
Methylation of DROSHA and DICER as a Biomarker for the Detection of Lung Cancer

 , , , , and
, , , , and 
Abstract
:Simple Summary
Abstract
1. Introduction
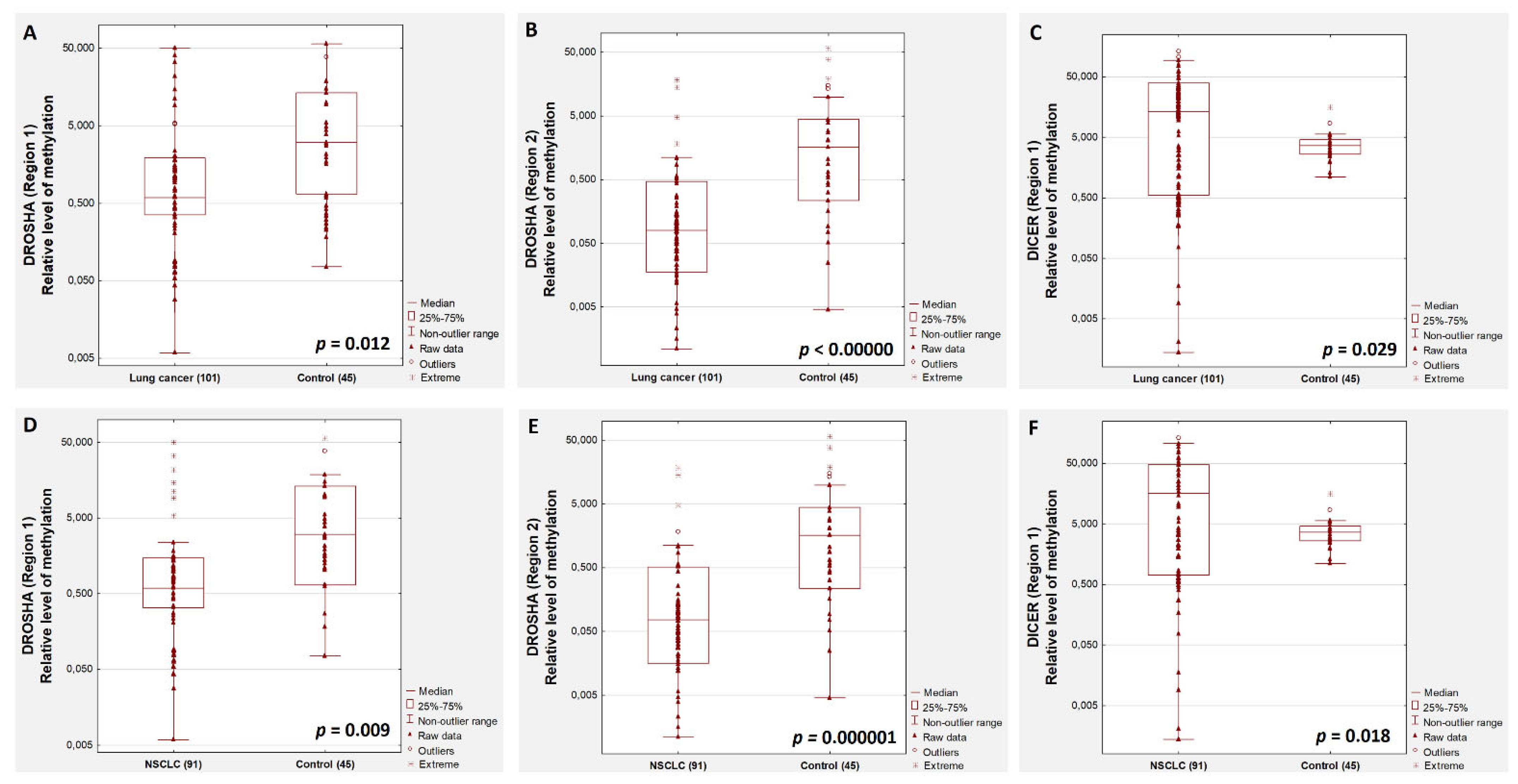
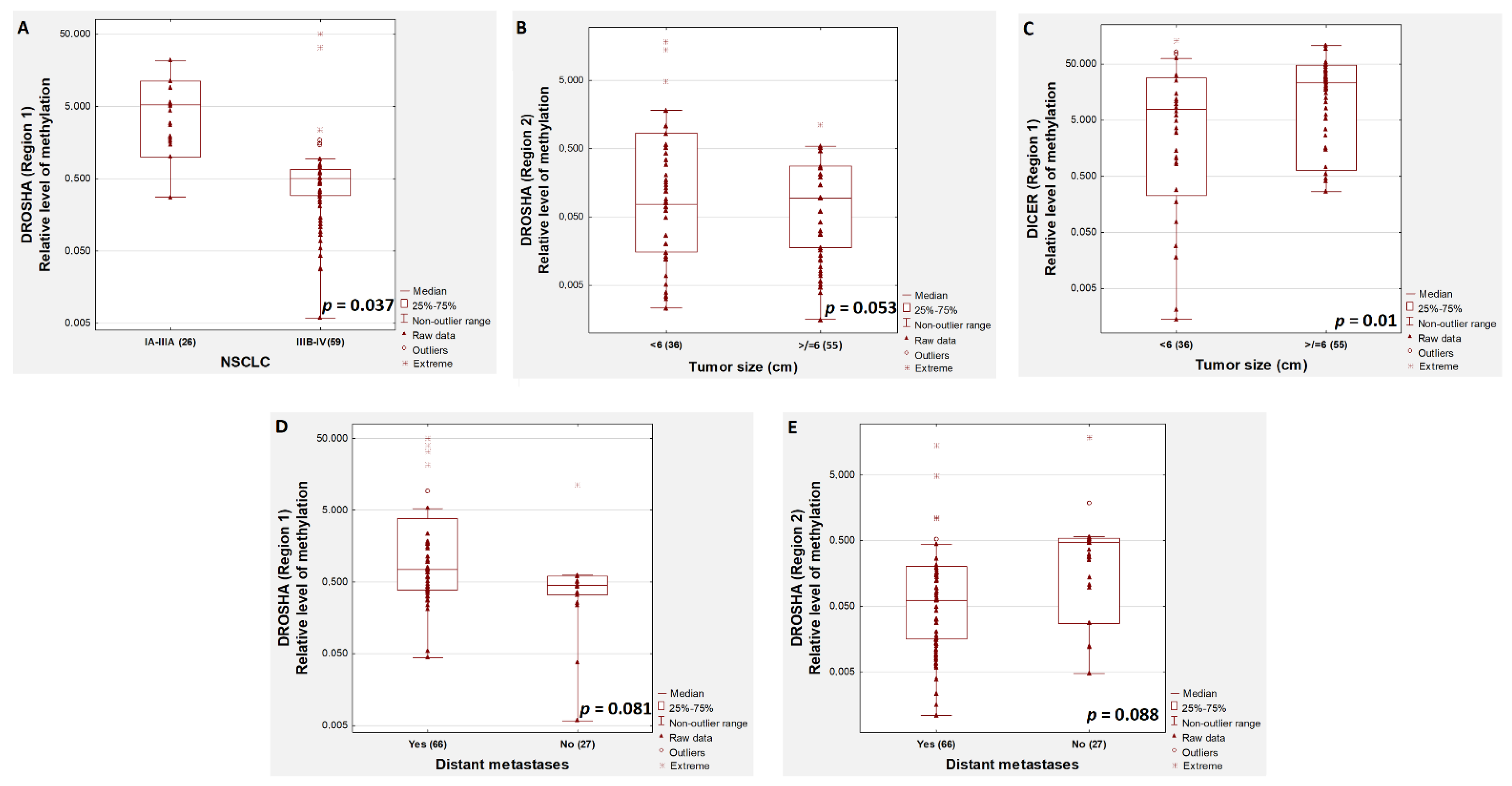
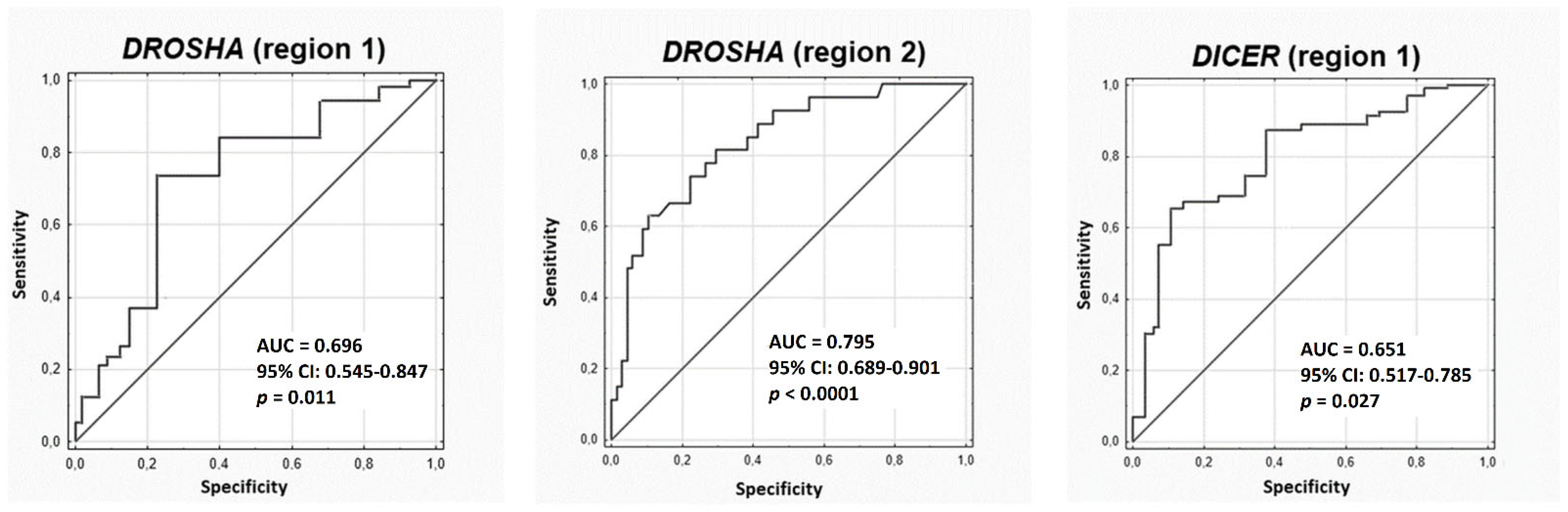
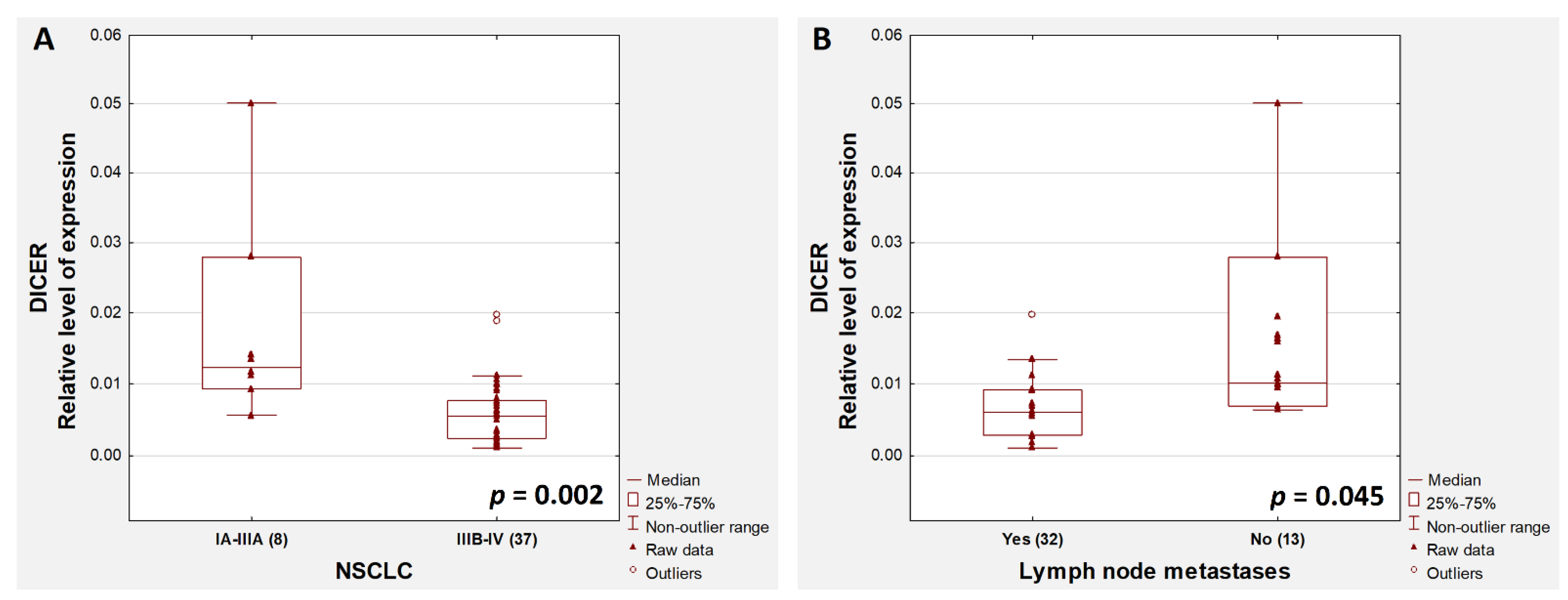
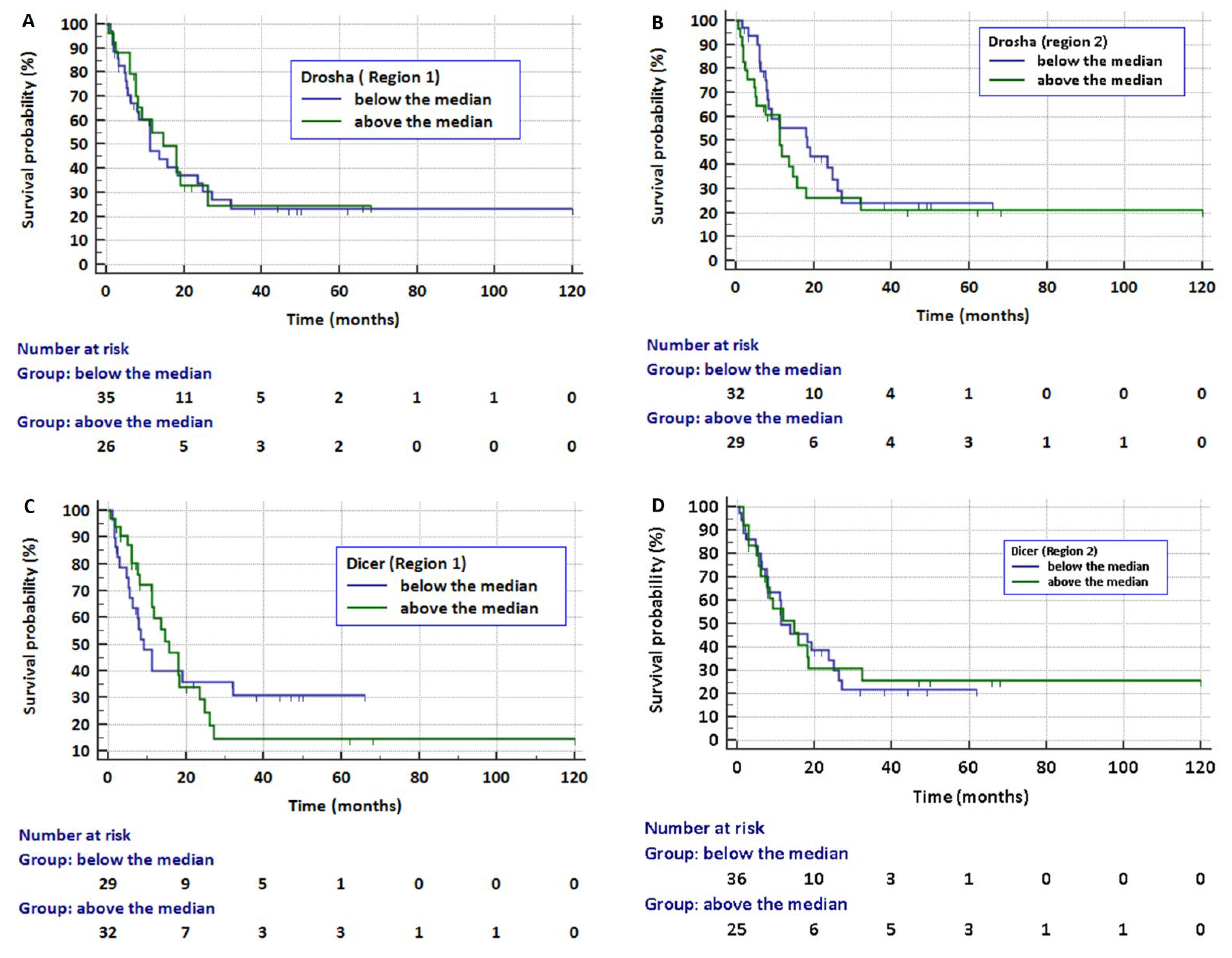
2. Results
3. Discussion
4. Materials and Methods
4.1. Study Group
4.2. Isolation of Circulating Free DNA from Plasma Samples
4.3. Bisulfitation of DNA and Real-Time PCR
4.4. Expression of DROSHA and DICER
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Wong, M.C.S.; Lao, X.Q.; Ho, K.F.; Goggins, W.B.; Tse, S.L.A. Incidence and mortality of lung cancer: Global trends and association with socioeconomic status. Sci. Rep. 2017, 7, 14300. [Google Scholar] [CrossRef] [Green Version]
- Lu, T.; Yang, X.; Huang, Y.; Zhao, M.; Li, M.; Ma, K.; Yin, J.; Zhan, C.; Wang, Q. Trends in the incidence, treatment, and survival of patients with lung cancer in the last four decades. Cancer Manag. Res. 2019, 11, 943–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melamed, M.R.; Flehinger, B.J. Screening for lung cancer. Chest 1984, 86, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Marcus, P.M.; Bergstralh, E.J.; Zweig, M.H.; Harris, A.; Offord, K.P.; Fontana, R.S. Extended lung cancer incidence follow-up in the Mayo Lung Project and overdiagnosis. J. Natl. Cancer Inst. 2006, 98, 748–756. [Google Scholar] [CrossRef]
- National Lung Screening Trial Research Team; Aberle, D.R.; Adams, A.M.; Berg, C.D.; Black, W.C.; Clapp, J.D.; Fagerstrom, R.M.; Gareen, I.F.; Gatsonis, C.; Marcus, P.M.; et al. Reduced lung-cancer mortality with low-dose computed tomographic screening. N. Engl. J. Med. 2011, 365, 395–409. [Google Scholar] [PubMed] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Slack, F.J.; Weidhaas, J.B. MicroRNA in cancer prognosis. N. Engl. J. Med. 2008, 359, 2720–2722. [Google Scholar] [CrossRef] [PubMed]
- Iorio, M.V.; Croce, C.M. microRNA involvement in human cancer. Carcinogenesis 2012, 33, 1126–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filipowicz, W.; Bhattacharyya, S.N.; Sonenberg, N. Mechanisms of post-transcriptional regulation by microRNAs: Are the answers in sight? Nat. Rev. Genet. 2008, 9, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Cedar, H.; Bergman, Y. Programming of DNA methylation patterns. Annu. Rev. Biochem. 2012, 81, 97–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aran, D.; Sabato, S.; Hellman, A. DNA methylation of distal regulatory sites characterizes dysregulation of cancer genes. Genome Biol. 2013, 14, R21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, R.E.; Golan, T.; Sheinboim, D.; Malcov, H.; Amar, D.; Salamon, A.; Liron, T.; Gelfman, S.; Gabet, Y.; Shamir, R.; et al. Enhancer methylation dynamics contribute to cancer plasticity and patient mortality. Genome Res. 2016, 26, 601–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esteller, M. Epigenetic gene silencing in cancer: The DNA hypermethylome. Hum. Mol. Genet. 2007, 16, R50–R59. [Google Scholar] [CrossRef] [PubMed]
- Hollander, D.; Naftelberg, S.; Lev-Maor, G.; Kornblihtt, A.R.; Ast, G. How are short exons flanked by long introns defined and committed to splicing? Trends Genet. 2016, 32, 596–606. [Google Scholar] [CrossRef]
- Maunakea, A.K.; Chepelev, I.; Cui, K.R.; Zhao, K.J.; Intragenic, D.N.A. Methylation modulates alternative splicing by recruiting MeCP2 to promote exon recognition. Cell Res. 2013, 23, 1256–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelfman, S.; Cohen, N.; Yearim, A.; Ast, G. DNA-methylation effect on cotranscriptional splicing is dependent on GC architecture of the exon-intron structure. Genome Res. 2013, 23, 789–799. [Google Scholar] [CrossRef] [Green Version]
- Young, J.I.; Hong, E.P.; Castle, J.C.; Crespo-Barreto, J.; Bowman, A.B.; Rose, M.F.; Kang, D.; Richman, R.; Johnson, J.M.; Berget, S.; et al. Regulation of RNA splicing by the methylation-dependent transcriptional repressor methyl-CpG binding protein 2. Proc. Natl. Acad. Sci. USA 2005, 102, 17551–17558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullany, L.E.; Herrick, J.S.; Wolff, R.K.; Slattery, M.L. Single nucleotide polymorphisms within MicroRNAs, MicroRNA targets, and MicroRNA biogenesis genes and their impact on colorectal cancer survival. Gen. Chromosomes Cancer. 2017, 56, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Bell, R.E.; Levy, C. The three M’s: Melanoma, microphthalmia-associated transcription factor and microRNA. Pigment. Cell Melanoma Res. 2011, 24, 1088–1106. [Google Scholar] [CrossRef] [PubMed]
- Voorhoeve, P.M.; Agami, R. Classifying microRNAs in cancer: The good, the bad and the ugly. Biochim. Biophys. Acta 2007, 1775, 274–282. [Google Scholar] [CrossRef]
- Karube, Y.; Tanaka, H.; Osada, H.; Tomida, S.; Tatematsu, Y.; Yanagisawa, K.; Yatabe, Y.; Takamizawa, J.; Miyoshi, S.; Mitsudomi, T.; et al. Reduced expression of Dicer associated with poor prognosis in lung cancer patients. Cancer Sci. 2005, 96, 111–115. [Google Scholar] [CrossRef]
- Chiosea, S.; Jelezcova, E.; Chandran, U.; Luo, J.; Mantha, G.; Sobol, R.W.; Dacic, S. Overexpression of Dicer in precursor lesions of lung adenocarcinoma. Cancer Res. 2007, 67, 2345–2350. [Google Scholar] [CrossRef] [Green Version]
- Prodromaki, E.; Korpetinou, A.; Giannopoulou, E.; Vlotinou, E.; Chatziathanasiadou, Μ.; Papachristou, N.I.; Scopa, C.D.; Papadaki, H.; Kalofonos, H.P.; Papachristou, D.J. Expression of the microRNA regulators Drosha, Dicer and Ago2 in non-small cell lung carcinomas. Cell. Oncol. (Dordr.) 2015, 38, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Chiosea, S.; Jelezcova, E.; Chandran, U.; Acquafondata, M.; McHale, T.; Sobol, R.W.; Dhir, R. Up-regulation of dicer, a component of the MicroRNA machinery, in prostate adenocarcinoma. Am. J. Pathol. 2006, 169, 1812–1820. [Google Scholar] [CrossRef] [Green Version]
- Flavin, R.J.; Smyth, P.C.; Finn, S.P.; Laios, A.; O'Toole, S.A.; Barrett, C.; Ring, M.; Denning, K.M.; Li, J.; Aherne, S.T.; et al. Altered eIF6 and Dicer expression is associated with clinicopathological features in ovarian serous carcinoma patients. Mod. Pathol. 2008, 21, 676–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faber, C.; Horst, D.; Hlubek, F.; Kirchner, T. Overexpression of Dicer predicts poor survival in colorectal cancer. Eur. J. Cancer 2011, 47, 1414–1419. [Google Scholar] [CrossRef] [PubMed]
- Papachristou, D.J.; Rao, U.N.; Korpetinou, A.; Giannopoulou, E.; Sklirou, E.; Kontogeorgakos, V.; Kalofonos, H.P. Prognostic significance of Dicer cellular levels in soft tissue sarcomas. Cancer Invest. 2012, 30, 172–179. [Google Scholar] [CrossRef]
- Potenza, N.; Papa, U.; Russo, A. Differential expression of Dicer and Argonaute genes during the differentiation of human neuroblastoma cells. Cell Biol. Int. 2009, 33, 734–738. [Google Scholar] [CrossRef]
- Sugito, N.; Ishiguro, H.; Kuwabara, Y.; Kimura, M.; Mitsui, A.; Kurehara, H.; Ando, T.; Mori, R.; Takashima, N.; Ogawa, R.; et al. RNASEN regulates cell proliferation and affects survival in esophageal cancer patients. Clin. Cancer Res. 2006, 12, 7322328. [Google Scholar] [CrossRef] [Green Version]
- Grelier, G.; Voirin, N.; Ay, A.S.; Cox, D.G.; Chabaud, S.; Treilleux, I.; Léon-Goddard, S.; Rimokh, R.; Mikaelian, I.; Venoux, C.; et al. Prognostic value of Dicer in human breast cancers and association with the mesenchymal phenotype. Br. J. Cancer 2009, 101, 673–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, G.S.; Yang, Z.L.; Liu, D.C. Immunohistochemical study of Dicer and Drosha expression in the benign and malignant lesions of gallbladder and their clinicopathological significances. Pathol. Res. Pr. 2012, 208, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Díaz-García, C.V.; Agudo-López, A.; Pérez, C.; López-Martín, J.A.; Rodríguez-Peralto, J.L.; de Castro, J.; Cortijo, A.; Martínez-Villanueva, M.; Iglesias, L.; García-Carbonero, R.; et al. DICER1, DROSHA and miRNAs in patients with non-small cell lung cancer: Implications for outcomes and histologic classification. Carcinogenesis 2013, 34, 1031–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | n (%) | DROSHA1 | DROSHA2 | DICER1 | DICER2 | ||||
|---|---|---|---|---|---|---|---|---|---|
| Below the Median | Above the Median | Below the Median | Above the Median | Below the Median | Above the Median | Below the Median | Above the Median | ||
| Age | |||||||||
| <65 | 46 (45.5) | 21 (45.7) | 25 (54.3) | 22 (47.8) | 24 (52.2) | 19 (41.3) | 27 (58.7) | 21 (45.6) | 25 (54.4) |
| ≥65 | 55 (54.5) | 27 (49.9) | 28 (50.1) | 28 (50.9) | 27 (49.1) | 29 (52.7) | 26 (47.3) | 30 (54.5) | 25 (45.5) |
| p-value | 0.730 | 0.758 | 0.252 | 0.433 | |||||
| χ2 | 0.119 | 0.095 | 1.311 | 0.615 | |||||
| Gender | |||||||||
| Male | 63 (62.4) | 34 (54.0) | 29 (46.0) | 32 (50.8) | 31 (49.2) | 28 (44.4) | 35 (55.56) | 27 (42.9) | 36 (57.1) |
| Female | 38 (37.6) | 14 (36.8) | 24 (63.2) | 18 (47.4) | 20 (52.6) | 20 (52.6) | 18 (47.4) | 24 (63.2) | 14 (36.8) |
| p-value | 0.095 | 0.680 | 0.425 | 0.048 | |||||
| χ2 | 2.788 | 0.170 | 0.637 | 3.908 | |||||
| Histopathology | |||||||||
| Adenocarcinoma (AC) | 52 (51.5) | 21 (40.4) | 31 (59.6) | 29 (55.8) | 23 (44.2) | 26 (50.0) | 26 (50.0) | 26 (50.0) | 26 (50.0) |
| Squamous cell carcinoma (SqSC) | 34 (33.7) | 20 (58.8) | 14 (41.2) | 16 (47.1) | 18 (52.9) | 14 (41.2) | 20 (58.8) | 19 (55.9) | 15 (44.1) |
| NSCLC NOS | 5 (4.9) | 3 (60) | 2 (40) | 3 (60) | 2 (40) | 2 (40) | 3 (60) | 3 (60) | 2 (40) |
| SCLC | 8 (7.9) | 3 (37.5) | 5 (62.5) | 1 (12.5) | 7 (87.5) | 6 (75) | 2 (25) | 3 (37.5) | 5 (62.5) |
| LCC | 2 (2.0) | 1 (50.0) | 1 (50.0) | 1 (50.0) | 1 (50.0) | 0 (0) | 2 (100) | 0 (0) | 2 (100) |
| p-value | 0.487 | 0.240 | 0.285 | 0.531 | |||||
| χ2 | 3.443 | 5.501 | 5.024 | 3.161 | |||||
| Tumour size | |||||||||
| <60 mm | 36 (39.6) | 22 (61.1) | 14 (38.9) | 16 (61.5) | 16 (44.4) | 20 (55.6) | 16 (44.4) | 16 (44.4) | 20 (55.6) |
| ≥60 mm | 55 (60.4) | 24 (43.6) | 31 (56.4) | 28 (50.9) | 27 (49.1) | 17 (30.9) | 38 (69.1) | 32 (58.2) | 23 (41.8) |
| p-value | 0.103 | 0.933 | 0.019 | 0.199 | |||||
| χ2 | 2.658 | 0.007 | 5.478 | 1.647 | |||||
| Stage | |||||||||
| IA-IIIA | 27 (29.0) | 4 (14.8) | 23 (85.2) | 18 (66.7) | 9 (33.3) | 19 (70.4) | 8 (29.6) | 10 (37.0) | 17 (63.0) |
| IIIB-IV | 66 (71.0) | 38 (57.6) | 28 (42.4) | 26 (39.4) | 40 (60.6) | 24 (36.4) | 42 (63.6) | 39 (59.1) | 27 (40.9) |
| p-value | 0.00005 | 0.017 | 0.003 | 0.053 | |||||
| χ2 | 16.405 | 5.717 | 8.914 | 3.739 | |||||
| Lymph nodes metastases | |||||||||
| No | 17 (31.5) | 6 (35.3) | 11 (64.7) | 11 (64.7) | 6 (35.3) | 12 (70.6) | 5 (29.4) | 4 (23.5) | 13 (76.5) |
| Yes | 37 (68.5) | 11 (29.7) | 26 (70.3) | 18 (48.6) | 19 (51.4) | 24 (64.9) | 13 (35.1) | 17 (45.9) | 20 (54.1) |
| p-value | 0.683 | 0.359 | 0.678 | 0.117 | |||||
| χ2 | 0.167 | 0.841 | 0.172 | 2.463 | |||||
| Distant metastases | |||||||||
| No | 27 (29.0) | 19 (70.4) | 8 (29.6) | 8 (29.6) | 19 (70.4) | 17 (63.0) | 10 (37) | 12 (44.4) | 15 (55.6) |
| Yes | 66 (71.0) | 25 (37.9) | 41 (62.1) | 37 (56.1) | 29 (43.9) | 29 (43.9) | 37 (56.1) | 36 (54.5) | 30 (45.5) |
| p-value | 0.004 | 0.021 | 0.096 | 0.376 | |||||
| χ2 | 8.115 | 5.360 | 2.774 | 0.783 | |||||
| DICER | DROSHA | |
|---|---|---|
| First region | MF1: | MF1: |
| AAGGTTTAGTTTAGGCGTTGGGTCGTAAAC | AGAAGATTCGGGAAAGTCGGCGTTT | |
| MR1: | MR1: | |
| GCTTAAAAAATCCCACTAACTCCCGCA | CCACCGCAAAACCTTATACGCGATAA | |
| UF1: | UF1: | |
| AGGTTTAGTTTAGGTGTTGGGTTGTAAATGT | AGTTGGAGAAGATTTGGGAAAGTTGGTGTTT | |
| UR1: | UR1: | |
| ACCACTTAAAAAATCCCACTAACTCCCACA | CAACCCACCACAAAACCTTATACACAATAA | |
| Second region | MF5: | MF2: |
| GGGTTAATTAGAGTGTTTGGGATTTTAATTC | AAGTGGAGTAGTTTTAAGGAATGGTC | |
| MR5: | MR2: | |
| AAATTTACTATCAATAAATTTTTTCCACCCG | AAAATCTAACTCCATATCCTCCTCG | |
| UF5: | UF2: | |
| GGGTTAATTAGAGTGTTTGGGATTTTAATTT | AGTGGAGTAGTTTTAAGGAATGGTT | |
| UR5: | UR2: | |
| AATTTACTATCAATAAATTTTTTCCACCCA | AAAATCTAACTCCATATCCTCCTCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szczyrek, M.; Grenda, A.; Kuźnar-Kamińska, B.; Krawczyk, P.; Sawicki, M.; Batura-Gabryel, H.; Mlak, R.; Szudy-Szczyrek, A.; Krajka, T.; Krajka, A.; et al. Methylation of DROSHA and DICER as a Biomarker for the Detection of Lung Cancer. Cancers 2021, 13, 6139. https://doi.org/10.3390/cancers13236139
Szczyrek M, Grenda A, Kuźnar-Kamińska B, Krawczyk P, Sawicki M, Batura-Gabryel H, Mlak R, Szudy-Szczyrek A, Krajka T, Krajka A, et al. Methylation of DROSHA and DICER as a Biomarker for the Detection of Lung Cancer. Cancers. 2021; 13(23):6139. https://doi.org/10.3390/cancers13236139
Chicago/Turabian StyleSzczyrek, Michał, Anna Grenda, Barbara Kuźnar-Kamińska, Paweł Krawczyk, Marek Sawicki, Halina Batura-Gabryel, Radosław Mlak, Aneta Szudy-Szczyrek, Tomasz Krajka, Andrzej Krajka, and et al. 2021. "Methylation of DROSHA and DICER as a Biomarker for the Detection of Lung Cancer" Cancers 13, no. 23: 6139. https://doi.org/10.3390/cancers13236139
APA StyleSzczyrek, M., Grenda, A., Kuźnar-Kamińska, B., Krawczyk, P., Sawicki, M., Batura-Gabryel, H., Mlak, R., Szudy-Szczyrek, A., Krajka, T., Krajka, A., & Milanowski, J. (2021). Methylation of DROSHA and DICER as a Biomarker for the Detection of Lung Cancer. Cancers, 13(23), 6139. https://doi.org/10.3390/cancers13236139