
Aminopeptidase Expression in Multiple Myeloma Associates with Disease Progression and Sensitivity to Melflufen

 ,
,  , , , ,
, , , ,  , , and
, , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. RNA Sequencing and Analysis
2.3. Data Validation Using the CoMMpass Dataset
2.4. Exome Sequencing and Cytogenetics
2.5. Liquid Chromatography–Tandem Mass Spectrometry-Based Proteomics
2.6. Proteomics Data Analysis
2.7. Flow Cytometry-Based Drug Sensitivity Testing
2.8. Cell Lines and Viability Assay
2.9. Hydrolysis Assay
3. Results
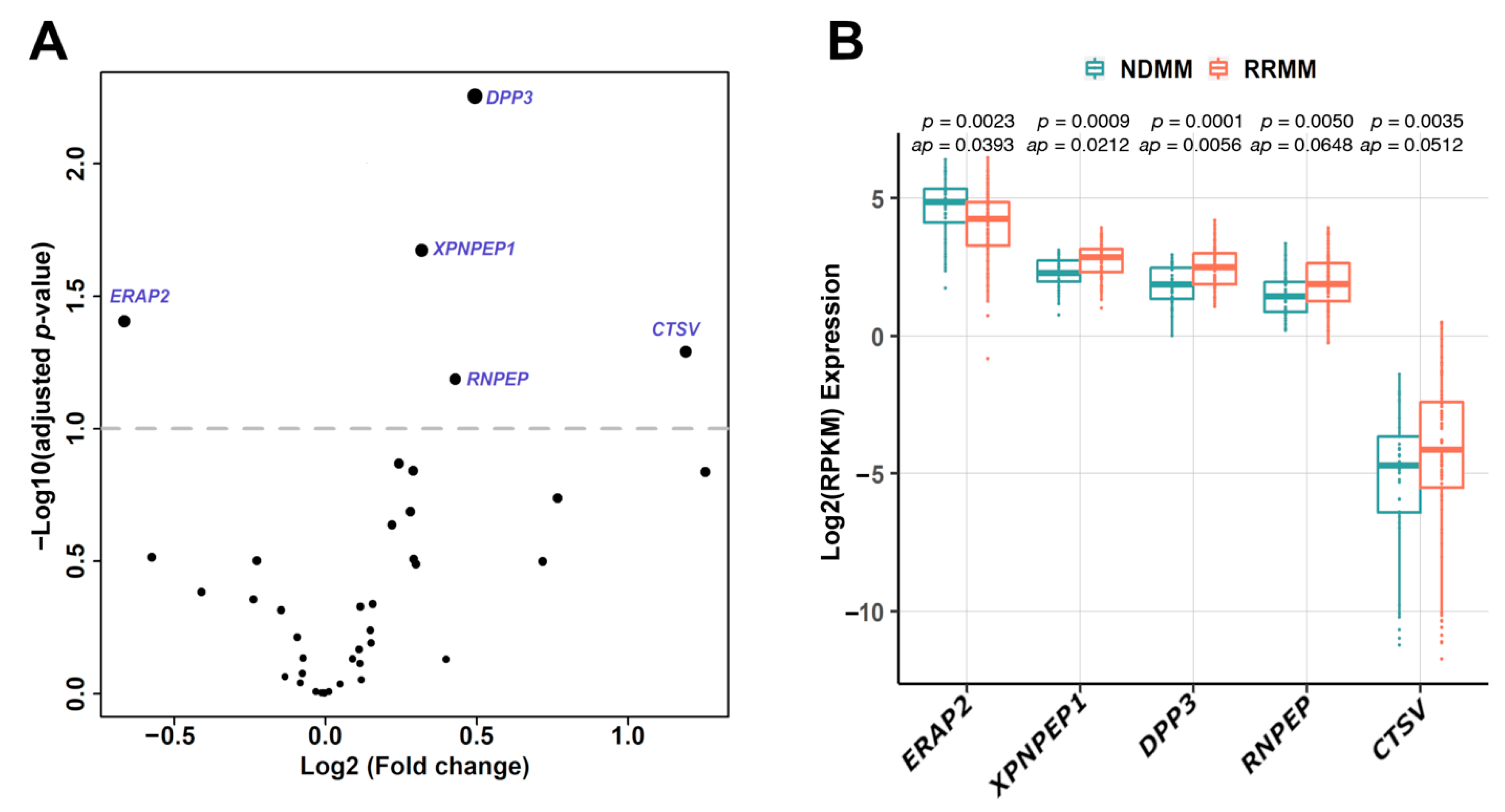
3.1. Differential Expression of Aminopeptidase Genes in MM
3.2. Association between Aminopeptidase Gene Expression and MM Disease Status
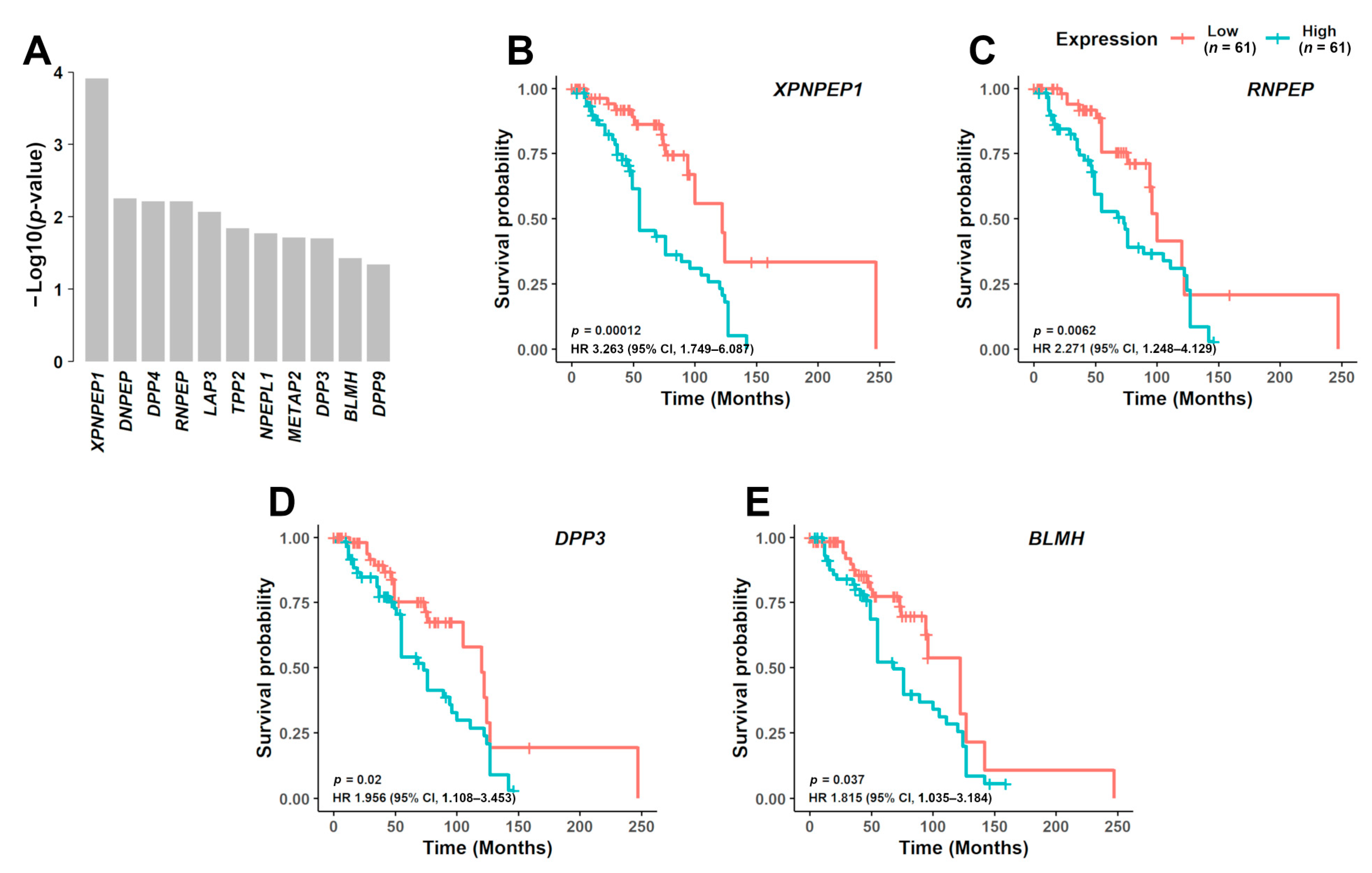
3.3. Prognostic Significance of Aminopeptidase Gene Expression
3.4. Aminopeptidase Somatic Mutation and CNV Characteristics in Myeloma
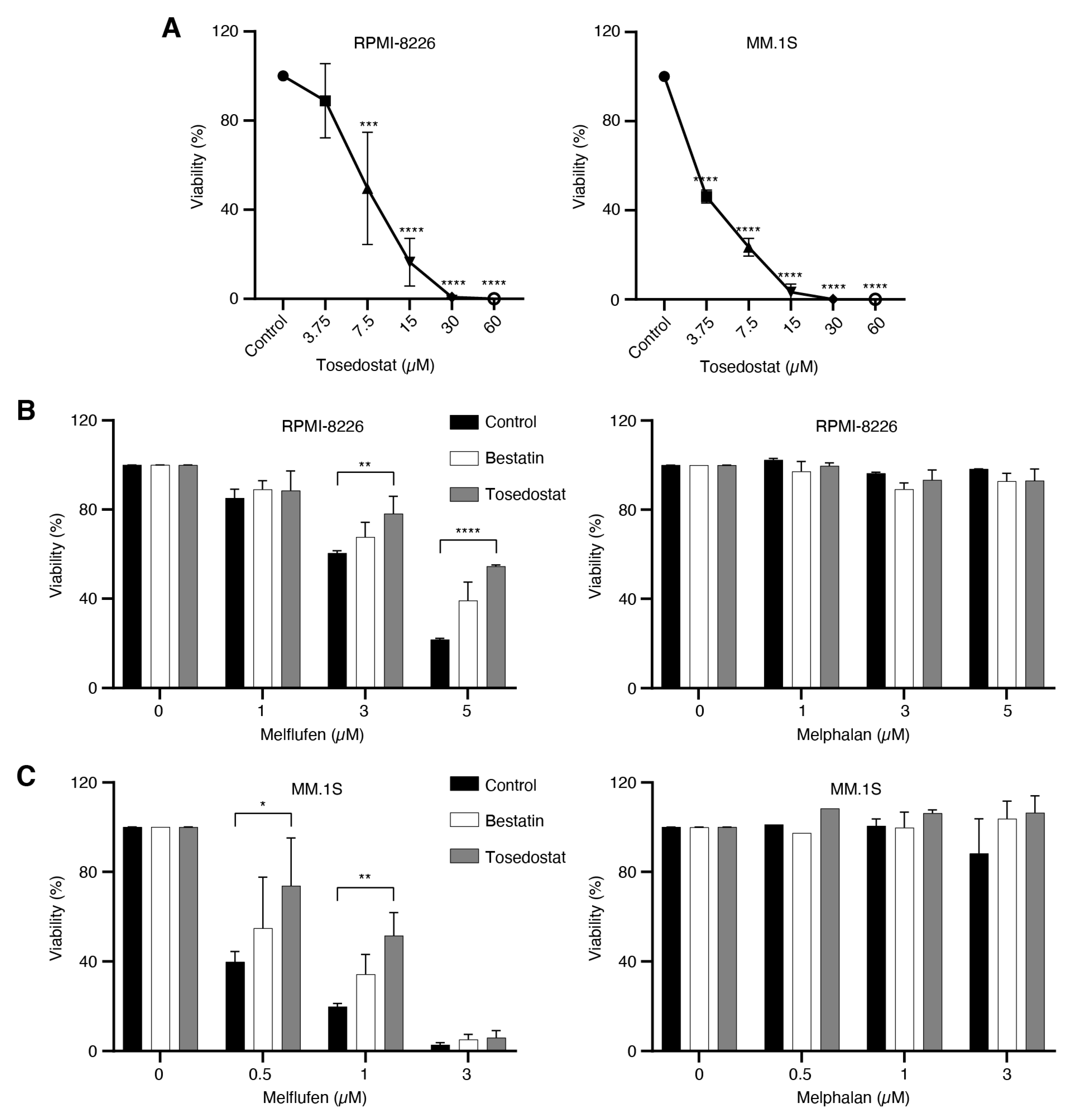
3.5. MM Patient Bone Marrow CD138+CD38+ Plasma Cells Are Sensitive to Melflufen
3.6. RRMM Samples Are More Sensitive to Melflufen than NDMM Samples
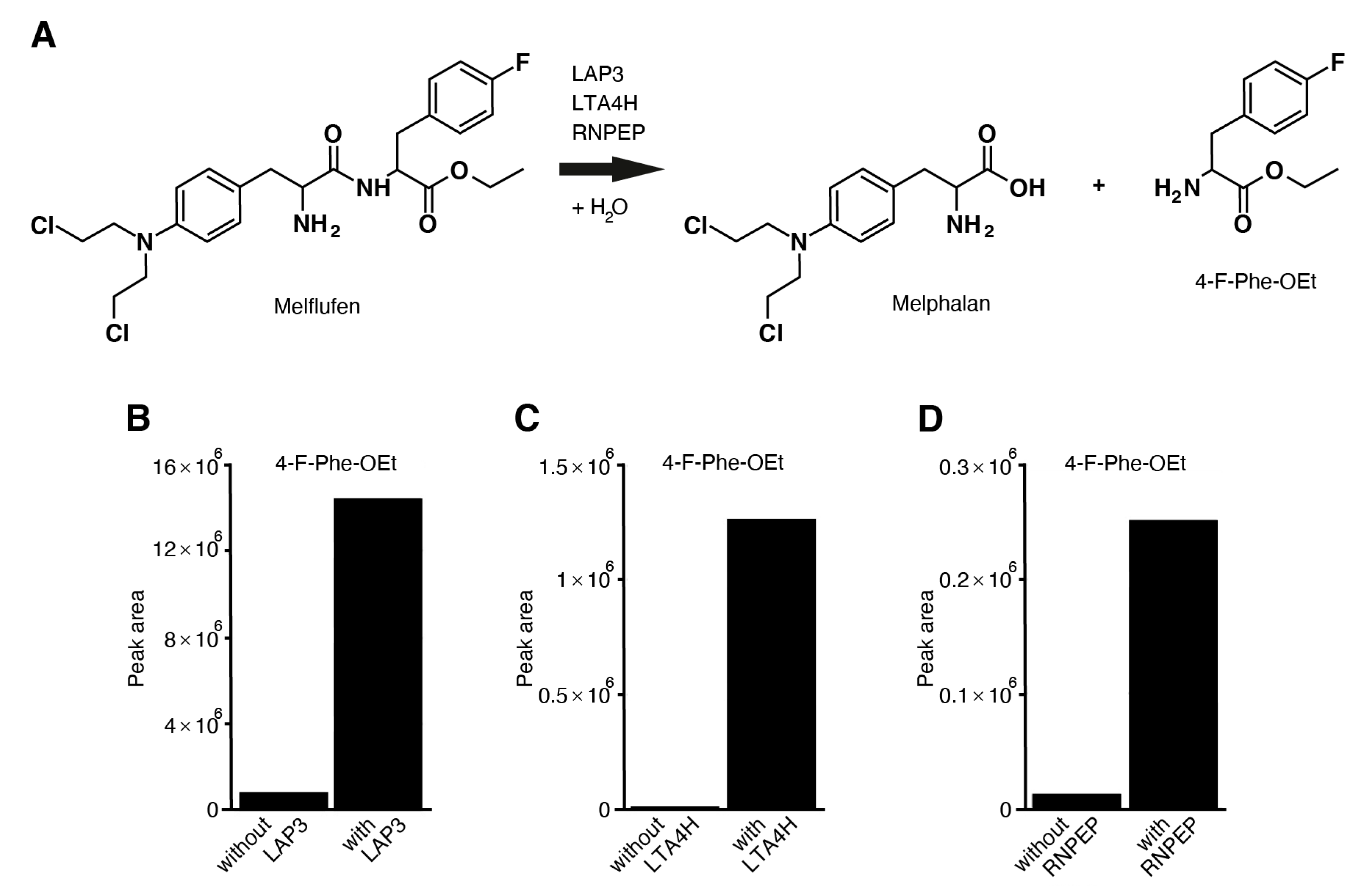
3.7. Melflufen Activity Is Dependent on Aminopeptidase Activity
3.8. Aminopeptidases LAP3, LTA4H, RNPEP, DPP7, and ANPEP Hydrolyze Melflufen
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moore, H.E.; Davenport, E.L.; Smith, E.M.; Muralikrishnan, S.; Dunlop, A.S.; Walker, B.A.; Krige, D.; Drummond, A.H.; Hooftman, L.; Morgan, G.J.; et al. Aminopeptidase inhibition as a targeted treatment strategy in myeloma. Mol. Cancer Ther. 2009, 8, 762–770. [Google Scholar] [CrossRef] [Green Version]
- Leleu, X.; Martin, T.G.; Einsele, H.; Lyons, R.M.; Durie, B.G.; Iskander, K.S.; Ailawadhi, S. Role of Proteasome Inhibitors in Relapsed and/or Refractory Multiple Myeloma. Clin. Lymphoma Myeloma Leuk. 2019, 19, 9–22. [Google Scholar] [CrossRef]
- Sanderink, G.-J.; Artur, Y.; Siest, G. Human Aminopeptidases: A Review of the Literature. Clin. Chem. Lab. Med. 1988, 26, 795–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ackerman, G.A. Histochemical demonstration of aminopeptidase activity in the leukocytes of blood and bone marrow. J. Histochem. Cytochem. 1960, 8, 386. [Google Scholar] [CrossRef] [Green Version]
- Ackerman, G.A. Histochemical Demonstration of Aminopeptidase Activity in the Cells of the Blood and Bone Marrow from Various Hæmatological Disorders. Nat. Cell Biol. 1963, 197, 189–190. [Google Scholar] [CrossRef] [PubMed]
- Dalal, B.I.; Al Mugairi, A.; Pi, S.; Lee, S.Y.; Khare, N.S.; Pal, J.; Bryant, A.; Vakil, A.P.; Lau, S.; Mourad, Y.R.A. Aberrant Expression of CD13 Identifies a Subgroup of Standard-Risk Adult Acute Lymphoblastic Leukemia With Inferior Survival. Clin. Lymphoma Myeloma Leuk. 2014, 14, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Craddock, K.J.; Chen, Y.; Brandwein, J.M.; Chang, H. CD13 expression is an independent adverse prognostic factor in adults with Philadelphia chromosome negative B cell acute lymphoblastic leukemia. Leuk. Res. 2013, 37, 759–764. [Google Scholar] [CrossRef]
- Lau, H.; Nagy, A.; Atwater, S.K.; Cascio, M.J.; Ohgami, R.S. An integrated flow cytometry analysis of 286 mature B cell neoplasms identifies CD13 as a useful marker for diagnostic subtyping. Int. J. Lab. Hematol. 2018, 40, 715–720. [Google Scholar] [CrossRef] [PubMed]
- García-Sanz, R.; Orfão, A.; González, M.; Tabernero, M.; Bladé, J.; Moro, M.; Fernández-Calvo, J.; Sanz, M.; Pérez-Simoón, J.; Rasillo, A.; et al. Primary Plasma Cell Leukemia: Clinical, Immunophenotypic, DNA Ploidy, and Cytogenetic Characteristics. Blood 1999, 93, 1032–1037. [Google Scholar] [CrossRef] [PubMed]
- Raimbault, A.; Machherndl-Spandl, S.; Itzykson, R.; Clauser, S.; Chapuis, N.; Mathis, S.; Lauf, J.; Alary, A.-S.; Burroni, B.; Kosmider, O.; et al. CD13 expression in B cell malignancies is a hallmark of plasmacytic differentiation. Br. J. Haematol. 2019, 184, 625–633. [Google Scholar] [CrossRef] [Green Version]
- Shim, H.; Ha, J.H.; Lee, H.; Sohn, J.Y.; Kim, H.J.; Eom, H.-S.; Kong, S.-Y. Expression of Myeloid Antigen in Neoplastic Plasma Cells Is Related to Adverse Prognosis in Patients with Multiple Myeloma. BioMed Res. Int. 2014, 2014, 1–8. [Google Scholar] [CrossRef]
- Kakodkar, P.; More, S.; András, K.; Papakonstantinou, N.; Kelly, S.; Makrooni, M.A.; Ortutay, C.; Szegezdi, E. Aspartic Aminopeptidase Is a Novel Biomarker of Aggressive Chronic Lymphocytic Leukemia. Cancers 2020, 12, 1876. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.W.; Chen-Harris, H.; Mayba, O.; Lianoglou, S.; Wuster, A.; Bhangale, T.; Khan, Z.; Mariathasan, S.; Daemen, A.; Reeder, J.; et al. Germline genetic polymorphisms influence tumor gene expression and immune cell infiltration. Proc. Natl. Acad. Sci. USA 2018, 115, E11701–E11710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, T.; Tatekoshi, A.; Takada, K.; Iyama, S.; Kamihara, Y.; Jawaid, P.; Rehman, M.U.; Noguchi, K.; Kondo, T.; Kajikawa, S.; et al. DPP8 is a novel therapeutic target for multiple myeloma. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.C.; Taabazuing, C.Y.; Okondo, M.C.; Chui, A.J.; Rao, S.D.; Brown, F.C.; Reed, C.; Peguero, E.; De Stanchina, E.; Kentsis, A.; et al. DPP8/DPP9 inhibitor-induced pyroptosis for treatment of acute myeloid leukemia. Nat. Med. 2018, 24, 1151–1156. [Google Scholar] [CrossRef]
- Dinardo, C.D.; Cortes, J.E. Tosedostat for the treatment of relapsed and refractory acute myeloid leukemia. Expert Opin. Investig. Drugs 2013, 23, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Mawad, R.; Becker, P.S.; Hendrie, P.; Scott, B.; Wood, B.L.; Dean, C.; Sandhu, V.; Deeg, H.J.; Walter, R.; Wang, L.; et al. Phase II study of tosedostat with cytarabine or decitabine in newly diagnosed older patients with acute myeloid leukaemia or high-risk MDS. Br. J. Haematol. 2015, 172, 238–245. [Google Scholar] [CrossRef]
- Cohen, A.D. Myeloma: Next generation immunotherapy. Hematology 2019, 2019, 266–272. [Google Scholar] [CrossRef]
- Gullbo, J.; Wickström, M.; Tullberg, M.; Ehrsson, H.; Lewensohn, R.; Nygren, P.; Luthman, K.; Larsson, R. Activity of Hydrolytic Enzymes in Tumour Cells is a Determinant for Anti-tumour Efficacy of the Melphalan Containing Prodrug J1. J. Drug Target. 2003, 11, 355–363. [Google Scholar] [CrossRef]
- Chauhan, D.; Ray, A.; Viktorsson, K.; Spira, J.; Paba-Prada, C.; Munshi, N.; Richardson, P.; Lewensohn, R.; Anderson, K.C. In Vitro and In Vivo Antitumor Activity of a Novel Alkylating Agent, Melphalan-Flufenamide, against Multiple Myeloma Cells. Clin. Cancer Res. 2013, 19, 3019–3031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, A.; Ravillah, D.; Das, D.S.; Song, Y.; Nordström, E.; Gullbo, J.; Richardson, P.G.; Chauhan, D.; Anderson, K.C. A novel alkylating agent Melflufen induces irreversible DNA damage and cytotoxicity in multiple myeloma cells. Br. J. Haematol. 2016, 174, 397–409. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Koboldt, D.C.; Zhang, Q.; Larson, D.E.; Shen, D.; McLellan, M.D.; Lin, L.; Miller, C.A.; Mardis, E.R.; Ding, L.; Wilson, R.K. VarScan 2: Somatic mutation and copy number alteration discovery in cancer by exome sequencing. Genome Res. 2012, 22, 568–576. [Google Scholar] [CrossRef] [Green Version]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [Green Version]
- Caers, J.; Garderet, L.; Kortüm, K.M.; O’Dwyer, M.E.; Van De Donk, N.W.; Binder, M.; Dold, S.M.; Gay, F.; Corre, J.; Beguin, Y.; et al. European Myeloma Network recommendations on tools for the diagnosis and monitoring of multiple myeloma: What to use and when. Haematology 2018, 103, 1772–1784. [Google Scholar] [CrossRef]
- Razak, K.; Newland, A. The significance of aminopeptidases and haematopoietic cell differentiation. Blood Rev. 1992, 6, 243–250. [Google Scholar] [CrossRef]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Compagnone, M.; Cifaldi, L.; Fruci, D. Regulation of ERAP1 and ERAP2 genes and their disfunction in human cancer. Hum. Immunol. 2019, 80, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Wickström, M.; Nygren, P.; Larsson, R.; Harmenberg, J.; Lindberg, J.; Sjöberg, P.; Jerling, M.; Lehmann, F.; Richardson, P.; Anderson, K.; et al. Melflufen—A peptidase-potentiated alkylating agent in clinical trials. Oncotarget 2017, 8, 66641–66655. [Google Scholar] [CrossRef] [Green Version]
- Matsui, M.; Fowler, J.H.; Walling, L.L. Leucine aminopeptidases: Diversity in structure and function. Biol. Chem. 2006, 387, 1535–1544. [Google Scholar] [CrossRef] [PubMed]
- Beninga, J.; Rock, K.L.; Goldberg, A.L. Interferon-γ Can Stimulate Post-proteasomal Trimming of the N Terminus of an Antigenic Peptide by Inducing Leucine Aminopeptidase. J. Biol. Chem. 1998, 273, 18734–18742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, C.; Zhang, J.; Yang, H.; Peng, L.; Wang, K.; Wang, Y.; Zhao, X.; Liu, H.; Dou, C.; Shi, L.; et al. Leucine aminopeptidase 3 promotes migration and invasion of breast cancer cells through upregulation of fascin and matrix metalloproteinases-2/9 expression. J. Cell. Biochem. 2019, 120, 3611–3620. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Huang, Q.; Qiu, X.; Liu, X.; Sun, G.; Guo, J.; Ding, Z.; Yang, L.; Ban, N.; Tao, T.; et al. LAP3 promotes glioma progression by regulating proliferation, migration and invasion of glioma cells. Int. J. Biol. Macromol. 2015, 72, 1081–1089. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.-Y.; Chen, S.-H.; Shao, B.-F.; Cai, H.-Y.; Zhou, Y.; Zhou, Y.-L.; Xu, A.-B. Expression of leucine aminopeptidase 3 (LAP3) correlates with prognosis and malignant development of human hepatocellular carcinoma (HCC). Int. J. Clin. Exp. Pathol. 2014, 7, 3752–3762. [Google Scholar]
- Zhang, S.; Yang, X.; Shi, H.; Li, M.; Xue, Q.; Ren, H.; Yao, L.; Chen, X.; Zhang, J.; Wang, H. Overexpression of leucine aminopeptidase 3 contributes to malignant development of human esophageal squamous cell carcinoma. J. Mol. Histol. 2014, 45, 283–292. [Google Scholar] [CrossRef]
- Hattori, A.; Tsujimoto, M. Endoplasmic reticulum aminopeptidases: Biochemistry, physiology and pathology. J. Biochem. 2013, 154, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Stratikos, E. Modulating antigen processing for cancer immunotherapy. OncoImmunology 2014, 3, e27568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warthan, M.D.; Washington, S.L.; Franzese, S.E.; Ramus, R.M.; Kim, K.-R.; York, T.P.; Stratikos, E.; Strauss, J.F.; Lee, E.D. The role of endoplasmic reticulum aminopeptidase 2 in modulating immune detection of choriocarcinoma†. Biol. Reprod. 2018, 98, 309–322. [Google Scholar] [CrossRef] [Green Version]
- Toss, M.; Miligy, I.; Gorringe, K.; Mittal, K.; Aneja, R.; Ellis, I.; Green, A.; Rakha, E. Prognostic significance of cathepsin V (CTSV/CTSL2) in breast ductal carcinoma in situ. J. Clin. Pathol. 2019, 73, 76–82. [Google Scholar] [CrossRef]
- Danilov, A.V.; Danilova, O.V.; Brown, J.R.; Rabinowitz, A.; Klein, A.K.; Huber, B.T. Dipeptidyl peptidase 2 apoptosis assay determines the B-cell activation stage and predicts prognosis in chronic lymphocytic leukemia. Exp. Hematol. 2010, 38, 1167–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mele, D.A.; Bista, P.; Baez, D.V.; Huber, B.T. Dipeptidyl Peptidase 2 is an essential survival factor in the regulation of cell quiescence. Cell Cycle 2009, 8, 2425–2434. [Google Scholar] [CrossRef] [Green Version]
- Shah, V.; Johnson, D.C.; Sherborne, A.L.; Ellis, S.; Aldridge, F.M.; Howard-Reeves, J.; Begum, F.; Price, A.; Kendall, J.; Chiecchio, L.; et al. Subclonal TP53 copy number is associated with prognosis in multiple myeloma. Blood 2018, 132, 2465–2469. [Google Scholar] [CrossRef] [Green Version]
- Thakurta, A.; Ortiz, M.; Blecua, P.; Towfic, F.; Corre, J.; Serbina, N.V.; Flynt, E.; Yu, Z.; Yang, Z.; Palumbo, A.; et al. High subclonal fraction of 17p deletion is associated with poor prognosis in multiple myeloma. Blood 2019, 133, 1217–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, K.; Alcivar, A.L.; Ma, J.; Foo, T.K.; Zywea, S.; Mahdi, A.; Huo, Y.; Kensler, T.W.; Gatza, M.L.; Xia, B. NRF2 Induction Supporting Breast Cancer Cell Survival Is Enabled by Oxidative Stress–Induced DPP3–KEAP1 Interaction. Cancer Res. 2017, 77, 2881–2892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šimaga, Š.; Babić, D.; Osmak, M.; Ilić-Forko, J.; Vitale, L.; Miličić, D.; Abramić, M. Dipeptidyl peptidase III in malignant and non-malignant gynaecological tissue. Eur. J. Cancer 1998, 34, 399–405. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, H.; He, Q. Involvement of bleomycin hydrolase and poly(ADP-ribose) polymerase-1 in Ubc9-mediated resistance to chemotherapy agents. Int. J. Oncol. 2016, 50, 223–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jóna, Á.; Miltenyi, Z.; Póliska, S.; Bálint, B.L.; Illes, A. Effect of Bleomycin Hydrolase Gene Polymorphism on Late Pulmonary Complications of Treatment for Hodgkin Lymphoma. PLoS ONE 2016, 11, e0157651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, P.; Li, Y.; Yu, J.; Durr, E.; Krasinska, K.M.; Carver, L.A.; Testa, J.E.; Schnitzer, J.E. Subtractive proteomic mapping of the endothelial surface in lung and solid tumours for tissue-specific therapy. Nat. Cell Biol. 2004, 429, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Perez, I.; Blanco, L.; Sanz, B.; Errarte, P.; Ariz, U.; Beitia, M.; Fernández, A.; Loizate, A.; Candenas, M.L.; Pinto, F.M.; et al. Altered Activity and Expression of Cytosolic Peptidases in Colorectal Cancer. Int. J. Med Sci. 2015, 12, 458–467. [Google Scholar] [CrossRef] [Green Version]
- Larrinaga, G.; Blanco, L.; Errarte, P.; Beitia, M.; Sanz, B.; Perez, I.; Irazusta, A.; Sánchez, C.E.; Santaolalla, F.; Andrés, L.; et al. Altered Peptidase Activities in Thyroid Neoplasia and Hyperplasia. Dis. Markers 2013, 35, 825–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, P.G.; Bringhen, S.; Voorhees, P.; Plesner, T.; Mellqvist, U.-H.; Reeves, B.; Paba-Prada, C.; Zubair, H.; Byrne, C.; Chauhan, D.; et al. Melflufen plus dexamethasone in relapsed and refractory multiple myeloma (O-12-M1): A multicentre, international, open-label, phase 1–2 study. Lancet Haematol. 2020, 7, e395–e407. [Google Scholar] [CrossRef]
- Richardson, P.G.; Oriol, A.; Larocca, A.; Bladé, J.; Cavo, M.; Rodriguez-Otero, P.; Leleu, X.; Nadeem, O.; Hiemenz, J.W.; Hassoun, H.; et al. Melflufen and Dexamethasone in Heavily Pretreated Relapsed and Refractory Multiple Myeloma. J. Clin. Oncol. 2021, 39, 757–767. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient and disease characteristics by disease stage a | |||
|---|---|---|---|
| NDMM (n = 57) | RRMM (n = 83) | Total (n = 140) | |
| Age at diagnosis, years, median (range) | 65 (46–84) | 63 (26–81) | 64 (26–84) |
| Sex, female/male, n | 27/30 | 31/52 | 58/82 |
| Cytogenetics, n (%) | |||
| t(11;14) | 15 (26.3) | 16 (19.3) | 31 (22.1) |
| t(4;14) | 9 (15.8) | 19 (22.9) | 28 (20.0) |
| t(14;16) | 2 (3.5) | 2 (2.4) | 4 (2.9) |
| t(14;20) | 0 | 2 (2.4) | 2 (1.4) |
| del(17p) | 5 (8.8) | 20 (24.1) | 25 (17.9) |
| del(13q)/-13 | 39 (68.4) | 42 (50.6) | 81 (57.9) |
| 1q gain | 18 (31.6) | 46 (55.4) | 64 (45.7) |
| No abnormalities found | 2 (3.5) | 0 | 2 (1.4) |
| International Staging System, n (%) | |||
| 1 | 11 (19.3) | 19 (22.9) | 30 (21.4) |
| 2 | 27 (47.4) | 23 (27.7) | 50 (35.7) |
| 3 | 11 (19.3) | 16 (19.3) | 27 (19.3) |
| Not available | 8 (14.0) | 25 (30.1) | 33 (23.6) |
| Treatment history of relapsed/refractory patients (n = 83) | |||
| Prior treatment, n (%) | Exposed | Refractory | Not exposed |
| Alkylating agents (MEL, CPM) | 63 (75.9) | 16 (19.3) | 4 (4.8) |
| Bortezomib | 44 (53.0) | 28 (33.7) | 11 (13.3) |
| IMiDs | 31 (37.3) | 34 (41.0) | 18 (21.7) |
| Gene Symbol a | Genomic Location | Median log2(RPKM) Expression (n = 122) | Gene Name a | Peptidase/Protein Family |
|---|---|---|---|---|
| LAP3 | 4p15.32 | 5.00 | Leucine aminopeptidase 3 | M |
| ERAP2 | 5q15 | 4.40 | Endoplasmic reticulum aminopeptidase 2 | M |
| METAP2 | 12q22 | 4.22 | Methionyl aminopeptidase 2 | M |
| TPP2 | 13q33.1 | 4.13 | Tripeptidyl peptidase 2 | S |
| DPP7 | 9q34.3 | 4.04 | Dipeptidyl peptidase 7 | S |
| ERAP1 | 5q15 | 4.03 | Endoplasmic reticulum aminopeptidase 1 | M |
| LTA4H | 12q23.1 | 3.44 | Leukotriene A4 hydrolase | M |
| LNPEP | 5q15 | 2.93 | Leucyl and cystinyl aminopeptidase | M |
| XPNPEP1 | 10q25.1 | 2.66 | X-prolyl aminopeptidase 1 | M |
| METAP1 | 4q23 | 2.46 | Methionyl aminopeptidase 1 | M |
| DPP3 | 11q13.2 | 2.28 | Dipeptidyl peptidase 3 | M |
| DPP8 | 15q22.31 | 2.21 | Dipeptidyl peptidase 8 | S |
| NPEPPS | 17q21.32 | 2.12 | Aminopeptidase puromycin sensitive | M |
| BLMH | 17q11.2 | 1.83 | Bleomycin hydrolase | C |
| JMJD7 | 15q15.1 | 1.78 | Jumonji domain containing 7 | * |
| RNPEP | 1q32.1 | 1.76 | Arginyl aminopeptidase | M |
| PGPEP1 | 19p13.11 | 1.72 | Pyroglutamyl-peptidase I | C |
| TPP1 | 11p15.4 | 1.59 | Tripeptidyl peptidase 1 | S |
| PEPD | 19q13.11 | 1.41 | Peptidase D | M |
| DNPEP | 2q35 | 1.16 | Aspartyl aminopeptidase | M |
| DPP9 | 19p13.3 | 1.16 | Dipeptidyl peptidase 9 | S |
| METAP1D | 2q31.1 | 0.38 | Methionyl aminopeptidase type 1D, mitochondrial | M |
| NPEPL1 | 20q13.32 | 0.37 | Aminopeptidase like 1 | M |
| CTSH | 15q25.1 | 0.36 | Cathepsin H | C |
| XPNPEP3 | 22q13.2 | −0.04 | X-prolyl aminopeptidase 3 | M |
| RNPEPL1 | 2q37.3 | −0.1 | Arginyl aminopeptidase like 1 | M |
| AOPEP | 9q22.32 | −0.38 | Aminopeptidase O (putative) | M |
| KDM8 | 16p12.1 | −1.3 | Lysine demethylase 8 | * |
| ANPEP | 15q26.1 | −2.13 | Alanyl aminopeptidase, membrane | M |
| ENPEP | 4q25 | −2.15 | Glutamyl Aminopeptidase | M |
| MMP14 | 14q11.2 | −2.83 | Matrix Metallopeptidase 14 | M |
| DPP4 | 2q24.2 | −2.84 | Dipeptidyl Peptidase 4 | S |
| NAALADL1 | 11q13.1 | −3.39 | N-Acetylated Alpha-Linked Acidic Dipeptidase Like 1 | M |
| AMZ1 | 7p22.3 | −3.6 | Archaelysin Family Metallopeptidase 1 | M |
| LVRN | 5q23.1 | −3.64 | Laeverin | M |
| CTSV | 9q22.33 | −4.39 | Cathepsin V | C |
| XPNPEP2 | Xq26.1 | −6.15 | X-Prolyl Aminopeptidase 2 | M |
| F11 | 4q35.2 | −6.22 | Coagulation Factor XI | S |
| TRHDE | 12q21.1 | −6.57 | Thyrotropin Releasing Hormone Degrading Enzyme | M |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miettinen, J.J.; Kumari, R.; Traustadottir, G.A.; Huppunen, M.-E.; Sergeev, P.; Majumder, M.M.; Schepsky, A.; Gudjonsson, T.; Lievonen, J.; Bazou, D.; et al. Aminopeptidase Expression in Multiple Myeloma Associates with Disease Progression and Sensitivity to Melflufen. Cancers 2021, 13, 1527. https://doi.org/10.3390/cancers13071527
Miettinen JJ, Kumari R, Traustadottir GA, Huppunen M-E, Sergeev P, Majumder MM, Schepsky A, Gudjonsson T, Lievonen J, Bazou D, et al. Aminopeptidase Expression in Multiple Myeloma Associates with Disease Progression and Sensitivity to Melflufen. Cancers. 2021; 13(7):1527. https://doi.org/10.3390/cancers13071527
Chicago/Turabian StyleMiettinen, Juho J., Romika Kumari, Gunnhildur Asta Traustadottir, Maiju-Emilia Huppunen, Philipp Sergeev, Muntasir M. Majumder, Alexander Schepsky, Thorarinn Gudjonsson, Juha Lievonen, Despina Bazou, and et al. 2021. "Aminopeptidase Expression in Multiple Myeloma Associates with Disease Progression and Sensitivity to Melflufen" Cancers 13, no. 7: 1527. https://doi.org/10.3390/cancers13071527
APA StyleMiettinen, J. J., Kumari, R., Traustadottir, G. A., Huppunen, M. -E., Sergeev, P., Majumder, M. M., Schepsky, A., Gudjonsson, T., Lievonen, J., Bazou, D., Dowling, P., O`Gorman, P., Slipicevic, A., Anttila, P., Silvennoinen, R., Nupponen, N. N., Lehmann, F., & Heckman, C. A. (2021). Aminopeptidase Expression in Multiple Myeloma Associates with Disease Progression and Sensitivity to Melflufen. Cancers, 13(7), 1527. https://doi.org/10.3390/cancers13071527