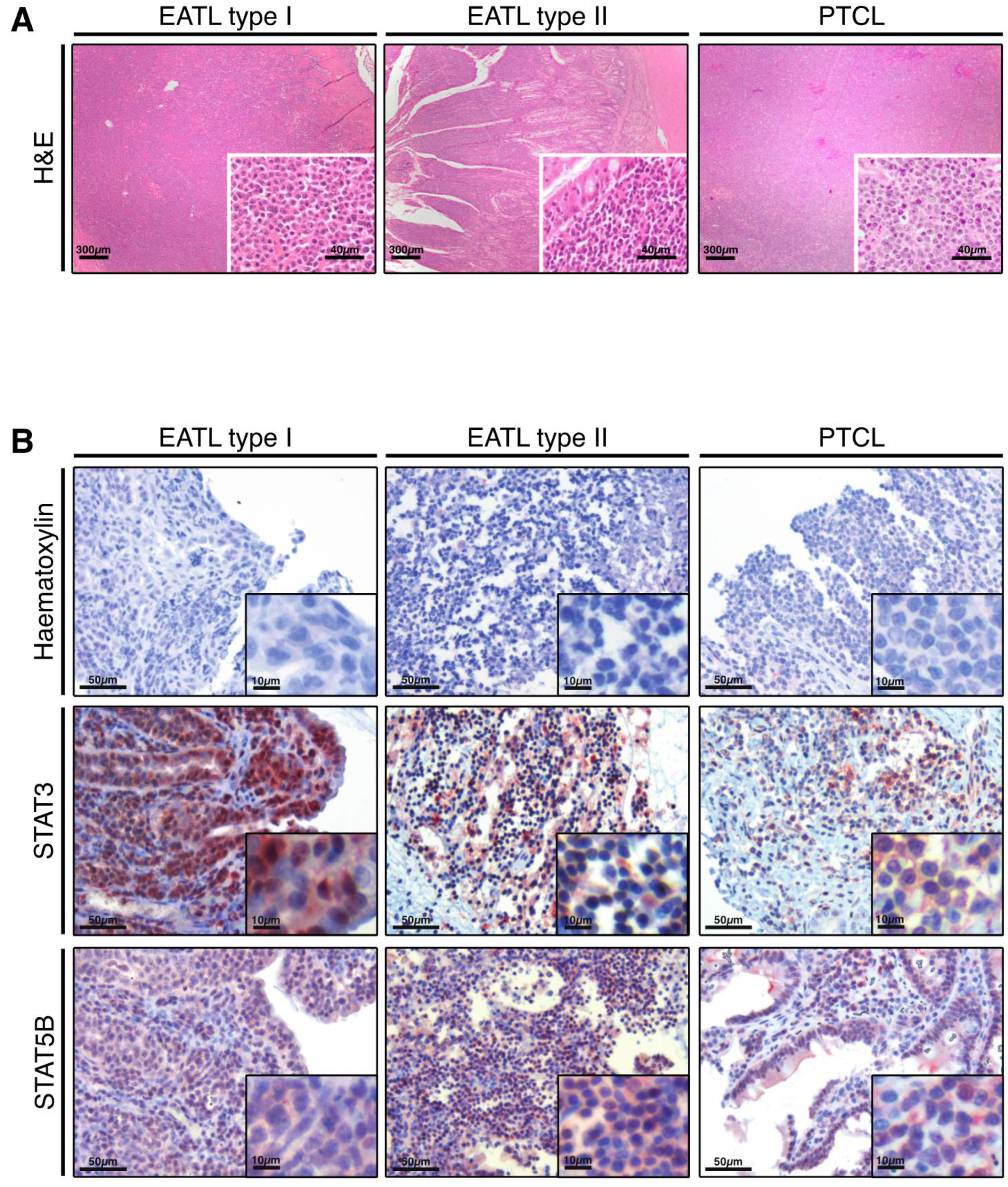
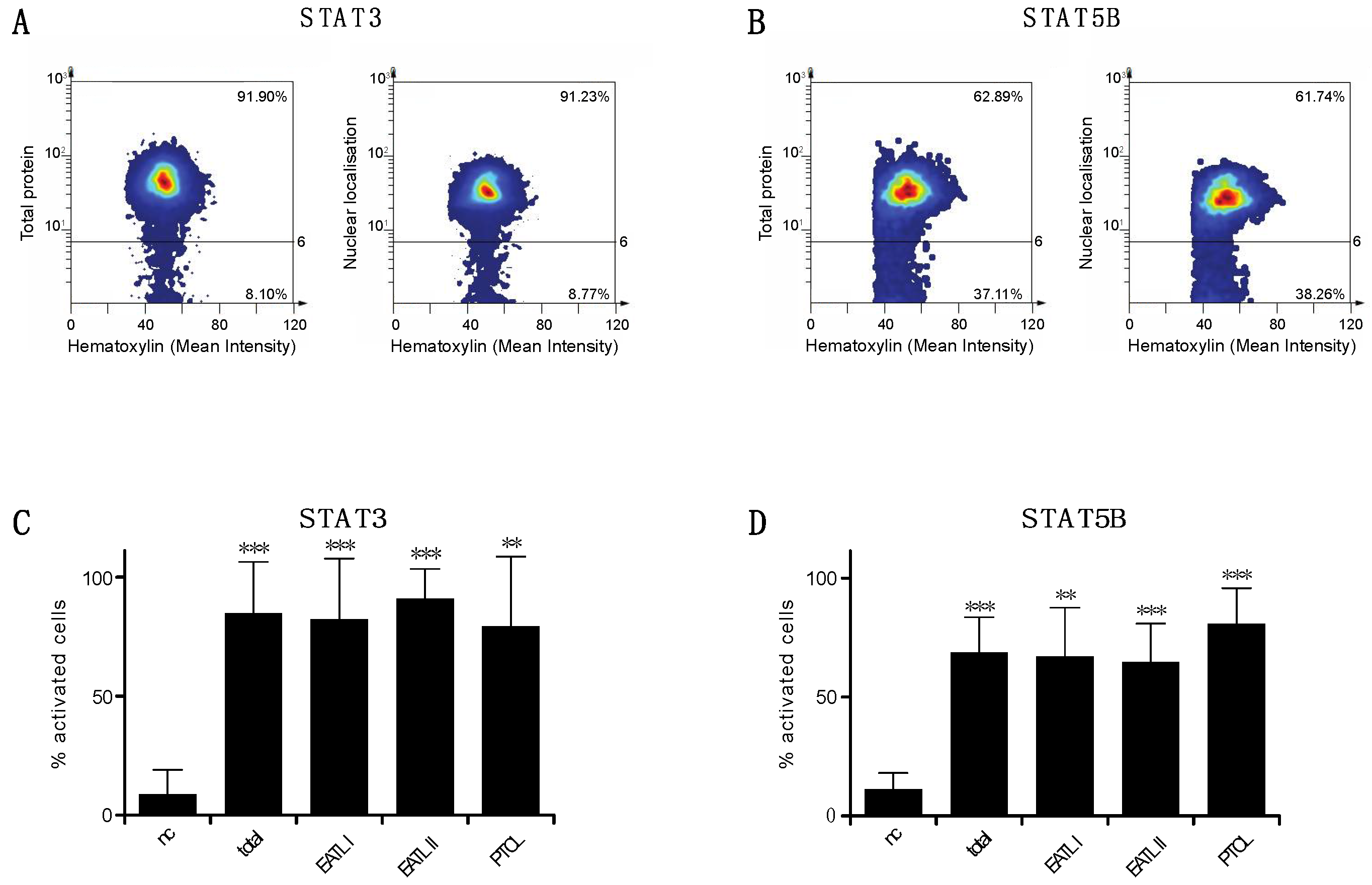
3.1. STAT3 and STAT5B Are Activated in Feline Alimentary Lymphoma
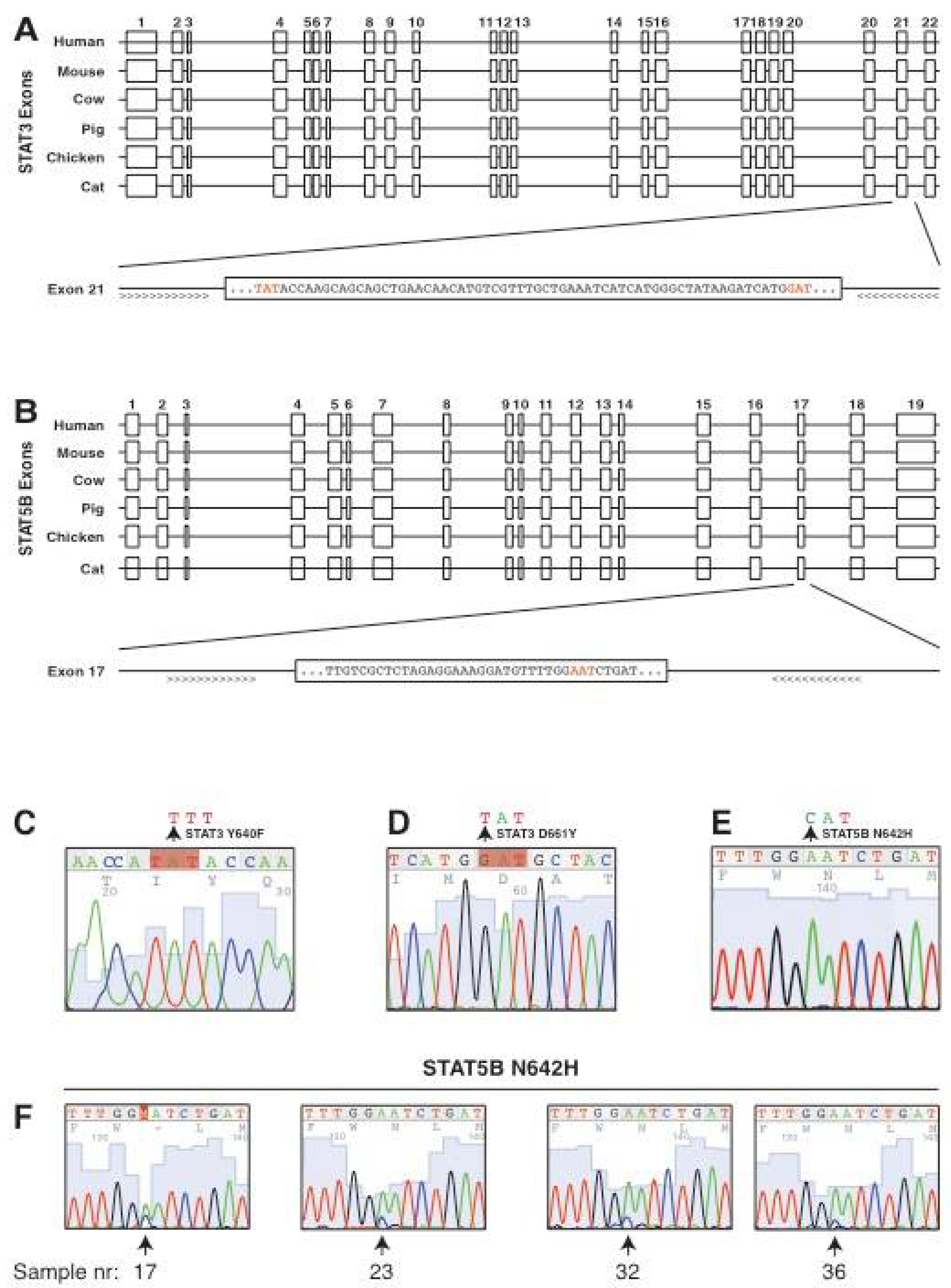
3.2. Conservation of the Genomic Locus and the Intron-Exon Structure of Feline STAT3 and STAT5B
3.3. Screening for Candidate Mutations of STAT3 and STAT5B in Feline Alimentary Lymphoma
3.4. Confirmation of STAT5B N642H Mutation in Feline Alimentary Lymphoma Cells
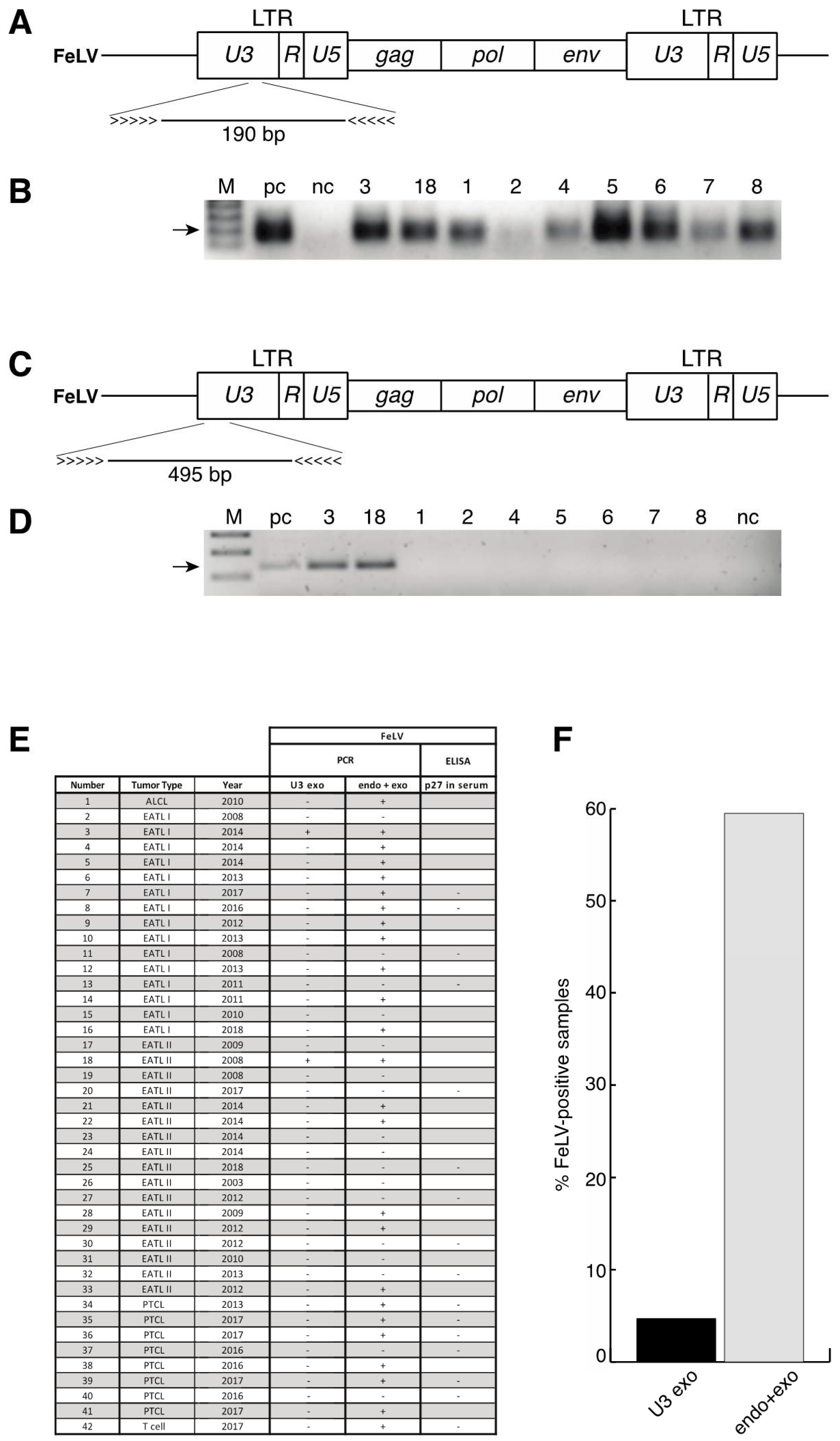
3.5. No Enrichment for Infection with Feline Leukaemia Virus (FeLV) in Patient Samples and Identification of a Novel Polymorphism of STAT5B
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Louwerens, M.; London, C.A.; Pedersen, N.C.; Lyons, L.A. Feline lymphoma in the post-feline leukemia virus era. J. Vet. Intern. Med. 2005, 19, 329–335. [Google Scholar]
- Withrow, S.J.; Vail, D.M. Withrow & MacEwen’s Small Animal Clinical Oncology, 5th ed.; Saunders Elsevier: Philadelphia, PA, USA, 2013. [Google Scholar]
- Barrs, V.R.; Beatty, J.A. Feline alimentary lymphoma: 1. Classification, risk factors, clinical signs and non-invasive diagnostics. J. Feline Med. Surg. 2012, 14, 182–190. [Google Scholar] [CrossRef]
- Valli, V.E.; Bienzle, D.; Meuten, D.J. Tumors of the Hemolymphatic System. Tumors in Domestic Animals, 5th ed.; Wiley & Sons: Hoboken, NJ, USA, 2018; pp. 203–321. [Google Scholar]
- Grover, S. Gastronintestinal lymphoma in cats. Oncol. Compend. 2005, 27, 741–751. [Google Scholar]
- Barrs, V.R.; Beatty, J.A. Feline alimentary lymphoma: 2. Further diagnostics, therapy and prognosis. J. Feline Med. Surg. 2012, 14, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Swanson, C.M.; Smedley, R.C.; Saavedra, P.V.; Kiupel, M.; Kitchell, B.E. Expression of the Bcl-2 apoptotic marker in cats diagnosed with inflammatory bowel disease and gastrointestinal lymphoma. J. Feline Med. Surg. 2012, 14, 741–745. [Google Scholar] [CrossRef]
- Foukas, P.G.; de Leval, L. Recent advances in intestinal lymphomas. Histopathology 2015, 66, 112–136. [Google Scholar] [CrossRef] [PubMed]
- Wolfesberger, B.; Fuchs-Baumgartinger, A.; Greß, V.; Hammer, S.; Gradner, G.; Knödl, K.; Tichy, A.; Rütgen, B.; Beham-Schmid, C. World Health Organisation classification of lymphoid tumours in veterinary and human medicine: A comparative evaluation of gastrointestinal lymphomas in 61 cats. J. Comp. Pathol. 2018, 159, 1–10. [Google Scholar] [CrossRef]
- Moore, P.F.; Rodriguez-Bertos, A.; Kass, P.H. Feline gastrointestinal lymphoma: Mucosal architecture, immunophenotype, and molecular clonality. Vet. Pathol. 2012, 49, 658–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulin, M.V.; Couronné, L.; Beguin, J.; Le Poder, S.; Delverdier, M.; Semin, M.-O.; Bruneau, J.; Cerf-Bensussan, N.; Malamut, G.; Cellier, C.; et al. Feline low-grade alimentary lymphoma: An emerging entity and a potential animal model for human disease. BMC Vet. Res. 2018, 14, 306. [Google Scholar] [CrossRef]
- Swerdlow, S.H.; Campo, E.; Pileri, S.A.; Harris, N.L.; Stein, H.; Siebert, R.; Advani, R.; Ghielmini, M.; Salles, G.A.; Zelenetz, A.D.; et al. The 2016 revision of the World Health Organization classification of lymphoid neoplasms. Blood 2016, 127, 2375–2390. [Google Scholar] [PubMed] [Green Version]
- Nijeboer, P.; De Baaij, L.R.; Visser, O.; Witte, B.I.; Cillessen, S.A.; Mulder, C.J.; Bouma, G. Treatment response in enteropathy associated T-cell lymphoma; survival in a large multicenter cohort. Am. J. Hematol. 2015, 90, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Vose, J.; Armitage, J.; Weisenburger, D.; International T-Cell Lymphoma Project. International peripheral T-cell and natural killer/T-cell lymphoma study: Pathology findings and clinical outcomes. J. Clin. Oncol. 2008, 26, 4124–4130. [Google Scholar]
- Margolskee, E.; Jobanputra, V.; Lewis, S.K.; Alobeid, B.; Green, P.H.R.; Bhagat, G. Indolent small intestinal CD4+ T-cell lymphoma is a distinct entity with unique biologic and clinical features. PLoS ONE 2013, 8, e68343. [Google Scholar] [CrossRef] [Green Version]
- Stark, G.R.; Darnell, J.E., Jr. The JAK-STAT pathway at twenty. Immunity 2012, 36, 503–514. [Google Scholar] [CrossRef] [Green Version]
- Wingelhofer, B.; Neubauer, H.A.; Valent, P.; Han, X.; Constantinescu, S.N.; Gunning, P.T.; Muller, M.; Morrigl, R. Implications of STAT3 and STAT5 signaling on gene regulation and chromatin remodeling in hematopoietic cancer. Leukemia 2018, 32, 1713–1726. [Google Scholar] [CrossRef] [Green Version]
- Fagard, R.; Metelev, V.; Souissi, I.; Baran-Marszak, F. STAT3 inhibitors for cancer therapy: Have all roads been explored? JAKSTAT 2013, 2, e22882. [Google Scholar] [CrossRef] [Green Version]
- Vainchenker, W.; Constantinescu, S.N. JAK/STAT signaling in hematological malignancies. Oncogene 2012, 32, 2601–2613. [Google Scholar] [CrossRef] [Green Version]
- O’Shea, J.J.; Schwartz, D.M.; Villarino, A.V.; Gadina, M.; McInnes, I.B.; Laurence, A. The JAK-STAT pathway: Impact on human disease and therapeutic intervention. Annu. Rev. Med. 2015, 66, 311–328. [Google Scholar] [CrossRef] [Green Version]
- Owen, K.L.; Brockwell, N.K.; Parker, B.S. JAK-STAT Signaling: A Double-Edged Sword of Immune Regulation and Cancer Progression. Cancers 2019, 11, 2002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orlova, A.; Wingelhofer, B.; Neubauer, H.A.; Maurer, B.; Berger-Becvar, A.; Keserű, G.M.; Gunning, P.T.; Valent, P.; Moriggl, R. Emerging therapeutic targets in myeloproliferative neoplasms and peripheral T-cell leukemia and lymphomas. Expert Opin. Ther. Targets 2018, 22, 45–57. [Google Scholar] [CrossRef] [Green Version]
- Deleeuw, R.J.; Zettl, A.; Klinker, E.; Haralambieva, E.; Trottier, M.; Chari, R.; Ge, Y.; Gascoyne, R.D.; Chott, A.; Müller–Hermelink, H.; et al. Whole-genome analysis and HLA genotyping of enteropathy-type T-cell lymphoma reveals 2 distinct lymphoma subtypes. Gastroenterology 2007, 132, 1902–1911. [Google Scholar] [CrossRef] [PubMed]
- Tomita, S.; Kikuti, Y.Y.; Carreras, J.; Kojima, M.; Ando, K.; Takasaki, H.; Sakai, R.; Takata, K.; Yoshino, T.; Bea, S.; et al. Genomic and immunohistochemical profiles of enteropathy-associated T-cell lymphoma in Japan. Mod. Pathol. 2015, 28, 1286–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberti, A.; Dobay, M.P.; Bisig, B.; Vallois, D.; Boéchat, C.; Lanitis, E.; Bouchindhomme, B.; Parrens, M.-C.; Bossard, C.; Quintanilla-Martinez, L.; et al. Type II enteropathy-associated T-cell lymphoma features a unique genomic profile with highly recurrent SETD2 alterations. Nat. Commun. 2016, 7, 12602. [Google Scholar] [CrossRef]
- Nairismägi, M.-L.; Tan, J.; Lim, J.Q.; Nagarajan, S.; Ng, C.C.Y.; Rajasegaran, V.; Huang, D.; Lim, W.K.; Laurensia, Y.; Wijaya, G.C.; et al. JAK-STAT and G-protein-coupled receptor signaling pathways are frequently altered in epitheliotropic intestinal T-cell lymphoma. Leukemia 2016, 30, 1311–1319. [Google Scholar] [CrossRef] [Green Version]
- Kucuk, C.; Jiang, B.; Hu, X.; Zhang, W.; Chan, J.K.; Xiao, W.; Lack, N.; Alkan, C.; Williams, J.C.; Avery, K.N.; et al. Activating mutations of STAT5B and STAT3 in lymphomas derived from gammadelta-T or NK cells. Nat. Commun. 2015, 6, 6025. [Google Scholar] [CrossRef] [Green Version]
- Nicolae, A.; Xi, L.; Pham, T.H.; Pham, T.-A.; Navarro, W.; Meeker, H.G.; Pittaluga, S.; Jaffe, E.S.; Raffeld, M. Mutations in the JAK/STAT and RAS signaling pathways are common in intestinal T-cell lymphomas. Leukemia 2016, 30, 2245–2247. [Google Scholar] [CrossRef] [Green Version]
- Moffitt, A.B.; Ondrejka, S.L.; McKinney, M.; Rempel, R.E.; Goodlad, J.R.; Teh, C.H.; Leppa, S.; Mannisto, S.; Kovanen, P.E.; Tse, E.; et al. Enteropathy-associated T cell lymphoma subtypes are characterized by loss of function of SETD2. J. Exp. Med. 2017, 214, 1371–1386. [Google Scholar] [CrossRef]
- Kadekar, D.; Agerholm, R.; Rizk, J.; Neubauer, H.A.; Suske, T.; Maurer, B.; Vinals, M.T.; Comelli, E.M.; Taibi, A.; Moriggl, L.; et al. The neonatal microenvironment programs innate gammadelta T cells through the transcription factor STAT5. J. Clin. Investig. 2020, 130, 2496–2508. [Google Scholar] [CrossRef]
- Valli, V.E.; Kiupel, M.; Bienzle, D. Pathology of Domestic Animals, 6th ed.; Academic Press: Cambridge, MA, USA, 2016; Volume 3, pp. 102–268. [Google Scholar]
- Meuten, D.J.; Moore, F.M. Tumors in Domestic Animals; Wiley Blackwell: Hoboken, NJ, USA, 2018; p. 944. [Google Scholar]
- Tandon, R.; Cattori, V.; Gomes-Keller, M.A.; Meli, M.L.; Golder, M.C.; Lutz, H.; Hofmann-Lehmann, R. Quantitation of feline leukaemia virus viral and proviral loads by TaqMan real-time polymerase chain reaction. J. Virol. Methods 2005, 130, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Little, S. Amplification-refractory mutation system (ARMS) analysis of point mutations. Curr. Protoc. Hum. Genet. 1995, 7, 9.8.1–9.8.12. [Google Scholar] [CrossRef]
- Burgstaller, J.P.; Schinogl, P.; Dinnyes, A.; Muller, M.; Steinborn, R. Mitochondrial DNA heteroplasmy in ovine fetuses and sheep cloned by somatic cell nuclear transfer. BMC Dev. Biol. 2007, 7, 141. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Desmet, F.O.; Hamroun, D.; Lalande, M.; Collod-Beroud, G.; Claustres, M.; Beroud, C. Human Splicing Finder: An online bioinformatics tool to predict splicing signals. Nucleic Acids Res. 2009, 37, e67. [Google Scholar] [CrossRef] [Green Version]
- Gordziel, C.; Bratsch, J.; Moriggl, R.; Knosel, T.; Friedrich, K. Both STAT1 and STAT3 are favourable prognostic determinants in colorectal carcinoma. Br. J. Cancer 2013, 109, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Maurer, B.; Nivarthi, H.; Wingelhofer, B.; Pham, H.T.T.; Schlederer, M.; Suske, T.; Grausenburger, R.; Schiefer, A.-I.; Prchal-Murphy, M.; Chen, D.; et al. High activation of STAT5A drives peripheral T-cell lymphoma and leukemia. Haematologica 2018, 105, 435–447. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.; Carritt, K.; Lane, J.; Jayappa, H.; Stahl, M.; Bourgeois, M. Comparative efficacy of feline leukemia virus (FeLV) inactivated whole-virus vaccine and canarypox virus-vectored vaccine during virulent FeLV challenge and immunosuppression. Clin. Vaccine Immunol. 2015, 22, 798–805. [Google Scholar] [CrossRef] [Green Version]
- Verbeek, W.H.; Van De Water, J.M.; Al-Toma, A.; Oudejans, J.J.; Mulder, C.J.; Coupe, V.M. Incidence of enteropathy-associated T-cell lymphoma: A nation-wide study of a population-based registry in The Netherlands. Scand. J. Gastroenterol. 2008, 43, 1322–1328. [Google Scholar] [CrossRef] [PubMed]
- Tse, E.; Gill, H.; Loong, F.; Kim, S.J.; Ng, S.B.; Tang, T.; Ko, Y.-H.; Chng, W.-J.; Lim, S.-T.; Kim, W.S. Type II enteropathy-associated T-cell lymphoma: A multicenter analysis from the Asia Lymphoma Study Group. Am. J. Hematol. 2012, 87, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Gagne, J.J.; Thompson, L.; O’Keefe, K.; Kesselheim, A.S. Innovative research methods for studying treatments for rare diseases: Methodological review. BMJ 2014, 349, g6802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pogue, R.; Cavalcanti, D.P.; Shanker, S.; Andrade, R.V.; Aguiar, L.R.; Carvalho, J.; Costa, F.F. Rare genetic diseases: Update on diagnosis, treatment and online resources. Drug Discov. Today 2018, 23, 187–195. [Google Scholar] [CrossRef]
- Wilson, H.M. Feline alimentary lymphoma: Demystifying the enigma. Top. Companion Anim. Med. 2008, 23, 177–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rissetto, K.; Villamil, J.A.; Selting, K.A.; Tyler, J.; Henry, C.J. Recent trends in feline intestinal neoplasia: An epidemiologic study of 1129 cases in the veterinary medical database from 1964 to 2004. J. Am. Anim. Hosp. Assoc. 2011, 47, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Brachet-Botineau, M.; Polomski, M.; Neubauer, H.A.; Juen, L.; Hédou, D.; Viaud-Massuard, M.C.; Prie, G.; Gouilleux, F. Pharmacological inhibition of oncogenic STAT3 and STAT5 signaling in hematopoietic cancers. Cancers 2020, 12, 240. [Google Scholar] [CrossRef] [Green Version]
- De Araujo, E.D.; Orlova, A.; Neubauer, H.A.; Bajusz, D.; Seo, H.-S.; Dhe-Paganon, S.; Keserű, G.M.; Moriggl, R.; Gunning, P.T. Structural implications of STAT3 and STAT5 SH2 domain mutations. Cancers 2019, 11, 1757. [Google Scholar] [CrossRef] [Green Version]
- Hohdatsu, T.; Hirabayashi, H.; Motokawa, K.; Koyama, H. Comparative study of the cell tropism of feline immunodeficiency virus isolates of subtypes A, B and D classified on the basis of the env gene V3-V5 sequence. J. Gen. Virol. 1996, 77, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, T.; Toyosaki, T.; Tomonaga, K.; Norimine, J.; Ohno, K.; Hasegawa, A.; Kai, C.; Mikami, T. Further characterization of a feline T-lymphoblastoid cell line (MYA-1 cells) highly sensitive for feline immunodeficiency virus. J. Vet. Med. Sci. 1992, 54, 173–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orlova, A.; Wagner, C.; De Araujo, E.D.; Bajusz, D.; Neubauer, H.A.; Herling, M.; Gunning, P.T.; Keserű, G.M.; Moriggl, R. Direct targeting options for STAT3 and STAT5 in cancer. Cancers 2019, 11, 1930. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Oishi, N.; Boddicker, R.L.; Hu, G.; Benson, H.K.; Ketterling, R.P.; Greipp, P.T.; Knutson, D.L.; Kloft-Nelson, S.M.; He, R.; et al. Recurrent STAT3-JAK2 fusions in indolent T-cell lymphoproliferative disorder of the gastrointestinal tract. Blood 2018, 131, 2262–2266. [Google Scholar] [CrossRef] [Green Version]
- Soderquist, C.R.; Patel, N.; Murty, V.V.; Betman, S.; Aggarwal, N.; Young, K.H.; Xerri, L.; Leeman-Neill, R.; Lewis, S.K.; Green, P.H.; et al. Genetic and phenotypic characterization of indolent T-cell lymphoproliferative disorders of the gastrointestinal tract. Haematologica 2020, 105, 1895–1906. [Google Scholar] [CrossRef] [Green Version]
- Wingelhofer, B.; Maurer, B.; Heyes, E.C.; Cumaraswamy, A.A.; Berger-Becvar, A.; De Araujo, E.D.; Orlova, A.; Freund, P.; Ruge, F.; Park, J.; et al. Pharmacologic inhibition of STAT5 in acute myeloid leukemia. Leukemia 2018, 32, 1135–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, B.D.G.; Khoury, H.; Laister, R.C.; Fletcher, S.; Vellozo, M.; Manzoli, A.; Yue, P.; Turkson, J.; Minden, M.D.; Gunning, P.T. Small molecule STAT5-SH2 domain inhibitors exhibit potent antileukemia activity. J. Med. Chem. 2012, 55, 1047–1055. [Google Scholar] [CrossRef] [PubMed]
- Pham, H.T.T.; Maurer, B.; Prchal-Murphy, M.; Grausenburger, R.; Grundschober, E.; Javaheri, T.; Nivarthi, H.; Boersma, A.; Kolbe, T.; Elabd, M.; et al. STAT5BN642H is a driver mutation for T cell neoplasia. J. Clin. Investig. 2017, 128, 387–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shelton, G.H.; Grant, C.K.; Cotter, S.M.; Gardner, M.B.; Hardy, W.D.; DiGiacomo, R.F., Jr. Feline immunodeficiency virus and feline leukemia virus infections and their relationships to lymphoid malignancies in cats: A retrospective study (1968–1988). J. Acquir. Immune Defic. Syndr. 1990, 3, 623–630. [Google Scholar] [PubMed]
- Stutzer, B.; Simon, K.; Lutz, H.; Majzoub, M.; Hermanns, W.; Hirschberger, J.; Sauter-Louis, C.; Hartmann, K. Incidence of persistent viraemia and latent feline leukaemia virus infection in cats with lymphoma. J. Feline Med. Surg. 2011, 13, 81–87. [Google Scholar] [CrossRef]
- Weiss, A.T.; Klopfleisch, R.; Gruber, A.D. Prevalence of feline leukaemia provirus DNA in feline lymphomas. J. Feline Med. Surg. 2010, 12, 929–935. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kieslinger, M.; Swoboda, A.; Kramer, N.; Freund, P.; Pratscher, B.; Neubauer, H.A.; Steinborn, R.; Wolfesberger, B.; Fuchs-Baumgartinger, A.; Moriggl, R.; et al. A Recurrent STAT5BN642H Driver Mutation in Feline Alimentary T Cell Lymphoma. Cancers 2021, 13, 5238. https://doi.org/10.3390/cancers13205238
Kieslinger M, Swoboda A, Kramer N, Freund P, Pratscher B, Neubauer HA, Steinborn R, Wolfesberger B, Fuchs-Baumgartinger A, Moriggl R, et al. A Recurrent STAT5BN642H Driver Mutation in Feline Alimentary T Cell Lymphoma. Cancers. 2021; 13(20):5238. https://doi.org/10.3390/cancers13205238
Chicago/Turabian StyleKieslinger, Matthias, Alexander Swoboda, Nina Kramer, Patricia Freund, Barbara Pratscher, Heidi A. Neubauer, Ralf Steinborn, Birgitt Wolfesberger, Andrea Fuchs-Baumgartinger, Richard Moriggl, and et al. 2021. "A Recurrent STAT5BN642H Driver Mutation in Feline Alimentary T Cell Lymphoma" Cancers 13, no. 20: 5238. https://doi.org/10.3390/cancers13205238
APA StyleKieslinger, M., Swoboda, A., Kramer, N., Freund, P., Pratscher, B., Neubauer, H. A., Steinborn, R., Wolfesberger, B., Fuchs-Baumgartinger, A., Moriggl, R., & Burgener, I. A. (2021). A Recurrent STAT5BN642H Driver Mutation in Feline Alimentary T Cell Lymphoma. Cancers, 13(20), 5238. https://doi.org/10.3390/cancers13205238