
Is EBV Associated with Breast Cancer in Specific Geographic Locations?



Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Epstein–Barr Virus Lifecycle and B-Lymphocytes
1.2. Epstein–Barr Virus and Epithelial Cells
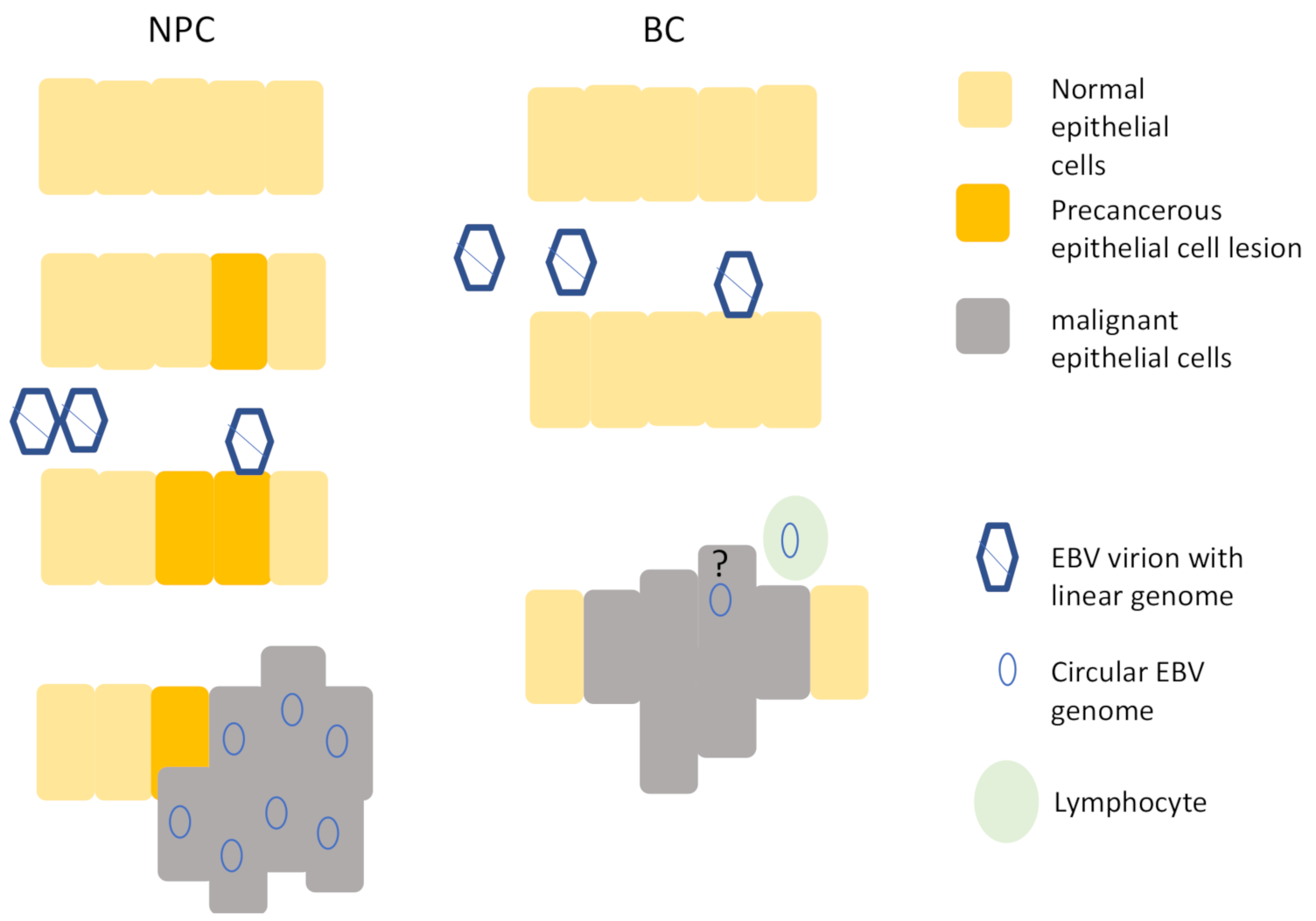
1.3. Established EBV-Associated Epithelial Cancers
“(i) A nucleic acid sequence belonging to a putative pathogen should be present in most cases of an infectious disease. Microbial nucleic acids should be found preferentially in those organs or gross anatomic sites known to be diseased (i.e., with anatomic, histologic, chemical, or clinical evidence of pathology) and not in those organs that lack pathology.
(ii) Fewer, or no, copy numbers of pathogen-associated nucleic acid sequences should occur in hosts or tissues without disease.”
1.4. The Potential of Epstein–Barr Virus in Breast Cancer
1.5. The Impact of EBV on Breast Epithelial Cells
2. Conclusions and Remaining Questions
- What is the geographic extent of BC biopsies with evidence of an EBV association and are EBV genomes in epithelial cells?
- Is there a specific sub-type of EBV that is more common in BC biopsies associated with EBV?
- What is the molecular signature of viral gene expression in BC biopsies associated with EBV?
- Do epidemiological factors relate to the association with EBV in these regions?
- Is there is a molecular signature of cell gene mutations in BC biopsies associated with EBV in these regions?
- What is the molecular signature of the expression of cell genes in BC biopsies associated with EBV in these regions?
Author Contributions
Funding
Conflicts of Interest
References
- Shannon-Lowe, C.; Rowe, M. Epstein Barr virus entry; kissing and conjugation. Curr. Opin. Virol. 2014, 4, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Farrell, P.J. Epstein–Barr Virus and Cancer. Annu. Rev. Pathol. Mech. Dis. 2019, 14, 29–53. [Google Scholar] [CrossRef] [PubMed]
- Pattle, S.B.; Farrell, P.J. The role of Epstein–Barr virus in cancer. Expert Opin. Biol. Ther. 2006, 6, 1193–1205. [Google Scholar] [CrossRef] [PubMed]
- Thompson, M.P.; Kurzrock, R. Epstein-Barr Virus and Cancer. Clin. Cancer Res. 2004, 10, 803–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epstein, M.A.; Achong, B.G.; Barr, Y.M. Virus Particles in Cultured Lymphoblasts from Burkitt’s Lymphoma. Lancet 1964, 1, 702–703. [Google Scholar] [CrossRef]
- Epstein, A. Why and How Epstein-Barr Virus Was Discovered 50 Years Ago. Curr. Top. Microbiol. Immunol. 2015, 390, 3–15. [Google Scholar] [CrossRef]
- Murray, P.G.; Young, L.S. An etiological role for the Epstein-Barr virus in the pathogenesis of classical Hodgkin lymphoma. Blood 2019, 134, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Saha, A.; Robertson, E.S. Mechanisms of B-Cell Oncogenesis Induced by Epstein-Barr Virus. J. Virol. 2019, 93, e00238-19. [Google Scholar] [CrossRef] [Green Version]
- Somasundaram, N.; Lim, J.Q.; Ong, C.K.; Lim, S.T. Pathogenesis and biomarkers of natural killer T cell lymphoma (NKTL). J. Hematol. Oncol. 2019, 12, 28. [Google Scholar] [CrossRef]
- Marquitz, A.R.; Mathur, A.; Edwards, R.H.; Raab-Traub, N. Host Gene Expression Is Regulated by Two Types of Noncoding RNAs Transcribed from the Epstein-Barr Virus BamHI A Rightward Transcript Region. J. Virol. 2015, 89, 11256–11268. [Google Scholar] [CrossRef] [Green Version]
- Tsao, S.W.; Tsang, C.M.; Lo, K.W. Epstein–Barr virus infection and nasopharyngeal carcinoma. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160270. [Google Scholar] [CrossRef]
- Xue, F.; He, X. Epstein–Barr Virus DNA in Nasopharyngeal Carcinoma: A Brief Review. Methods Mol. Biol. 2020, 2204, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-P.; Chan, A.T.C.; Le, Q.-T.; Blanchard, P.; Sun, Y.; Ma, J. Nasopharyngeal carcinoma. Lancet 2019, 394, 64–80. [Google Scholar] [CrossRef]
- Morales-Sanchez, A.; Fuentes-Panana, E.M. Epstein-Barr Virus-associated Gastric Cancer and Potential Mechanisms of Oncogenesis. Curr. Cancer Drug Targets 2017, 17, 534–554. [Google Scholar] [CrossRef]
- Naseem, M.; Barzi, A.; Brezden-Masley, C.; Puccini, A.; Berger, M.D.; Tokunaga, R.; Battaglin, F.; Soni, S.; McSkane, M.; Zhang, W.; et al. Outlooks on Epstein-Barr virus associated gastric cancer. Cancer Treat. Rev. 2018, 66, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Fukayama, M.; Abe, H.; Kunita, A.; Shinozaki-Ushiku, A.; Matsusaka, K.; Ushiku, T.; Kaneda, A. Thirty years of Epstein-Barr virus-associated gastric carcinoma. Virchows Archiv 2020, 476, 353–365. [Google Scholar] [CrossRef] [PubMed]
- Glaser, S.L.; Ambinder, R.F.; DiGiuseppe, J.A.; Horn-Ross, P.L.; Hsu, J.L. Absence of Epstein-Barr virus EBER-1 transcripts in an epidemiologically diverse group of breast cancers. Int. J. Cancer 1998, 75, 555–558. [Google Scholar] [CrossRef]
- Dadmanesh, F.; Peterse, J.L.; Sapino, A.; Fonelli, A.; Eusebi, V. Lymphoepithelioma-like carcinoma of the breast: Lack of evidence of Epstein-Barr virus infection. Histopathology 2001, 38, 54–61. [Google Scholar] [CrossRef]
- Herrmann, K.; Niedobitek, G. Lack of evidence for an association of Epstein–Barr virus infection with breast carcinoma. Breast Cancer Res. 2002, 5, R13–R17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, P.G.; Lissauer, D.; Junying, J.; Davies, G.; Moore, S.; Bell, A.; Timms, J.; Rowlands, D.; McConkey, C.; Reynolds, G.M.; et al. Reactivity with A monoclonal antibody to Epstein-Barr virus (EBV) nuclear antigen 1 defines a subset of aggressive breast cancers in the absence of the EBV genome. Cancer Res. 2003, 63, 2338–2343. [Google Scholar] [PubMed]
- Fina, F.; Romain, S.; Ouafik, L.; Palmari, J.; Ben Ayed, F.; Benharkat, S.; Bonnier, P.; Spyratos, F.; A Foekens, J.; Rose, C.; et al. Frequency and genome load of Epstein-Barr virus in 509 breast cancers from different geographical areas. Br. J. Cancer 2001, 84, 783–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preciado, M.V. Lack of evidence for an association of Epstein-Barr virus infection with breast carcinoma—another point of view. Breast Cancer Res. 2003, 5, E6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vetsika, E.-K.; Callan, M. Infectious mononucleosis and Epstein-Barr virus. Expert Rev. Mol. Med. 2004, 6, 1–16. [Google Scholar] [CrossRef]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Epstein-Barr Virus and Kaposi’s Sarcoma Herpes-virus/Human Herpesvirus 8; International Agency for Research on Cancer: Lyon, France, 1997; Volume 70. [Google Scholar]
- Laichalk, L.L.; Thorley-Lawson, D.A. Terminal Differentiation into Plasma Cells Initiates the Replicative Cycle of Epstein-Barr Virus In Vivo. J. Virol. 2005, 79, 1296–1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, W.-M.; Chan, K.H.; Chen, H.-L.; Luo, R.-X.; Ng, S.P.; Luk, W.; Zheng, B.-J.; Ji, M.-F.; Liang, J.-S.; Sham, J.S.T.; et al. Assessing the risk of nasopharyngeal carcinoma on the basis of EBV antibody spectrum. Int. J. Cancer 2001, 97, 489–492. [Google Scholar] [CrossRef]
- Shannon-Lowe, C.; Rowe, M. Epstein-Barr Virus Infection of Polarized Epithelial Cells via the Basolateral Surface by Memory B Cell-Mediated Transfer Infection. PLoS Pathog. 2011, 7, e1001338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, S.; Nishikawa, J.; Takada, K. Cell-to-Cell Contact as an Efficient Mode of Epstein-Barr Virus Infection of Diverse Human Epithelial Cells. J. Virol. 1998, 72, 4371–4378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tugizov, S.M.; Berline, J.W.; Palefsky, J.M. Epstein-Barr virus infection of polarized tongue and nasopharyngeal epithelial cells. Nat. Med. 2003, 9, 307–314. [Google Scholar] [CrossRef]
- Chen, J.; Sathiyamoorthy, K.; Zhang, X.; Schaller, S.; White, B.E.P.; Jardetzky, T.S.; Longnecker, R. Ephrin receptor A2 is a functional entry receptor for Epstein–Barr virus. Nat. Microbiol. 2018, 3, 172–180. [Google Scholar] [CrossRef]
- Zhang, H.; Li, Y.; Wang, H.-B.; Zhang, A.; Chen, M.-L.; Fang, Z.-X.; Dong, X.-D.; Li, S.-B.; Du, Y.; Xiong, D.; et al. Ephrin receptor A2 is an epithelial cell receptor for Epstein–Barr virus entry. Nat. Microbiol. 2018, 3, 164–171. [Google Scholar] [CrossRef]
- Young, L.S.; Lau, R.; Rowe, M.; Niedobitek, G.; Packham, G.; Shanahan, F.; Rowe, D.T.; Greenspan, D.; Greenspan, J.S.; Rickinson, A.B. Differentiation-associated expression of the Epstein-Barr virus BZLF1 transactivator protein in oral hairy leukoplakia. J. Virol. 1991, 65, 2868–2874. [Google Scholar] [CrossRef] [Green Version]
- Tsai, M.-H.; Raykova, A.; Klinke, O.; Bernhardt, K.; Gärtner, K.; Leung, C.S.; Geletneky, K.; Sertel, S.; Münz, C.; Feederle, R.; et al. Spontaneous Lytic Replication and Epitheliotropism Define an Epstein-Barr Virus Strain Found in Carcinomas. Cell Rep. 2013, 5, 458–470. [Google Scholar] [CrossRef] [Green Version]
- Fredericks, D.N.; A Relman, D. Sequence-based identification of microbial pathogens: A reconsideration of Koch’s postulates. Clin. Microbiol. Rev. 1996, 9, 18–33. [Google Scholar] [CrossRef]
- Wolf, H.; Hausen, H.Z.; Klein, G.; Becker, V.; Henle, G.; Henle, W. Attempts to detect virus-specific DNA sequences in human tumors. Med. Microbiol. Immunol. 1975, 161, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Young, L.S.; Dawson, C.W. Epstein-Barr virus and nasopharyngeal carcinoma. Chin. J. Cancer 2014, 33, 581–590. [Google Scholar] [CrossRef]
- Pathmanathan, R.; Prasad, U.; Chandrika, G.; Sadler, R.; Flynn, K.; Raab-Traub, N. Undifferentiated, nonkeratinizing, and squamous cell carcinoma of the nasopharynx. Variants of Epstein-Barr virus-infected neoplasia. Am. J. Pathol. 1995, 146, 1355–1367. [Google Scholar]
- Cai, W.; Rudolph, J.L.; Harrison, S.M.W.; Jin, L.; Frantz, A.L.; Harrison, D.A.; Andres, D.A. An evolutionarily conserved Rit GTPase–p38 MAPK signaling pathway mediates oxidative stress resistance. Mol. Biol. Cell 2011, 22, 3231–3241. [Google Scholar] [CrossRef]
- Aung, H.T.; Schroder, K.; Himes, S.R.; Brion, K.; Van Zuylen, W.; Trieu, A.; Suzuki, H.; Hayashizaki, Y.; Hume, D.A.; Sweet, M.J.; et al. LPS regulates proinflammatory gene expression in macrophages by altering histone deacetylase expression. FASEB J. 2006, 20, 1315–1327. [Google Scholar] [CrossRef]
- Nishikawa, J.; Iizasa, H.; Yoshiyama, H.; Shimokuri, K.; Kobayashi, Y.; Sasaki, S.; Nakamura, M.; Yanai, H.; Sakai, K.; Suehiro, Y.; et al. Clinical Importance of Epstein–Barr Virus-Associated Gastric Cancer. Cancers 2018, 10, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earl, H.M.; Vallier, A.-L.; Hiller, L.; Fenwick, N.; Young, J.; Iddawela, M.; Abraham, J.; Hughes-Davies, L.; Gounaris, I.; McAdam, K.; et al. Effects of the addition of gemcitabine, and paclitaxel-first sequencing, in neoadjuvant sequential epirubicin, cyclophosphamide, and paclitaxel for women with high-risk early breast cancer (Neo-tAnGo): An open-label, 2×2 factorial randomised phase 3 trial. Lancet Oncol. 2014, 15, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Lindahl, T.; Adams, A.; Bjursell, G.; Bornkamm, G.W.; Kaschka-Dierich, C.; Jehn, U. Covalently closed circular duplex DNA of Epstein-Barr virus in a human lymphoid cell line. J. Mol. Biol. 1976, 102, 511–530. [Google Scholar] [CrossRef]
- Raabtraub, N.; Flynn, K. The Structure of the Termini of the Epstein-Barr-Virus as a Marker of Clonal Cellular Proliferation. Cell 1986, 47, 883–889. [Google Scholar] [CrossRef]
- Fukayama, M.; Hayashi, Y.; Iwasaki, Y.; Chong, J.; Ooba, T.; Takizawa, T.; Koike, M.; Mizutani, S.; Miyaki, M.; Hirai, K. Epstein-Barr virus-associated gastric carcinoma and Epstein-Barr virus infection of the stomach. Lab. Investig. 1994, 71, 73–81. [Google Scholar] [PubMed]
- Labrecque, L.G.; Barnes, D.M.; Fentiman, I.S.; E Griffin, B. Epstein-Barr virus in epithelial cell tumors: A breast cancer study. Cancer Res. 1995, 55, 39–45. [Google Scholar]
- Huo, Q.; Zhang, N.; Yang, Q. Epstein-Barr Virus Infection and Sporadic Breast Cancer Risk: A Meta-Analysis. PLoS ONE 2012, 7, e31656. [Google Scholar] [CrossRef] [Green Version]
- Jin, Q.E.; Su, J.R.; Yan, D.H.; Wu, S.N. Epstein-Barr Virus Infection and Increased Sporadic Breast Carcinoma Risk: A Meta-Analysis. Med. Princ. Pract. 2019, 29, 2. [Google Scholar] [CrossRef] [PubMed]
- Farahmand, M.; Monavari, S.H.; Shoja, Z.; Ghaffari, H.; Tavakoli, M.; Tavakoli, A. Epstein–Barr virus and risk of breast cancer: A systematic review and meta-analysis. Futur. Oncol. 2019, 15, 2873–2885. [Google Scholar] [CrossRef] [PubMed]
- Niedobitek, G.; Herbst, H.; Ian, D.A.; Tim, H.D. In Situ Detection of Epstein-Barr Virus and Phenotype Determination of EBV-Infected Cells. In Situ Hybrid. Protoc. 2006, 326, 115–138. [Google Scholar] [CrossRef]
- Gulley, M.L.; Tang, W. Laboratory Assays for Epstein-Barr Virus-Related Disease. J. Mol. Diagn. 2008, 10, 279–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshpande, C.G.; Badve, S.; Kidwai, N.; Longnecker, R. Lack of Expression of the Epstein-Barr Virus (EBV) Gene Products, EBERs, EBNA1, LMP1, and LMP2A, in Breast Cancer Cells. Lab. Investig. 2002, 82, 1193–1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pai, T.; Gupta, S.; Gurav, M.; Nag, S.; Shet, T.; Patil, A.; Desai, S. Evidence for the association of Epstein-Barr Virus in breast cancer in Indian patients using in-situ hybridization technique. Breast J. 2017, 24, 16–22. [Google Scholar] [CrossRef]
- Fessahaye, G.; Elhassan, A.M.; Elamin, E.M.; Adam, A.A.M.; Ghebremedhin, A.; Ibrahim, M.E. Association of Epstein - Barr virus and breast cancer in Eritrea. Infect. Agents Cancer 2017, 12, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- A Yahia, Z.; Adam, A.A.; Elgizouli, M.; Hussein, A.; A Masri, M.; Kamal, M.; Mohamed, H.S.; Alzaki, K.; Elhassan, A.M.; Hamad, K.; et al. Epstein Barr virus: A prime candidate of breast cancer aetiology in Sudanese patients. Infect. Agents Cancer 2014, 9, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zekri, A.-R.N.; Bahnassy, A.A.; Mohamed, W.S.; El-Kassem, F.A.; El-Khalidi, S.J.; Hafez, M.M.; Hassan, Z.K. Epstein-Barr virus and breast cancer: Epidemiological and Molecular study on Egyptian and Iraqi women. J. Egypt. Natl. Cancer Inst. 2012, 24, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baltzell, K.; Buehring, G.C.; Krishnamurthy, S.; Kuerer, H.; Shen, H.M.; Sison, J.D. Epstein–Barr virus is seldom found in mammary epithelium of breast cancer tissue using in situ molecular methods. Breast Cancer Res. Treat. 2011, 132, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Luo, M.-L.M.; Desmedt, C.; Nabavi, S.S.; Yadegarynia, S.S.; A Hong, A.; Konstantinopoulos, P.A.; Gabrielson, E.E.; Hines-Boykin, R.R.; Pihan, G.G.; et al. Epstein–Barr Virus Infection of Mammary Epithelial Cells Promotes Malignant Transformation. EBioMedicine 2016, 9, 148–160. [Google Scholar] [CrossRef] [Green Version]
- Arbach, H.; Viglasky, V.; Lefeu, F.; Guinebretière, J.-M.; Ramirez, V.; Bride, N.; Boualaga, N.; Bauchet, T.; Peyrat, J.-P.; Mathieu, M.-C.; et al. Epstein-Barr Virus (EBV) Genome and Expression in Breast Cancer Tissue: Effect of EBV Infection of Breast Cancer Cells on Resistance to Paclitaxel (Taxol). J. Virol. 2006, 80, 845–853. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sinclair, A.J.; Moalwi, M.H.; Amoaten, T. Is EBV Associated with Breast Cancer in Specific Geographic Locations? Cancers 2021, 13, 819. https://doi.org/10.3390/cancers13040819
Sinclair AJ, Moalwi MH, Amoaten T. Is EBV Associated with Breast Cancer in Specific Geographic Locations? Cancers. 2021; 13(4):819. https://doi.org/10.3390/cancers13040819
Chicago/Turabian StyleSinclair, Alison J., Manal H. Moalwi, and Thomas Amoaten. 2021. "Is EBV Associated with Breast Cancer in Specific Geographic Locations?" Cancers 13, no. 4: 819. https://doi.org/10.3390/cancers13040819
APA StyleSinclair, A. J., Moalwi, M. H., & Amoaten, T. (2021). Is EBV Associated with Breast Cancer in Specific Geographic Locations? Cancers, 13(4), 819. https://doi.org/10.3390/cancers13040819