
Genetic Analysis Reveals the Prognostic Significance of the DNA Mismatch Repair Gene MSH2 in Advanced Prostate Cancer

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Response Evaluation
2.2. SNP Selection and Genotyping
2.3. Bioinformatics Analysis
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chinegwundoh, F.; Enver, M.; Lee, A.; Nargund, V.; Oliver, T.; Ben-Shlomo, Y. Risk and presenting features of prostate cancer amongst African-Caribbean, South Asian and European men in North-east London. BJU Int. 2006, 98, 1216–1220. [Google Scholar] [CrossRef] [PubMed]
- Mucci, L.A.; Hjelmborg, J.B.; Harris, J.R.; Czene, K.; Havelick, D.J.; Scheike, T.; Graff, R.E.; Holst, K.; Moller, S.; Unger, R.H.; et al. Familial Risk and Heritability of Cancer Among Twins in Nordic Countries. JAMA 2016, 315, 68–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aune, D.; Navarro Rosenblatt, D.A.; Chan, D.S.; Vieira, A.R.; Vieira, R.; Greenwood, D.C.; Vatten, L.J.; Norat, T. Dairy products, calcium, and prostate cancer risk: A systematic review and meta-analysis of cohort studies. Am. J. Clin. Nutr. 2015, 101, 87–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowles, J.L., 3rd; Ranard, K.M.; Smith, J.W.; An, R.; Erdman, J.W., Jr. Increased dietary and circulating lycopene are associated with reduced prostate cancer risk: A systematic review and meta-analysis. Prostate Cancer Prostatic Dis. 2017, 20, 361–377. [Google Scholar] [CrossRef]
- Fletcher, S.A.; von Landenberg, N.; Cole, A.P.; Gild, P.; Choueiri, T.K.; Lipsitz, S.R.; Trinh, Q.D.; Kibel, A.S. Contemporary national trends in prostate cancer risk profile at diagnosis. Prostate Cancer Prostatic Dis. 2020, 23, 81–87. [Google Scholar] [CrossRef]
- Huggins, C.; Hodges, C.V. Studies on prostatic cancer. I. The effect of castration, of estrogen and androgen injection on serum phosphatases in metastatic carcinoma of the prostate. CA Cancer J. Clin. 1972, 22, 232–240. [Google Scholar] [CrossRef]
- Frank, S.; Nelson, P.; Vasioukhin, V. Recent advances in prostate cancer research: Large-scale genomic analyses reveal novel driver mutations and DNA repair defects. F1000Research 2018, 7, 1173. [Google Scholar] [CrossRef]
- Lockett, K.L.; Hall, M.C.; Clark, P.E.; Chuang, S.C.; Robinson, B.; Lin, H.Y.; Su, L.J.; Hu, J.J. DNA damage levels in prostate cancer cases and controls. Carcinogenesis 2006, 27, 1187–1193. [Google Scholar] [CrossRef] [Green Version]
- Miyake, H.; Hara, I.; Kamidono, S.; Eto, H. Oxidative DNA damage in patients with prostate cancer and its response to treatment. J. Urol. 2004, 171, 1533–1536. [Google Scholar] [CrossRef]
- Dronkert, M.L.; Kanaar, R. Repair of DNA interstrand cross-links. Mutat. Res. 2001, 486, 217–247. [Google Scholar] [CrossRef]
- Dall’Era, M.A.; McPherson, J.D.; Gao, A.C.; DeVere White, R.W.; Gregg, J.P.; Lara, P.N., Jr. Germline and somatic DNA repair gene alterations in prostate cancer. Cancer 2020, 126, 2980–2985. [Google Scholar] [CrossRef]
- Marshall, C.H.; Fu, W.; Wang, H.; Baras, A.S.; Lotan, T.L.; Antonarakis, E.S. Prevalence of DNA repair gene mutations in localized prostate cancer according to clinical and pathologic features: Association of Gleason score and tumor stage. Prostate Cancer Prostatic Dis. 2019, 22, 59–65. [Google Scholar] [CrossRef]
- Robinson, D.; Van Allen, E.M.; Wu, Y.M.; Schultz, N.; Lonigro, R.J.; Mosquera, J.M.; Montgomery, B.; Taplin, M.E.; Pritchard, C.C.; Attard, G.; et al. Integrative clinical genomics of advanced prostate cancer. Cell 2015, 161, 1215–1228. [Google Scholar] [CrossRef] [Green Version]
- Moro, L.; Arbini, A.A.; Yao, J.L.; di Sant’Agnese, P.A.; Marra, E.; Greco, M. Loss of BRCA2 promotes prostate cancer cell invasion through up-regulation of matrix metalloproteinase-9. Cancer Sci. 2008, 99, 553–563. [Google Scholar] [CrossRef]
- Kim, S.H.; Park, W.S.; Yun, S.I.; Joo, J.; Joung, J.Y.; Seo, H.K.; Chung, J.; Lee, G.K.; Lee, K.H. Overexpression of BRCA1 or BRCA2 in prostatectomy specimens is predictive of biochemical recurrence after radical prostatectomy. Histopathology 2016, 68, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Leongamornlert, D.A.; Saunders, E.J.; Wakerell, S.; Whitmore, I.; Dadaev, T.; Cieza-Borrella, C.; Benafif, S.; Brook, M.N.; Donovan, J.L.; Hamdy, F.C.; et al. Germline DNA Repair Gene Mutations in Young-onset Prostate Cancer Cases in the UK: Evidence for a More Extensive Genetic Panel. Eur. Urol. 2019, 76, 329–337. [Google Scholar] [CrossRef] [Green Version]
- Na, R.; Zheng, S.L.; Han, M.; Yu, H.; Jiang, D.; Shah, S.; Ewing, C.M.; Zhang, L.; Novakovic, K.; Petkewicz, J.; et al. Germline Mutations in ATM and BRCA1/2 Distinguish Risk for Lethal and Indolent Prostate Cancer and are Associated with Early Age at Death. Eur. Urol. 2017, 71, 740–747. [Google Scholar] [CrossRef] [Green Version]
- Castro, E.; Goh, C.; Olmos, D.; Saunders, E.; Leongamornlert, D.; Tymrakiewicz, M.; Mahmud, N.; Dadaev, T.; Govindasami, K.; Guy, M.; et al. Germline BRCA mutations are associated with higher risk of nodal involvement, distant metastasis, and poor survival outcomes in prostate cancer. J. Clin. Oncol. 2013, 31, 1748–1757. [Google Scholar] [CrossRef] [Green Version]
- Leongamornlert, D.; Saunders, E.; Dadaev, T.; Tymrakiewicz, M.; Goh, C.; Jugurnauth-Little, S.; Kozarewa, I.; Fenwick, K.; Assiotis, I.; Barrowdale, D.; et al. Frequent germline deleterious mutations in DNA repair genes in familial prostate cancer cases are associated with advanced disease. Br. J. Cancer 2014, 110, 1663–1672. [Google Scholar] [CrossRef] [PubMed]
- Marshall, C.H.; Sokolova, A.O.; McNatty, A.L.; Cheng, H.H.; Eisenberger, M.A.; Bryce, A.H.; Schweizer, M.T.; Antonarakis, E.S. Differential Response to Olaparib Treatment Among Men with Metastatic Castration-resistant Prostate Cancer Harboring BRCA1 or BRCA2 Versus ATM Mutations. Eur. Urol. 2019, 76, 452–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mateo, J.; Carreira, S.; Sandhu, S.; Miranda, S.; Mossop, H.; Perez-Lopez, R.; Nava Rodrigues, D.; Robinson, D.; Omlin, A.; Tunariu, N.; et al. DNA-Repair Defects and Olaparib in Metastatic Prostate Cancer. N. Engl. J. Med. 2015, 373, 1697–1708. [Google Scholar] [CrossRef] [PubMed]
- Damaraju, S.; Murray, D.; Dufour, J.; Carandang, D.; Myrehaug, S.; Fallone, G.; Field, C.; Greiner, R.; Hanson, J.; Cass, C.E.; et al. Association of DNA repair and steroid metabolism gene polymorphisms with clinical late toxicity in patients treated with conformal radiotherapy for prostate cancer. Clin. Cancer Res. 2006, 12, 2545–2554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, R.; Price, D.K.; Dahut, W.L.; Reed, E.; Figg, W.D. Genetic polymorphisms in XRCC1 associated with radiation therapy in prostate cancer. Cancer Biol. Ther. 2010, 10, 13–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanusso, C.; Bortolus, R.; Dreussi, E.; Polesel, J.; Montico, M.; Cecchin, E.; Gagno, S.; Rizzolio, F.; Arcicasa, M.; Novara, G.; et al. Impact of DNA repair gene polymorphisms on the risk of biochemical recurrence after radiotherapy and overall survival in prostate cancer. Oncotarget 2017, 8, 22863–22875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Little, J.; Wilson, B.; Carter, R.; Walker, K.; Santaguida, P.; Tomiak, E.; Beyene, J.; Usman Ali, M.; Raina, P. Multigene panels in prostate cancer risk assessment: A systematic review. Genet. Med. 2016, 18, 535–544. [Google Scholar] [CrossRef] [Green Version]
- Bao, B.Y.; Pao, J.B.; Huang, C.N.; Pu, Y.S.; Chang, T.Y.; Lan, Y.H.; Lu, T.L.; Lee, H.Z.; Juang, S.H.; Chen, L.M.; et al. Polymorphisms inside microRNAs and microRNA target sites predict clinical outcomes in prostate cancer patients receiving androgen-deprivation therapy. Clin. Cancer Res. 2011, 17, 928–936. [Google Scholar] [CrossRef] [Green Version]
- Geng, J.H.; Lin, V.C.; Yu, C.C.; Huang, C.Y.; Yin, H.L.; Chang, T.Y.; Lu, T.L.; Huang, S.P.; Bao, B.Y. Inherited Variants in Wnt Pathway Genes Influence Outcomes of Prostate Cancer Patients Receiving Androgen Deprivation Therapy. Int. J. Mol. Sci. 2016, 17, 1970. [Google Scholar] [CrossRef] [Green Version]
- Gong, J.; Posadas, E.; Bhowmick, N.; Kim, H.; Daskivich, T.; Gupta, A.; Sandler, H.; Kamrava, M.; Zumsteg, Z.; Freedland, S.; et al. Integrating PARP Inhibitors Into Advanced Prostate Cancer Therapeutics. Oncology 2021, 35, 119–125. [Google Scholar] [CrossRef]
- Genomes Project, C.; Abecasis, G.R.; Auton, A.; Brooks, L.D.; De Pristo, M.A.; Durbin, R.M.; Handsaker, R.E.; Kang, H.M.; Marth, G.T.; McVean, G.A. An integrated map of genetic variation from 1092 human genomes. Nature 2012, 491, 56–65. [Google Scholar]
- de Bakker, P.I.; Yelensky, R.; Pe’er, I.; Gabriel, S.B.; Daly, M.J.; Altshuler, D. Efficiency and power in genetic association studies. Nat. Genet. 2005, 37, 1217–1223. [Google Scholar] [CrossRef]
- Bao, B.Y.; Pao, J.B.; Huang, C.N.; Pu, Y.S.; Chang, T.Y.; Lan, Y.H.; Lu, T.L.; Lee, H.Z.; Chen, L.M.; Ting, W.C.; et al. Significant associations of prostate cancer susceptibility variants with survival in patients treated with androgen-deprivation therapy. Int. J. Cancer 2012, 130, 876–884. [Google Scholar] [CrossRef] [PubMed]
- Ward, L.D.; Kellis, M. HaploReg v4: Systematic mining of putative causal variants, cell types, regulators and target genes for human complex traits and disease. Nucleic Acids Res. 2016, 44, D877–D881. [Google Scholar] [CrossRef] [PubMed]
- Consortium, G.T. The Genotype-Tissue Expression (GTEx) project. Nat. Genet. 2013, 45, 580–585. [Google Scholar]
- The Cancer Genome Atlas (TCGA) Research Network. Comprehensive genomic characterization defines human glioblastoma genes and core pathways. Nature 2008, 455, 1061–1068. [Google Scholar] [CrossRef]
- Rhodes, D.R.; Yu, J.; Shanker, K.; Deshpande, N.; Varambally, R.; Ghosh, D.; Barrette, T.; Pandey, A.; Chinnaiyan, A.M. ONCOMINE: A cancer microarray database and integrated data-mining platform. Neoplasia 2004, 6, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Park, S.J.; Yoon, B.H.; Kim, S.K.; Kim, S.Y. GENT2: An updated gene expression database for normal and tumor tissues. BMC Med. Genom. 2019, 12, 101. [Google Scholar] [CrossRef]
- Barr, F.G. Chromosomal translocations involving paired box transcription factors in human cancer. Int. J. Biochem. Cell Biol. 1997, 29, 1449–1461. [Google Scholar] [CrossRef]
- Wu, X.; Xiao, Y.; Zhou, Y.; Zhou, Z.; Yan, W. LncRNA FOXP4-AS1 is activated by PAX5 and promotes the growth of prostate cancer by sequestering miR-3184-5p to upregulate FOXP4. Cell Death Dis. 2019, 10, 472. [Google Scholar] [CrossRef] [Green Version]
- Acharya, S.; Wilson, T.; Gradia, S.; Kane, M.F.; Guerrette, S.; Marsischky, G.T.; Kolodner, R.; Fishel, R. hMSH2 forms specific mispair-binding complexes with hMSH3 and hMSH6. Proc. Natl. Acad. Sci. USA 1996, 93, 13629–13634. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Frederiksen, J.H.; Liberti, S.E.; Lutzen, A.; Keijzers, G.; Pena-Diaz, J.; Rasmussen, L.J. Human DNA polymerase delta double-mutant D316A;E318A interferes with DNA mismatch repair in vitro. Nucleic Acids Res. 2017, 45, 9427–9440. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, F.C.; Jager, A.C.; Lutzen, A.; Bundgaard, J.R.; Rasmussen, L.J. Characterization of human exonuclease 1 in complex with mismatch repair proteins, subcellular localization and association with PCNA. Oncogene 2004, 23, 1457–1468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albero-Gonzalez, R.; Hernandez-Llodra, S.; Juanpere, N.; Lorenzo, M.; Lloret, A.; Segales, L.; Duran, X.; Fumado, L.; Cecchini, L.; Lloreta-Trull, J. Immunohistochemical expression of mismatch repair proteins (MSH2, MSH6, MLH1, and PMS2) in prostate cancer: Correlation with grade groups (WHO 2016) and ERG and PTEN status. Virchows Arch. 2019, 475, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Guedes, L.B.; Antonarakis, E.S.; Schweizer, M.T.; Mirkheshti, N.; Almutairi, F.; Park, J.C.; Glavaris, S.; Hicks, J.; Eisenberger, M.A.; De Marzo, A.M.; et al. MSH2 Loss in Primary Prostate Cancer. Clin. Cancer Res. 2017, 23, 6863–6874. [Google Scholar] [CrossRef] [Green Version]
- Burger, M.; Denzinger, S.; Hammerschmied, C.G.; Tannapfel, A.; Obermann, E.C.; Wieland, W.F.; Hartmann, A.; Stoehr, R. Elevated microsatellite alterations at selected tetranucleotides (EMAST) and mismatch repair gene expression in prostate cancer. J. Mol. Med. 2006, 84, 833–841. [Google Scholar] [CrossRef]
- Prtilo, A.; Leach, F.S.; Markwalder, R.; Kappeler, A.; Burkhard, F.C.; Cecchini, M.G.; Studer, U.E.; Thalmann, G.N. Tissue microarray analysis of hMSH2 expression predicts outcome in men with prostate cancer. J. Urol. 2005, 174, 1814–1818. [Google Scholar] [CrossRef]
- Velasco, A.; Hewitt, S.M.; Albert, P.S.; Hossein, M.; Rosenberg, H.; Martinez, C.; Sagalowsky, A.I.; McConnell, J.D.; Marston, W.; Leach, F.S. Differential expression of the mismatch repair gene hMSH2 in malignant prostate tissue is associated with cancer recurrence. Cancer 2002, 94, 690–699. [Google Scholar] [CrossRef]
- Kauffmann, A.; Rosselli, F.; Lazar, V.; Winnepenninckx, V.; Mansuet-Lupo, A.; Dessen, P.; van den Oord, J.J.; Spatz, A.; Sarasin, A. High expression of DNA repair pathways is associated with metastasis in melanoma patients. Oncogene 2008, 27, 565–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Liu, L.; Wang, Z.; Wang, L.; Liu, Z.; Xu, G.; Lu, S. Overexpression of hMSH2 and hMLH1 protein in certain gastric cancers and their surrounding mucosae. Oncol. Rep. 2008, 19, 401–406. [Google Scholar] [CrossRef] [Green Version]
- Wagner, V.P.; Webber, L.P.; Salvadori, G.; Meurer, L.; Fonseca, F.P.; Castilho, R.M.; Squarize, C.H.; Vargas, P.A.; Martins, M.D. Overexpression of MutSalpha Complex Proteins Predicts Poor Prognosis in Oral Squamous Cell Carcinoma. Medicine 2016, 95, e3725. [Google Scholar] [CrossRef]
- Wilczak, W.; Rashed, S.; Hube-Magg, C.; Kluth, M.; Simon, R.; Buscheck, F.; Clauditz, T.S.; Grupp, K.; Minner, S.; Tsourlakis, M.C.; et al. Up-regulation of mismatch repair genes MSH6, PMS2 and MLH1 parallels development of genetic instability and is linked to tumor aggressiveness and early PSA recurrence in prostate cancer. Carcinogenesis 2017, 38, 19–27. [Google Scholar] [CrossRef]
- James, N.D.; Sydes, M.R.; Clarke, N.W.; Mason, M.D.; Dearnaley, D.P.; Spears, M.R.; Ritchie, A.W.; Parker, C.C.; Russell, J.M.; Attard, G.; et al. Addition of docetaxel, zoledronic acid, or both to first-line long-term hormone therapy in prostate cancer (STAMPEDE): Survival results from an adaptive, multiarm, multistage, platform randomised controlled trial. Lancet 2016, 387, 1163–1177. [Google Scholar] [CrossRef] [Green Version]
- Sweeney, C.J.; Chen, Y.H.; Carducci, M.; Liu, G.; Jarrard, D.F.; Eisenberger, M.; Wong, Y.N.; Hahn, N.; Kohli, M.; Cooney, M.M.; et al. Chemohormonal Therapy in Metastatic Hormone-Sensitive Prostate Cancer. N. Engl. J. Med. 2015, 373, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Abida, W.; Campbell, D.; Patnaik, A.; Shapiro, J.D.; Sautois, B.; Vogelzang, N.J.; Voog, E.G.; Bryce, A.H.; McDermott, R.; Ricci, F.; et al. Non-BRCA DNA Damage Repair Gene Alterations and Response to the PARP Inhibitor Rucaparib in Metastatic Castration-Resistant Prostate Cancer: Analysis From the Phase II TRITON2 Study. Clin. Cancer Res. 2020, 26, 2487–2496. [Google Scholar] [CrossRef] [Green Version]
- de Bono, J.; Mateo, J.; Fizazi, K.; Saad, F.; Shore, N.; Sandhu, S.; Chi, K.N.; Sartor, O.; Agarwal, N.; Olmos, D.; et al. Olaparib for Metastatic Castration-Resistant Prostate Cancer. N. Engl. J. Med. 2020, 382, 2091–2102. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, J.F.; Schiewer, M.J.; Dean, J.L.; Schrecengost, R.S.; de Leeuw, R.; Han, S.; Ma, T.; Den, R.B.; Dicker, A.P.; Feng, F.Y.; et al. A hormone-DNA repair circuit governs the response to genotoxic insult. Cancer Discov. 2013, 3, 1254–1271. [Google Scholar] [CrossRef] [Green Version]
- Thompson, T.C.; Li, L.; Broom, B.M. Combining enzalutamide with PARP inhibitors: Pharmaceutically induced BRCAness. Oncotarget 2017, 8, 93315–93316. [Google Scholar] [CrossRef]







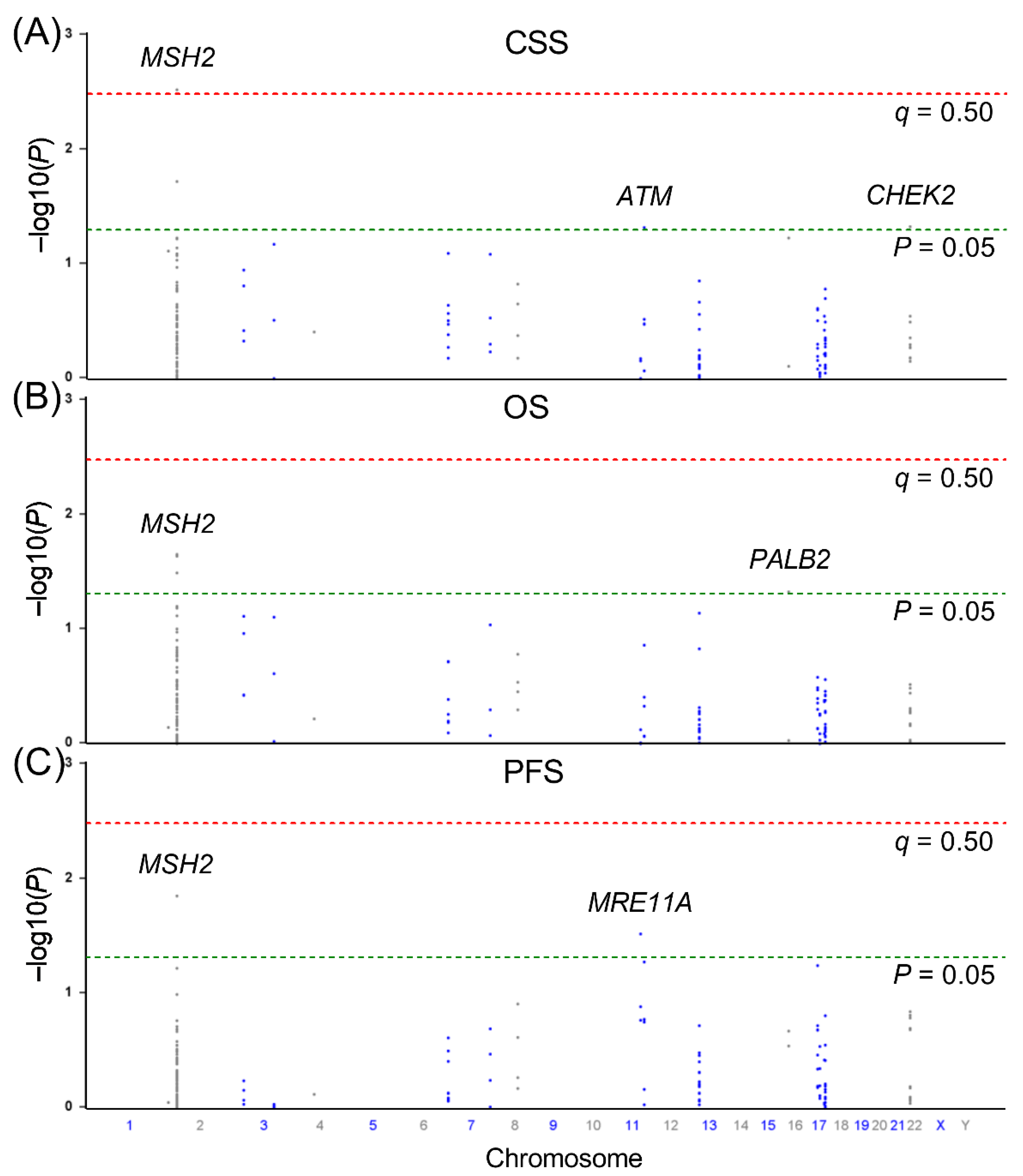{kind=link}
{kind=link}
{kind=link}
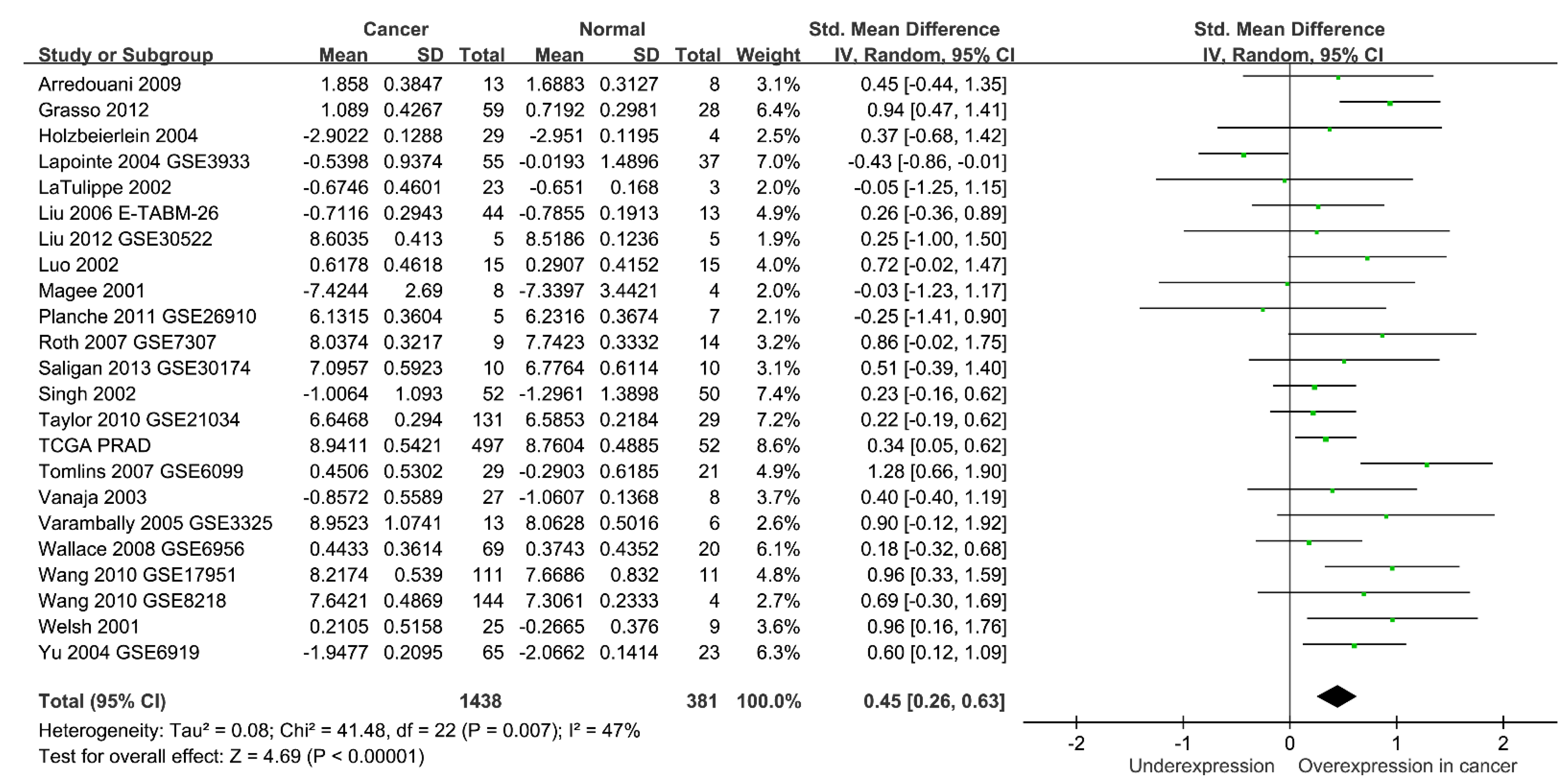{kind=link}
{kind=link}
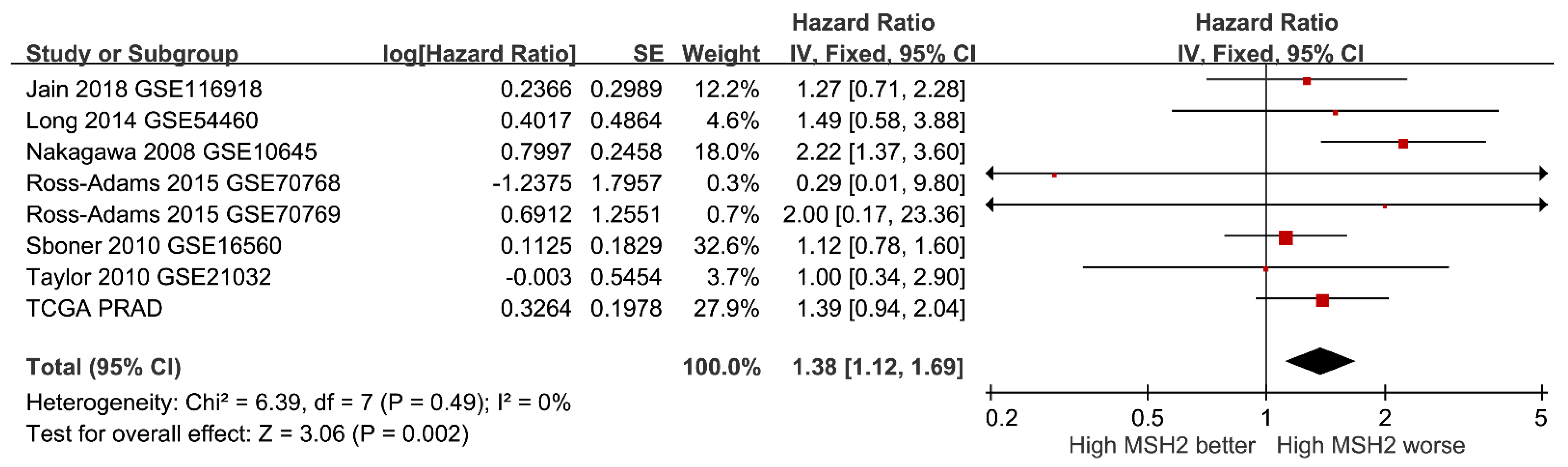{kind=link}
| Characteristics | CSS a | OS a | PFS a | ||||
|---|---|---|---|---|---|---|---|
| HR (95% CI) | p | HR (95% CI) | p | HR (95% CI) | p | ||
| Total, n (%) | 630 | 314 (49.8) | 414 (65.7) | 518 (82.6) | |||
| Age at diagnosis, years | |||||||
| Median (IQR) | 73 (67–79) | 1.012 (0.999–1.026) | 0.078 | 1.029 (1.016–1.041) | <0.001 | 0.980 (0.970–0.990) | <0.001 |
| PSA at ADT initiation, ng/mL | |||||||
| Median (IQR) | 34.5 (11.25–129) | 1.000 (1.000–1.000) | <0.001 | 1.000 (1.000–1.000) | <0.001 | 1.000 (1.000–1.000) | 0.001 |
| PSA nadir, ng/mL | |||||||
| Median (IQR) | 0.14 (0.01–1.16) | 1.002 (1.001–1.002) | <0.001 | 1.002 (1.001–1.002) | <0.001 | 1.001 (1.000–1.001) | 0.019 |
| Time to PSA nadir, months | |||||||
| Median (IQR) | 11 (5–20) | 0.972 (0.962–0.981) | <0.001 | 0.982 (0.975–0.990) | <0.001 | 0.956 (0.948–0.963) | <0.001 |
| Clinical stage at diagnosis | |||||||
| T1/T2, n (%) | 187 (29.9) | Reference | Reference | Reference | |||
| T3/T4/N1, n (%) | 205 (32.8) | 1.080 (0.785–1.488) | 0.636 | 1.065 (0.818–1.386) | 0.642 | 0.860 (0.686–1.078) | 0.190 |
| M1, n (%) | 233 (37.3) | 3.096 (2.334–4.106) | <0.001 | 2.508 (1.968–3.194) | <0.001 | 1.406 (1.136–1.739) | 0.002 |
| Gleason score at diagnosis | |||||||
| 2–6, n (%) | 188 (30.6) | Reference | Reference | Reference | |||
| 7, n (%) | 194 (31.6) | 1.063 (0.783–1.443) | 0.694 | 1.064 (0.820–1.381) | 0.640 | 1.162 (0.929–1.455) | 0.189 |
| 8–10, n (%) | 232 (37.8) | 2.144 (1.629–2.820) | <0.001 | 1.957 (1.542–2.484) | <0.001 | 1.483 (1.194–1.841) | <0.001 |
| Genotype | Frequency | CSS | OS | ||||||
|---|---|---|---|---|---|---|---|---|---|
| HR (95% CI) | p | HR (95% CI) a | pa | HR (95% CI) | p | HR (95% CI) a | pa | ||
| CC/CG/GG | 244/269/84 | 0.78 (0.65–0.92) | 0.003 | 0.75 (0.63–0.90) | 0.002 | 0.84 (0.73–0.98) | 0.023 | 0.84 (0.72–0.98) | 0.029 |
| Variant | Position | LD (r2) | Reference Allele | Alternate Allele | ASN Frquency | Promoter Histone Marks | Enhancer Histone Marks | DNase | Proteins Bound | Motifs Changed |
|---|---|---|---|---|---|---|---|---|---|---|
| rs1882450 | 47647464 | 0.82 | G | C | 0.64 | 2 altered motifs | ||||
| rs2091750 | 47647822 | 0.8 | T | G | 0.65 | 5 altered motifs | ||||
| rs1400633 | 47656863 | 1 | G | C | 0.65 | 4 tissues | 5 tissues | 3 bound proteins | 2 altered motifs | |
| rs2969775 | 47658337 | 1 | G | T | 0.65 | 2 tissues | 7 altered motifs | |||
| rs2969774 | 47661684 | 0.94 | T | C | 0.65 | 2 tissues | 2 altered motifs | |||
| rs2705766 | 47663192 | 0.95 | T | C | 0.66 | 2 tissues | 3 tissues | 1 tissue | 2 altered motifs | |
| rs2705767 | 47664866 | 0.92 | G | A | 0.65 | 2 tissues | 1 bound protein | 5 altered motifs | ||
| rs2944783 | 47665860 | 0.92 | G | C | 0.65 | 8 altered motifs | ||||
| rs2969772 | 47667333 | 0.9 | A | G | 0.65 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, H.-H.; Lee, C.-H.; Chen, Y.-T.; Huang, C.-Y.; Yu, C.-C.; Lin, V.C.; Geng, J.-H.; Lu, T.-L.; Huang, S.-P.; Bao, B.-Y. Genetic Analysis Reveals the Prognostic Significance of the DNA Mismatch Repair Gene MSH2 in Advanced Prostate Cancer. Cancers 2022, 14, 223. https://doi.org/10.3390/cancers14010223
Chang H-H, Lee C-H, Chen Y-T, Huang C-Y, Yu C-C, Lin VC, Geng J-H, Lu T-L, Huang S-P, Bao B-Y. Genetic Analysis Reveals the Prognostic Significance of the DNA Mismatch Repair Gene MSH2 in Advanced Prostate Cancer. Cancers. 2022; 14(1):223. https://doi.org/10.3390/cancers14010223
Chicago/Turabian StyleChang, Hao-Han, Cheng-Hsueh Lee, Yei-Tsung Chen, Chao-Yuan Huang, Chia-Cheng Yu, Victor C. Lin, Jiun-Hung Geng, Te-Ling Lu, Shu-Pin Huang, and Bo-Ying Bao. 2022. "Genetic Analysis Reveals the Prognostic Significance of the DNA Mismatch Repair Gene MSH2 in Advanced Prostate Cancer" Cancers 14, no. 1: 223. https://doi.org/10.3390/cancers14010223
APA StyleChang, H.-H., Lee, C.-H., Chen, Y.-T., Huang, C.-Y., Yu, C.-C., Lin, V. C., Geng, J.-H., Lu, T.-L., Huang, S.-P., & Bao, B.-Y. (2022). Genetic Analysis Reveals the Prognostic Significance of the DNA Mismatch Repair Gene MSH2 in Advanced Prostate Cancer. Cancers, 14(1), 223. https://doi.org/10.3390/cancers14010223