Taccalonolides: A Novel Class of Microtubule-Stabilizing Anticancer Agents


Abstract
:Simple Summary
Abstract
1. Introduction
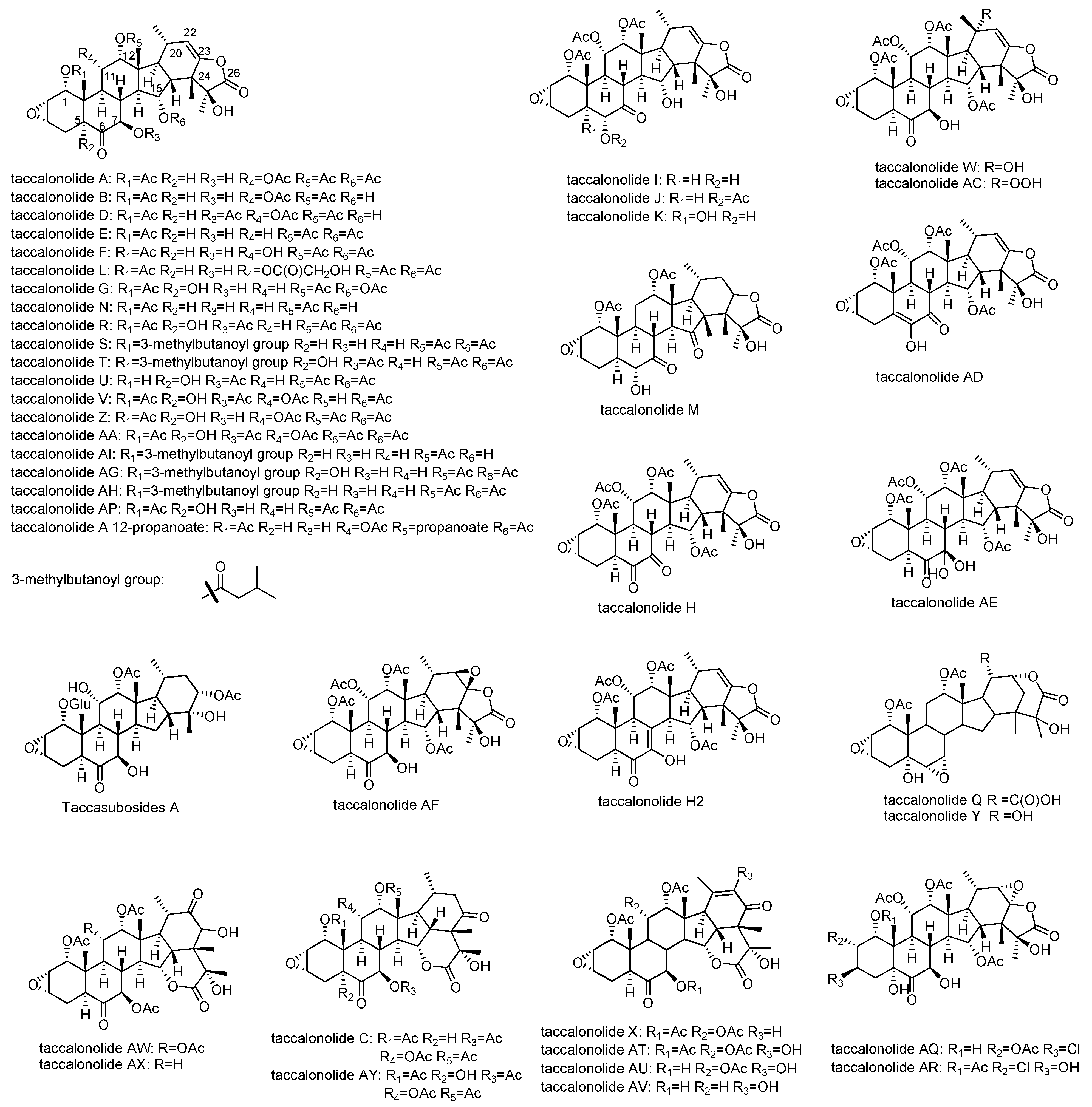
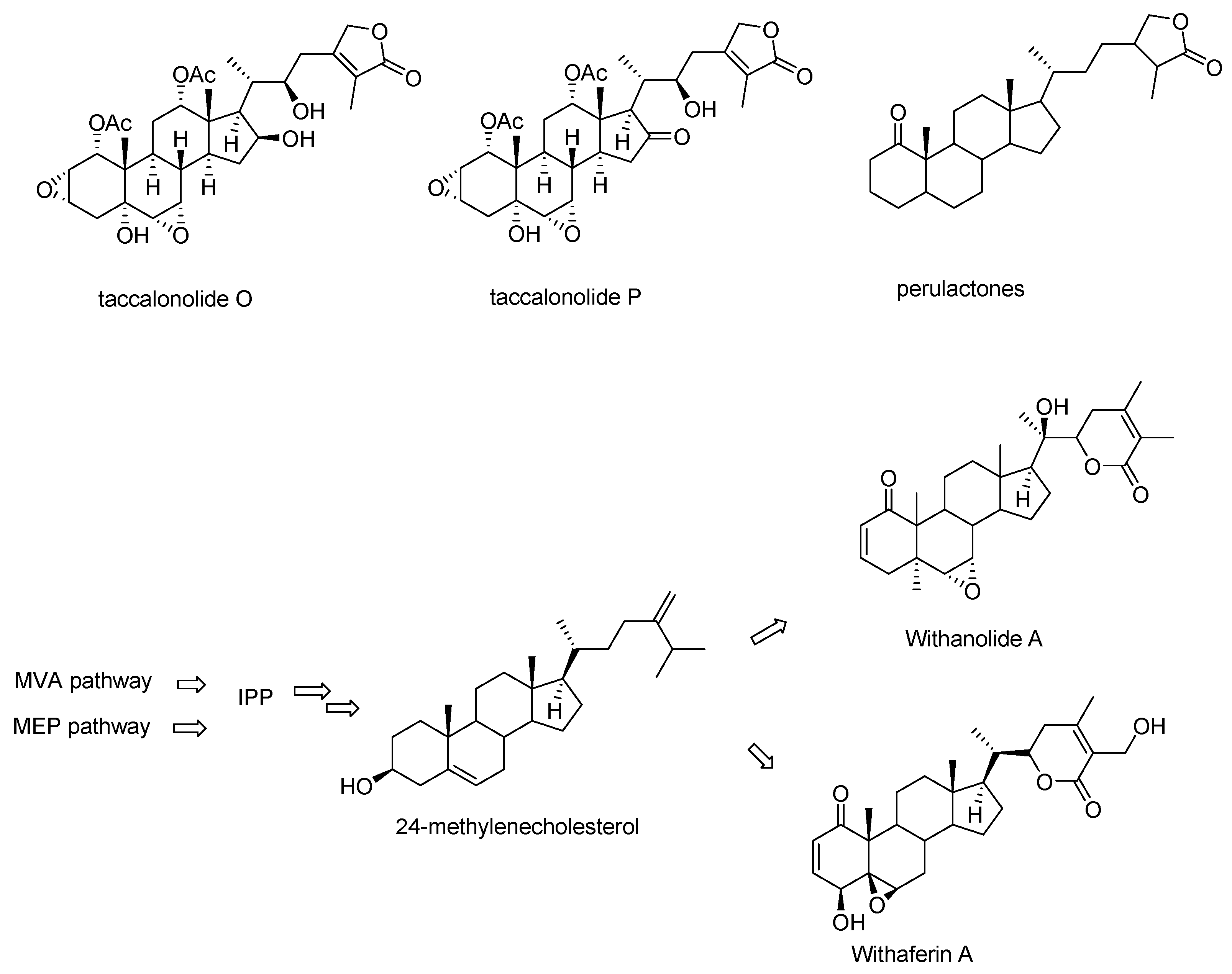
2. Structures of Natural Taccalonolides (1987–2020)
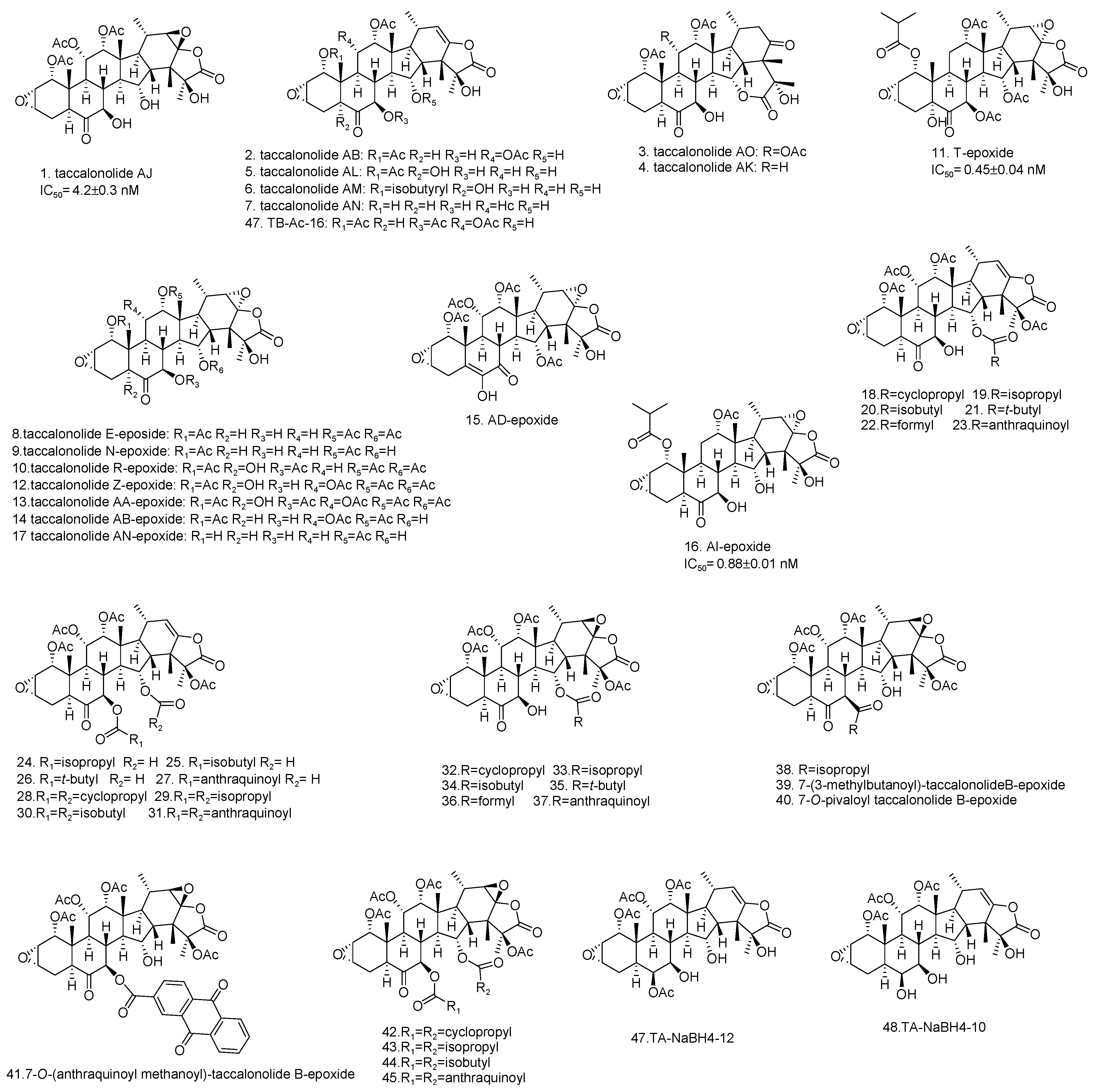
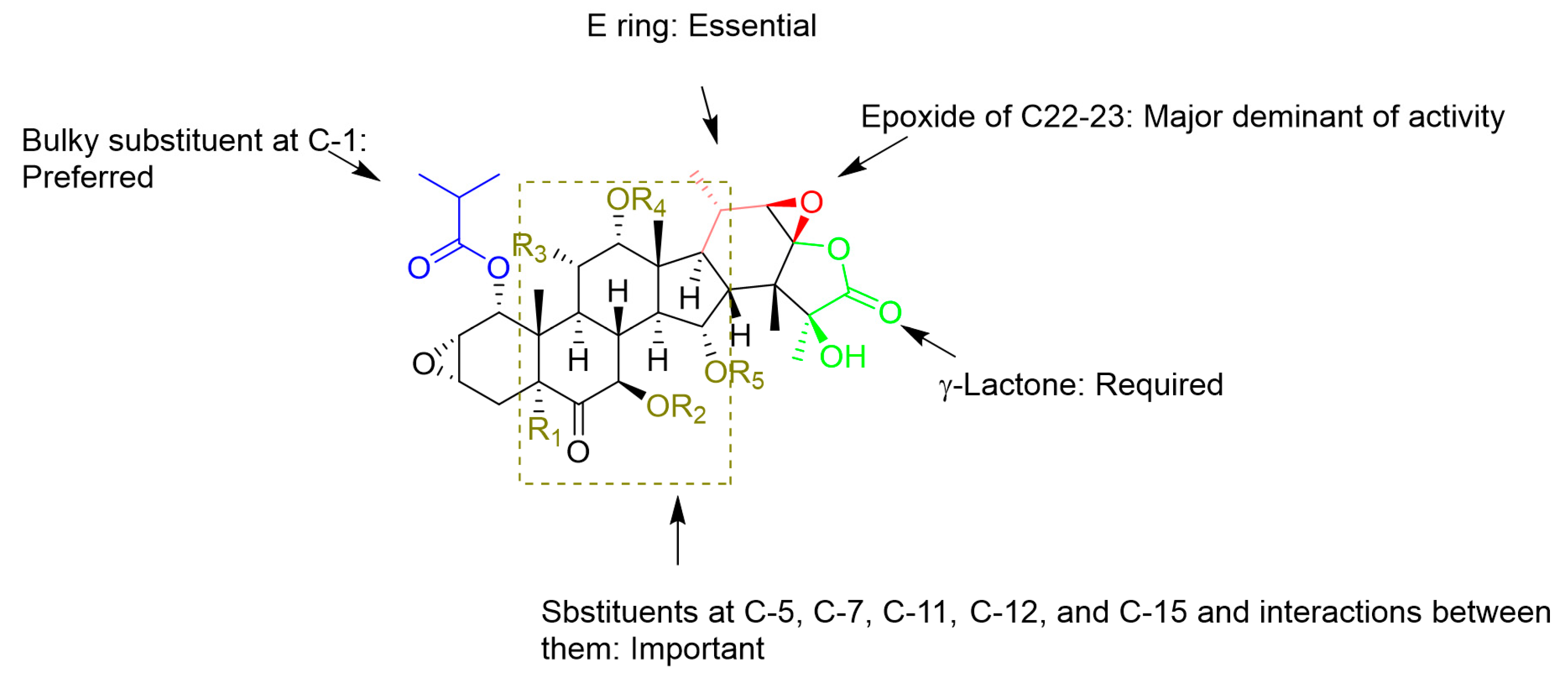
3. Semisynthetic Taccalonolides and Structure-Activity Relationships (SARs)
4. Pharmacological Effects and Mechanism of Action of Taccalonolides
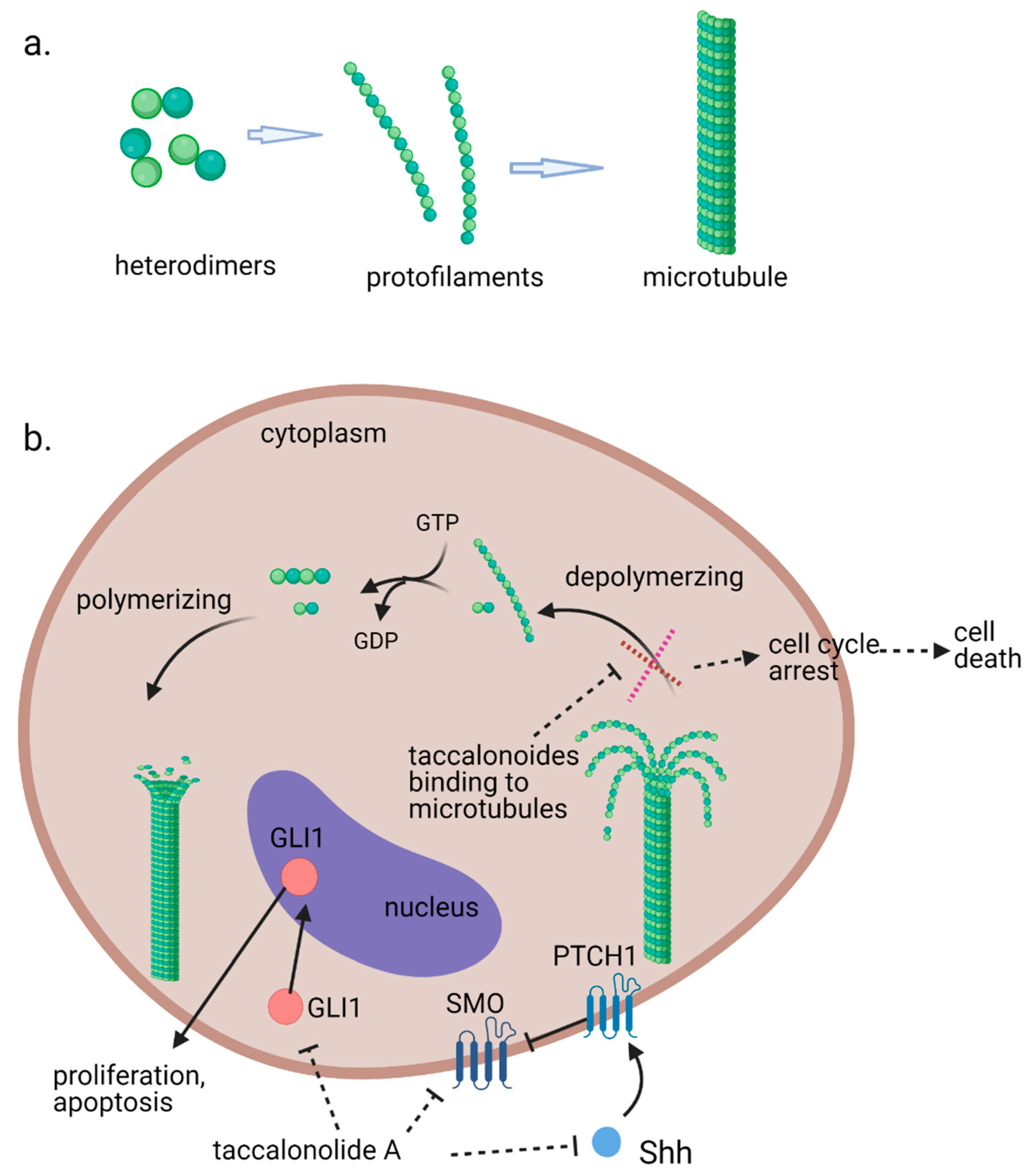
4.1. Cellular Actions on Interphase Microtubules, Mitosis, and Cell Cycle
4.2. In Vitro Antiproliferative Effects and In Vivo Antitumor Efficacy
4.3. Tubulin Polymerization and Microtubule Stabilization Caused by Taccalonolides
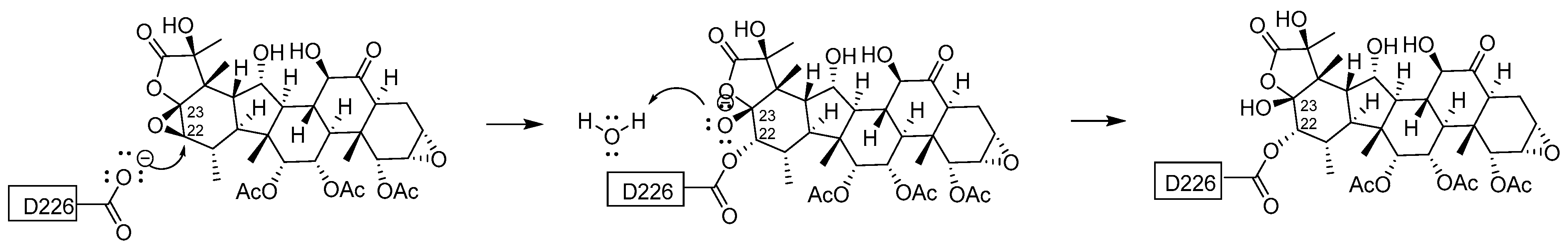
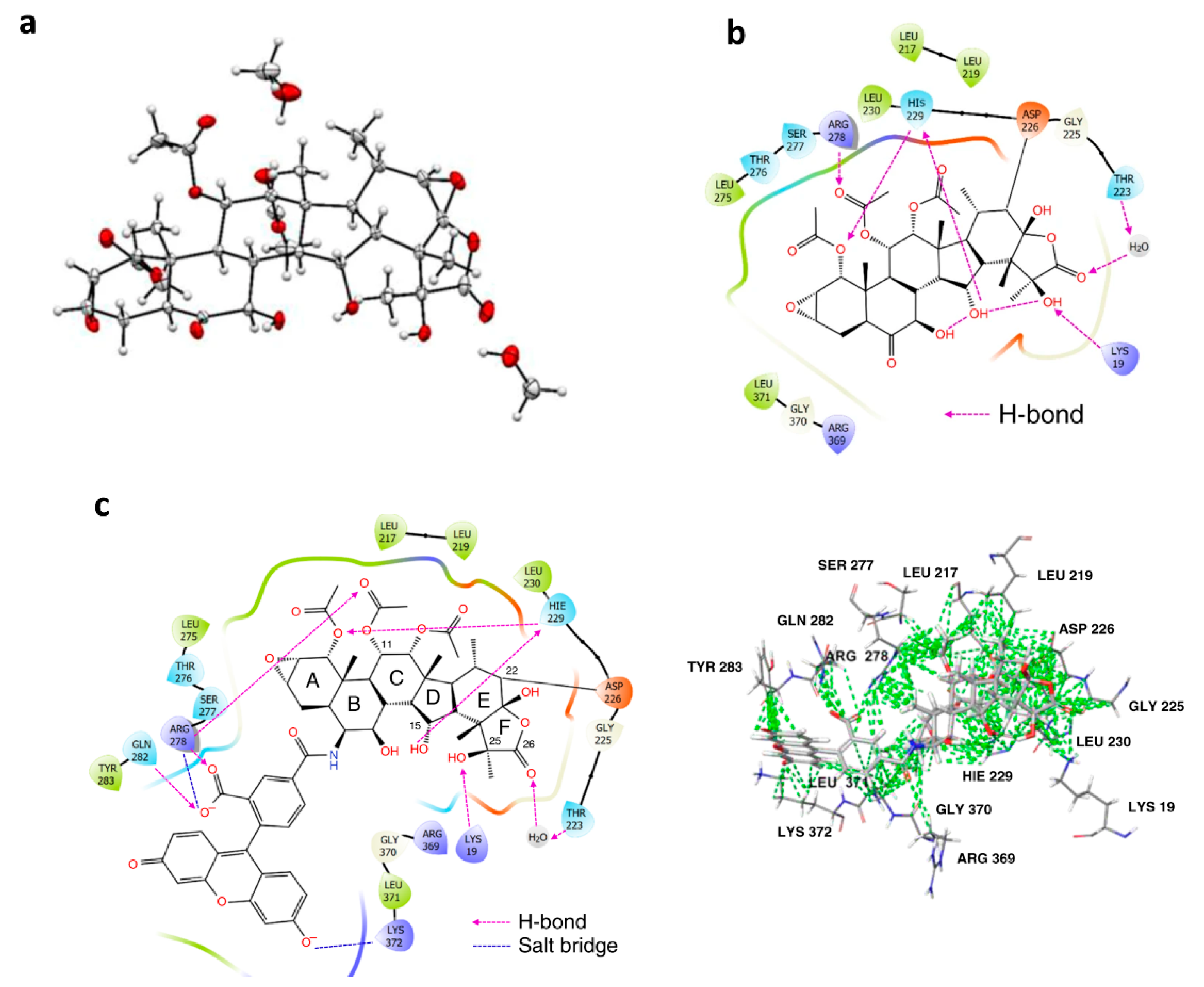
4.4. Define the Covalent Binding
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Margolin, G.; Gregoretti, I.V.; Cickovski, T.M.; Li, C.; Shi, W.; Alber, M.S.; Goodson, H.V. The mechanisms of microtubule catastrophe and rescue: Implications from analysis of a dimer-scale computational model. Mol. Biol. Cell 2012, 23, 642–656. [Google Scholar] [CrossRef] [PubMed]
- Cooper, G.; Hausman, R.E. The Cell: A Molecular Approach; Sinauer Associates: Oxford, UK, 2000. [Google Scholar]
- Jordan, M.A.; Wilson, L. Microtubules as a target for anticancer drugs. Nat. Rev. Cancer 2004, 4, 253–265. [Google Scholar] [CrossRef]
- Kavallaris, M.; Verrills, N.M.; Hill, B.T. Anticancer therapy with novel tubulin-interacting drugs. Drug Resist. Updat. 2001, 4, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Conde, C.; Cáceres, A. Microtubule assembly, organization and dynamics in axons and dendrites. Nat. Rev. Neurosci. 2009, 10, 319–332. [Google Scholar] [CrossRef]
- Palaparthi, S.; Pidaparti, R. Mimicking Sub-Structures Self-Organization in Microtubules. Biomimetics 2019, 4, 71. [Google Scholar] [CrossRef] [Green Version]
- Rohena, C.C.; Mooberry, S.L. Recent progress with microtubule stabilizers: New compounds, binding modes and cellular activities. Nat. Prod. Rep. 2014, 31, 335–355. [Google Scholar] [CrossRef] [Green Version]
- Horio, T.; Murata, T. The role of dynamic instability in microtubule organization. Front. Plant Sci. 2014, 5, 511. [Google Scholar] [CrossRef] [Green Version]
- Stanton, R.A.; Gernert, K.M.; Nettles, J.H.; Aneja, R. Drugs that target dynamic microtubules: A new molecular perspective. Med. Res. Rev. 2011, 31, 443–481. [Google Scholar] [CrossRef] [Green Version]
- Wilmes, A.; Bargh, K.; Kelly, C.; Northcote, A.P.T.; Miller, J.H. Peloruside A Synergizes with Other Microtubule Stabilizing Agents in Cultured Cancer Cell Lines. Mol. Pharm. 2007, 4, 269–280. [Google Scholar] [CrossRef]
- Dumontet, C.; Jordan, M.A. Microtubule-binding agents: A dynamic field of cancer therapeutics. Nat. Rev. Drug Discov. 2010, 9, 790–803. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.-L.; Wang, B.-D.; Chen, M.-Q. Steroidal bitter principles from Tacca plantaginea structures of taccalonolide A and B. Tetrahedron Lett. 1987, 28, 1673–1675. [Google Scholar] [CrossRef]
- Tinley, T.L.; Randall-Hlubek, D.A.; Leal, R.M.; Jackson, E.M.; Cessac, J.W.; Quada, J.C.; Hemscheidt, T.K.; Mooberry, S.L. Taccalonolides E and A. Cancer Res. 2003, 63, 3211. [Google Scholar]
- Rimkus, T.K.; Carpenter, R.L.; Qasem, S.; Chan, M.; Lo, H.-W. Targeting the Sonic Hedgehog Signaling Pathway: Review of Smoothened and GLI Inhibitors. Cancers 2016, 8, 22. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Risinger, A.L.; Mooberry, S.L. Taccalonolide microtubule stabilizers. Bioorganic Med. Chem. 2014, 22, 5091–5096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Risinger, A.L.; Peng, J.; Chen, Z.; Hu, L.; Mooberry, S.L. Potent Taccalonolides, AF and AJ, Inform Significant Structure–Activity Relationships and Tubulin as the Binding Site of These Microtubule Stabilizers. J. Am. Chem. Soc. 2011, 133, 19064–19067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Murcia, P.A.; Mills, A.; Cortés-Cabrera, Á.; Gago, F. Unravelling the covalent binding of zampanolide and taccalonolide AJ to a minimalist representation of a human microtubule. J. Comput. Mol. Des. 2019, 33, 627–644. [Google Scholar] [CrossRef]
- Du, L.; Yee, S.S.; Ramachandran, K.; Risinger, A.L. Elucidating target specificity of the taccalonolide covalent microtubule stabilizers employing a combinatorial chemical approach. Nat. Commun. 2020, 11, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Yang, H.; Wang, Y.; Chen, Y. Phytochemical and Pharmacological Studies of the Genus Tacca: A Review. Trop. J. Pharm. Res. 2014, 13, 635. [Google Scholar] [CrossRef] [Green Version]
- Misico, R.I.; Nicotra, V.E.; Oberti, J.C.; Barboza, G.; Gil, R.R.; Burton, G. Withanolides and Related Steroids. Fortschr. Chem. Org. Nat. Prog. Chem. Org. Nat. Prod. 2011, 94, 127–229. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Liu, J.-K.; Mühlbauer, A.; Henkel, T. Three Novel Taccalonolides from the Tropical Plant Tacca subflaellata. Helv. Chim. Acta 2002, 85, 2553–2558. [Google Scholar] [CrossRef]
- Ray, A.B.; Gupta, M. Withasteroids, a growing group of naturally occurring steroidal lactones. In Fortschritte der Chemie organischer Naturstoffe/Progress in the Chemistry of Organic Natural Products; Springer: London, UK, 1994; pp. 1–106. [Google Scholar]
- Ni, G.; Yang, H.-Z.; Fu, N.-J.; Zhang, L.-L.; Wang, M.-C.; Chen, J.; Zhang, C.-L.; Li, Y.; Chen, X.-G.; Chen, R.-Y.; et al. Cytotoxic Taccalonolides and Withanolides from Tacca chantrieri. Planta Med. 2015, 81, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Knoch, E.; Sugawara, S.; Mori, T.; Poulsen, C.; Fukushima, A.; Harholt, J.; Fujimoto, Y.; Umemoto, N.; Saito, K. Third DWF1 paralog in Solanaceae, sterol Δ24-isomerase, branches withanolide biosynthesis from the general phytosterol pathway. Proc. Natl. Acad. Sci. 2018, 115, E8096–E8103. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Rather, G.A.; Misra, P.; Dhar, M.K.; Lattoo, S.K. Jasmonate responsive transcription factor WsMYC2 regulates the biosynthesis of triterpenoid withanolides and phytosterol via key pathway genes in Withania somnifera (L.) Dunal. Plant Mol. Biol. 2019, 100, 543–560. [Google Scholar] [CrossRef]
- Dhar, N.; Razdan, S.; Rana, S.; Bhat, W.W.; A Vishwakarma, R.; Lattoo, S.K. A Decade of Molecular Understanding of Withanolide Biosynthesis and In vitro Studies in Withania somnifera (L.) Dunal: Prospects and Perspectives for Pathway Engineering. Front. Plant Sci. 2015, 6, 1031. [Google Scholar] [CrossRef] [Green Version]
- Stein, A.; Compera, D.; Karge, B.; Brönstrup, M.; Franke, J. Isolation and characterisation of irinans, androstane-type withanolides from Physalis peruviana L. Beilstein J. Org. Chem. 2019, 15, 2003–2012. [Google Scholar] [CrossRef] [Green Version]
- Risinger, A.L.; Mooberry, S.L. Cellular studies reveal mechanistic differences between taccalonolide A and paclitaxel. Cell Cycle 2011, 10, 2162–2171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mühlbauer, A.; Seip, S.; Nowak, A.; Tran, V.S. Five Novel Taccalonolides from the Roots of the Vietnamese Plant Tacca paxiana. Helv. Chim. Acta 2003, 86, 2065–2072. [Google Scholar] [CrossRef]
- Peng, J.; Risinger, A.L.; Fest, G.A.; Jackson, E.M.; Helms, G.; Polin, L.A.; Mooberry, S.L. Identification and Biological Activities of New Taccalonolide Microtubule Stabilizers. J. Med. Chem. 2011, 54, 6117–6124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Peng, J.; Risinger, A.L.; Mooberry, S.L. Hydrolysis Reactions of the Taccalonolides Reveal Structure–Activity Relationships. J. Nat. Prod. 2013, 76, 1369–1375. [Google Scholar] [CrossRef] [Green Version]
- Tian, H.; He, Z. Anti-hepatoma effect of taccalonolide A through suppression of sonic hedgehog pathway. Artif. Cells Nanomed. Biotechnol. 2020, 48, 939–947. [Google Scholar] [CrossRef]
- Risinger, A.L.; Jackson, E.M.; Polin, L.A.; Helms, G.L.; Leboeuf, D.A.; Joe, P.A.; Hopper-Borge, E.; Ludueña, R.F.; Kruh, G.D.; Mooberry, S.L. The Taccalonolides: Microtubule Stabilizers That Circumvent Clinically Relevant Taxane Resistance Mechanisms. Cancer Res. 2008, 68, 8881–8888. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.-L.; Wang, B.-D.; Shen, J.-H. Taccalonolide C and D, two pentacyclic steroids of Tacca plantaginea. Phytochemistry 1988, 27, 2999–3001. [Google Scholar] [CrossRef]
- Shen, J.-H.; Chen, Z.-L.; Gao, Y.-S. The pentacyclic steroidal constituents of Tacca plantaginea: Taccalonolide E and F. Chin. J. Chem. 2010, 9, 92–94. [Google Scholar] [CrossRef]
- Shen, J.; Chen, Z.; Gao, Y. Taccalonolides from Tacca plantaginea. Phytochemistry 1996, 42, 891–893. [Google Scholar] [CrossRef]
- Chen, Z.-L.; Shen, J.-H.; Gao, Y.-S.; Wichtl, M. Five Taccalonolides from Tacca plantaginea. Planta Med. 1997, 63, 40–43. [Google Scholar] [CrossRef]
- Yang, J.; Zhao, R.; Chen, C.; Ni, W.; Teng, F.; Hao, X.; Liu, H. Taccalonolides W–Y, Three New Pentacyclic Steroids from Tacca plantaginea. Helv. Chim. Acta 2008, 91, 1077–1082. [Google Scholar] [CrossRef]
- Ola, A.R.B.; Risinger, A.L.; Du, L.; Zammiello, C.L.; Peng, J.; Cichewicz, R.H.; Mooberry, S.L. Taccalonolide Microtubule Stabilizers Generated Using Semisynthesis Define the Effects of Mono Acyloxy Moieties at C-7 or C-15 and Disubstitutions at C-7 and C-25. J. Nat. Prod. 2017, 81, 579–593. [Google Scholar] [CrossRef] [PubMed]
- Risinger, A.L.; Li, J.; Bennett, M.J.; Rohena, C.C.; Peng, J.; Schriemer, D.C.; Mooberry, S.L. Taccalonolide Binding to Tubulin Imparts Microtubule Stability and Potent In Vivo Activity. Cancer Res. 2013, 73, 6780–6792. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Ni, W.; Li, X.-R.; Hua, Y.; Fang, P.-L.; Kong, L.-M.; Pan, L.-L.; Li, Y.; Chen, C.-X.; Liu, H.-Y. Taccasubosides A–D, four new steroidal glycosides from Tacca subflabellata. Steroids 2011, 76, 1037–1042. [Google Scholar] [CrossRef]
- Peng, J.; Risinger, A.L.; Li, J.; Mooberry, S.L. Synthetic Reactions with Rare Taccalonolides Reveal the Value of C-22,23 Epoxidation for Microtubule Stabilizing Potency. J. Med. Chem. 2014, 57, 6141–6149. [Google Scholar] [CrossRef]
- Dike, V.T.; Vihiior, B.; Bosha, J.A.; Yin, T.M.; Ebiloma, G.U.; de Koning, H.P.; Igoli, J.O.; Gray, A.I. Antitrypanosomal Activity of a Novel Taccalonolide from the Tubers of Tacca leontopetaloides. Phytochem. Anal. 2016, 27, 217–221. [Google Scholar] [CrossRef] [PubMed]
- The Board of Regents of the University of Texas System. US Patent for Taccalonolide Microtubule Stabilizers. Patent No. 10,501,490 B2, 10 December 2019.
- Wang, Y.; Yu, Y.; Li, G.-B.; Li, S.-A.; Wu, C.; Gigant, B.; Qin, W.; Chen, H.; Wu, Y.; Chen, Q.; et al. Mechanism of microtubule stabilization by taccalonolide AJ. Nat. Commun. 2017, 8, 15787. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Risinger, A.L.; Yee, S.S.; Ola, A.R.B.; Zammiello, C.L.; Cichewicz, R.H.; Mooberry, S.L. Identification of C-6 as a New Site for Linker Conjugation to the Taccalonolide Microtubule Stabilizers. J. Nat. Prod. 2019, 82, 583–588. [Google Scholar] [CrossRef]
- Buey, R.M.; Barasoain, I.; Jackson, E.; Meyer, A.; Giannakakou, P.; Paterson, I.; Mooberry, S.; Andreu, J.M.; Díaz, J.F. Microtubule Interactions with Chemically Diverse Stabilizing Agents: Thermodynamics of Binding to the Paclitaxel Site Predicts Cytotoxicity. Chem. Biol. 2005, 12, 1269–1279. [Google Scholar] [CrossRef] [Green Version]
- Rohena, C.C.; Peng, J.; Johnson, T.A.; Crews, P.; Mooberry, S.L. Chemically diverse microtubule stabilizing agents initiate distinct mitotic defects and dysregulated expression of key mitotic kinases. Biochem. Pharmacol. 2013, 85, 1104–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Risinger, A.L.; Riffle, S.M.; Lopus, M.; A Jordan, M.; Wilson, L.; Mooberry, S.L. The taccalonolides and paclitaxel cause distinct effects on microtubule dynamics and aster formation. Mol. Cancer 2014, 13, 41. [Google Scholar] [CrossRef] [Green Version]
- Risinger, A.L.; Li, J.; Du, L.; Benavides, R.; Robles, A.J.; Cichewicz, R.H.; Kuhn, J.G.; Mooberry, S.L. Pharmacokinetic Analysis and in Vivo Antitumor Efficacy of Taccalonolides AF and AJ. J. Nat. Prod. 2017, 80, 409–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.; Zhang, S.; Niu, J.; Zhang, C.; Dai, W.; Wu, Y.; Hu, L. Development of Taccalonolide AJ-Hydroxypropyl-β-Cyclodextrin Inclusion Complexes for Treatment of Clear Cell Renal-Cell Carcinoma. Molecules 2020, 25, 5586. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound Name | Species | Biological Activities (Year) | Ref. |
|---|---|---|---|---|
| 1 | Taccalonolide A | T. plantaginea, T. chantrieri, T. paxiana | Bundling of interphase microtubules in HeLa cells at 250 nM. (2011) 5 µM taccalonolide A induced bundles, multipolar spindles, and multiple micronuclei on Human lung carcinoma A549 cells. (2005) Initiating Bcl-2 phosphorylation, MAPK activation, and apoptosis. (2003) In vivo antitumor effects on the syngeneic murine mammary carcinoma 16/C model. (2011) Inhibiting cell proliferation of two drug-sensitive cell lines SK-OV-3 and SK-OV-3/MDR-1-6/6 with IC50s of 0.6 µM and 2.5 µM. (2008) Inhibiting cell proliferation of taxol-resistant cell lines, PTX 10 and PTX 22 with IC50s of 7.05 µM and 6.40 µM, and the epothilone-resistant cell line, 1A9/A8 with IC50 of 8.89 µM. (2003) Inhibiting proliferation of HepG2 and Huh7 cells with IC50 values for HepG2 and Huh7 cells 11.9 µM and 16.8 µM, respectively. (2020) In vivo against a doxorubicin- and paclitaxel-resistant Pgp-expressing tumor in syngeneic mammary 17/ADR model. (2008) | [12,13,16,28,29,30,31,32,33] |
| 2 | Taccalonolide B | T. plantaginea, T. paxiana | Antiproliferative effects in HeLa cells. (2011) Inhibiting cell proliferation of two drug-sensitive cell lines SK-OV-3 and SK-OV-3/MDR-1-6/6 with IC50s of 0.2 µM and 2.5 µM. (2008) Mitotic arrest and bundling of interphase microtubules in HeLa cells at 0.8 μM. (2011) | [12,16,29,30,31,33] |
| 3 | Taccalonolide C | T. plantaginea | N/A 1 | [34] |
| 4 | Taccalonolide D | T. plantaginea | N/A | [34] |
| 5 | Taccalonolide E | T. plantaginea T. chantrieri, T. paxiana, | Mitotic accumulating in the G2-M phase of the cell cycle. (2003, 2005) Forming of multiple aberrant mitotic spindles and initiating micronucleation in interphase A-10 cells at 1µM. (2003) Bundling of interphase microtubules. Antiproliferative effects in HeLa cells. (2003) Inhibiting cell proliferation of two drug-sensitive cell lines SK-OV-3 and SK-OV-3/MDR-1-6/6 with IC50s of 0.7 µM and 3.6 µM. Inhibiting cell proliferation of taxol-resistant cell lines, PTX 10 and PTX 22 with IC50s of 1.64 µM and 4.01 µM, and the epothilone-resistant cell line, 1A9/A8 with IC50 of 1.42 µM. (2003, 2008) In vivo antitumor effects on the syngeneic murine mammary carcinoma 16/C model. (2011) In vivo against a doxorubicin- and paclitaxel- resistant Pgp-expressing tumor in syngeneic mammary 17/ADR model. (2008) | [13,29,30,31,33,35] |
| 6 | Taccalonolide F | T. plantaginea | N/A | [35] |
| 7–8 | Taccalonolides L and M | T. plantaginea | N/A | [36] |
| 9–11 | Taccalonolides G, H, and J | T. plantaginea | N/A | [37] |
| 12 | Taccalonolide I | T. plantaginea | Antiproliferative effects in HeLa cells (2013) | [31,37] |
| 13 | Taccalonolide K | T. plantaginea, T. paxiana | N/A | [29,37] |
| 14 | Taccalonolide N | T. paxiana | Antiproliferative effects in HeLa cells. (2013) Inhibiting cell proliferation of two drug-sensitive cell lines SK-OV-3 and SK-OV-3/MDR-1-6/6 with IC50s of 0.2 µM and 1.2 µM. Mitotic arrest and bundling of microtubules in HeLa cells at 1.3 μM. (2011) In vivo antitumor effects on the syngeneic murine mammary carcinoma 16/C model. (2011) | [29,30,31,33] |
| 15 | Taccalonolide Q | T. subflaellata | N/A | [21] |
| 16 | Taccalonolide R | T. paxiana T. chantrieri | Mitotic arrest and bundling of microtubules in HeLa cells at 57 μM. (2011) | [29,30] |
| 17 | Taccalonolide S | T. paxiana | N/A | [29] |
| 18 | Taccalonolide T | T. paxiana T. chantrieri | Antiproliferative effects in HeLa cells. Mitotic arrest and bundling of microtubules in HeLa cells at 3.5 μM. (2011) | [29,30] |
| 19–20 | Taccalonolide U and V | T. paxiana | N/A | [29] |
| 21–23 | Taccalonolides W, X, Y | T. plantaginea | N/A | [38] |
| 24 | Taccalonolide Z | T. integrifolia | Antiproliferative effects in HeLa cells using the SRB assay. Mitotic arrest and bundling of microtubules in HeLa cells at 0.6 μM. (2011) | [30] |
| 25 | Taccalonolides AA | T. chantrieri | Antiproliferative effects in HeLa cells. (2011) Mitotic arrest and bundling of interphase microtubules in HeLa cells at 0.32 μM. (2011) | [30] |
| 26 | Taccalonolides AC | T. plantaginea | Lack of potency (IC50>50,000 nM against HeLa cells). (2013) | [16,31] |
| 27–28 | Taccalonolides AD-AE | T. plantaginea | Increasing in cellular microtubule density and microtubule bundling in HeLa cells at 17 and 25 μM. Antiproliferative actions in HeLa cells with IC50 of 3.48 and 5.01 μM. (2011) | [16] |
| 29 | Taccalonolid AF | T. plantaginea | Increasing the density of interphase microtubules HeLa cells. (2011) Causing HeLa cells to arrest in the G2/M phase of the cell cycle with multiple aberrant mitotic spindles at 100 nM. (2011) Stimulating the polymerization of purified tubulin. (2011) High antiproliferative potency in HeLa cells. (2011) In vivo antitumor effect in the MDA-MB-231 breast cancer xenograft model. (2013) | [16,39,40] |
| 30 | Taccalonolides H2 | T. plantaginea | Increasing the density of interphase microtubules HeLa cells. Causing HeLa cells to arrest in the G2/M phase of the cell cycle with multiple aberrant mitotic spindles. Antiproliferative potency in HeLa cells using the SRB assay. (2011) | [16] |
| 31 | Taccasuboside A | T. subflabellata | Lack of potency (2011) | [41] |
| 32 | Taccalonolid AI | T. chantrieri | Potent antiproliferative effect in HeLa cells. (2014) | [42] |
| 33–38 | Taccalonolides AT-AY | T. chantrieri | Devoid of cytotoxicity. (2015) | [23] |
| 39 | Taccalonolide A 12-propanoate | T. leontopetaloides | Antitrypanosomal activity against Trypanosoma brucei brucei with the EC50 value of 3.13 + 0.089 μg/mL. (2016) | [43] |
| 40–41 | Taccalonolides AG, AH | T. chantrieri | Antiproliferative in HeLa cells. (2019) | [44] |
| 42–44 | Taccalonolides AP, AQ, and AR | T. chantrieri |
| Substituent | Compound Name |
|---|---|
| 11-H | taccalonolides E, G, N, U, AI, AG, AH, AP |
| 6-OH or 6-OAc | Taccalonolide I, J, K, M, AD |
| 6=O and 7=O | Taccalonolide H |
| 7=O and 15=O | Taccalonolide M |
| 20-OOH | Taccalonolide AC |
| δ-lactone between C15 and C24 | Taccalonolides Q, Y |
| δ-lactone be-tweenC22 and C24 | Taccalonolides C, X, AY, AW, AT, AU, AV, AX, |
| unsaturated ring B | Taccalonolides H2 and AD |
| unsaturated ring E | Taccalonolides M, F, AQ, AR and |
| No δ-lactone or γ-lactone on E ring | Taccasubosides A |
| No. | Compound Name | Biological Activities (Year) | Ref. |
|---|---|---|---|
| 1 | Taccalonolide AJ | Stimulating the polymerization of purified tubulin. (2011) Increasing the density of interphase microtubules in HeLa cells at 30 nM. (2011) Causing HeLa cells to arrest in the G2/M phase of the cell cycle with multiple aberrant mitotic spindles. (2011) High antiproliferative potency in HeLa cells. (2011) In vivo antitumor effect in the MDA-MB-231 breast cancer xenograft model with the absence of therapeutic window. Taccalonolide AJ showed excellent antitumor efficacy only when directly administered to the tumor. (2013) | [16,39,40] |
| 2 | Taccalonolide AB | Mitotic arrest and bundling of interphase microtubules in HeLa cells at 13.5 μM. (2011) Antiproliferative action in HeLa cells. (2014) | [30,42] |
| 3–4 | Taccalonolides AO, AK | Lack of potency (IC50>50 μM against HeLa cells). No microtubule stabilizing effects at 50 μM. (2013) | [31] |
| 5–7 | Taccalonolides AL, AM, AN | Antiproliferative actions in HeLa cells. Increase in the cellular density of interphase microtubules in HeLa cells at 10-50 μM. (2013) | [31] |
| 8–17 | Taccalonolide E-epoxide, Taccalonolide N-epoxide, Taccalonolide R-epoxide, Taccalonolide T-epoxide, Taccalonolide Z-epoxide, Taccalonolide AA-epoxide, Taccalonolide AB-epoxide, Taccalonolide AD-epoxide, Taccalonolide AI-epoxide, Taccalonolide AN-epoxide, | Causing interphase microtubule bundling in HeLa cells. Antiproliferative activities in HeLa cells. Taccalonolide T-epoxide and Taccalonolide AI-epoxide enhanced polymerization of purified tubulin in turbidimetric assays. Taccalonolide T-epoxide and Taccalonolide AI-epoxide possess antitumor efficacy in the MDA-MB-231 triple negative breast cancer xenograft mice model. (2014) | [42] |
| 18–45 | 7-(3-methylbutanoyl)-taccalonolide B-epoxide and other C-7, C-15, and C-7, C-25 acyloxy taccalonolide B and their corresponding C-22, C-23 epoxides | Antiproliferative effects against the HeLa cell line. Eliciting the tubulin polymerization in HeLa cells. In vivo antitumor activities in female athymic nude mice with transplanted NCI/ADR-RES tumor fragments. (2018) | [39] |
| 46 | TB-AC-16 | Antiproliferative effects in HeLa cells. (2019) | [45] |
| 47 | TA-NaBH4-12 | ||
| 48 | TA-NaBH4-10 | ||
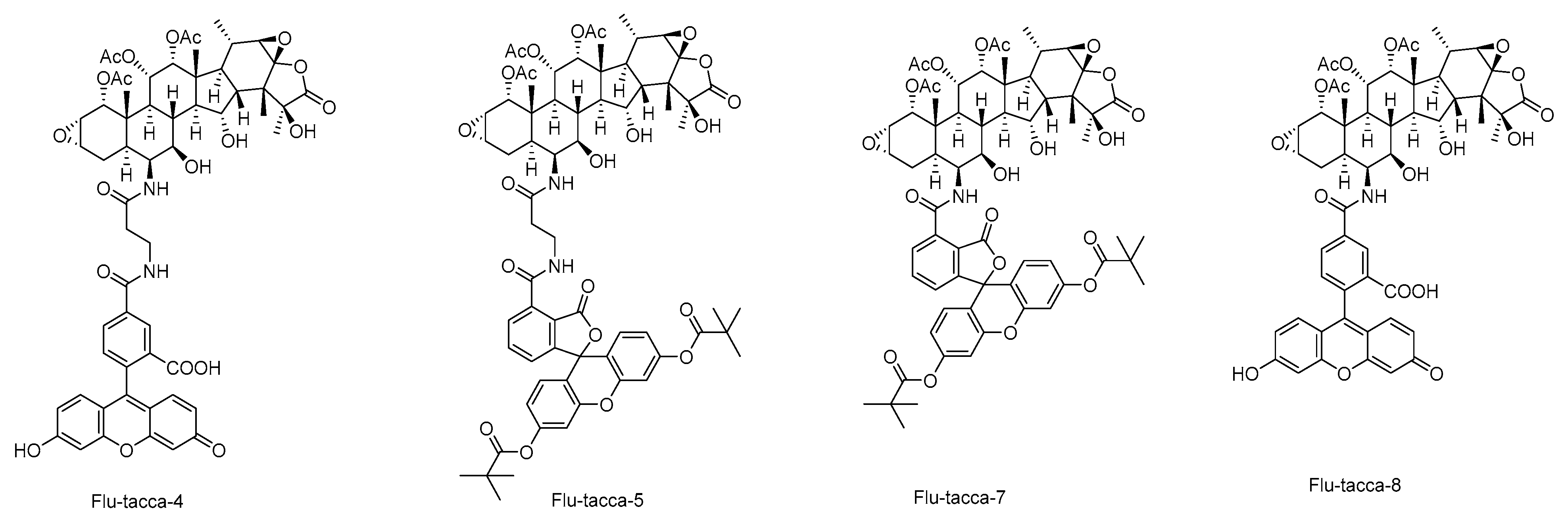
| 49–56 | Flu-tacca-1 ~Flu-tacca-8 | Causing a decrease in the proliferation of HeLa or SK-OV-3 cells. Polymerizing purified tubulin. Binding to endogenous β-tubulin of HCC1937 cells evaluated by immunoblotting. (2020) | [18] |
| Compound | IC50 (nM) 1 | Compound | IC50 (nM) |
|---|---|---|---|
| Taccalonolide A | 5380 ± 230 | Taccalonolide AE | 5010 ± 210 |
| Taccalonolide B | 3120 ± 180 | Taccalonolide AF | 23 ± 3 |
| Taccalonolide I | 49,200 ± 2800 | Taccalonolide AJ | 4.2 ± 0.3 |
| Taccalonolide T | 335 ± 24 | Taccalonolide H2 | 730 ± 20 |
| Taccalonolide R | 13,144 ± 1390 | Taccalonolide AL | 34,400 ± 7500 |
| Taccalonolide E | 39,500 ± 4700 | Taccalonolide AO | >50,000 |
| Taccalonolide N | 8500 ± 400 | Taccalonolide AK | >50,000 |
| Taccalonolide Z | 120 ± 7.5 | Taccalonolide AM | 2000 ± 100 |
| Taccalonolide AA | 32.3 ± 1.9 | Taccalonolide AN | 1500 ± 100 |
| Taccalonolide AB | 2767 ± 107 | Taccalonolide AI | 47 ± 3 |
| Taccalonolide AC | >50,000 | Taccalonolide AI-epoxide | 0.88 ± 0.01 |
| Taccalonolide AD | 3480 ± 230 | Taccalonolide T-epoxide | 0.45 ± 0.04 |
| Taccalonolide Z-epoxide | 17.2 ± 0.3 | Taccalonolide AN-epoxide | 685 ± 19 |
| 7- O- pivaloyl taccalonolide B-epoxide | 9 ± 1 | 7-O-(anthraquinoyl methanoyl)-taccalonolide B-epoxide | 22 ± 3 |
| 7-O-(3-methylbutanoyl)-taccalonolide B-epoxide | 2.4 ± 0.7 | Flu-tacca-7 | 31 ± 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Winstead, A.; Yu, H.; Peng, J. Taccalonolides: A Novel Class of Microtubule-Stabilizing Anticancer Agents. Cancers 2021, 13, 920. https://doi.org/10.3390/cancers13040920
Chen X, Winstead A, Yu H, Peng J. Taccalonolides: A Novel Class of Microtubule-Stabilizing Anticancer Agents. Cancers. 2021; 13(4):920. https://doi.org/10.3390/cancers13040920
Chicago/Turabian StyleChen, Xiaoyan, Angela Winstead, Hongtao Yu, and Jiangnan Peng. 2021. "Taccalonolides: A Novel Class of Microtubule-Stabilizing Anticancer Agents" Cancers 13, no. 4: 920. https://doi.org/10.3390/cancers13040920
APA StyleChen, X., Winstead, A., Yu, H., & Peng, J. (2021). Taccalonolides: A Novel Class of Microtubule-Stabilizing Anticancer Agents. Cancers, 13(4), 920. https://doi.org/10.3390/cancers13040920