
Targeting CD38 in Neoplasms and Non-Cancer Diseases

 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
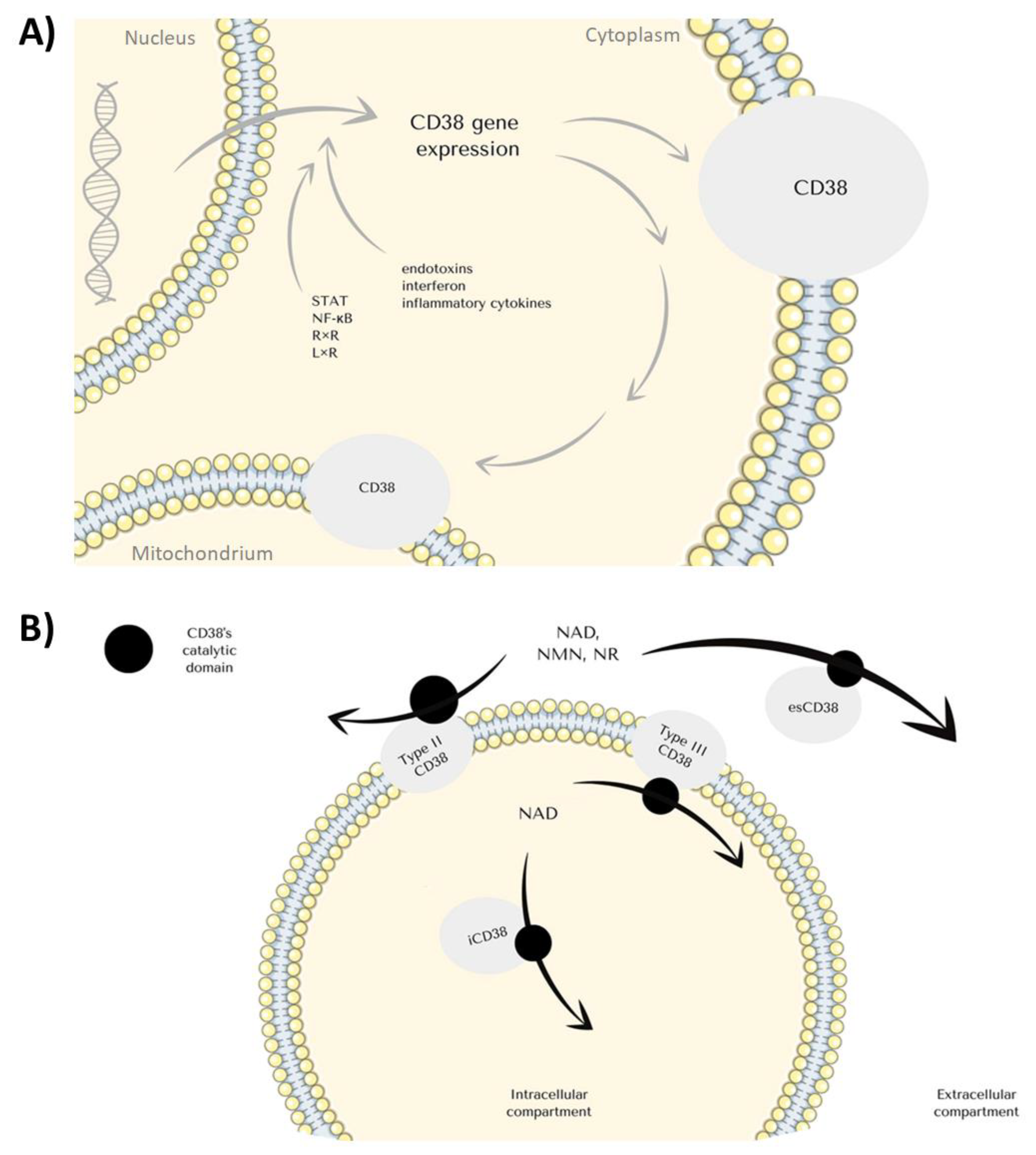
1. CD38 Structure and Function
2. CD38 Expression
2.1. The Biological Consequences of CD38 Stimulation End Expression
2.2. CD38 Expression and Inflammation
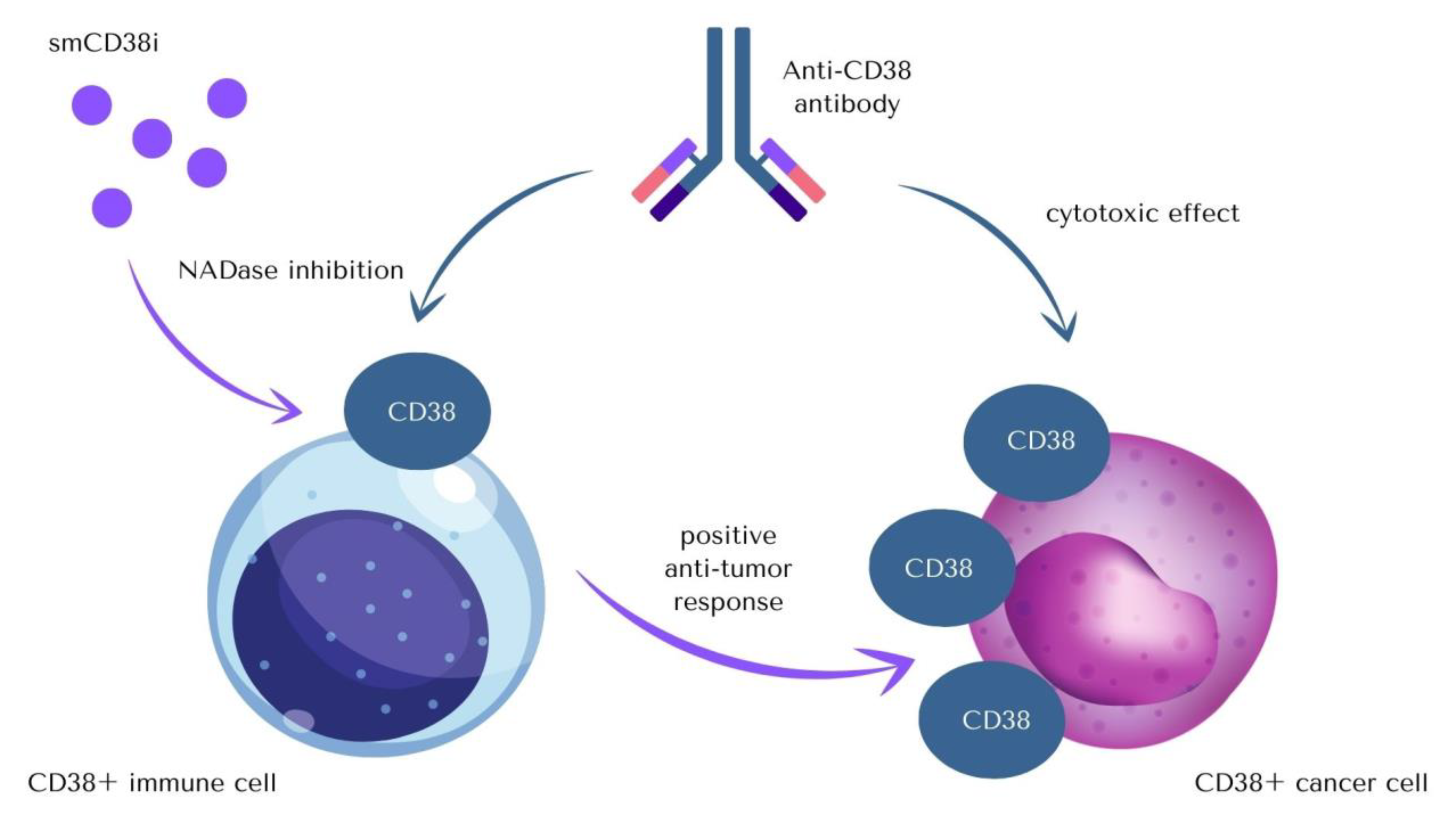
3. Anti-CD38 Therapeutics
4. CD38 in Hematological Neoplasms and Diseases
4.1. Acute Myeloid Leukemia
4.2. Acute Lymphoblastic Leukaemia
4.3. Chronic Lymphocytic Leukaemia/Small Lymphoma (CLL/SLL), B-Cell Prolymphocytic Leukaemia (B-PLL)
4.4. NK-T Cell Lymphoma
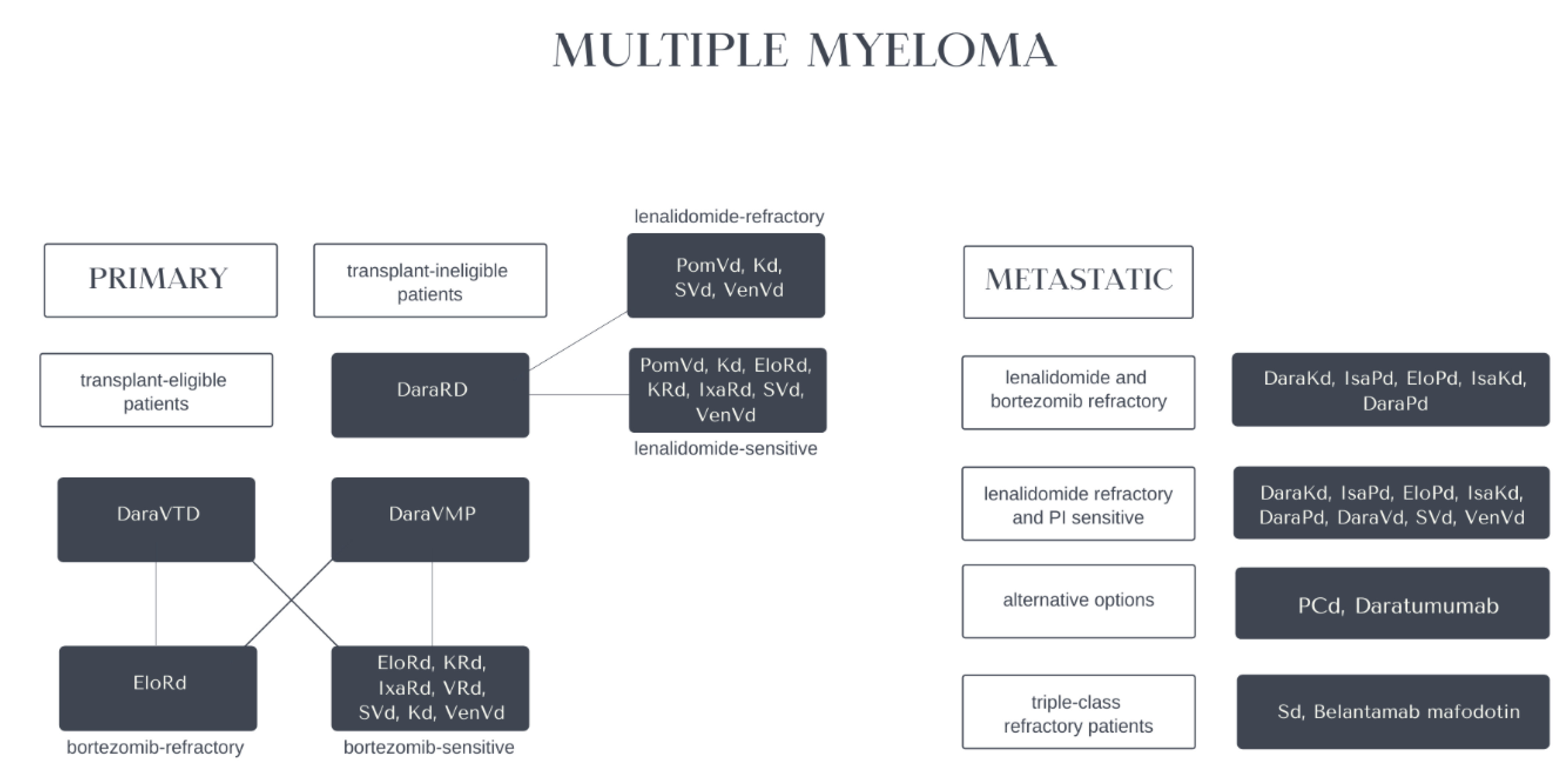
4.5. Multiple Myeloma
4.6. Aplastic Anaemia
4.7. Immune Thrombocytopenic Purpura
4.8. Immunoglobulin Light-Chain (AL) Amyloidosis
4.9. Other Hematological Abnormalities
5. CD38 Outside Hematology
5.1. Non-Hematological Malignancies
5.2. HIV Infection, AIDS
5.3. COVID-19
5.4. Gastrointestinal Diseases
5.5. Pulmonary Injury Induced by Escherichia Coli
5.6. Neurological Diseases
5.7. Systemic Lupus Erythematosus
5.8. Transplant Rejection
5.9. Arthritis
6. Experimental Medicine Involving CD38—Clinical Trials
6.1. Daratumumab and Isatuximab in Multiple Myeloma, NK- and T-Cell Lymphomas, and B-Cell Malignancies
6.2. Daratumumab and Isatuximab in Lymphomas and Acute Lymphoblastic Leukemias
6.3. Daratumumab and Isatuximab in AML
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Amici, S.A.; Young, N.A.; Narvaez-Miranda, J.; Jablonski, K.A.; Arcos, J.; Rosas, L.; Papenfuss, T.L.; Torrelles, J.B.; Jarjour, W.N.; Guerau-de-Arellano, M. CD38 Is Robustly Induced in Human Macrophages and Monocytes in Inflammatory Conditions. Front. Immunol. 2018, 9, 1593. [Google Scholar] [CrossRef] [PubMed]
- Kar, A.; Mehrotra, S.; Chatterjee, S. CD38: T Cell Immuno-Metabolic Modulator. Cells 2020, 9, 1716. [Google Scholar] [CrossRef] [PubMed]
- Deaglio, S.; Capobianco, A.; Bergui, L.; Dürig, J.; Morabito, F.; Dührsen, U.; Malavasi, F. CD38 Is a Signaling Molecule in B-Cell Chronic Lymphocytic Leukemia Cells. Blood 2003, 102, 2146–2155. [Google Scholar] [CrossRef] [PubMed]
- Pittner, B.T.; Shanafelt, T.D.; Kay, N.E.; Jelinek, D.F. CD38 Expression Levels in Chronic Lymphocytic Leukemia B Cells Are Associated with Activation Marker Expression and Differential Responses to Interferon Stimulation. Leukemia 2005, 19, 2264–2272. [Google Scholar] [CrossRef]
- Le Gars, M.; Seiler, C.; Kay, A.W.; Bayless, N.L.; Sola, E.; Starosvetsky, E.; Moore, L.; Shen-Orr, S.S.; Aziz, N.; Khatri, P.; et al. CD38 Contributes to Human Natural Killer Cell Responses through a Role in Immune Synapse Formation. bioRxiv 2019, 349084. [Google Scholar] [CrossRef]
- Benito, J.M.; López, M.; Lozano, S.; Martinez, P.; González-Lahoz, J.; Soriano, V. CD38 Expression on CD8 T Lymphocytes as a Marker of Residual Virus Replication in Chronically HIV-Infected Patients Receiving Antiretroviral Therapy. AIDS Res. Hum. Retrovir. 2004, 20, 227–233. [Google Scholar] [CrossRef]
- Cannizzo, E.S.; Bellistrì, G.M.; Casabianca, A.; Tincati, C.; Iannotti, N.; Barco, A.; Orlandi, C.; Monforte, A.D.A.; Marchetti, G. Immunophenotype and Function of CD38-Expressing CD4+ and CD8+ T Cells in HIV-Infected Patients Undergoing Suppressive Combination Antiretroviral Therapy. J. Infect. Dis. 2015, 211, 1511–1513. [Google Scholar] [CrossRef]
- Tohgos, A.; Takasawa, S.; Noguchi, N.; Koguma, T.; Nata, K.; Sugimoto, T.; Furuya, Y.; Yonekura, H.; Okamotos, H. Essential Cysteine Residues for Cyclic ADP-Ribose Synthesis and Hydrolysis by CD38*. J. Biol. Chem. 1994, 269, 28555–28557. [Google Scholar] [CrossRef]
- Lanner, J.T.; Georgiou, D.K.; Joshi, A.D.; Hamilton, S.L. Ryanodine Receptors: Structure, Expression, Molecular Details, and Function in Calcium Release. Cold Spring Harb. Perspect. Biol. 2010, 2, a003996. [Google Scholar] [CrossRef]
- Galione, A.; Sethi, J. Cyclic ADP-Ribose and Calcium Signaling. In Biochemistry of Smooth Muscle Contraction; Academic Press: Cambridge, MA, USA, 1996; pp. 295–307.e. [Google Scholar]
- Malavasi, F.; Funaro, A.; Roggero, S.; Horenstein, A.; Calosso, L.; Mehta, K. Human CD38: A Glycoprotein in Search of a Function. Immunol. Today 1994, 15, 95–97. [Google Scholar] [CrossRef]
- Deaglio, S.; Aydin, S.; Grand, M.M.; Vaisitti, T.; Bergui, L.; D’Arena, G.; Chiorino, G.; Malavasi, F. CD38/CD31 Interactions Activate Genetic Pathways Leading to Proliferation and Migration in Chronic Lymphocytic Leukemia Cells. Mol. Med. 2010, 16, 87–91. [Google Scholar] [CrossRef]
- Lee, H.C.; Deng, Q.W.; Zhao, Y.J. The Calcium Signaling Enzyme CD38—A Paradigm for Membrane Topology Defining Distinct Protein Functions. Cell Calcium 2022, 101, 102514. [Google Scholar] [CrossRef]
- Zambello, R.; Barilà, G.; Manni, S.; Piazza, F.; Semenzato, G. NK Cells and CD38: Implication for (Immuno)Therapy in Plasma Cell Dyscrasias. Cells 2020, 9, 768. [Google Scholar] [CrossRef]
- Fedele, G.; Frasca, L.; Palazzo, R.; Ferrero, E.; Malavasi, F.; Ausiello, C.M. CD38 Is Expressed on Human Mature Monocyte-Derived Dendritic Cells and Is Functionally Involved in CD83 Expression and IL-12 Induction. Eur. J. Immunol. 2004, 34, 1342–1350. [Google Scholar] [CrossRef]
- Fujita, T.; Kantarci, A.; Warbington, M.L.; Zawawi, K.H.; Hasturk, H.; Kurihara, H.; Van Dyke, T.E. CD38 Expression in Neutrophils from Patients with Localized Aggressive Periodontitis. J. Periodontol. 2005, 76, 1960–1965. [Google Scholar] [CrossRef]
- Clavarino, G.; Delouche, N.; Vettier, C.; Laurin, D.; Pernollet, M.; Raskovalova, T.; Cesbron, J.Y.; Dumestre-Pérard, C.; Jacob, M.C. Novel Strategy for Phenotypic Characterization of Human B Lymphocytes from Precursors to Effector Cells by Flow Cytometry. PLoS ONE 2016, 11, e0162209. [Google Scholar] [CrossRef]
- Deterre, P.; Berthelier, V.; Bauvois, B.; Dalloul, A.; Schuber, F.; Lund, F. CD38 in T- and B-Cell Functions. Chem. Immunol. 2000, 75, 146–168. [Google Scholar] [CrossRef]
- Chini, E.N.; Chini, C.C.S.; Espindola Netto, J.M.; de Oliveira, G.C.; van Schooten, W. The Pharmacology of CD38/NADase: An Emerging Target in Cancer and Diseases of Aging. Trends Pharm. Sci. 2018, 39, 424–436. [Google Scholar] [CrossRef]
- Piedra-Quintero, Z.L.; Wilson, Z.; Nava, P.; Guerau-de-Arellano, M. CD38: An Immunomodulatory Molecule in Inflammation and Autoimmunity. Front. Immunol. 2020, 11, 597959. [Google Scholar] [CrossRef]
- Van De Donk, N.W.C.J.; Richardson, P.G.; Malavasi, F. CD38 Antibodies in Multiple Myeloma: Back to the Future. Blood 2018, 131, 13–29. [Google Scholar] [CrossRef] [Green Version]
- Fernàndez, J.E.; Deaglio, S.; Donati, D.; Beusan, I.S.; Corno, F.; Aranega, A.; Forni, M.; Falini, B.; Malavasi, F. Analysis of the Distribution of Human CD38 and of Its Ligand CD31 in Normal Tissues. J. Biol. Regul. Homeost. Agents 1998, 12, 81–91. [Google Scholar] [PubMed]
- Martin, T.G.; Corzo, K.; Chiron, M.; van de Velde, H.; Abbadessa, G.; Campana, F.; Solanki, M.; Meng, R.; Lee, H.; Wiederschain, D.; et al. Therapeutic Opportunities with Pharmacological Inhibition of CD38 with Isatuximab. Cells 2019, 8, 1522. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, P.; Mittelbrunn, M.; De La Fuente, H.; Perez-Martinez, M.; Garcfa-Perez, A.; Ariza-Veguillas, A.; Malavasi, F.; Zubiaur, M.; Sanchez-Madrid, F.; Sancho, J. Antigen-Induced Clustering of Surface CD38 and Recruitment of Intracellular CD38 to the Immunologic Synapse. Blood 2008, 111, 3653–3664. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhao, Y.J.; Li, W.H.; Hou, Y.N.; Li, T.; Zhao, Z.Y.; Fang, C.; Li, S.L.; Lee, H.C. Cytosolic Interaction of Type III Human CD38 with CIB1 Modulates Cellular Cyclic ADP-Ribose Levels. Proc. Natl. Acad. Sci. USA 2017, 114, 8283–8288. [Google Scholar] [CrossRef] [PubMed]
- Shrimp, J.H.; Hu, J.; Dong, M.; Wang, B.S.; Macdonald, R.; Jiang, H.; Hao, Q.; Yen, A.; Lin, H. Revealing CD38 Cellular Localization Using a Cell Permeable, Mechanism-Based Fluorescent Small-Molecule Probe. J. Am. Chem Soc. 2014, 136, 5656–5663. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.J.; Lam, C.M.C.; Lee, H.C. The Membrane-Bound Enzyme CD38 Exists in Two Opposing Orientations. Sci. Signal. 2012, 5, ra67. [Google Scholar] [CrossRef] [PubMed]
- Grozio, A.; Sociali, G.; Sturla, L.; Caffa, I.; Soncini, D.; Salis, A.; Raffaelli, N.; De Flora, A.; Nencioni, A.; Bruzzone, S. CD73 Protein as a Source of Extracellular Precursors for Sustained NAD+ Biosynthesis in FK866-Treated Tumor Cells. J. Biol. Chem. 2013, 288, 25938–25949. [Google Scholar] [CrossRef]
- Camacho-Pereira, J.; Tarragó, M.G.; Chini, C.C.S.; Nin, V.; Escande, C.; Warner, G.M.; Puranik, A.S.; Schoon, R.A.; Reid, J.M.; Galina, A.; et al. CD38 Dictates Age-Related NAD Decline and Mitochondrial Dysfunction through an SIRT3-Dependent Mechanism. Cell Metab. 2016, 23, 1127–1139. [Google Scholar] [CrossRef]
- Chini, C.C.S.; Peclat, T.R.; Warner, G.M.; Kashyap, S.; Espindola-Netto, J.M.; de Oliveira, G.C.; Gomez, L.S.; Hogan, K.A.; Tarragó, M.G.; Puranik, A.S.; et al. CD38 Ecto-Enzyme in Immune Cells Is Induced during Aging and Regulates NAD + and NMN Levels. Nat. Metab. 2020, 2, 1284–1304. [Google Scholar] [CrossRef]
- Yoshino, J.; Baur, J.A.; Imai, S. ichiro NAD + Intermediates: The Biology and Therapeutic Potential of NMN and NR. Cell Metab. 2018, 27, 513–528. [Google Scholar] [CrossRef] [Green Version]
- Hogan, K.A.; Chini, C.C.S.; Chini, E.N. The Multi-Faceted Ecto-Enzyme CD38: Roles in Immunomodulation, Cancer, Aging, and Metabolic Diseases. Front. Immunol. 2019, 10, 1187. [Google Scholar] [CrossRef]
- Ning, L.G.; Shan, G.; Sun, Z.; Zhang, F.; Xu, C.; Lou, X.; Li, S.; Du, H.; Chen, H.; Xu, G. Quantitative Proteomic Analysis Reveals the Deregulation of Nicotinamide Adenine Dinucleotide Metabolism and CD38 in Inflammatory Bowel Disease. BioMed Res. Int. 2019, 2019, 3950628. [Google Scholar] [CrossRef]
- Morandi, F.; Airoldi, I.; Marimpietri, D.; Bracci, C.; Faini, A.C.; Gramignoli, R. CD38, a Receptor with Multifunctional Activities: From Modulatory Functions on Regulatory Cell Subsets and Extracellular Vesicles, to a Target for Therapeutic Strategies. Cells 2019, 8, 1527. [Google Scholar] [CrossRef]
- Morra, M.; Zubiaur, M.; Terhorst, C.; Sancho, J.; Malavasi, F. CD38 Is Functionally Dependent on the TCR/CD3 Complex in Human T Cells. FASEB J. 1998, 12, 581–592. [Google Scholar] [CrossRef]
- Deaglio, S.; Zubiaur, M.; Gregorini, A.; Bottarel, F.; Ausiello, C.M.; Dianzani, U.; Sancho, J.; Malavasi, F. Human CD38 and CD16 Are Functionally Dependent and Physically Associated in Natural Killer Cells. Blood 2002, 99, 2490–2498. [Google Scholar] [CrossRef]
- Martins, T.C.; Águast, A.P. A Role for CD45RBlow CD38+ T Cells and Costimulatory Pathways of T-Cell Activation in Protection of Non-Obese Diabetic (NOD) Mice from Diabetes. Immunology 1999, 96, 600–605. [Google Scholar] [CrossRef]
- Matalonga, J.; Glaria, E.; Bresque, M.; Escande, C.; Carbó, J.M.; Kiefer, K.; Vicente, R.; León, T.E.; Beceiro, S.; Pascual-García, M.; et al. The Nuclear Receptor LXR Limits Bacterial Infection of Host Macrophages through a Mechanism That Impacts Cellular NAD Metabolism. Cell Rep. 2017, 18, 1241–1255. [Google Scholar] [CrossRef]
- Chini, E. CD38 as a Regulator of Cellular NAD: A Novel Potential Pharmacological Target for Metabolic Conditions. Curr. Pharm. Des. 2009, 15, 57–63. [Google Scholar] [CrossRef]
- Malavasi, F.; Deaglio, S.; Funaro, A.; Ferrero, E.; Horenstein, A.L.; Ortolan, E.; Vaisitti, T.; Aydin, S. Evolution and Function of the ADP Ribosyl Cyclase/CD38 Gene Family in Physiology and Pathology. Physiol. Rev. 2008, 88, 841–886. [Google Scholar] [CrossRef]
- Bauvois, B.; Durant, L.; Laboureau, J.; Barthélémy, E.; Rouillard, D.; Boulla, G.; Deterre, P. Upregulation of CD38 Gene Expression in Leukemic B Cells by Interferon Types I and II. J. Interferon Cytokine Res. 1999, 19, 1059–1066. [Google Scholar] [CrossRef]
- Deaglio, S.; Morra, M.; Mallone, R.; Ausiello, C.M.; Prager, E.; Garbarino, G.; Dianzani, U.; Stockinger, H.; Malavasi, F. Human CD38 (ADP-Ribosyl Cyclase) Is a Counter-Receptor of CD31, an Ig Superfamily Member. J. Immunol. 1998, 160, 395–402. [Google Scholar]
- Partida-Sánchez, S.; Goodrich, S.; Kusser, K.; Oppenheimer, N.; Randall, T.D.; Lund, F.E. Regulation of Dendritic Cell Trafficking by the ADP-Ribosyl Cyclase CD38: Impact on the Development of Humoral Immunity. Immunity 2004, 20, 279–291. [Google Scholar] [CrossRef]
- Najibi, M.; Honwad, H.H.; Moreau, J.A.; Becker, S.M.; Irazoqui, J.E. A novel nox/phox-cd38-naadp-tfeb axis important for macrophage activation during bacterial phagocytosis. Autophagy 2022, 18, 124–141. [Google Scholar] [CrossRef] [PubMed]
- Roboon, J.; Hattori, T.; Ishii, H.; Takarada-Iemata, M.; Le, T.M.; Shiraishi, Y.; Ozaki, N.; Yamamoto, Y.; Sugawara, A.; Okamoto, H.; et al. Deletion of CD38 Suppresses Glial Activation and Neuroinflammation in a Mouse Model of Demyelination. Front. Cell. Neurosci. 2019, 13, 258. [Google Scholar] [CrossRef] [PubMed]
- Glaría, E.; Valledor, A.F. Roles of CD38 in the Immune Response to Infection. Cells 2020, 9, 228. [Google Scholar] [CrossRef] [PubMed]
- Vaishnani, J. Superantigen. Indian J. Derm. Venereol. Leprol. 2009, 75, 540–544. [Google Scholar] [CrossRef]
- Zilber, M.T.; Setterblad, N.; Vasselon, T.; Doliger, C.; Charron, D.; Mooney, N.; Gelin, C. MHC Class II/CD38/CD9: A Lipid-Raft-Dependent Signaling Complex in Human Monocytes. Blood 2005, 106, 3074–3081. [Google Scholar] [CrossRef]
- Lischke, T.; Heesch, K.; Schumacher, V.; Schneider, M.; Haag, F.; Koch-Nolte, F.; Mittrücker, H.W. CD38 Controls the Innate Immune Response against Listeria Monocytogenes. Infect. Immun. 2013, 81, 4091–4099. [Google Scholar] [CrossRef]
- Kidd, P. Th1/Th2 Balance: The Hypothesis, Its Limitations, and Implications for Health and Disease. Altern. Med. Rev. 2003, 8, 223–246. [Google Scholar]
- Chang, X.; Yue, L.; Liu, W.; Wang, Y.; Wang, L.; Xu, B.; Wang, Y.; Pan, J.; Yan, X. CD38 and E2F Transcription Factor 2 Have Uniquely Increased Expression in Rheumatoid Arthritis Synovial Tissues. Clin. Exp. Immunol. 2014, 176, 222–231. [Google Scholar] [CrossRef]
- Wang, H.; Fang, K.; Yan, W.; Chang, X. T-Cell Immune Imbalance in Rheumatoid Arthritis Is Associated with Alterations in NK Cells and NK-Like T Cells Expressing CD38. J. Innate Immun. 2021, 14, 148–166. [Google Scholar] [CrossRef]
- Peclat, T.R.; Shi, B.; Varga, J.; Chini, E.N. The NADase Enzyme CD38: An Emerging Pharmacological Target for Systemic Sclerosis, Systemic Lupus Erythematosus and Rheumatoid Arthritis. Curr. Opin. Rheumatol. 2020, 32, 488–496. [Google Scholar] [CrossRef]
- Pérez-Lara, J.C.; Espinosa, E.; Santos-Argumedo, L.; Romero-Ramírez, H.; López-Herrera, G.; García-García, F.; Sandoval-Montes, C.; Ortiz-Navarrete, V.; Flores-Muñoz, M.; Rodríguez-Alba, J.C. CD38 Correlates with an Immunosuppressive Treg Phenotype in Lupus-Prone Mice. Int. J. Mol. Sci. 2021, 22, 11977. [Google Scholar] [CrossRef]
- Katsuyama, E.; Suarez-Fueyo, A.; Bradley, S.J.; Mizui, M.; Marin, A.V.; Mulki, L.; Krishfield, S.; Malavasi, F.; Yoon, J.; Sui, S.J.H.; et al. The CD38/NAD/SIRTUIN1/EZH2 Axis Mitigates Cytotoxic CD8 T Cell Function and Identifies Patients with SLE Prone to Infections. Cell Rep. 2020, 30, 112–123.e4. [Google Scholar] [CrossRef]
- Lokhorst, H.M.; Plesner, T.; Laubach, J.P.; Nahi, H.; Gimsing, P.; Hansson, M.; Minnema, M.C.; Lassen, U.; Krejcik, J.; Palumbo, A.; et al. Targeting CD38 with Daratumumab Monotherapy in Multiple Myeloma. N. Engl. J. Med. 2015, 373, 1207–1219. [Google Scholar] [CrossRef]
- de Weers, M.; Tai, Y.-T.; van der Veer, M.S.; Bakker, J.M.; Vink, T.; Jacobs, D.C.H.; Oomen, L.A.; Peipp, M.; Valerius, T.; Slootstra, J.W.; et al. Daratumumab, a Novel Therapeutic Human CD38 Monoclonal Antibody, Induces Killing of Multiple Myeloma and Other Hematological Tumors. J. Immunol. 2011, 186, 1840–1848. [Google Scholar] [CrossRef]
- Horenstein, A.L.; Chillemi, A.; Quarona, V.; Zito, A.; Roato, I.; Morandi, F.; Marimpietri, D.; Bolzoni, M.; Toscani, D.; Oldham, R.J.; et al. NAD+-Metabolizing Ectoenzymes in Remodeling Tumor–Host Interactions: The Human Myeloma Model. Cells 2015, 4, 520. [Google Scholar] [CrossRef]
- Feng, X.; Zhang, L.; Acharya, C.; An, G.; Wen, K.; Qiu, L.; Munshi, N.C.; Tai, Y.T.; Anderson, K.C. Targeting CD38 Suppresses Induction and Function of T Regulatory Cells to Mitigate Immunosuppression in Multiple Myeloma. Clin. Cancer Res. 2017, 23, 4290–4300. [Google Scholar] [CrossRef]
- Nijhof, I.S.; Casneuf, T.; Van Velzen, J.; Van Kessel, B.; Axel, A.E.; Syed, K.; Groen, R.W.J.; Van Duin, M.; Sonneveld, P.; Minnema, M.C.; et al. CD38 Expression and Complement Inhibitors Affect Response and Resistance to Daratumumab Therapy in Myeloma. Blood 2016, 128, 959–970. [Google Scholar] [CrossRef]
- Deckert, J.; Wetzel, M.C.; Bartle, L.M.; Skaletskaya, A.; Goldmacher, V.S.; Vallée, F.; Zhou-Liu, Q.; Ferrari, P.; Pouzieux, S.; Lahoute, C.; et al. SAR650984, a Novel Humanized CD38-Targeting Antibody, Demonstrates Potent Antitumor Activity in Models of Multiple Myeloma and Other CD38+ Hematologic Malignancies. Clin. Cancer Res. 2014, 20, 4574–4583. [Google Scholar] [CrossRef]
- Martin, T.; Baz, R.; Benson, D.M.; Lendvai, N.; Wolf, J.; Munster, P.; Lesokhin, A.M.; Wack, C.; Charpentier, E.; Campana, F.; et al. A Phase 1b Study of Isatuximab plus Lenalidomide and Dexamethasone for Relapsed/Refractory Multiple Myeloma. Blood 2017, 129, 3294–3303. [Google Scholar] [CrossRef] [PubMed]
- Smithson, G.; Zalevsky, J.; Korver, W.; Roepcke, S.; McLean, L. CD38+ Cell Depletion with TAK-079 Reduces Arthritis in a Cynomolgus Collagen-Induced Arthritis (CIA) Model. J. Immunol. 2017, 198, 127.17. [Google Scholar]
- Ugamraj, H.S.; Dang, K.; Ouisse, L.-H.; Buelow, B.; Chini, E.N.; Castello, G.; Allison, J.; Clarke, S.C.; Davison, L.M.; Buelow, R.; et al. TNB-738, a Biparatopic Antibody, Boosts Intracellular NAD+ by Inhibiting CD38 Ecto-Enzyme Activity. MAbs 2022, 14, 2095949. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, S. Isatuximab: First Approval. Drugs 2020, 80, 905–912. [Google Scholar] [CrossRef]
- Richardson, P.G.; Beksaç, M.; Špička, I.; Mikhael, J. Isatuximab for the Treatment of Relapsed/Refractory Multiple Myeloma. Expert Opin. Biol. 2020, 20, 1395–1404. [Google Scholar] [CrossRef]
- Moreno, L.; Perez, C.; Zabaleta, A.; Manrique, I.; Alignani, D.; Ajona, D.; Blanco, L.; Lasa, M.; Maiso, P.; Rodriguez, I.; et al. The Mechanism of Action of the Anti-CD38 Monoclonal Antibody Isatuximab in Multiple Myeloma. Clin. Cancer Res. 2019, 25, 3176–3187. [Google Scholar] [CrossRef]
- Krejcik, J.; Casneuf, T.; Nijhof, I.S.; Verbist, B.; Bald, J.; Plesner, T.; Syed, K.; Liu, K.; Van De Donk, N.W.C.J.; Weiss, B.M.; et al. Daratumumab Depletes CD38+ Immune Regulatory Cells, Promotes T-Cell Expansion, and Skews T-Cell Repertoire in Multiple Myeloma. Blood 2016, 128, 384–394. [Google Scholar] [CrossRef]
- van de Donk, N.W.C.J. Immunomodulatory Effects of CD38-Targeting Antibodies. Immunol. Lett. 2018, 199, 16–22. [Google Scholar] [CrossRef]
- Li, Y.; Yang, R.; Chen, L.; Wu, S. CD38 as an Immunomodulator in Cancer. Future Oncol. 2020, 16, 2853–2861. [Google Scholar] [CrossRef]
- Kassem, S.; Diallo, B.K.; El-Murr, N.; Carrié, N.; Tang, A.; Fournier, A.; Bonnevaux, H.; Nicolazzi, C.; Cuisinier, M.; Arnould, I.; et al. SAR442085, a Novel Anti-CD38 Antibody with Enhanced Antitumor Activity against Multiple Myeloma. Blood 2022, 139, 1160–1176. [Google Scholar] [CrossRef]
- Chatterjee, S.; Daenthanasanmak, A.; Chakraborty, P.; Wyatt, M.W.; Dhar, P.; Selvam, S.P.; Fu, J.; Zhang, J.; Nguyen, H.; Kang, I.; et al. CD38-NAD+Axis Regulates Immunotherapeutic Anti-Tumor T Cell Response. Cell Metab. 2018, 27, 85–100. [Google Scholar] [CrossRef]
- Chini, C.C.S.; Tarragó, M.G.; Chini, E.N. NAD and the Aging Process: Role in Life, Death and Everything in Between. Mol. Cell Endocrinol 2017, 455, 62–74. [Google Scholar] [CrossRef]
- Schultz, M.B.; Sinclair, D.A. Why NAD(+) Declines during Aging: It’s Destroyed. Cell Metab. 2016, 23, 965–966. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Xue, X.; Zhang, L.; Zhang, L.; Liu, Z. Comparative Analysis of Pharmacophore Features and Quantitative Structure-Activity Relationships for CD38 Covalent and Non-Covalent Inhibitors. Chem. Biol. Drug Des. 2015, 86, 1411–1424. [Google Scholar] [CrossRef]
- Liu, Z.; Graeff, R.M.; Jin, H.; Zhang, L.; Zhang, L. Studies on CD38 Inhibitors and Their Application to CADPR-Mediated Ca2 Signaling. Messenger 2013, 2, 19–32. [Google Scholar] [CrossRef]
- Boslett, J.; Hemann, C.; Zhao, Y.J.; Lee, H.C.; Zweier, J.L. Luteolinidin Protects the Postischemic Heart through CD38 Inhibition with Preservation of NAD(P)(H). J. Pharmacol. Exp. Ther. 2017, 361, 99. [Google Scholar] [CrossRef]
- Shu, B.; Feng, Y.; Gui, Y.; Lu, Q.; Wei, W.; Xue, X.; Sun, X.; He, W.; Yang, J.; Dai, C. Blockade of CD38 Diminishes Lipopolysaccharide-Induced Macrophage Classical Activation and Acute Kidney Injury Involving NF-ΚB Signaling Suppression. Cell Signal. 2018, 42, 249–258. [Google Scholar] [CrossRef]
- Blacher, E.; Ben Baruch, B.; Levy, A.; Geva, N.; Green, K.D.; Garneau-Tsodikova, S.; Fridman, M.; Stein, R. Inhibition of Glioma Progression by a Newly Discovered CD38 Inhibitor. Int. J. Cancer 2015, 136, 1422–1433. [Google Scholar] [CrossRef]
- Blacher, E.; Levy, A.; Baruch, B.; Green, K.D.; Garneau-Tsodikova, S.; Fridman, M.; Stein, R. Targeting CD38 in the Tumor Microenvironment: A Novel Approach to Treat Glioma. Cancer Cell Microenviron. 2015, 2, 486. [Google Scholar] [CrossRef]
- Schiavoni, I.; Scagnolari, C.; Horenstein, A.L.; Leone, P.; Pierangeli, A.; Malavasi, F.; Ausiello, C.M.; Fedele, G. CD38 Modulates Respiratory Syncytial Virus-driven Proinflammatory Processes in Human Monocyte-derived Dendritic Cells. Immunology 2018, 154, 122. [Google Scholar] [CrossRef]
- Deshpande, D.A.; Guedes, A.G.P.; Lund, F.E.; Subramanian, S.; Walseth, T.F.; Kannan, M.S. CD38 in the Pathogenesis of Allergic Airway Disease: Potential Therapeutic Targets. Pharmacol. Ther. 2017, 172, 116. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Feng, K.; Tong, C.; Jia, H.; Liu, Y.; Wang, Y.; Ti, D.; Yang, Q.; Wu, Z.; Han, W. Efficiency and Side Effects of Anti-CD38 CAR T Cells in an Adult Patient with Relapsed B-ALL after Failure of Bi-Specific CD19/CD22 CAR T Cell Treatment. Cell. Mol. Immunol. 2020, 17, 430–432. [Google Scholar] [CrossRef] [PubMed]
- Cui, Q.; Qian, C.; Xu, N.; Kang, L.; Dai, H.; Cui, W.; Song, B.; Yin, J.; Li, Z.; Zhu, X.; et al. CD38-Directed CAR-T Cell Therapy: A Novel Immunotherapy Strategy for Relapsed Acute Myeloid Leukemia after Allogeneic Hematopoietic Stem Cell Transplantation. J. Hematol. Oncol. 2021, 14, 82. [Google Scholar] [CrossRef] [PubMed]
- Roex, G.; Timmers, M.; Wouters, K.; Campillo-Davo, D.; Flumens, D.; Schroyens, W.; Chu, Y.; Berneman, Z.N.; Lion, E.; Luo, F.; et al. Safety and Clinical Efficacy of BCMA CAR-T-Cell Therapy in Multiple Myeloma. J. Hematol. Oncol. 2020, 13, 164. [Google Scholar] [CrossRef]
- Mei, H.; Li, C.; Jiang, H.; Zhao, X.; Huang, Z.; Jin, D.; Guo, T.; Kou, H.; Liu, L.; Tang, L.; et al. A Bispecific CAR-T Cell Therapy Targeting BCMA and CD38 in Relapsed or Refractory Multiple Myeloma. J. Hematol. Oncol. 2021, 14, 930. [Google Scholar] [CrossRef]
- Naik, J.; Themeli, M.; de Jong-Korlaar, R.; Ruiter, R.W.J.; Poddighe, P.J.; Yuan, H.; de Bruijn, J.D.; Ossenkoppele, G.J.; Zweegman, S.; Smit, L.; et al. CD38 as a Therapeutic Target for Adult Acute Myeloid Leukemia and T-Cell Acute Lymphoblastic Leukemia. Haematologica 2019, 104, e100–e103. [Google Scholar] [CrossRef]
- Gudgin, E.J.; Huntly, B.J.P. Acute Myeloid Leukemia: Leukemia Stem Cells Write a Prognostic Signature. Stem Cell Res. Ther. 2011, 2, 21. [Google Scholar] [CrossRef]
- Tang, L.; Bergevoet, S.M.; Gilissen, C.; de Witte, T.; Jansen, J.H.; van der Reijden, B.A.; Raymakers, R.A.P. Hematopoietic Stem Cells Exhibit a Specific ABC Transporter Gene Expression Profile Clearly Distinct from Other Stem Cells. BMC Pharm. 2010, 10, 12. [Google Scholar] [CrossRef]
- de Jonge-Peeters, S.D.P.W.M.; Kuipers, F.; de Vries, E.G.E.; Vellenga, E. ABC Transporter Expression in Hematopoietic Stem Cells and the Role in AML Drug Resistance. Crit. Rev. Oncol. Hematol. 2007, 62, 214–226. [Google Scholar] [CrossRef]
- Zeijlemaker, W.; Grob, T.; Meijer, R.; Hanekamp, D.; Kelder, A.; Carbaat-Ham, J.C.; Oussoren-Brockhoff, Y.J.M.; Snel, A.N.; Veldhuizen, D.; Scholten, W.J.; et al. CD34+CD38− Leukemic Stem Cell Frequency to Predict Outcome in Acute Myeloid Leukemia. Leukemia 2018, 33, 1102–1112. [Google Scholar] [CrossRef]
- Witte, K.E.; Ahlers, J.; Schäfer, I.; André, M.; Kerst, G.; Scheel-Walter, H.G.; Schwarze, C.P.; Pfeiffer, M.; Lang, P.; Handgretinger, R.; et al. High Proportion of Leukemic Stem Cells at Diagnosis Is Correlated with Unfavorable Prognosis in Childhood Acute Myeloid Leukemia. Pediatr. Hematol. Oncol. 2011, 28, 91–99. [Google Scholar] [CrossRef]
- Ebinger, M.; Witte, K.E.; Ahlers, J.; Schäfer, I.; André, M.; Kerst, G.; Scheel-Walter, H.G.; Lang, P.; Handgretinger, R. High Frequency of Immature Cells at Diagnosis Predicts High Minimal Residual Disease Level in Childhood Acute Lymphoblastic Leukemia. Leuk. Res. 2010, 34, 1139–1142. [Google Scholar] [CrossRef]
- Farber, M.; Chen, Y.; Arnold, L.; Möllmann, M.; Boog-Whiteside, E.; Lin, Y.A.; Reinhardt, H.C.; Dührsen, U.; Hanoun, M. Targeting CD38 in Acute Myeloid Leukemia Interferes with Leukemia Trafficking and Induces Phagocytosis. Sci. Rep. 2021, 11, 22062. [Google Scholar] [CrossRef]
- Wynne, J.; Stock, W. “Dar”-Ing to Target CD38 in T-ALL. Blood 2018, 131, 948–949. [Google Scholar] [CrossRef]
- Li, X.M.; Zhang, L.P.; Wang, Y.Z.; Lu, A.D.; Chang, Y.; Zhu, H.H.; Qin, Y.Z.; Lai, Y.Y.; Kong, Y.; Huang, X.J.; et al. CD38+ CD58− Is an Independent Adverse Prognostic Factor in Paediatric Philadelphia Chromosome Negative B Cell Acute Lymphoblastic Leukaemia Patients. Leuk. Res. 2016, 43, 33–38. [Google Scholar] [CrossRef]
- Ganzel, C.; Kharit, M.; Duksin, C.; Rowe, J.M. Daratumumab for Relapsed/Refractory Philadelphia-Positive Acute Lymphoblastic Leukemia. Haematologica 2018, 103, e489–e490. [Google Scholar] [CrossRef]
- Bride, K.L.; Vincent, T.L.; Im, S.Y.; Aplenc, R.; Barrett, D.M.; Carroll, W.L.; Carson, R.; Dai, Y.; Devidas, M.; Dunsmore, K.P.; et al. Preclinical Efficacy of Daratumumab in T-Cell Acute Lymphoblastic Leukemia. Blood 2018, 131, 995–999. [Google Scholar] [CrossRef]
- Karawajew, L.; Dworzak, M.; Ratei, R.; Rhein, P.; Gaipa, G.; Buldini, B.; Basso, G.; Hrusak, O.; Ludwig, W.D.; Henze, G.; et al. Minimal Residual Disease Analysis by Eight-Color Flow Cytometry in Relapsed Childhood Acute Lymphoblastic Leukemia. Haematologica 2015, 100, 935–944. [Google Scholar] [CrossRef]
- Malavasi, F.; Deaglio, S.; Damle, R.; Cutrona, G.; Ferrarini, M.; Chiorazzi, N. CD38 and Chronic Lymphocytic Leukemia: A Decade Later. Blood 2011, 118, 3470–3478. [Google Scholar] [CrossRef]
- Yang, S.M.; Li, J.Y.; Gale, R.P.; Huang, X.J. The Mystery of Chronic Lymphocytic Leukemia (CLL): Why Is It Absent in Asians and What Does This Tell Us about Etiology, Pathogenesis and Biology? Blood Rev. 2015, 29, 205–213. [Google Scholar] [CrossRef]
- Hallek, M.; Cheson, B.D.; Catovsky, D.; Caligaris-Cappio, F.; Dighiero, G.; Döhner, H.; Hillmen, P.; Keating, M.; Montserrat, E.; Chiorazzi, N.; et al. IwCLL Guidelines for Diagnosis, Indications for Treatment, Response Assessment, and Supportive Management of CLL. Blood 2018, 131, 2745–2760. [Google Scholar] [CrossRef]
- Morilla, A.; Gonzalez de Castro, D.; Del Giudice, I.; Osuji, N.; Else, M.; Morilla, R.; Brito Babapulle, V.; Rudenko, H.; Matutes, E.; Dearden, C.; et al. Combinations of ZAP-70, CD38 and IGHV Mutational Status as Predictors of Time to First Treatment in CLL. Leuk. Lymphoma 2008, 49, 2108–2115. [Google Scholar] [CrossRef]
- Raponi, S.; Ilari, C.; Della Starza, I.; Cappelli, L.V.; Cafforio, L.; Piciocchi, A.; Arena, V.; Mariglia, P.; Mauro, F.R.; Gentile, M.; et al. Redefining the Prognostic Likelihood of Chronic Lymphocytic Leukaemia Patients with Borderline Percentage of Immunoglobulin Variable Heavy Chain Region Mutations. Br. J. Haematol. 2020, 189, 853–859. [Google Scholar] [CrossRef]
- Cramer, P.; Hallek, M. Prognostic Factors in Chronic Lymphocytic Leukemia—What Do We Need to Know? Nat. Rev. Clin. Oncol. 2010, 8, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Greiner, S.M.; Märklin, M.; Holzmayer, S.; Kaban, K.; Meyer, S.; Hinterleitner, C.; Tandler, C.; Hagelstein, I.; Jung, G.; Salih, H.R.; et al. Identification of CD105 (Endoglin) as Novel Risk Marker in CLL. Ann. Hematol. 2022, 101, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, H.; Li, P.F.; Lu, Y.; Xia, Z.J.; Huang, H.Q.; Zhang, Y.J. CD38 Expression Predicts Poor Prognosis and Might Be a Potential Therapy Target in Extranodal NK/T Cell Lymphoma, Nasal Type. Ann. Hematol. 2015, 94, 1381–1388. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, N.; Nee, A.H.F.; Chooi, J.Y.; Toh, S.H.M.; Chung, T.H.; Selvarajan, V.; Fan, S.; Ng, S.B.; Poon, M.; Chan, E.; et al. Determinants of Response to Daratumumab in Epstein-Barr Virus-Positive Natural Killer and T-Cell Lymphoma. J. Immunother. Cancer 2021, 9, e002123. [Google Scholar] [CrossRef] [PubMed]
- Dimopoulos, M.A.; Moreau, P.; Terpos, E.; Mateos, M.V.; Zweegman, S.; Cook, G.; Delforge, M.; Hájek, R.; Schjesvold, F.; Cavo, M.; et al. Multiple Myeloma: EHA-ESMO Clinical Practice Guidelines for Diagnosis, Treatment and Follow-Up†. Ann. Oncol. 2021, 32, 309–322. [Google Scholar] [CrossRef]
- Oostvogels, R.; Jak, M.; Raymakers, R.; Mous, R.; Minnema, M.C. Efficacy of Retreatment with Immunomodulatory Drugs and Proteasome Inhibitors Following Daratumumab Monotherapy in Relapsed and Refractory Multiple Myeloma Patients. Br. J. Haematol. 2018, 183, 60–67. [Google Scholar] [CrossRef]
- Gandhi, U.H.; Cornell, R.F.; Lakshman, A.; Gahvari, Z.J.; McGehee, E.; Jagosky, M.H.; Gupta, R.; Varnado, W.; Fiala, M.A.; Chhabra, S.; et al. Outcomes of Patients with Multiple Myeloma Refractory to CD38-Targeted Monoclonal Antibody Therapy. Leukemia 2019, 33, 2266–2275. [Google Scholar] [CrossRef]
- D’Agostino, M.; Mina, R.; Gay, F. Anti-CD38 Monoclonal Antibodies in Multiple Myeloma: Another Cook in the Kitchen? Lancet Haematol. 2020, 7, e355–e357. [Google Scholar] [CrossRef]
- Overdijk, M.B.; Jansen, J.H.M.; Nederend, M.; Lammerts van Bueren, J.J.; Groen, R.W.J.; Parren, P.W.H.I.; Leusen, J.H.W.; Boross, P. The Therapeutic CD38 Monoclonal Antibody Daratumumab Induces Programmed Cell Death via Fcγ Receptor-Mediated Cross-Linking. J. Immunol. 2016, 197, 807–813. [Google Scholar] [CrossRef]
- Overdijk, M.B.; Verploegen, S.; Bögels, M.; Van Egmond, M.; Lammerts Van Bueren, J.J.; Mutis, T.; Groen, R.W.J.; Breij, E.; Martens, A.C.M.; Bleeker, W.K.; et al. Antibody-Mediated Phagocytosis Contributes to the Anti-Tumor Activity of the Therapeutic Antibody Daratumumab in Lymphoma and Multiple Myeloma. MAbs 2015, 7, 311–320. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Hughes, T.; Zhang, J.; Caligiuri, M.A.; Benson, D.M.; Yu, J. Fratricide of NK Cells in Daratumumab Therapy for Multiple Myeloma Overcome by Ex Vivo-Expanded Autologous NK Cells. Clin. Cancer Res. 2018, 24, 4006–4017. [Google Scholar] [CrossRef]
- Casneuf, T.; Xu, X.S.; Adams, H.C.; Axel, A.E.; Chiu, C.; Khan, I.; Ahmadi, T.; Yan, X.; Lonial, S.; Plesner, T.; et al. Effects of Daratumumab on Natural Killer Cells and Impact on Clinical Outcomes in Relapsed or Refractory Multiple Myeloma. Blood Adv. 2017, 1, 2105–2114. [Google Scholar] [CrossRef]
- Patiño-Escobar, B.; Ramos, R.; Linares, M.; Mejía, A.; Alcalá, S. CD38: From Positive to Negative Expression after Daratumumab Treatment. Cureus 2020, 12, e7627. [Google Scholar] [CrossRef]
- Lonial, S.; Weiss, B.M.; Usmani, S.Z.; Singhal, S.; Chari, A.; Bahlis, N.J.; Belch, A.; Krishnan, A.; Vescio, R.A.; Mateos, M.V.; et al. Daratumumab Monotherapy in Patients with Treatment-Refractory Multiple Myeloma (SIRIUS): An Open-Label, Randomised, Phase 2 Trial. Lancet 2016, 387, 1551–1560. [Google Scholar] [CrossRef]
- Terpos, E. Multiple Myeloma: Clinical Updates From the American Society of Hematology Annual Meeting 2016. Clin. Lymphoma Myeloma Leuk. 2017, 17, 329–339. [Google Scholar] [CrossRef]
- Moreau, P.; Hulin, C.; Perrot, A.; Arnulf, B.; Belhadj, K.; Benboubker, L.; Béné, M.C.; Zweegman, S.; Caillon, H.; Caillot, D.; et al. Maintenance with Daratumumab or Observation Following Treatment with Bortezomib, Thalidomide, and Dexamethasone with or without Daratumumab and Autologous Stem-Cell Transplant in Patients with Newly Diagnosed Multiple Myeloma (CASSIOPEIA): An Open-Label, Randomised, Phase 3 Trial. Lancet Oncol. 2021, 22, 1378–1390. [Google Scholar] [CrossRef]
- Bahlis, N.J.; Baz, R.; Harrison, S.J.; Quach, H.; Ho, S.J.; Vangsted, A.J.; Plesner, T.; Moreau, P.; Gibbs, S.D.; Coppola, S.; et al. Phase I Study of Venetoclax Plus Daratumumab and Dexamethasone, With or Without Bortezomib, in Patients With Relapsed or Refractory Multiple Myeloma With and Without t(11;14). J. Clin. Oncol. 2021, 39, 3602–3612. [Google Scholar] [CrossRef]
- Mateos, M.-V.; Dimopoulos, M.A.; Cavo, M.; Suzuki, K.; Jakubowiak, A.; Knop, S.; Doyen, C.; Lucio, P.; Nagy, Z.; Kaplan, P.; et al. Daratumumab plus Bortezomib, Melphalan, and Prednisone for Untreated Myeloma. N. Engl. J. Med. 2018, 378, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Mateos, M.V.; Cavo, M.; Blade, J.; Dimopoulos, M.A.; Suzuki, K.; Jakubowiak, A.; Knop, S.; Doyen, C.; Lucio, P.; Nagy, Z.; et al. Overall Survival with Daratumumab, Bortezomib, Melphalan, and Prednisone in Newly Diagnosed Multiple Myeloma (ALCYONE): A Randomised, Open-Label, Phase 3 Trial. Lancet 2020, 395, 132–141. [Google Scholar] [CrossRef]
- Facon, T.; Kumar, S.; Plesner, T.; Orlowski, R.Z.; Moreau, P.; Bahlis, N.; Basu, S.; Nahi, H.; Hulin, C.; Quach, H.; et al. Daratumumab plus Lenalidomide and Dexamethasone for Untreated Myeloma. N. Engl. J. Med. 2019, 380, 2104–2115. [Google Scholar] [CrossRef] [PubMed]
- Voorhees, P.M.; Kaufman, J.L.; Laubach, J.P.; Sborov, D.W.; Reeves, B.; Rodriguez, C.; Chari, A.; Silbermann, R.; Costa, L.J.; Anderson, L.D., Jr.; et al. Daratumumab, Lenalidomide, Bortezomib, and Dexamethasone for Transplant-Eligible Newly Diagnosed Multiple Myeloma: The GRIFFIN Trial. Blood 2020, 136, 936–945. [Google Scholar] [CrossRef] [PubMed]
- Mateos, M.V.; Nahi, H.; Legiec, W.; Grosicki, S.; Vorobyev, V.; Spicka, I.; Hungria, V.; Korenkova, S.; Bahlis, N.; Flogegard, M.; et al. Subcutaneous versus Intravenous Daratumumab in Patients with Relapsed or Refractory Multiple Myeloma (COLUMBA): A Multicentre, Open-Label, Non-Inferiority, Randomised, Phase 3 Trial. Lancet Haematol. 2020, 7, e370–e380. [Google Scholar] [CrossRef]
- Dimopoulos, M.A.; Terpos, E.; Boccadoro, M.; Delimpasi, S.; Beksac, M.; Katodritou, E.; Moreau, P.; Baldini, L.; Symeonidis, A.; Bila, J.; et al. Daratumumab plus Pomalidomide and Dexamethasone versus Pomalidomide and Dexamethasone Alone in Previously Treated Multiple Myeloma (APOLLO): An Open-Label, Randomised, Phase 3 Trial. Lancet Oncol. 2021, 22, 801–812. [Google Scholar] [CrossRef]
- Attal, M.; Richardson, P.G.; Rajkumar, S.V.; San-Miguel, J.; Beksac, M.; Spicka, I.; Leleu, X.; Schjesvold, F.; Moreau, P.; Dimopoulos, M.A.; et al. Isatuximab plus Pomalidomide and Low-Dose Dexamethasone versus Pomalidomide and Low-Dose Dexamethasone in Patients with Relapsed and Refractory Multiple Myeloma (ICARIA-MM): A Randomised, Multicentre, Open-Label, Phase 3 Study. Lancet 2019, 394, 2096–2107. [Google Scholar] [CrossRef]
- Szemraj-Rogucka, Z.; Szemraj, J.; Grzybowska-Izydorczyk, O.; Robak, T.; Jamroziak, K. CD38 Gene Polymorphisms and Genetic Predisposition to Multiple Myeloma. Acta Haematol. Pol. 2013, 44, 58–62. [Google Scholar] [CrossRef]
- You, X.; Yang, Q.; Yan, K.; Wang, S.-R.; Huang, R.-R.; Wang, S.-Q.; Gao, C.-Y.; Li, L.; Lian, Z.-X. Multi-Omics Profiling Identifies Pathways Associated With CD8+ T-Cell Activation in Severe Aplastic Anemia. Front. Genet. 2022, 12, 790990. [Google Scholar] [CrossRef]
- Liszewski, M.K.; Kolev, M.; Le Friec, G.; Leung, M.; Bertram, P.G.; Fara, A.F.; Subias, M.; Pickering, M.C.; Drouet, C.; Meri, S.; et al. Intracellular Complement Activation Sustains T Cell Homeostasis and Mediates Effector Differentiation. Immunity 2013, 39, 1143–1157. [Google Scholar] [CrossRef]
- Hoff, P.; Maschmeyer, P.; Gaber, T.; Schütze, T.; Raue, T.; Schmidt-Bleek, K.; Dziurla, R.; Schellmann, S.; Lohanatha, F.L.; Röhner, E.; et al. Human Immune Cells’ Behavior and Survival under Bioenergetically Restricted Conditions in an in Vitro Fracture Hematoma Model. Cell Mol. Immunol. 2013, 10, 151–158. [Google Scholar] [CrossRef]
- Behzad, M.M.; Asnafi, A.A.; Jalalifar, M.A.; Moghtadaei, M.; Jaseb, K.; Saki, N. Cellular Expression of CD Markers in Immune Thrombocytopenic Purpura: Implications for Prognosis. APMIS 2018, 126, 523–532. [Google Scholar] [CrossRef]
- Kastritis, E.; Palladini, G.; Minnema, M.C.; Wechalekar, A.D.; Jaccard, A.; Lee, H.C.; Sanchorawala, V.; Gibbs, S.; Mollee, P.; Venner, C.P.; et al. Daratumumab-Based Treatment for Immunoglobulin Light-Chain Amyloidosis. N. Engl. J. Med. 2021, 385, 46–58. [Google Scholar] [CrossRef]
- Palladini, G.; Kastritis, E.; Maurer, M.S.; Zonder, J.; Minnema, M.C.; Wechalekar, A.D.; Jaccard, A.; Lee, H.C.; Bumma, N.; Kaufman, J.L.; et al. Daratumumab plus CyBorD for Patients with Newly Diagnosed AL Amyloidosis: Safety Run-in Results of ANDROMEDA. Blood 2020, 136, 71–80. [Google Scholar] [CrossRef]
- Dispenzieri, A.; Kastritis, E.; Wechalekar, A.D.; Schönland, S.O.; Kim, K.; Sanchorawala, V.; Landau, H.J.; Kwok, F.; Suzuki, K.; Comenzo, R.L.; et al. A Randomized Phase 3 Study of Ixazomib–Dexamethasone versus Physician’s Choice in Relapsed or Refractory AL Amyloidosis. Leukemia 2021, 36, 225–235. [Google Scholar] [CrossRef]
- Liu, Y.; Bian, T.; Zhang, Y.; Zheng, Y.; Zhang, J.; Zhou, X.; Xie, J. A Combination of LMO2 Negative and CD38 Positive Is Useful for the Diagnosis of Burkitt Lymphoma. Diagn. Pathol. 2019, 14, 100. [Google Scholar] [CrossRef]
- Chatzidimitriou, C.; Vassilakopoulos, T.P. Rapidly Deteriorating B-CLL at Presentation: Clonally Related Burkitt Leukemia as an Atypical Richter Transformation? Blood 2021, 137, 2272. [Google Scholar] [CrossRef]
- Kumar, D.; Prince, C.; Bennett, C.M.; Briones, M.; Lucas, L.; Russell, A.; Patel, K.; Chonat, S.; Graciaa, S.; Edington, H.; et al. T-Follicular Helper Cell Expansion and Chronic T-Cell Activation Are Characteristic Immune Anomalies in Evans Syndrome. Blood 2022, 139, 369–383. [Google Scholar] [CrossRef]
- Kallen, M.E.; Dulau-Florea, A.; Wang, W.; Calvo, K.R. Acquired and Germline Predisposition to Bone Marrow Failure: Diagnostic Features and Clinical Implications. Semin. Hematol. 2019, 56, 69–82. [Google Scholar] [CrossRef]
- Chaturvedi, V.; Marsh, R.A.; Zoref-Lorenz, A.; Owsley, E.; Chaturvedi, V.; Nguyen, T.C.; Goldman, J.R.; Henry, M.M.; Greenberg, J.N.; Ladisch, S.; et al. T-Cell Activation Profiles Distinguish Hemophagocytic Lymphohistiocytosis and Early Sepsis. Blood 2021, 137, 2337–2346. [Google Scholar] [CrossRef] [PubMed]
- Poret, N.; Fu, Q.; Guihard, S.; Cheok, M.; Miller, K.; Zeng, G.; Quesnel, B.; Troussard, X.; Galiègue-Zouitina, S.; Shelley, C.S. CD38 in Hairy Cell Leukemia Is a Marker of Poor Prognosis and a New Target for Therapy. Cancer Res. 2015, 75, 3902–3911. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Y.; Fuda, F.S.; Chen, W.; Karandikar, N.J. Notch1 in Primary Effusion Lymphoma: A Clinicopathological Study. Mod. Pathol. 2010, 23, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Armitage, J.O. The Aggressive Peripheral T-Cell Lymphomas: 2017. Am. J. Hematol. 2017, 92, 706–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taddesse-Heath, L.; Meloni-Ehrig, A.; Scheerle, J.; Kelly, J.C.; Jaffe, E.S. Plasmablastic Lymphoma with MYC Translocation: Evidence for a Common Pathway in the Generation of Plasmablastic Features. Mod. Pathol. 2010, 23, 991–999. [Google Scholar] [CrossRef]
- Rodig, S.J.; Vergilio, J.A.; Shahsafaei, A.; Dorfman, D.M. Characteristic Expression Patterns of TCL1, CD38, and CD44 Identify Aggressive Lymphomas Harboring a MYC Translocation. Am. J. Surg. Pathol. 2008, 32, 113–122. [Google Scholar] [CrossRef]
- Kastritis, E.; Leblond, V.; Dimopoulos, M.A.; Kimby, E.; Staber, P.; Kersten, M.J.; Tedeschi, A.; Buske, C. Waldenström’s Macroglobulinaemia: ESMO Clinical Practice Guidelines for Diagnosis, Treatment and Follow-Up†. Ann. Oncol. 2018, 29, iv41–iv50. [Google Scholar] [CrossRef]
- Ge, Y.; Long, Y.; Xiao, S.; Liang, L.; He, Z.; Yue, C.; Wei, X.; Zhou, Y. CD38 Affects the Biological Behavior and Energy Metabolism of Nasopharyngeal Carcinoma Cells. Int. J. Oncol. 2019, 54, 585–599. [Google Scholar] [CrossRef]
- Zucali, P.A.; Lin, C.-C.; Carthon, B.C.; Bauer, T.M.; Tucci, M.; Italiano, A.; Iacovelli, R.; Su, W.-C.; Massard, C.; Saleh, M.; et al. Targeting CD38 and PD-1 with Isatuximab plus Cemiplimab in Patients with Advanced Solid Malignancies: Results from a Phase I/II Open-Label, Multicenter Study. J. Immunother. Cancer 2022, 10, e003697. [Google Scholar] [CrossRef]
- Guo, C.; Crespo, M.; Gurel, B.; Dolling, D.; Rekowski, J.; Sharp, A.; Petremolo, A.; Sumanasuriya, S.; Rodrigues, D.N.; Ferreira, A.; et al. CD38 in Advanced Prostate Cancers. Eur. Urol. 2021, 79, 736–746. [Google Scholar] [CrossRef]
- Chmielewski, J.P.; Bowlby, S.C.; Wheeler, F.B.; Shi, L.; Sui, G.; Davis, A.L.; Howard, T.D.; D’Agostino, R.B.; Miller, L.D.; Sirintrapun, S.J.; et al. CD38 Inhibits Prostate Cancer Metabolism and Proliferation by Reducing Cellular NAD+ Pools. Mol. Cancer Res. 2018, 16, 1687–1700. [Google Scholar] [CrossRef]
- Karakasheva, T.A.; Dominguez, G.A.; Hashimoto, A.; Lin, E.W.; Chiu, C.; Sasser, K.; Lee, J.W.; Beatty, G.L.; Gabrilovich, D.I.; Rustgi, A.K. CD38+ M-MDSC Expansion Characterizes a Subset of Advanced Colorectal Cancer Patients. JCI Insight 2018, 3, e97022. [Google Scholar] [CrossRef]
- Ma, Y.; Jiang, J.; Ying, W. CD38 Mediates the Intracellular ATP Levels and Cell Survival of C6 Glioma Cells. Neuroreport 2014, 25, 569–573. [Google Scholar] [CrossRef]
- Lu, L.; Wang, J.; Yang, Q.; Xie, X.; Huang, Y. The Role of CD38 in HIV Infection. AIDS Res. Ther. 2021, 18, 11. [Google Scholar] [CrossRef]
- Židovec Lepej, S.; Vince, A.; Daković Rode, O.; Remenar, A.; Jeren, T. Increased Numbers of CD38 Molecules on Bright CD8+ T Lymphocytes in Infectious Mononucleosis Caused by Epstein–Barr Virus Infection. Clin. Exp. Immunol. 2003, 133, 384–390. [Google Scholar] [CrossRef]
- Booiman, T.; Wit, F.W.; Girigorie, A.F.; Maurer, I.; De Francesco, D.; Sabin, C.A.; Harskamp, A.M.; Prins, M.; Franceschi, C.; Deeks, S.G.; et al. Terminal Differentiation of T Cells Is Strongly Associated with CMV Infection and Increased in HIV-Positive Individuals on ART and Lifestyle Matched Controls. PLoS ONE 2017, 12, e0183357. [Google Scholar] [CrossRef]
- Hunt, P.W.; Martin, J.N.; Sinclair, E.; Epling, L.; Teague, J.; Jacobson, M.A.; Tracy, R.P.; Corey, L.; Deeks, S.G. Valganciclovir Reduces T Cell Activation in HIV-Infected Individuals With Incomplete CD4+ T Cell Recovery on Antiretroviral Therapy. J. Infect. Dis. 2011, 203, 1474–1483. [Google Scholar] [CrossRef]
- Shah, N.N.; Singavi, A.K.; Harrington, A. Daratumumab in Primary Effusion Lymphoma. N. Engl. J. Med. 2018, 379, 689–690. [Google Scholar] [CrossRef]
- Horenstein, A.L.; Faini, A.C.; Malavasi, F. CD38 in the Age of Covid-19: Amedical Perspective. Physiol. Rev. 2021, 101, 1457–1486. [Google Scholar] [CrossRef]
- Zeidler, J.D.; Kashyap, S.; Hogan, K.A.; Chini, E.N. Implications of the nadase cd38 in covid pathophysiology. Physiol. Rev. 2022, 102, 339–341. [Google Scholar] [CrossRef]
- Partida-Sánchez, S.; Cockayne, D.A.; Monard, S.; Jacobson, E.L.; Oppenheimer, N.; Garvy, B.; Kusser, K.; Goodrich, S.; Howard, M.; Harmsen, A.; et al. Cyclic ADP-Ribose Production by CD38 Regulates Intracellular Calcium Release, Extracellular Calcium Influx and Chemotaxis in Neutrophils and Is Required for Bacterial Clearance in Vivo. Nat. Med. 2001, 7, 1209–1216. [Google Scholar] [CrossRef]
- Nahi, H.; Chrobok, M.; Gran, C.; Lund, J.; Gruber, A.; Gahrton, G.; Ljungman, P.; Wagner, A.K.; Alici, E. Infectious Complications and NK Cell Depletion Following Daratumumab Treatment of Multiple Myeloma. PLoS ONE 2019, 14, e0211927. [Google Scholar] [CrossRef]
- al Saleh, A.S.; Sher, T.; Gertz, M.A. Multiple Myeloma in the Time of COVID-19. Acta Haematol. 2020, 143, 410–416. [Google Scholar] [CrossRef]
- Terpos, E.; Engelhardt, M.; Cook, G.; Gay, F.; Mateos, M.V.; Ntanasis-Stathopoulos, I.; van de Donk, N.W.C.J.; Avet-Loiseau, H.; Hajek, R.; Vangsted, A.J.; et al. Management of Patients with Multiple Myeloma in the Era of COVID-19 Pandemic: A Consensus Paper from the European Myeloma Network (EMN). Leukemia 2020, 34, 2000–2011. [Google Scholar] [CrossRef]
- Pimpinelli, F.; Marchesi, F.; Piaggio, G.; Giannarelli, D.; Papa, E.; Falcucci, P.; Pontone, M.; di Martino, S.; Laquintana, V.; la Malfa, A.; et al. Fifth-Week Immunogenicity and Safety of Anti-SARS-CoV-2 BNT162b2 Vaccine in Patients with Multiple Myeloma and Myeloproliferative Malignancies on Active Treatment: Preliminary Data from a Single Institution. J. Hematol. Oncol. 2021, 14, 81. [Google Scholar] [CrossRef]
- Bitoun, S.; Henry, J.; Vauloup-Fellous, C.; Dib, N.; Belkhir, R.; Mouna, L.; Joly, C.; Desjardins, D.; Bitu, M.; le Grand, R.; et al. Response to COVID-19 MRNA Vaccination in Multiple Myeloma Is Conserved but Impaired Compared to Controls. J. Hematol. Oncol. 2021, 14, 166. [Google Scholar] [CrossRef]
- Abdallah, A.; Mahmoudjafari, Z.; Atieh, T.; Ahmed, N.; Cui, W.; Shune, L.; Mohan, M.; McGuirk, J.; Remker, C.; Foss, M.; et al. Neutralizing Antibody Responses against SARS-CoV-2 in Patients with Plasma Cell Disorders Who Are on Active Treatment after Two Doses of MRNA Vaccination. Blood 2021, 138, 3804. [Google Scholar] [CrossRef]
- Zühlke, S.; Risnes, L.F.; Dahal-Koirala, S.; Christophersen, A.; Sollid, L.M.; Lundin, K.E.A. CD38 Expression on Gluten-Specific T Cells Is a Robust Marker of Gluten Re-Exposure in Coeliac Disease. United Eur. Gastroenterol. J. 2019, 7, 1337–1344. [Google Scholar] [CrossRef]
- Du, Y.; Zhang, H.; Guo, Y.; Song, K.; Zeng, L.; Chen, Y.; Xie, Z.; Li, R. CD38 Deficiency Up-Regulated IL-1β and MCP-1 through TLR4/ERK/NF-ΚB Pathway in Sepsis Pulmonary Injury. Microbes Infect. 2021, 23, 104845. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Du, Y.; Guo, Y.; Wang, Z.; Li, H.; Lv, Z.; Zeng, L.; Chen, Y.; Xie, Z.; Li, R. TLR4-NLRP3-GSDMD-Mediated Pyroptosis Plays an Important Role in Aggravated Liver Injury of CD38 -/- Sepsis Mice. J. Immunol Res. 2021, 2021, 104845. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Ma, L.; Li, R.; Huang, S.; Xie, R.; Chen, Y.; Zhao, B.; Fei, J.; Qu, H.; Chen, H.; et al. DC-SIGN Expression in Intestinal Epithelial Cells Regulates Sepsis-Associated Acute Intestinal Injury Via Activating ERK1/2-NF-ΚB/P65 Signaling. Shock 2019, 52, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Guerreiro, S.; Privat, A.L.; Bressac, L.; Toulorge, D. CD38 in Neurodegeneration and Neuroinflammation. Cells 2020, 9, 471. [Google Scholar] [CrossRef]
- Milo, R. Therapies for Multiple Sclerosis Targeting B Cells. Croat. Med. J. 2019, 60, 87–98. [Google Scholar] [CrossRef]
- Roboon, J.; Hattori, T.; Ishii, H.; Takarada-Iemata, M.; Nguyen, D.T.; Heer, C.D.; O’Meally, D.; Brenner, C.; Yamamoto, Y.; Okamoto, H.; et al. Inhibition of CD38 and Supplementation of Nicotinamide Riboside Ameliorate Lipopolysaccharide-Induced Microglial and Astrocytic Neuroinflammation by Increasing NAD. J. Neurochem. 2021, 158, 311–327. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, G.; Shin, H.J.; Hyun, J.W.; Kim, S.H.; Lee, E.; Kim, H.J. Restoration of Regulatory B Cell Deficiency Following Alemtuzumab Therapy in Patients with Relapsing Multiple Sclerosis. J. Neuroinflammation 2018, 15, 300. [Google Scholar] [CrossRef]
- Nakamura, M.; Matsuoka, T.; Chihara, N.; Miyake, S.; Sato, W.; Araki, M.; Okamoto, T.; Lin, Y.; Ogawa, M.; Murata, M.; et al. Differential Effects of Fingolimod on B-Cell Populations in Multiple Sclerosis. Mult. Scler. 2014, 20, 1371–1380. [Google Scholar] [CrossRef]
- Hawke, S.; Zinger, A.; Juillard, P.G.; Holdaway, K.; Byrne, S.N.; Grau, G.E. Selective Modulation of Trans-Endothelial Migration of Lymphocyte Subsets in Multiple Sclerosis Patients under Fingolimod Treatment. J. Neuroimmunol. 2020, 349, 577392. [Google Scholar] [CrossRef]
- Grützke, B.; Hucke, S.; Gross, C.C.; Herold, M.V.B.; Posevitz-Fejfar, A.; Wildemann, B.T.; Kieseier, B.C.; Dehmel, T.; Wiendl, H.; Klotz, L. Fingolimod Treatment Promotes Regulatory Phenotype and Function of B Cells. Ann. Clin. Transl. Neurol. 2015, 2, 119–130. [Google Scholar] [CrossRef]
- Pavelek, Z.; Vyšata, O.; Klímová, B.; Andrýs, C.; Vokurková, D.; Vališ, M. Lymphocytes in the Treatment with Interferon Beta-1 b. Mult. Scler. Relat. Disord. 2017, 18, 29–32. [Google Scholar] [CrossRef]
- Schubert, R.D.; Hu, Y.; Kumar, G.; Szeto, S.; Abraham, P.; Winderl, J.; Guthridge, J.M.; Pardo, G.; Dunn, J.; Steinman, L.; et al. IFN-β Treatment Requires B Cells for Efficacy in Neuroautoimmunity. J. Immunol. 2015, 194, 2110–2116. [Google Scholar] [CrossRef]
- Lundy, S.K.; Wu, Q.; Wang, Q.; Dowling, C.A.; Taitano, S.H.; Mao, G.; Mao-Draayer, Y. Dimethyl Fumarate Treatment of Relapsing-Remitting Multiple Sclerosis Influences B-Cell Subsets. Neurol. (R) Neuroimmunol. Neuroinflammation 2016, 3, e211. [Google Scholar] [CrossRef]
- Khodaverdian, S.; Dashtban-Moghadam, E.; Dabirmanesh, B.; Mirnajafi-Zadeh, J.; Taleb, M.; Khajeh, K.; Fathollahi, Y. CD38 and MGluR1 as Possible Signaling Molecules Involved in Epileptogenesis: A Potential Role for NAD+ Homeostasis. Brain Res. 2021, 1765, 147509. [Google Scholar] [CrossRef] [PubMed]
- Perry, D.J.; Titov, A.A.; Sobel, E.S.; Brusko, T.M.; Morel, L. Immunophenotyping Reveals Distinct Subgroups of Lupus Patients Based on Their Activated T Cell Subsets. Clin. Immunol. 2020, 221, 108602. [Google Scholar] [CrossRef] [PubMed]
- Ostendorf, L.; Burns, M.; Durek, P.; Heinz, G.A.; Heinrich, F.; Garantziotis, P.; Enghard, P.; Richter, U.; Biesen, R.; Schneider, U.; et al. Targeting CD38 with Daratumumab in Refractory Systemic Lupus Erythematosus. N. Engl. J. Med. 2020, 383, 1149–1155. [Google Scholar] [CrossRef] [PubMed]
- Cole, S.; Walsh, A.; Yin, X.; Wechalekar, M.D.; Smith, M.D.; Proudman, S.M.; Veale, D.J.; Fearon, U.; Pitzalis, C.; Humby, F.; et al. Integrative Analysis Reveals CD38 as a Therapeutic Target for Plasma Cell-Rich Pre-Disease and Established Rheumatoid Arthritis and Systemic Lupus Erythematosus. Arthritis Res. 2018, 20, 85. [Google Scholar] [CrossRef]
- Humbel, M.; Bellanger, F.; Fluder, N.; Horisberger, A.; Suffiotti, M.; Fenwick, C.; Ribi, C.; Comte, D. Restoration of NK Cell Cytotoxic Function With Elotuzumab and Daratumumab Promotes Elimination of Circulating Plasma Cells in Patients With SLE. Front. Immunol. 2021, 12, 645478. [Google Scholar] [CrossRef]
- Liu, J.; Tang, T.; Qu, Z.; Wang, L.; Si, R.; Wang, H.; Jiang, Y. Elevated Number of IL-21+ TFH and CD86+CD38+ B Cells in Blood of Renal Transplant Recipients with AMR under Conventional Immuno-Suppression. Int. J. Immunopathol. Pharmacol. 2022, 36, 20587384211048027. [Google Scholar] [CrossRef]
- Hulbert, A.L.; Pavlisko, E.N.; Palmer, S.M. Current Challenges and Opportunities in the Management of Antibody-Mediated Rejection in Lung Transplantation. Curr. Opin. Organ. Transpl. 2018, 23, 308–315. [Google Scholar] [CrossRef]
- Joher, N.; Matignon, M.; Grimbert, P. HLA Desensitization in Solid Organ Transplantation: Anti-CD38 to Across the Immunological Barriers. Front. Immunol. 2021, 12, 688301. [Google Scholar] [CrossRef]
- Spica, D.; Junker, T.; Dickenmann, M.; Schaub, S.; Steiger, J.; Rüfli, T.; Halter, J.; Hopfer, H.; Holbro, A.; Hirt-Minkowski, P. Daratumumab for Treatment of Antibody-Mediated Rejection after ABO-Incompatible Kidney Transplantation. Case Rep. Nephrol. Dial. 2019, 9, 149–157. [Google Scholar] [CrossRef]
- Aguilera Agudo, C.; Gómez Bueno, M.; Krsnik Castello, I. Daratumumab for Antibody-Mediated Rejection in Heart Transplant-A Novel Therapy: Successful Treatment of Antibody-Mediated Rejection. Transplantation 2021, 105, E30–E31. [Google Scholar] [CrossRef]
- Doberer, K.; Kläger, J.; Gualdoni, G.A.; Mayer, K.A.; Eskandary, F.; Farkash, E.A.; Agis, H.; Reiter, T.; Reindl-Schwaighofer, R.; Wahrmann, M.; et al. CD38 Antibody Daratumumab for the Treatment of Chronic Active Antibody-Mediated Kidney Allograft Rejection. Transplantation 2021, 105, 451–457. [Google Scholar] [CrossRef]
- Fenton, M.; Shaw, K.; Murchan, H.; Duignan, S.; Dunne, E.; McMahon, C.J. Daratumumab Provides Transient Response of Antibody Mediated Rejection Post Pediatric Orthotopic Heart Transplantation. J. Heart Lung Transpl. 2022. S1053-2498(22)01995-7. [Google Scholar] [CrossRef]
- Du, Y.; Dai, Q.; Zhang, H.; Li, Q.; Song, K.; Fu, Y.; Min, W.; Liu, Z.; Li, R. CD38 Deficiency Downregulates the Onset and Pathogenesis of Collagen-Induced Arthritis through the NF-ΚB Pathway. J. Immunol. Res. 2019, 2019, 7026067. [Google Scholar] [CrossRef] [Green Version]
- Korver, W.; Carsillo, M.; Yuan, J.; Idamakanti, N.; Wagoner, M.; Shi, P.; Xia, C.Q.; Smithson, G.; McLean, L.; Zalevsky, J.; et al. A Reduction in B, T, and Natural Killer Cells Expressing CD38 by TAK-079 Inhibits the Induction and Progression of Collagen-Induced Arthritis in Cynomolgus Monkeys. J. Pharmacol. Exp. Ther. 2019, 370, 182–196. [Google Scholar] [CrossRef]
- Mikhael, J.; Richter, J.; Vij, R.; Cole, C.; Zonder, J.; Kaufman, J.L.; Bensinger, W.; Dimopoulos, M.; Lendvai, N.; Hari, P.; et al. A Dose-Finding Phase 2 Study of Single Agent Isatuximab (Anti-CD38 MAb) in Relapsed/Refractory Multiple Myeloma. Leukemia 2020, 34, 3298–3309. [Google Scholar] [CrossRef]
- Moreau, P.; Dimopoulos, M.A.; Yong, K.; Mikhael, J.; Risse, M.-L.; Asset, G.; Martin, T. Isatuximab plus Carfilzomib/Dexamethasone versus Carfilzomib/Dexamethasone in Patients with Relapsed/Refractory Multiple Myeloma: IKEMA Phase III Study Design. Future Oncol. 2020, 16, 4347–4358. [Google Scholar] [CrossRef]
- Eslick, R.; Talaulikar, D. Multiple Myeloma: From Diagnosis to Treatment. Aust. Fam. Physician 2013, 42, 684–688. [Google Scholar]
- Sun, H.; Martin, T.G.; Marra, J.; Kong, D.; Keats, J.; Macé, S.; Chiron, M.; Wolf, J.L.; Venstrom, J.M.; Rajalingam, R. Individualized Genetic Makeup That Controls Natural Killer Cell Function Influences the Efficacy of Isatuximab Immunotherapy in Patients with Multiple Myeloma. J. ImmunoTherapy Cancer 2021, 9, 2958. [Google Scholar] [CrossRef]
- Kazandjian, D. Multiple Myeloma Epidemiology and Survival: A Unique Malignancy. Semin. Oncol. 2016, 43, 676–681. [Google Scholar] [CrossRef]
- Chari, A.; Martinez-Lopez, J.; Mateos, M.-V.; Bladé, J.; Benboubker, L.; Oriol, A.; Arnulf, B.; Rodriguez-Otero, P.; Pineiro, L.; Jakubowiak, A.; et al. Daratumumab plus Carfilzomib and Dexamethasone in Patients with Relapsed or Refractory Multiple Myeloma. Blood 2019, 134, 421–431. [Google Scholar] [CrossRef]
- Xu, X.S.; Moreau, P.; Usmani, S.Z.; Lonial, S.; Jakubowiak, A.; Oriol, A.; Krishnan, A.; Bladé, J.; Luo, M.; Sun, Y.N.; et al. Split First Dose Administration of Intravenous Daratumumab for the Treatment of Multiple Myeloma (MM): Clinical and Population Pharmacokinetic Analyses. Adv. Ther. 2020, 37, 1464. [Google Scholar] [CrossRef] [PubMed]
- Leleu, X.; Beksac, M.; Chou, T.; Dimopoulos, M.; Yoon, S.-S.; Prince, H.M.; Pour, L.; Shelekhova, T.; Chari, A.; Khurana, M.; et al. Efficacy and Safety of Weekly Carfilzomib (70 Mg/m(2)), Dexamethasone, and Daratumumab (KdD70) Is Comparable to Twice-Weekly KdD56 While Being a More Convenient Dosing Option: A Cross-Study Comparison of the CANDOR and EQUULEUS Studies. Leuk Lymphoma 2021, 62, 358–367. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of an Antibody | Mechanism of Action | References |
|---|---|---|
| Isatuximab | Allosteric inhibition of NADase activity | [14,21,62,65,66,67] |
| Daratumumab | Cytotoxic effect or clearance of CD38+ cells | [56,57,68] |
| TAK-079 (Takeda) | Cytotoxic effect or clearance of CD38+ cells | [21,63] |
| MOR-202 (Morphosys) | Cytotoxic effect or clearance of CD38+ cells | [21,69] |
| Affliction | Role of CD38 | Progression in Treatment | References |
|---|---|---|---|
| Acute myeloid leukaemia | Antiproliferative effect, autophagy induction | Several effective therapy attempts | [87,88] |
| Acute lymphoblastic leukaemia | Independent adverse prognostic factor in B-ALL Ph(−) | The promising target of a novel therapy for paediatric T-ALL patients | [97] |
| Chronic lymphoblastic leukaemia (CLL), Small lymphocytic lymphoma (SLL), B-cell prolymphocytic leukaemia (B-PLL) | Associated with advanced disease stage, higher incidence of lymphadenopathy, hepatomegaly, high-risk cytogenetics, short lymphocyte doubling time, short time to initiation of the first treatment, poor response to therapy, shorter progression-free survival | Prognostic marker–presence correlates with the worse course | [100] |
| NK-T cell lymphoma | Poor prognostic marker, CD31/CD38, promotes the activation and proliferation of different lymphocyte groups | Therapeutic target | [107] |
| Multiple myeloma | Expressed on plasma cells to a higher degree than physiologically | Anti-CD38 immunotherapy action, which leads to depletion of immunosuppressive cells and clonal expansion of cytotoxic T cells | [126] |
| Aplastic anaemia | Higher proinflammatory and proliferative capacity | Prognostic marker | [132] |
| Immune thrombocytopenic purpura | CD38+ Breg cells induce peripheral tolerance and diminish the function of autoreactive T-helper CD4+ cells via the production of IL-10 | Prognostic marker presence correlates with the worse course | [133] |
| Immunoglobin light-chain amyloidosis | Clonal expansion of CD38+ plasma cells that produce misfolded immunoglobulin light chains | Daratumumab registered in combination with VCd in the treatment of AL amyloidosis | [136] |
| Burkitt lymphoma | CD38+ cells regulate the potential of CD8+ cells involved in the pathogenesis of the disease | Prognostic marker | [137,138,140,141,142,143,144,145,146,147] |
| Evans syndrome | |||
| Lymphoplasmacytic lymphoma | |||
| MYC translocated aggressive B-cells lymphoma | |||
| ALK-positive large B-cells lymphoma | |||
| High-grade B-cell lymphoma (HGBL) | |||
| Plasmablastic lymphoma | |||
| Plasmablastic plasma cell myeloma | |||
| Anaplastic large cell lymphoma | |||
| Primary effusion diffused large B-cell lymphoma |
| Affliction | Role of CD38 | References |
|---|---|---|
| Non-hematological malignancies | Nasopharyngeal cancer (NPC): cell aging inhibition, cell proliferation promotion, cell metastasis, and conversion to the S phase; increase in the concentration of ATP, lactic acid, cAMP, and human ADP/acrp30 concentration in NPC cells, regulation of the metabolic associated signaling pathways associated with tumor protein 53 (TP53), hypoxia-inducible factor 1α (HIF-1α) and sirtuin 1 prostate cancer (PC): progression to castration-resistant PC, OS shortening, upregulation of excretion of IL-12, IL-23, and IL-27, immunosuppressive adenosine signaling and T cell exhaustion signatures, decrease in glycolytic and mitochondrial metabolism, activating AMP-activated protein kinase and inhibiting fatty acid and lipid synthesis colorectal cancer: expansion of CD38+ monocytic and polymorphonuclear myeloid-derived suppressor cells (M-MDSCs and PMN-MDSCs) with a tendency to increase CD38 expression on M- and PMN-MDSCs glioma: impact on the intracellular ATP levels and the survival of C6 glioma cells | [29,72,148,149,151,169,170] |
| AIDS | Sensitizing cells to HIV infection and enhancing HIV replication | [150] |
| COVID-19 | CD38-mediated thrombosis, enhancing symptoms of SARS-CoV-2 infection | [159,160] |
| Gastrointestinal diseases | Expression is visibly higher in patients with ulcerative colitis and Crohn’s disease | [168] |
| Pulmonary injury induced by Escherichia coli | Activation of MAPK/NF-kB signaling pathway | [170] |
| Neurological diseases | Controlling NAD bioavailability in the brain; expression crucial in epileptogenesis | [174] |
| Systemic lupus erythematosus | Undermining cytotoxic function of CD8+ T lymphocytes | [185,186] |
| Transplant rejection | Biomarker in renal transplant recipients with antibody-mediated acute rejection; promotion of disease progression | [188] |
| Collagen-induced arthritis | Regulation of collagen-induced arthritis | [185,194] |
| Identifier | Patients Number | Recruitment Status | Condition or Disease | Target Antigen | Therapy Protocol | Short Description |
|---|---|---|---|---|---|---|
| NCT01084252 | 351 | Active, not recruiting | Hematological Malignancy | CD38 | Drug: Isatuximab SAR650984 Drug: Dexamethasone | Multiple Intravenous Administrations of a Humanized Monoclonal Antibody (SAR650984) Against CD38 in Patients with Selected CD38+ Hematological Malignancies |
| NCT02332850 | 89 | Recruiting | Multiple Myeloma | CD38 | Biological: Isatuximab Drug: Carfilzomib Drug: Dexamethasone | SAR650984 in Combination with Carfilzomib for Treatment of Relapsed or Refractory Multiple Myeloma |
| NCT01749969 | 60 | Active, not recruiting | Plasma Cell Myeloma | CD38 | Drug: isatuximab SAR650984, lenalidomide and dexamethasone | SAR650984 (Isatuximab), Lenalidomide, and Dexamethasone in Combination in RRMM Patients |
| NCT04763616 | 37 | Recruiting | Natural Killer/T-cell Lymphoma, Relapsed Natural Killer/T-cell Lymphoma, Refractory Natural Killer/T-cell Lymphoma | CD38 | Drug: Isatuximab Drug: Cemiplimab | Isatuximab and Cemiplimab in Relapsed or Refractory Natural Killer/T-cell Lymphoid Malignancy (ICING) |
| NCT04751877 | 270 | Recruiting | Multiple Myeloma, Myeloma | CD38 | Drug: Isatuximab Drug: Lenalidomide Drug: Bortezomib Drug: Dexamethasone | Isatuximab + Lenalidomide + Dexamethasone With/Without Bortezomib in de Novo Non-Frail NTE Multiple Myeloma Elderly Patients |
| NCT02513186 | 90 | Active, not recruiting | Plasma Cell Myeloma | CD38 | Drug: isatuximab SAR650984 Drug: lenalidomide Drug: bortezomib Drug: cyclophosphamide Drug: dexamethasone | Isatuximab in combination with CyBorD in Newly Diagnosed Multiple Myeloma (MM) |
| NCT00574288 | 104 | Completed | Multiple Myeloma | CD38 | Drug: Part 1: Daratumumab Drug: Part 2: Daratumumab Other: Methylprednisolone Other: Dexamethasone | Daratumumab Safety Study in Multiple Myeloma |
| NCT01998971 | 242 | Active, not recruiting | Multiple Myeloma | CD38 | Drugs: Daratumumab, Velcade, Pomalidomide, and others | JNJ-54767414 (HuMax CD38) (Anti-CD38 Monoclonal Antibody) in Combination with Backbone Treatments for the Treatment of Patients with Multiple Myeloma |
| NCT02195479 | 706 | Active, not recruiting | Multiple Myeloma | CD38 | Drug: Velcade Drug: Melphalan Drug: Prednisone Drug: Daratumumab IV Drug: Dexamethasone Drug: Daratumumab SC | Combination of Daratumumab and Velcade (Bortezomib) Melphalan-Prednisone (DVMP) Compared to Velcade Melphalan-Prednisone (VMP) in Participants with Previously Untreated Multiple Myeloma |
| NCT03236428 | 42 | Active, not recruiting | Monoclonal Gammopathy, Smoldering Multiple Myeloma | CD38 | Drug: Daratumumab | CD38 Antibody Daratumumab in Patients with High-Risk MGUS and Low-Risk Smoldering Multiple Myeloma |
| NCT05182073 | 168 | Recruiting | Multiple Myeloma, Myeloma | CD38 | Drug: FT576 Drug: Cyclophosphamide Drug: Fludarabine Drug: Daratumumab | FT576 as Monotherapy and in Combination with Daratumumab in Subjects with Relapsed/Refractory Multiple Myeloma |
| NCT04430530 | 100 | Recruiting | CD19 Negative B-cell Malignancies | CD38, CD22, CD123, CD10, CD20 | Biological: Infusion of 4SCAR-T specific to CD22/CD123/CD38/CD10/CD20 | 4SCAR-T Therapy Post CD19-targeted Immunotherapy |
| NCT03767751 | 80 | Recruiting | Multiple Myeloma | CD38, BCMA | Biological: Dual Specificity CD38 and BCMA CAR-T Cells | Dual Specificity CD38 and BCMA CAR-T Cell Immunotherapy for Relapsed or Refractory Multiple Myeloma |
| NCT03754764 | 80 | Recruiting | Relapsed B-cell Acute Lymphoblastic Leukaemia After CD19 CAR-T ACI | CD38 | Biological: Specificity CD38 CAR-T Cells | CD38 CAR-T Cell Immunotherapy for Relapsed B-cell Acute Lymphoblastic Leukaemia After CD19 CAR-T Adoptive Cellular Immunotherapy |
| NCT03464916 | 72 | Active, not recruiting | Relapsed or Refractory Multiple Myeloma | CD38 | Biological: CAR2 Anti-CD38 A2 CAR-T Cells | CAR2 Anti-CD38 A2 CAR-T Cells in patients with Relapsed or Refractory Multiple Myeloma |
| NCT03439280 | 100 | Active, not recruiting | Relapsed/Refractory Multiple Myeloma | CD38 | Drug: TAK-079 Drug: Pomalidomide Drug: Dexamethasone | TAK-079 Administered Subcutaneously as a Single Agent in Participants with Relapsed/Refractory (r/r) Multiple Myeloma (MM) |
| NCT03309111 | 197 | Recruiting | Relapsed/Refractory Multiple Myeloma | CD38, CD3 | Biological: ISB 1342 | Single-Agent ISB 1342 in Subjects with Previously Treated Multiple Myeloma |
| NCT04000282 | 78 | Recruiting | Plasma Cell Myeloma | CD38 | Drug: SAR442085 | SAR442085 in Patients with Relapsed or Refractory Multiple Myeloma (RRMM) |
| NCT01421186 | 91 | Completed | Multiple Myeloma | CD38 | Drug: MOR03087 phase 1 dose escalation Drug: MOR03087 Drug: Dexamethasone Drug: Pomalidomide Drug: Lenalidomide | Human Anti-CD 38 Antibody MOR03087 (MOR202) in Relapsed/Refractory Multiple Myeloma |
| NCT04466475 | 24 | Not yet recruiting | Plasma Cell Myeloma | CD38 | Biological: Astatine At 211 Anti-CD38 Monoclonal Antibody OKT10-B10 Drug: Melphalan Procedure: Peripheral Blood Stem Cell Transplantation | Anti-CD38 Monoclonal Antibody (211At-OKT10-B10) Combined with Melphalan as Conditioning Before Autologous Hematopoietic Cell Transplantation for Patients with Multiple Myeloma |
| NCT02136134 | 500 | Active, not recruiting | Relapsed or Refractory Multiple Myeloma | CD38 | Bortezomib and dexamethasone with or without daratumumab | An approach with the addition of daratumumab resulted in longer PFS and was linked with infusion-related reactions and higher rates of neutropenia and thrombocytopenia |
| NCT02076009 | 570 | Active, not recruiting | Relapsed or Refractory Multiple Myeloma | CD38 | Dexamethasone and lenalidomide with or without daratumumab | Adding daratumumab to the treatment increased PFS, the occurrence of infusion-related reactions, and a higher rate of neutropenia |
| Identifier | Number of Patients | Status | Drugs Used | Disease | Short Description |
|---|---|---|---|---|---|
| NCT04251065 | 8 | Active, not recruiting | Daratumumab, Gemcitabine, Dexamethasone, and Cisplatin | Relapsed or refractory T-Cell Lymphoma | |
| NCT04972942 | 40 (up to 39 years old) | Not yet recruiting | Daratumumab after total body irradiation (TBI)-based myeloablative conditioning and allogeneic hematopoietic cell transplantation | T-ALL | |
| NCT03432741 | 39 | Recruiting | Direct tumor microinjection (into the skin or in lymph nodes) with drugs such as Belinostat, Carfilzomib, Copanlisib Hydrochloride, Daratumumab, Fludeoxyglucose F-18, Gemcitabine Hydrochloride, Nivolumab, Obinutuzumab, Pembrolizumab, Rituximab, Romidepsin. Trastuzumab. Patients will be also undergoing FDG-PET with | non-Hodgkin lymphoma, Hodgkin lymphoma, | |
| NCT03384654 | 47 (up to 30 years old) | Active, not recruiting | Daratumumab, vincristine, prednisone. Doxorubicin, Peg-asparaginase, cyclophosphamide, cytarabine, 6-mercaptopurine, methotrexate | B-cell ALL/LL, T-cell ALL/LL | |
| NCT04045028 | 60 | recruiting | Tiragolumab, Daratumumab, Rituximab, Atezolizumab | Relapsed or Refractory B-Cell non-Hodgkin lymphoma | |
| NCT04136756 | 118 | Recruiting | NKTR-225, NKTR-255 Q21, Rituximab, Daratumumab, | relapsed/refractory multiple myeloma and Non-Hodgkin’s Lymphoma, indolent Non-Hodgkin’s Lymphoma | |
| NCT01592370 | 316 | Active, not recruiting | Nivolumab, Ipilimumab, Lirilumab, Pomalidomide, Dexamethasone. Daratumumab | Non-Hodgkin’s lymphoma, Hodgkin lymphoma | Nivolumab showed significant activity against B and T-cell lymphomas, while also being well-tolerated |
| NCT04017130 | 198 | Recruiting | MT-0169 | RRMM, RR NHL | |
| NCT03860844 | 96 (up to 17 years old) | Recruiting | Montelukast, Isatuximab, Dexamethasone, Fludarabine, Cytarabine, Liposomal daunonrubicin, daunorubicin, idarubicin, filgrastim, mitoxantrone. Doxorubicin, vincristine, PEG asparaginase, cyclophosphamide, etoposide, methotrexate, L-asparaginase, hydroxyurea, tocilizumab, L-asparaginase (erwinase) | Acute lymphoblastic leukaemia (ALL) and acute myeloblastic leukaemia (AML) | |
| NCT02999633 | 14 | Terminated | Isatuximab, dexamethasone, acetaminophen, ranitidine, diphenhydramine | RR T-ALL or T-LL | The trial was terminated due to an especially high benefit/risk ratio. |
| Identifier | Number of Patients | Status | Drugs Used | Disease Treated |
|---|---|---|---|---|
| NCT04714372 | 50 | Recruiting | Daratumumab, FT538, Fludarabine, Cyclophosphamide | AML |
| NCT02807558 | 155 | Active, not recruiting | Tamibarotene, azacitidine, daratumumab | AML, Myelodysplastic syndrome |
| NCT04614636 | 105 | Recruiting | FT538, Cyclophosphamide, Fludarabine, Daratumumab, Elotuzumab | AML |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szlasa, W.; Czarny, J.; Sauer, N.; Rakoczy, K.; Szymańska, N.; Stecko, J.; Kołodziej, M.; Kaźmierczak, M.; Barg, E. Targeting CD38 in Neoplasms and Non-Cancer Diseases. Cancers 2022, 14, 4169. https://doi.org/10.3390/cancers14174169
Szlasa W, Czarny J, Sauer N, Rakoczy K, Szymańska N, Stecko J, Kołodziej M, Kaźmierczak M, Barg E. Targeting CD38 in Neoplasms and Non-Cancer Diseases. Cancers. 2022; 14(17):4169. https://doi.org/10.3390/cancers14174169
Chicago/Turabian StyleSzlasa, Wojciech, Jakub Czarny, Natalia Sauer, Katarzyna Rakoczy, Natalia Szymańska, Jakub Stecko, Maksymilian Kołodziej, Maciej Kaźmierczak, and Ewa Barg. 2022. "Targeting CD38 in Neoplasms and Non-Cancer Diseases" Cancers 14, no. 17: 4169. https://doi.org/10.3390/cancers14174169
APA StyleSzlasa, W., Czarny, J., Sauer, N., Rakoczy, K., Szymańska, N., Stecko, J., Kołodziej, M., Kaźmierczak, M., & Barg, E. (2022). Targeting CD38 in Neoplasms and Non-Cancer Diseases. Cancers, 14(17), 4169. https://doi.org/10.3390/cancers14174169