
NKL-Code in Normal and Aberrant Hematopoiesis

Abstract
:Simple Summary
Abstract
1. Hematopoiesis
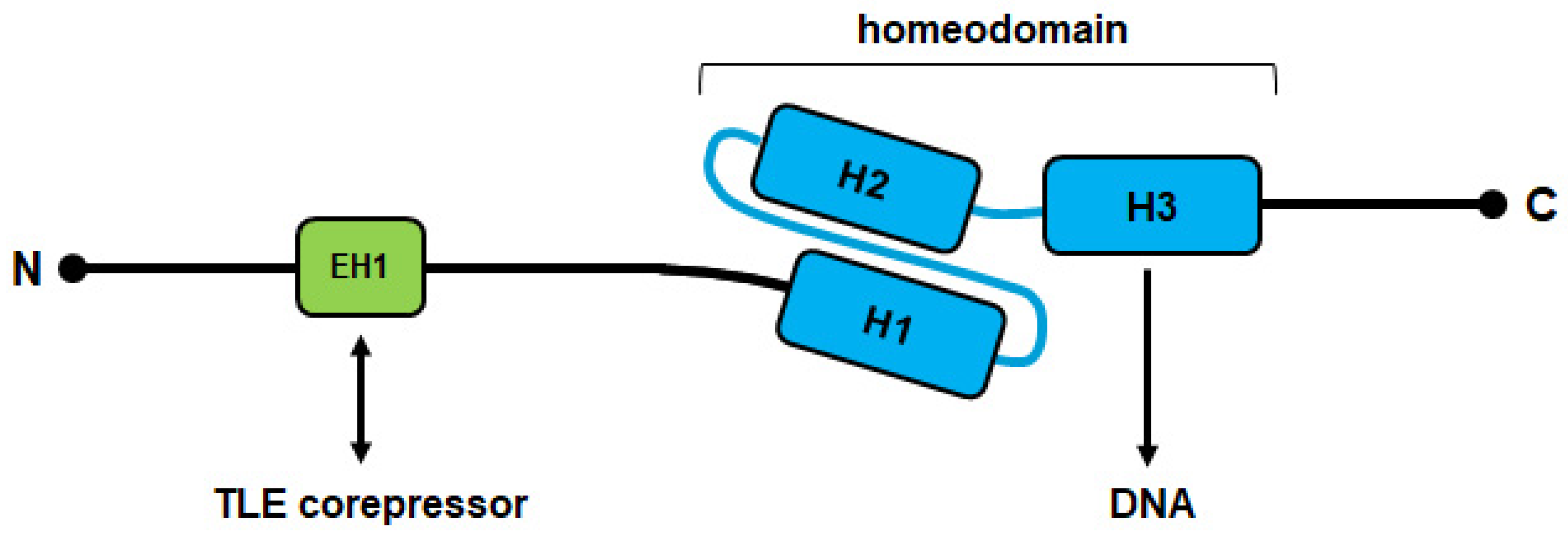
2. Classification of Homeobox Genes
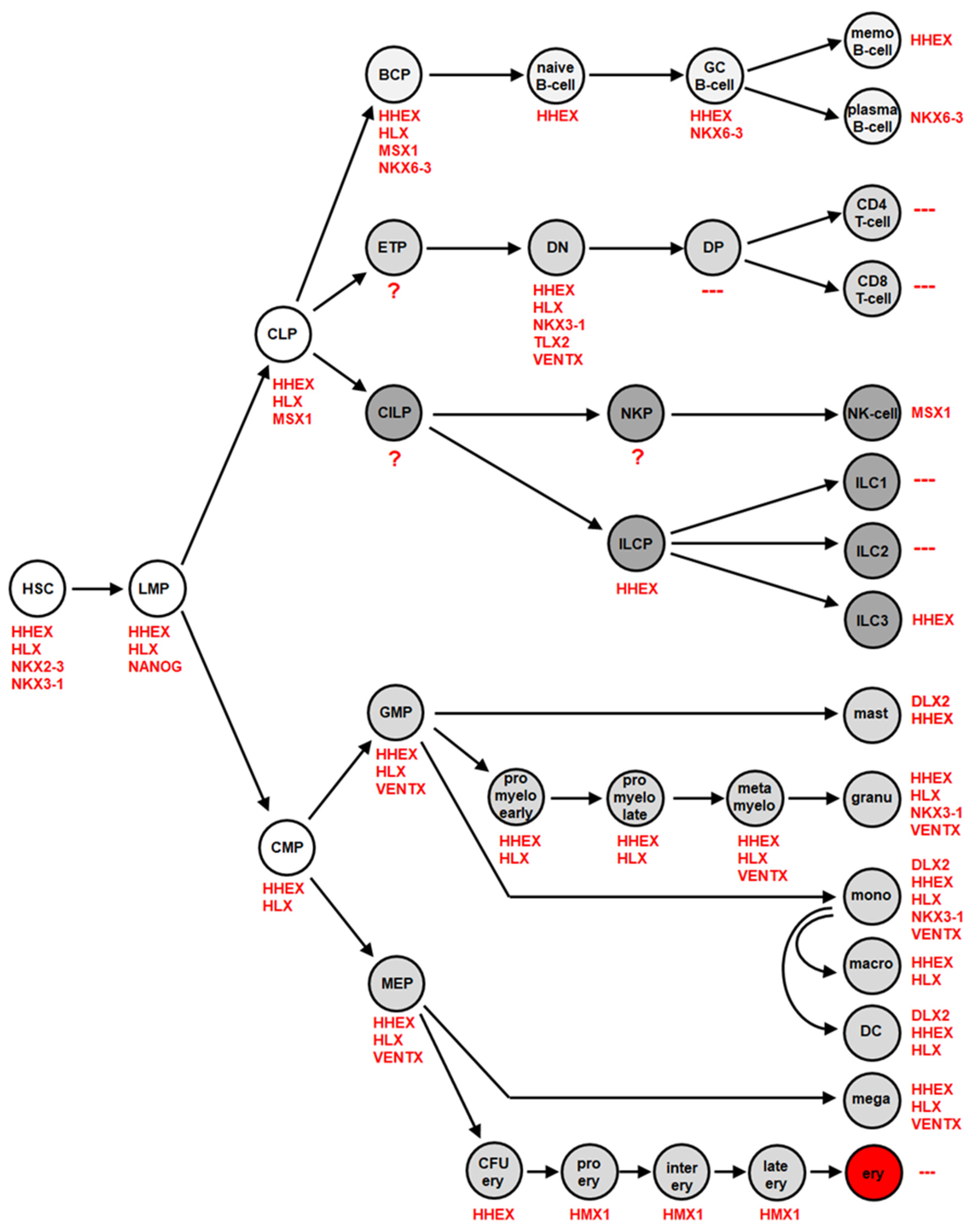
3. NKL-Code in Hematopoiesis
4. NKL Homeobox Genes in Non-Hematopoietic Tissues
5. Deregulated NKL Homeobox Genes in Hematopoietic Malignancies
6. Tumor Cell Lines as Models for Deregulated NKL Homeobox Genes
Funding
Acknowledgments
Conflicts of Interest
References
- Uhlen, M.; Karlsson, M.J.; Zhong, W.; Tebani, A.; Pou, C.; Mikes, J.; Lakshmikanth, T.; Forsström, B.; Edfors, F.; Odeberg, J.; et al. A genome-wide transcriptomic analysis of protein-coding genes in human blood cells. Science 2019, 366, eaax9198. [Google Scholar] [CrossRef] [PubMed]
- Liggett, L.A.; Sankaran, V.G. Unraveling Hematopoiesis through the lens of genomics. Cell 2020, 182, 1384–1400. [Google Scholar] [CrossRef] [PubMed]
- Kucinski, I.; Wilson, N.K.; Hannah, R.; Kinston, S.J.; Cauchy, P.; Lenaerts, A.; Grosschedl, R.; Göttgens, B. Interactions between lineage-associated transcription factors govern haematopoietic progenitor states. EMBO J. 2020, 39, e104983. [Google Scholar] [CrossRef]
- Rothenberg, E.V. Transcriptional control of early T and B cell developmental choices. Annu. Rev. Immunol. 2014, 32, 283–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souroullas, G.P.; Salmon, J.M.; Sablitzky, F.; Curtis, D.J.; Goodell, M.A. Adult hematopoietic stem and progenitor cells require either Lyl1 or Scl for survival. Cell Stem Cell 2009, 4, 180–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crispino, J.D.; Horwitz, M.S. GATA factor mutations in hematologic disease. Blood 2017, 129, 2103–2110. [Google Scholar] [CrossRef]
- Van de Walle, I.; Dolens, A.C.; Durinck, K.; De Mulder, K.; Van Loocke, W.; Damle, S.; Waegemans, E.; De Medts, J.; Velghe, I.; De Smedt, M.; et al. GATA3 induces human T-cell commitment by restraining Notch activity and repressing NK-cell fate. Nat. Commun. 2016, 7, 11171. [Google Scholar] [CrossRef] [Green Version]
- Boller, S.; Grosschedl, R. The regulatory network of B-cell differentiation: A focused view of early B-cell factor 1 function. Immunol. Rev. 2014, 261, 102–115. [Google Scholar] [CrossRef]
- Avram, D.; Califano, D. The multifaceted roles of Bcl11b in thymic and peripheral T cells: Impact on immune diseases. J. Immunol. 2014, 193, 2059–2065. [Google Scholar] [CrossRef] [Green Version]
- Kostrzewski, T.; Borg, A.J.; Meng, Y.; Filipovic, I.; Male, V.; Wack, A.; DiMaggio, P.A.; Brady, H.J.M. Multiple levels of control determine how E4bp4/Nfil3 regulates NK cell development. J. Immunol. 2018, 200, 1370–1381. [Google Scholar] [CrossRef]
- Tanriver, Y.; Diefenbach, A. Transcription factors controlling development and function of innate lymphoid cells. Int. Immunol. 2014, 26, 119–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liew, C.W.; Rand, K.D.; Simpson, R.J.; Yung, W.W.; Mansfield, R.E.; Crossley, M.; Proetorius-Ibba, M.; Nerlov, C.; Poulsen, F.M.; Mackay, J.P. Molecular analysis of the interaction between the hematopoietic master transcription factors GATA-1 and PU.1. J. Biol. Chem. 2006, 281, 28296–28306. [Google Scholar] [CrossRef] [Green Version]
- Vaquerizas, J.M.; Kummerfeld, S.K.; Teichmann, S.A.; Luscombe, N.M. A census of human transcription factors: Function, expression and evolution. Nat. Rev. Genet. 2009, 10, 252–263. [Google Scholar] [CrossRef]
- Bürglin, T.R.; Affolter, M. Homeodomain proteins: An update. Chromosoma 2016, 125, 497–521. [Google Scholar] [CrossRef] [Green Version]
- Gehring, W.J.; Müller, M.; Affolter, M.; Percival-Smith, A.; Billeter, M.; Qian, Y.Q.; Otting, G.; Wüthrich, K. The structure of the homeodomain and its functional implications. Trends Genet. 1990, 6, 323–329. [Google Scholar] [CrossRef]
- Holland, P.W.; Booth, H.A.; Bruford, E.A. Classification and nomenclature of all human homeobox genes. BMC Biol. 2007, 5, 47. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Nirenberg, M. Drosophila NK-homeobox genes. Proc. Natl. Acad. Sci. USA 1989, 86, 7716–7720. [Google Scholar] [CrossRef] [Green Version]
- Pollard, S.L.; Holland, P.W. Evidence for 14 homeobox gene clusters in human genome ancestry. Curr. Biol. 2000, 10, 1059–1062. [Google Scholar] [CrossRef] [Green Version]
- Harvey, R.P. NK-2 homeobox genes and heart development. Dev. Biol. 1996, 178, 203–216. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.T.; Jaynes, J.B. A conserved region of engrailed, shared among all en-, gsc-, Nk1-, Nk2- and msh-class homeoproteins, mediates active transcriptional repression in vivo. Development 1996, 122, 3141–3150. [Google Scholar]
- Jennings, B.H.; Ish-Horowicz, D. The Groucho/TLE/Grg family of transcriptional co-repressors. Genome Biol. 2008, 9, 205. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Fernàndez, J. The genesis and evolution of homeobox gene clusters. Nat. Rev. Genet. 2005, 6, 881–892. [Google Scholar] [CrossRef]
- Jagla, K.; Bellard, M.; Frasch, M. A cluster of Drosophila homeobox genes involved in mesoderm differentiation programs. Bioessays 2001, 23, 125–133. [Google Scholar] [CrossRef]
- Lints, T.J.; Parsons, L.M.; Hartley, L.; Lyons, I.; Harvey, R.P. Nkx-2.5: A novel murine homeobox gene expressed in early heart progenitor cells and their myogenic descendants. Development 1993, 119, 419–431. [Google Scholar]
- Lazzaro, D.; Price, M.; de Felice, M.; Di Lauro, R. The transcription factor TTF-1 is expressed at the onset of thyroid and lung morphogenesis and in restricted regions of the foetal brain. Development 1991, 113, 1093–1104. [Google Scholar]
- Nagel, S.; Pommerenke, C.; Scherr, M.; Meyer, C.; Kaufmann, M.; Battmer, K.; MacLeod, R.A.; Drexler, H.G. NKL homeobox gene activities in hematopoietic stem cells, T-cell development and T-cell leukemia. PLoS ONE 2017, 12, e0171164. [Google Scholar] [CrossRef]
- Nagel, S.; MacLeod, R.A.F.; Meyer, C.; Kaufmann, M.; Drexler, H.G. NKL homeobox gene activities in B-cell development and lymphomas. PLoS ONE 2018, 13, e0205537. [Google Scholar] [CrossRef] [PubMed]
- Nagel, S.; Pommerenke, C.; Meyer, C.; Kaufmann, M.; MacLeod, R.A.F.; Drexler, H.G. NKL homeobox gene MSX1 acts like a tumor suppressor in NK-cell leukemia. Oncotarget 2017, 8, 66815–66832. [Google Scholar] [CrossRef] [Green Version]
- Nagel, S.; Pommerenke, C.; MacLeod, R.A.F.; Meyer, C.; Kaufmann, M.; Drexler, H.G. The NKL-code for innate lymphoid cells reveals deregulated expression of NKL homeobox genes HHEX and HLX in anaplastic large cell lymphoma (ALCL). Oncotarget 2020, 11, 3208–3226. [Google Scholar] [CrossRef]
- Nagel, S.; Scherr, M.; MacLeod, R.A.F.; Pommerenke, C.; Koeppel, M.; Meyer, C.; Kaufmann, M.; Dallmann, I.; Drexler, H.G. NKL homeobox gene activities in normal and malignant myeloid cells. PLoS ONE 2019, 14, e0226212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farlik, M.; Halbritter, F.; Müller, F.; Choudry, F.A.; Ebert, P.; Klughammer, J.; Farrow, S.; Santoro, A.; Ciaurro, V.; Mathur, A.; et al. DNA methylation dynamics of human hematopoietic stem cell differentiation. Cell Stem Cell 2016, 19, 808–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, J.D.; Lints, T.; Jenkins, N.A.; Copeland, N.G.; Strasser, A.; Harvey, R.P.; Adams, J.M. Novel murine homeo box gene on chromosome 1 expressed in specific hematopoietic lineages and during embryogenesis. Genes Dev. 1991, 5, 509–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crompton, M.R.; Bartlett, T.J.; MacGregor, A.D.; Manfioletti, G.; Buratti, E.; Giancotti, V.; Goodwin, G.H. Identification of a novel vertebrate homeobox gene expressed in haematopoietic cells. Nucleic Acids Res. 1992, 20, 5661–5667. [Google Scholar] [CrossRef] [Green Version]
- Bedford, F.K.; Ashworth, A.; Enver, T.; Wiedemann, L.M. HEX: A novel homeobox gene expressed during haematopoiesis and conserved between mouse and human. Nucleic Acids Res. 1993, 21, 1245–1249. [Google Scholar] [CrossRef] [Green Version]
- Manfioletti, G.; Gattei, V.; Buratti, E.; Rustighi, A.; De Iuliis, A.; Aldinucci, D.; Goodwin, G.H.; Pinto, A. Differential expression of a novel proline-rich homeobox gene (Prh) in human hematolymphopoietic cells. Blood 1995, 85, 1237–1245. [Google Scholar] [CrossRef] [PubMed]
- Mack, D.L.; Leibowitz, D.S.; Cooper, S.; Ramsey, H.; Broxmeyer, H.E.; Hromas, R. Down-regulation of the myeloid homeobox protein Hex is essential for normal T-cell development. Immunology 2002, 107, 444–451. [Google Scholar] [CrossRef]
- Jackson, J.T.; Nasa, C.; Shi, W.; Huntington, N.D.; Bogue, C.W.; Alexander, W.S.; McCormack, M.P. A crucial role for the homeodomain transcription factor Hhex in lymphopoiesis. Blood 2015, 125, 803–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laidlaw, B.J.; Duan, L.; Xu, Y.; Vazquez, S.E.; Cyster, J.G. The transcription factor Hhex cooperates with the corepressor Tle3 to promote memory B cell development. Nat. Immunol. 2020, 21, 1082–1093. [Google Scholar] [CrossRef]
- Allen, J.D.; Harris, A.W.; Bath, M.L.; Strasser, A.; Scollay, R.; Adams, J.M. Perturbed development of T and B cells in mice expressing an Hlx homeobox transgene. J. Immunol. 1995, 154, 1531–1542. [Google Scholar]
- Gentner, E.; Vegi, N.M.; Mulaw, M.A.; Mandal, T.; Bamezai, S.; Claus, R.; Tasdogan, A.; Quintanilla-Martinez, L.; Grunenberg, A.; Döhner, K.; et al. VENTX induces expansion of primitive erythroid cells and contributes to the development of acute myeloid leukemia in mice. Oncotarget 2016, 7, 86889–86901. [Google Scholar] [CrossRef] [Green Version]
- Lewis, K.E. How do genes regulate simple behaviours? Understanding how different neurons in the vertebrate spinal cord are genetically specified. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 45–66. [Google Scholar] [CrossRef] [PubMed]
- Hunt, P.; Gulisano, M.; Cook, M.; Sham, M.H.; Faiella, A.; Wilkinson, D.; Boncinelli, E.; Krumlauf, R. A distinct Hox code for the branchial region of the vertebrate head. Nature 1991, 353, 861–864. [Google Scholar] [CrossRef]
- Depew, M.J.; Simpson, C.A.; Morasso, M.; Rubenstein, J.L. Reassessing the Dlx code: The genetic regulation of branchial arch skeletal pattern and development. J. Anat. 2005, 207, 501–561. [Google Scholar] [CrossRef] [PubMed]
- Streit, A. The cranial sensory nervous system: Specification of sensory progenitors and placodes. In StemBook; Harvard Stem Cell Institute: Cambridge, MA, USA, 2008. [Google Scholar]
- Nagel, S.; Pommerenke, C.; Meyer, C.; MacLeod, R.A.F.; Drexler, H.G. Establishment of the TALE-code reveals aberrantly activated homeobox gene PBX1 in Hodgkin lymphoma. PLoS ONE 2021, 16, e0246603. [Google Scholar] [CrossRef] [PubMed]
- Bogue, C.W.; Ganea, G.R.; Sturm, E.; Ianucci, R.; Jacobs, H.C. Hex expression suggests a role in the development and function of organs derived from foregut endoderm. Dev. Dyn. 2000, 219, 84–89. [Google Scholar] [CrossRef]
- Hentsch, B.; Lyons, I.; Li, R.; Hartley, L.; Lints, T.J.; Adams, J.M.; Harvey, R.P. Hlx homeo box gene is essential for an inductive tissue interaction that drives expansion of embryonic liver and gut. Genes Dev. 1996, 10, 70–79. [Google Scholar] [CrossRef] [Green Version]
- Lints, T.J.; Hartley, L.; Parsons, L.M.; Harvey, R.P. Mesoderm-specific expression of the divergent homeobox gene Hlx during murine embryogenesis. Dev. Dyn. 1996, 205, 457–470. [Google Scholar] [CrossRef]
- Chambers, I.; Colby, D.; Robertson, M.; Nichols, J.; Lee, S.; Tweedie, S.; Smith, A. Functional expression cloning of Nanog, a pluripotency sustaining factor in embryonic stem cells. Cell 2003, 113, 643–655. [Google Scholar] [CrossRef] [Green Version]
- Mitsui, K.; Tokuzawa, Y.; Itoh, H.; Segawa, K.; Murakami, M.; Takahashi, K.; Maruyama, M.; Maeda, M.; Yamanaka, S. The homeoprotein Nanog is required for maintenance of pluripotency in mouse epiblast and ES cells. Cell 2003, 113, 631–642. [Google Scholar] [CrossRef] [Green Version]
- Wernig, M.; Meissner, A.; Foreman, R.; Brambrink, T.; Ku, M.; Hochedlinger, K.; Bernstein, B.E.; Jaenisch, R. In vitro reprogramming of fibroblasts into a pluripotent ES-cell-like state. Nature 2007, 448, 318–324. [Google Scholar] [CrossRef]
- Pan, G.; Thomson, J.A. Nanog and transcriptional networks in embryonic stem cell pluripotency. Cell Res. 2007, 17, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Andrianakos, R.; Yang, Y.; Liu, C.; Lu, W. Kruppel-like factor 4 (Klf4) prevents embryonic stem (ES) cell differentiation by regulating Nanog gene expression. J. Biol. Chem. 2010, 285, 9180–9189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grocott, T.; Tambalo, M.; Streit, A. The peripheral sensory nervous system in the vertebrate head: A gene regulatory perspective. Dev. Biol. 2012, 370, 3–23. [Google Scholar] [CrossRef]
- Scerbo, P.; Monsoro-Burq, A.H. The vertebrate-specific VENTX/NANOG gene empowers neural crest with ectomesenchyme potential. Sci. Adv. 2020, 6, eaaz1469. [Google Scholar] [CrossRef] [PubMed]
- Mair, B.; Tomic, J.; Masud, S.N.; Tonge, P.; Weiss, A.; Usaj, M.; Tong, A.H.Y.; Kwan, J.J.; Brown, K.R.; Titus, E.; et al. Essential gene profiles for human pluripotent stem cells identify uncharacterized genes and substrate dependencies. Cell Rep. 2019, 27, 599–615. [Google Scholar] [CrossRef] [Green Version]
- Ramos, C.; Robert, B. msh/Msx gene family in neural development. Trends Genet. 2005, 21, 624–632. [Google Scholar] [CrossRef]
- Vastardis, H.; Karimbux, N.; Guthua, S.W.; Seidman, J.G.; Seidman, C.E. A human MSX1 homeodomain missense mutation causes selective tooth agenesis. Nat. Genet. 1996, 13, 417–421. [Google Scholar] [CrossRef]
- Liang, J.; Von den Hoff, J.; Lange, J.; Ren, Y.; Bian, Z.; Carels, C.E. MSX1 mutations and associated disease phenotypes: Genotype-phenotype relations. Eur. J. Hum. Genet. 2016, 24, 1663–1670. [Google Scholar] [CrossRef] [Green Version]
- Lan, Y.; Jia, S.; Jiang, R. Molecular patterning of the mammalian dentition. Semin. Cell Dev. Biol. 2014, 25–26, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Song, Y.; Zhao, X.; Zhang, X.; Fermin, C.; Chen, Y. Rescue of cleft palate in Msx1-deficient mice by transgenic Bmp4 reveals a network of BMP and Shh signaling in the regulation of mammalian palatogenesis. Development 2002, 129, 4135–4146. [Google Scholar]
- Odelberg, S.J.; Kollhoff, A.; Keating, M.T. Dedifferentiation of mammalian myotubes induced by msx1. Cell 2000, 103, 1099–1109. [Google Scholar] [CrossRef] [Green Version]
- Abate-Shen, C.; Shen, M.M.; Gelmann, E. Integrating differentiation and cancer: The Nkx3.1 homeobox gene in prostate organogenesis and carcinogenesis. Differentiation 2008, 76, 717–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alasti, F.; Van Camp, G. Genetics of microtia and associated syndromes. J. Med. Genet. 2009, 46, 361–369. [Google Scholar] [CrossRef]
- Brown, S.T.; Wang, J.; Groves, A.K. Dlx gene expression during chick inner ear development. J. Comp. Neurol. 2005, 483, 48–65. [Google Scholar] [CrossRef] [PubMed]
- O’Neil, J.; Look, A.T. Mechanisms of transcription factor deregulation in lymphoid cell transformation. Oncogene 2007, 26, 6838–6849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eswaran, J.; Sinclair, P.; Heidenreich, O.; Irving, J.; Russell, L.J.; Hall, A.; Calado, D.P.; Harrison, C.J.; Vormoor, J. The pre-B-cell receptor checkpoint in acute lymphoblastic leukaemia. Leukemia 2015, 29, 1623–1631. [Google Scholar] [CrossRef] [PubMed]
- Tomolonis, J.A.; Agarwal, S.; Shohet, J.M. Neuroblastoma pathogenesis: Deregulation of embryonic neural crest development. Cell Tissue Res. 2018, 372, 245–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrando, A.A.; Neuberg, D.S.; Staunton, J.; Loh, M.L.; Huard, C.; Raimondi, S.C.; Behm, F.G.; Pui, C.H.; Downing, J.R.; Gilliland, D.G.; et al. Gene expression signatures define novel oncogenic pathways in T cell acute lymphoblastic leukemia. Cancer Cell 2002, 1, 75–87. [Google Scholar] [CrossRef] [Green Version]
- Hatano, M.; Roberts, C.W.; Minden, M.; Crist, W.M.; Korsmeyer, S.J. Deregulation of a homeobox gene, HOX11, by the t(10;14) in T cell leukemia. Science 1991, 253, 79–82. [Google Scholar] [CrossRef]
- Brendolan, A.; Ferretti, E.; Salsi, V.; Moses, K.; Quaggin, S.; Blasi, F.; Cleary, M.L.; Selleri, L. A Pbx1-dependent genetic and transcriptional network regulates spleen ontogeny. Development 2005, 132, 3113–3126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernard, O.A.; Busson-LeConiat, M.; Ballerini, P.; Mauchauffé, M.; Della Valle, V.; Monni, R.; Nguyen Khac, F.; Mercher, T.; Penard-Lacronique, V.; Pasturaud, P.; et al. A new recurrent and specific cryptic translocation, t(5;14)(q35;q32), is associated with expression of the Hox11L2 gene in T acute lymphoblastic leukemia. Leukemia 2001, 15, 1495–1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meijerink, J.P. Genetic rearrangements in relation to immunophenotype and outcome in T-cell acute lymphoblastic leukaemia. Best Pract. Res. Clin. Haematol. 2010, 23, 307–318. [Google Scholar] [CrossRef] [PubMed]
- De Keersmaecker, K.; Real, P.J.; Gatta, G.D.; Palomero, T.; Sulis, M.L.; Tosello, V.; Van Vlierberghe, P.; Barnes, K.; Castillo, M.; Sole, X.; et al. The TLX1 oncogene drives aneuploidy in T cell transformation. Nat. Med. 2010, 16, 1321–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dadi, S.; Le Noir, S.; Payet-Bornet, D.; Lhermitte, L.; Zacarias-Cabeza, J.; Bergeron, J.; Villarèse, P.; Vachez, E.; Dik, W.A.; Millien, C.; et al. TLX homeodomain oncogenes mediate T cell maturation arrest in T-ALL via interaction with ETS1 and suppression of TCRα gene expression. Cancer Cell 2012, 21, 563–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagel, S.; Kaufmann, M.; Drexler, H.G.; MacLeod, R.A. The cardiac homeobox gene NKX2-5 is deregulated by juxtaposition with BCL11B in pediatric T-ALL cell lines via a novel t(5;14)(q35.1;q32.2). Cancer Res. 2003, 63, 5329–5334. [Google Scholar] [PubMed]
- Villarese, P.; Lours, C.; Trinquand, A.; Le Noir, S.; Belhocine, M.; Lhermitte, L.; Cieslak, A.; Tesio, M.; Petit, A.; LeLorch, M.; et al. TCRα rearrangements identify a subgroup of NKL-deregulated adult T-ALLs associated with favorable outcome. Leukemia 2018, 32, 61–71. [Google Scholar] [CrossRef]
- Przybylski, G.K.; Dik, W.A.; Grabarczyk, P.; Wanzeck, J.; Chudobska, P.; Jankowski, K.; von Bergh, A.; van Dongen, J.J.; Schmidt, C.A.; Langerak, A.W. The effect of a novel recombination between the homeobox gene NKX2-5 and the TRD locus in T-cell acute lymphoblastic leukemia on activation of the NKX2-5 gene. Haematologica 2006, 91, 317–321. [Google Scholar]
- Skerjanc, I.S.; Petropoulos, H.; Ridgeway, A.G.; Wilton, S. Myocyte enhancer factor 2C and Nkx2-5 up-regulate each other’s expression and initiate cardiomyogenesis in P19 cells. J. Biol. Chem. 1998, 273, 34904–34910. [Google Scholar] [CrossRef] [Green Version]
- Nagel, S.; Meyer, C.; Quentmeier, H.; Kaufmann, M.; Drexler, H.G.; MacLeod, R.A. MEF2C is activated by multiple mechanisms in a subset of T-acute lymphoblastic leukemia cell lines. Leukemia 2008, 22, 600–607. [Google Scholar] [CrossRef] [Green Version]
- Homminga, I.; Pieters, R.; Langerak, A.W.; de Rooi, J.J.; Stubbs, A.; Verstegen, M.; Vuerhard, M.; Buijs-Gladdines, J.; Kooi, C.; Klous, P.; et al. Integrated transcript and genome analyses reveal NKX2-1 and MEF2C as potential oncogenes in T cell acute lymphoblastic leukemia. Cancer Cell 2011, 19, 484–497. [Google Scholar] [CrossRef]
- Nagel, S.; Venturini, L.; Meyer, C.; Kaufmann, M.; Scherr, M.; Drexler, H.G.; Macleod, R.A. Transcriptional deregulation of oncogenic myocyte enhancer factor 2C in T-cell acute lymphoblastic leukemia. Leuk. Lymphoma 2011, 52, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Nagel, S.; Ehrentraut, S.; Tomasch, J.; Quentmeier, H.; Meyer, C.; Kaufmann, M.; Drexler, H.G.; MacLeod, R.A. Ectopic expression of homeobox gene NKX2-1 in diffuse large B-cell lymphoma is mediated by aberrant chromatin modifications. PLoS ONE 2013, 8, e61447. [Google Scholar] [CrossRef]
- Kusy, S.; Gerby, B.; Goardon, N.; Gault, N.; Ferri, F.; Gérard, D.; Armstrong, F.; Ballerini, P.; Cayuela, J.M.; Baruchel, A.; et al. NKX3.1 is a direct TAL1 target gene that mediates proliferation of TAL1-expressing human T cell acute lymphoblastic leukemia. J. Exp. Med. 2010, 207, 2141–2156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soulier, J.; Clappier, E.; Cayuela, J.M.; Regnault, A.; García-Peydró, M.; Dombret, H.; Baruchel, A.; Toribio, M.L.; Sigaux, F. HOXA genes are included in genetic and biologic networks defining human acute T-cell leukemia (T-ALL). Blood 2005, 106, 274–286. [Google Scholar] [CrossRef]
- Nagel, S.; Ehrentraut, S.; Tomasch, J.; Lienenklaus, S.; Schneider, B.; Geffers, R.; Meyer, C.; Kaufmann, M.; Drexler, H.G.; MacLeod, R.A. Transcriptional activation of prostate specific homeobox gene NKX3-1 in subsets of T-cell lymphoblastic leukemia (T-ALL). PLoS ONE 2012, 7, e40747. [Google Scholar] [CrossRef] [Green Version]
- Nagel, S.; Meyer, C.; Kaufmann, M.; Zaborski, M.; MacLeod, R.A.F.; Drexler, H.G. Aberrant activity of NKL homeobox gene NKX3-2 in a T-ALL subset. PLoS ONE 2018, 13, e0197194. [Google Scholar] [CrossRef]
- Nagel, S.; Ehrentraut, S.; Meyer, C.; Kaufmann, M.; Drexler, H.G.; MacLeod, R.A. Oncogenic deregulation of NKL homeobox gene MSX1 in mantle cell lymphoma. Leuk. Lymphoma 2014, 55, 1893–1903. [Google Scholar] [CrossRef]
- Nagel, S.; Ehrentraut, S.; Meyer, C.; Kaufmann, M.; Drexler, H.G.; MacLeod, R.A. Repressed BMP signaling reactivates NKL homeobox gene MSX1 in a T-ALL subset. Leuk. Lymphoma 2015, 56, 480–491. [Google Scholar] [CrossRef]
- Nagel, S.; Pommerenke, C.; Meyer, C.; Kaufmann, M.; Drexler, H.G.; MacLeod, R.A. Deregulation of polycomb repressor complex 1 modifier AUTS2 in T-cell leukemia. Oncotarget 2016, 7, 45398–45413. [Google Scholar] [CrossRef] [Green Version]
- Nagel, S.; Pommerenke, C.; Meyer, C.; Kaufmann, M.; MacLeod, R.A.F.; Drexler, H.G. Aberrant expression of NKL homeobox gene HLX in Hodgkin lymphoma. Oncotarget 2018, 9, 14338–14353. [Google Scholar] [CrossRef] [Green Version]
- Nagel, S.; Uphoff, C.C.; Dirks, W.G.; Pommerenke, C.; Meyer, C.; Drexler, H.G. Epstein-Barr virus (EBV) activates NKL homeobox gene HLX in DLBCL. PLoS ONE 2019, 14, e0216898. [Google Scholar] [CrossRef] [PubMed]
- Crescenzo, R.; Abate, F.; Lasorsa, E.; Tabbo’, F.; Gaudiano, M.; Chiesa, N.; Di Giacomo, F.; Spaccarotella, E.; Barbarossa, L.; Ercole, E.; et al. T-Cell Project: Prospective Collection of Data in Patients with Peripheral T-Cell Lymphoma and the AIRC 5xMille Consortium “Genetics-Driven Targeted Management of Lymphoid Malignancies”. Convergent mutations and kinase fusions lead to oncogenic STAT3 activation in anaplastic large cell lymphoma. Cancer Cell 2015, 27, 516–532. [Google Scholar] [PubMed] [Green Version]
- Robles, E.F.; Mena-Varas, M.; Barrio, L.; Merino-Cortes, S.V.; Balogh, P.; Du, M.Q.; Akasaka, T.; Parker, A.; Roa, S.; Panizo, C.; et al. Homeobox NKX2-3 promotes marginal-zone lymphomagenesis by activating B-cell receptor signalling and shaping lymphocyte dynamics. Nat. Commun. 2016, 7, 11889. [Google Scholar] [CrossRef]
- Nagel, S.; Pommerenke, C.; MacLeod, R.A.F.; Meyer, C.; Kaufmann, M.; Fähnrich, S.; Drexler, H.G. Deregulated expression of NKL homeobox genes in T-cell lymphomas. Oncotarget 2019, 10, 3227–3247. [Google Scholar] [CrossRef] [PubMed]
- Garg, N.; Po, A.; Miele, E.; Campese, A.F.; Begalli, F.; Silvano, M.; Infante, P.; Capalbo, C.; De Smaele, E.; Canettieri, G.; et al. microRNA-17-92 cluster is a direct Nanog target and controls neural stem cell through Trp53inp1. EMBO J. 2013, 32, 2819–2832. [Google Scholar] [CrossRef] [PubMed]
- Starkova, J.; Gadgil, S.; Qiu, Y.H.; Zhang, N.; Hermanova, I.; Kornblau, S.M.; Drabkin, H.A. Up-regulation of homeodomain genes, DLX1 and DLX2, by FLT3 signaling. Haematologica 2011, 96, 820–828. [Google Scholar] [CrossRef]
- Nagel, S.; Pommerenke, C.; Meyer, C.; MacLeod, R.A.F.; Drexler, H.G. Aberrant expression of NKL homeobox genes HMX2 and HMX3 interferes with cell differentiation in acute myeloid leukemia. PLoS ONE 2020, 15, e0240120. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Hosono, Y.; Yanagisawa, K.; Takahashi, T. NKX2-1/TTF-1: An enigmatic oncogene that functions as a double-edged sword for cancer cell survival and progression. Cancer Cell 2013, 23, 718–723. [Google Scholar] [CrossRef] [Green Version]
- Drexler, H.G. Guide to Leukemia-Lymphoma Cell Lines, 2nd ed.; DSMZ: Braunschweig, Germany, 2010. [Google Scholar]
- Drexler, H.G.; MacLeod, R.A. Malignant hematopoietic cell lines: In vitro models for the study of anaplastic large-cell lymphoma. Leukemia 2004, 18, 1569–1571. [Google Scholar] [CrossRef]
- Drexler, H.G.; Matsuo, Y.; MacLeod, R.A. Malignant hematopoietic cell lines: In vitro models for the study of erythroleukemia. Leuk. Res. 2004, 28, 1243–1251. [Google Scholar] [CrossRef] [PubMed]
- Drexler, H.G.; Quentmeier, H.; MacLeod, R.A.; Uphoff, C.C.; Hu, Z.B. Leukemia cell lines: In vitro models for the study of acute promyelocytic leukemia. Leuk. Res. 1995, 19, 681–691. [Google Scholar] [CrossRef]
- Drexler, H.G. Leukemia cell lines: In vitro models for the study of chronic myeloid leukemia. Leuk. Res. 1994, 18, 919–927. [Google Scholar] [CrossRef]
- Matsuo, Y.; Drexler, H.G. Establishment and characterization of human B cell precursor-leukemia cell lines. Leuk. Res. 1998, 22, 567–579. [Google Scholar] [CrossRef]
- Drexler, H.G.; Eberth, S.; Nagel, S.; MacLeod, R.A. Malignant hematopoietic cell lines: In vitro models for double-hit B-cell lymphomas. Leuk. Lymphoma 2016, 57, 1015–1020. [Google Scholar] [CrossRef] [PubMed]
- Drexler, H.G.; Pommerenke, C.; Eberth, S.; Nagel, S. Hodgkin lymphoma cell lines: To separate the wheat from the chaff. Biol. Chem. 2018, 399, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Drexler, H.G. Malignant hematopoietic cell lines: In vitro models for the study of myelodysplastic syndromes. Leuk. Res. 2000, 24, 109–115. [Google Scholar] [CrossRef]
- Drexler, H.G.; Matsuo, Y. Malignant hematopoietic cell lines: In vitro models for the study of multiple myeloma and plasma cell leukemia. Leuk. Res. 2000, 24, 681–703. [Google Scholar] [CrossRef]
- Drexler, H.G.; Matsuo, Y. Malignant hematopoietic cell lines: In vitro models for the study of natural killer cell leukemia-lymphoma. Leukemia 2000, 14, 777–782. [Google Scholar] [CrossRef] [Green Version]
- Carbone, A.; Cesarman, E.; Gloghini, A.; Drexler, H.G. Understanding pathogenetic aspects and clinical presentation of primary effusion lymphoma through its derived cell lines. AIDS 2010, 24, 479–490. [Google Scholar] [CrossRef] [Green Version]
- Drexler, H.G.; Ehrentraut, S.; Nagel, S.; Eberth, S.; MacLeod, R.A. Malignant hematopoietic cell lines: In vitro models for the study of primary mediastinal B-cell lymphomas. Leuk. Res. 2015, 39, 18–29. [Google Scholar] [CrossRef]
- Quentmeier, H.; Pommerenke, C.; Dirks, W.G.; Eberth, S.; Koeppel, M.; MacLeod, R.A.F.; Nagel, S.; Steube, K.; Uphoff, C.C.; Drexler, H.G. The LL-100 panel: 100 cell lines for blood cancer studies. Sci. Rep. 2019, 9, 8218. [Google Scholar] [CrossRef] [PubMed]
- Nagel, S.; Schneider, B.; Meyer, C.; Kaufmann, M.; Drexler, H.G.; Macleod, R.A. Transcriptional deregulation of homeobox gene ZHX2 in Hodgkin lymphoma. Leuk. Res. 2012, 36, 646–655. [Google Scholar] [CrossRef]
- Nagel, S.; MacLeod, R.A.F.; Pommerenke, C.; Meyer, C.; Kaufmann, M.; Drexler, H.G. NKL homeobox gene NKX2-2 is aberrantly expressed in Hodgkin lymphoma. Oncotarget 2018, 9, 37480–37496. [Google Scholar] [CrossRef]
- Nagel, S.; Scherr, M.; Kel, A.; Hornischer, K.; Crawford, G.E.; Kaufmann, M.; Meyer, C.; Drexler, H.G.; MacLeod, R.A. Activation of TLX3 and NKX2-5 in t(5;14)(q35;q32) T-cell acute lymphoblastic leukemia by remote 3’-BCL11B enhancers and coregulation by PU.1 and HMGA1. Cancer Res. 2007, 67, 1461–1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacLeod, R.A.; Nagel, S.; Kaufmann, M.; Janssen, J.W.; Drexler, H.G. Activation of HOX11L2 by juxtaposition with 3’-BCL11B in an acute lymphoblastic leukemia cell line (HPB-ALL) with t(5;14)(q35;q32.2). Genes Chromosomes Cancer 2003, 37, 84–91. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| DLBCL | FL | HCL | HL | MCL | SMZL | AITL | ALCL | ATLL | HSTL | NKTL | PTCL | T-ALL | AML | MDS | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BARHL1 | |||||||||||||||
| BARHL2 | |||||||||||||||
| BARX1 | |||||||||||||||
| BARX2 | |||||||||||||||
| BSX | |||||||||||||||
| DBX1 | |||||||||||||||
| DBX2 | |||||||||||||||
| DLX1 | |||||||||||||||
| DLX2 | |||||||||||||||
| DLX3 | |||||||||||||||
| DLX4 | |||||||||||||||
| DLX5 | |||||||||||||||
| DLX6 | |||||||||||||||
| EMX1 | |||||||||||||||
| EMX2 | |||||||||||||||
| EN1 | |||||||||||||||
| EN2 | |||||||||||||||
| HHEX | |||||||||||||||
| HLX | |||||||||||||||
| HMX1 | |||||||||||||||
| HMX2 | |||||||||||||||
| HMX3 | |||||||||||||||
| LBX1 | |||||||||||||||
| LBX2 | |||||||||||||||
| MSX1 | |||||||||||||||
| MSX2 | |||||||||||||||
| NANOG | |||||||||||||||
| NKX1-1 | |||||||||||||||
| NKX1-2 | |||||||||||||||
| NKX2-1 | |||||||||||||||
| NKX2-2 | |||||||||||||||
| NKX2-3 | |||||||||||||||
| NKX2-4 | |||||||||||||||
| NKX2-5 | |||||||||||||||
| NKX2-6 | |||||||||||||||
| NKX2-8 | |||||||||||||||
| NKX3-1 | |||||||||||||||
| NKX3-2 | |||||||||||||||
| NKX6-1 | |||||||||||||||
| NKX6-2 | |||||||||||||||
| NKX6-3 | |||||||||||||||
| NOTO | |||||||||||||||
| TLX1 | |||||||||||||||
| TLX2 | |||||||||||||||
| TLX3 | |||||||||||||||
| VAX1 | |||||||||||||||
| VAX2 | |||||||||||||||
| VENTX | |||||||||||||||
| 48 (35) | 3 | 3 | 2 | 6 | 5 | 3 | 7 | 9 | 6 | 6 | 11 | 11 | 24 | 18 | 14 |
| Gene | Cell Line | Disease | Regulated by | Effect, Target Genes | Reference |
|---|---|---|---|---|---|
| BARHL1 | |||||
| BARHL2 | |||||
| BARX1 | |||||
| BARX2 | RPMI-8226 | MM | [113] | ||
| BSX | |||||
| DBX1 | |||||
| DBX2 | |||||
| DLX1 | EOL-1 | AML | [98] | ||
| DLX2 | EOL-1 | AML | ERK-signaling | [98] | |
| HPB-ALL | T-ALL | [113] | |||
| DLX3 | |||||
| DLX4 | |||||
| DLX5 | NOMO-1 | AML | NANOG | [30] | |
| DLX6 | NOMO-1 | AML | NANOG | [30] | |
| EMX1 | |||||
| EMX2 | |||||
| EN1 | |||||
| EN2 | |||||
| HHEX | CCRF-CEM | T-ALL | [113] | ||
| RPMI-8402 | T-ALL | [113] | |||
| HLX | L-540 | HL | aberrant STAT3 activity | [91] | |
| DOHH-2 | DLBCL | EBV-mediated STAT3 | [92] | ||
| OCI-LY19 | DLBCL | [113] | |||
| NU-DHL-1 | DLBCL | [113] | |||
| SEM | BCP-ALL | [113] | |||
| DEL | ALCL | aberrant STAT3 activity | [29] | ||
| KI-JK | ALCL | aberrant STAT3 activity | [29] | ||
| L-82 | ALCL | aberrant STAT3 activity | [29] | ||
| SR-786 | ALCL | aberrant STAT3 activity | [29] | ||
| SU-DHL-1 | ALCL | aberrant STAT3 activity | [29] | ||
| SUP-M2 | ALCL | aberrant STAT3 activity | [29] | ||
| HMX1 | |||||
| HMX2 | EOL-1 | AML | inhibits EPX, activates HRT7 and ERK | [98] | |
| MOLM-13 | AML | [98] | |||
| MV4-11 | AML | [98] | |||
| 697 | BCP-ALL | [98] | |||
| REH | BCP-ALL | [98] | |||
| HMX3 | EOL-1 | AML | inhibits EPX, activates HRT7 and ERK | [98] | |
| MOLM-13 | AML | [98] | |||
| MV4-11 | AML | [98] | |||
| SEM | BCP-ALL | [113] | |||
| LBX1 | |||||
| LBX2 | |||||
| MSX1 | L-1236 | HL | suppressed BMP-pathway | cofactor H1C, target ZHX2 | [114] |
| GRANTA-519 | MCL | CCND1 in t(11;14)(q13;q32) | [88] | ||
| JEKO-1 | MCL | CCND1 in t(11;14)(q13;q32) | [88] | ||
| REC-1 | MCL | CCND1 in t(11;14)(q13;q32) | [88] | ||
| LOUCY | ETP-ALL | [89] | |||
| MSX2 | |||||
| NANOG | NOMO-1 | AML | NOTCH, CDK6, MIR17HG | [30] | |
| NKX1-1 | |||||
| NKX1-2 | |||||
| NKX2-1 | SU-DHL-5 | DLBCL | KMT2A, H2B | [83] | |
| RPMI-8402 | T-ALL | [113] | |||
| NKX2-2 | DEV | HL | IL17RB-signaling | [115] | |
| NKX2-3 | |||||
| NKX2-4 | |||||
| NKX2-5 | CCRF-CEM | T-ALL | t(5;14)(q35;q32) | activates MEF2C | [76,80] |
| PEER | T-ALL | t(5;14)(q35;q32) | activates MEF2C | [76,80] | |
| NKX2-6 | |||||
| NKX2-8 | |||||
| NKX3-1 | JURKAT | T-ALL | TAL1 and GATA3 | activates SIX6 | [84,86] |
| MOLT-4 | T-ALL | [86] | |||
| PER-117 | T-ALL | [86] | |||
| PF-392 | T-ALL | [86] | |||
| RPMI-8402 | T-ALL | [86] | |||
| OCI-LY9 | DLBCL | [113] | |||
| NKX3-2 | CCRF-CEM | T-ALL | suppressed BMP-pathway | activates SIX6 | [87] |
| NKX6-1 | |||||
| NKX6-2 | |||||
| NKX6-3 | DOHH-2 | DLBCL | repressed by HLX and HHEX, activated by PAX5 and MYB | inhibits PAX5 and MSX1 | [27] |
| NOTO | |||||
| TLX1 | ALL-SIL | T-ALL | t(10;14)(q24;q11) | ETS1-interaction | [70,75] |
| TLX2 | SUP-HD1 | HL | activated by PBX1 | activates TBX15 | [45] |
| TLX3 | DND-41 | T-ALL | t(5;14)(q35;q32) | [116] | |
| HPB-ALL | T-ALL | t(5;14)(q35;q32) | [117] | ||
| VAX1 | |||||
| VAX2 | |||||
| VENTX | SEM | BCP-ALL | [113] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagel, S. NKL-Code in Normal and Aberrant Hematopoiesis. Cancers 2021, 13, 1961. https://doi.org/10.3390/cancers13081961
Nagel S. NKL-Code in Normal and Aberrant Hematopoiesis. Cancers. 2021; 13(8):1961. https://doi.org/10.3390/cancers13081961
Chicago/Turabian StyleNagel, Stefan. 2021. "NKL-Code in Normal and Aberrant Hematopoiesis" Cancers 13, no. 8: 1961. https://doi.org/10.3390/cancers13081961
APA StyleNagel, S. (2021). NKL-Code in Normal and Aberrant Hematopoiesis. Cancers, 13(8), 1961. https://doi.org/10.3390/cancers13081961