
SPP1 Derived from Macrophages Is Associated with a Worse Clinical Course and Chemo-Resistance in Lung Adenocarcinoma

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. In Situ Hybridization (ISH)
2.3. IHC
2.4. Cell Culture of Macrophages and Cancer Cell Lines
2.5. 3D Cell Culture
2.6. Quantitative Real-Time Reverse Transcription PCR (qRT-PCR) Analysis
2.7. Western Blot Analysis
2.8. ELISA for SPP1
2.9. RNA-Sequencing (RNA-Seq) Transcriptome Analysis
2.10. Statistical Analysis
3. Results
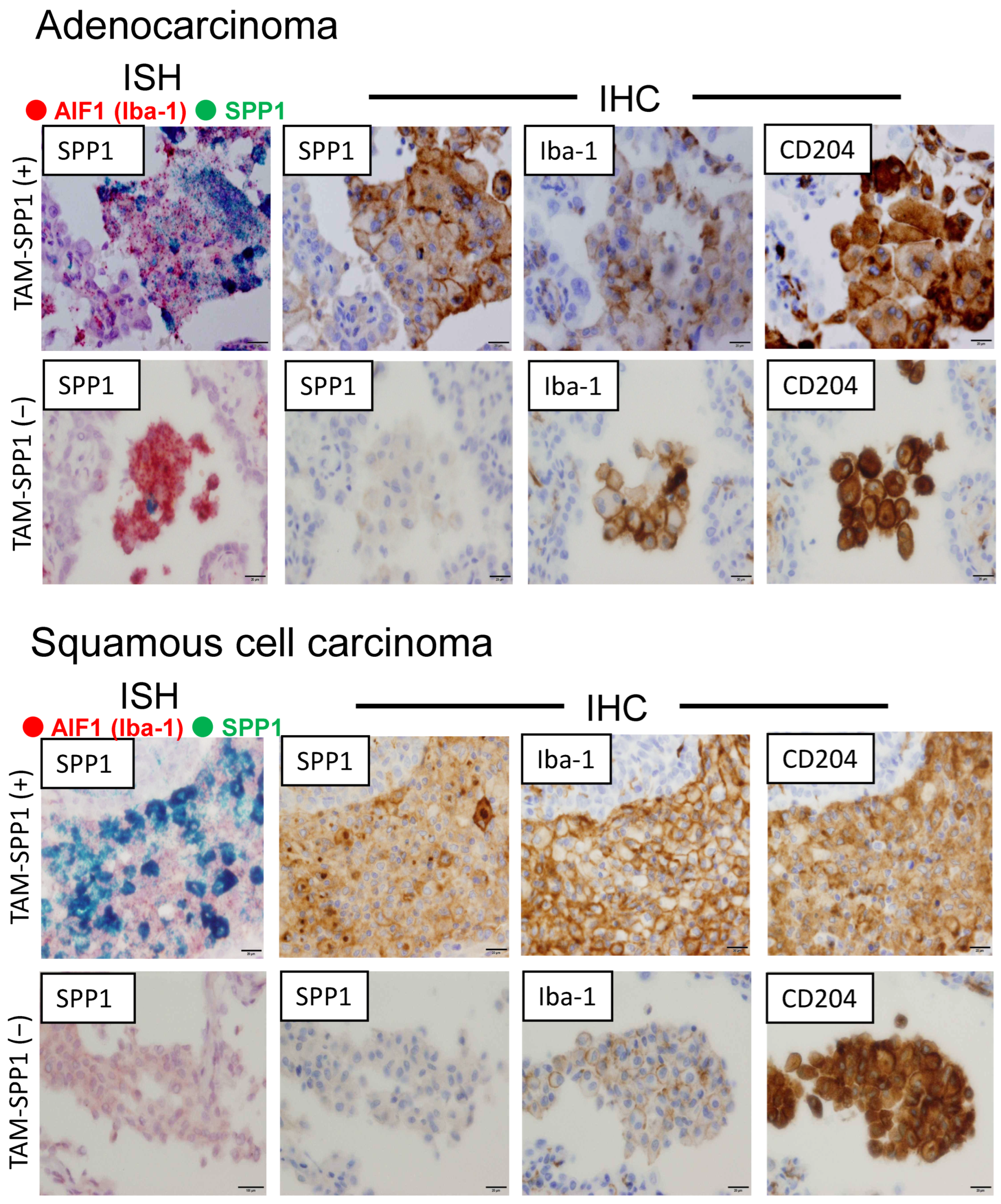
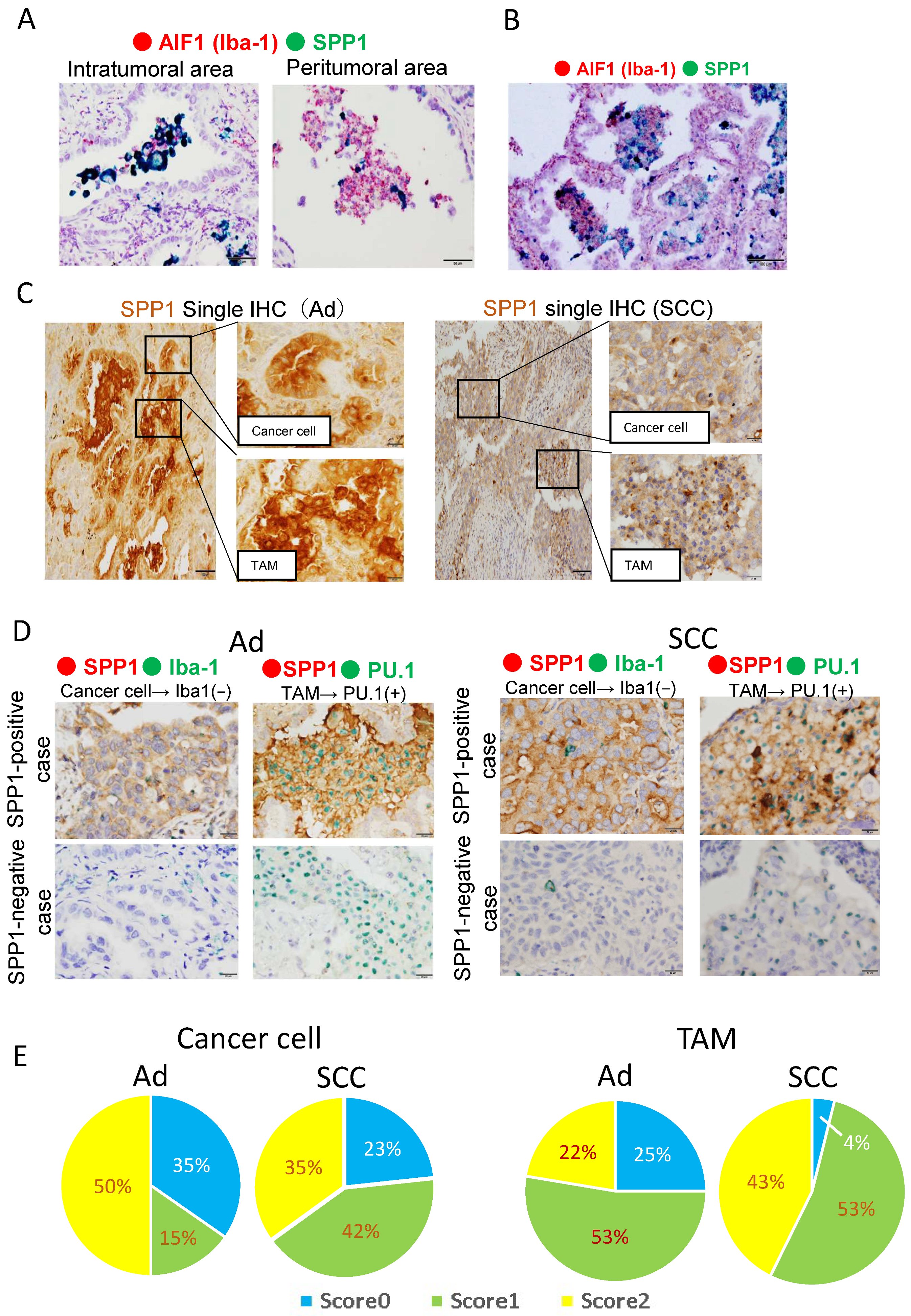
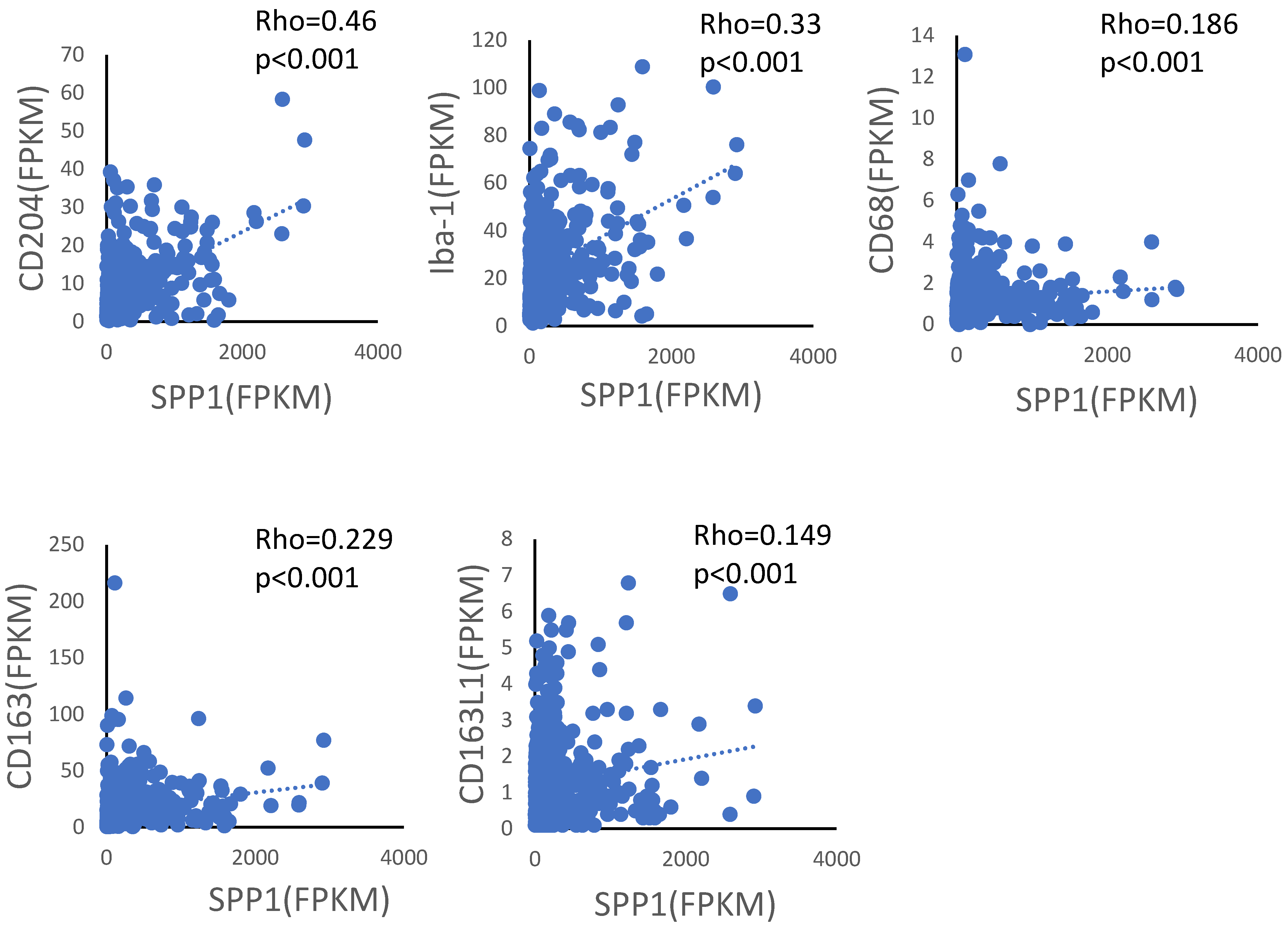
3.1. SPP1 Is Expressed on TAMs Infiltrating the Tumor, but Not on Alveolar Macrophages in Non-Tumor Areas
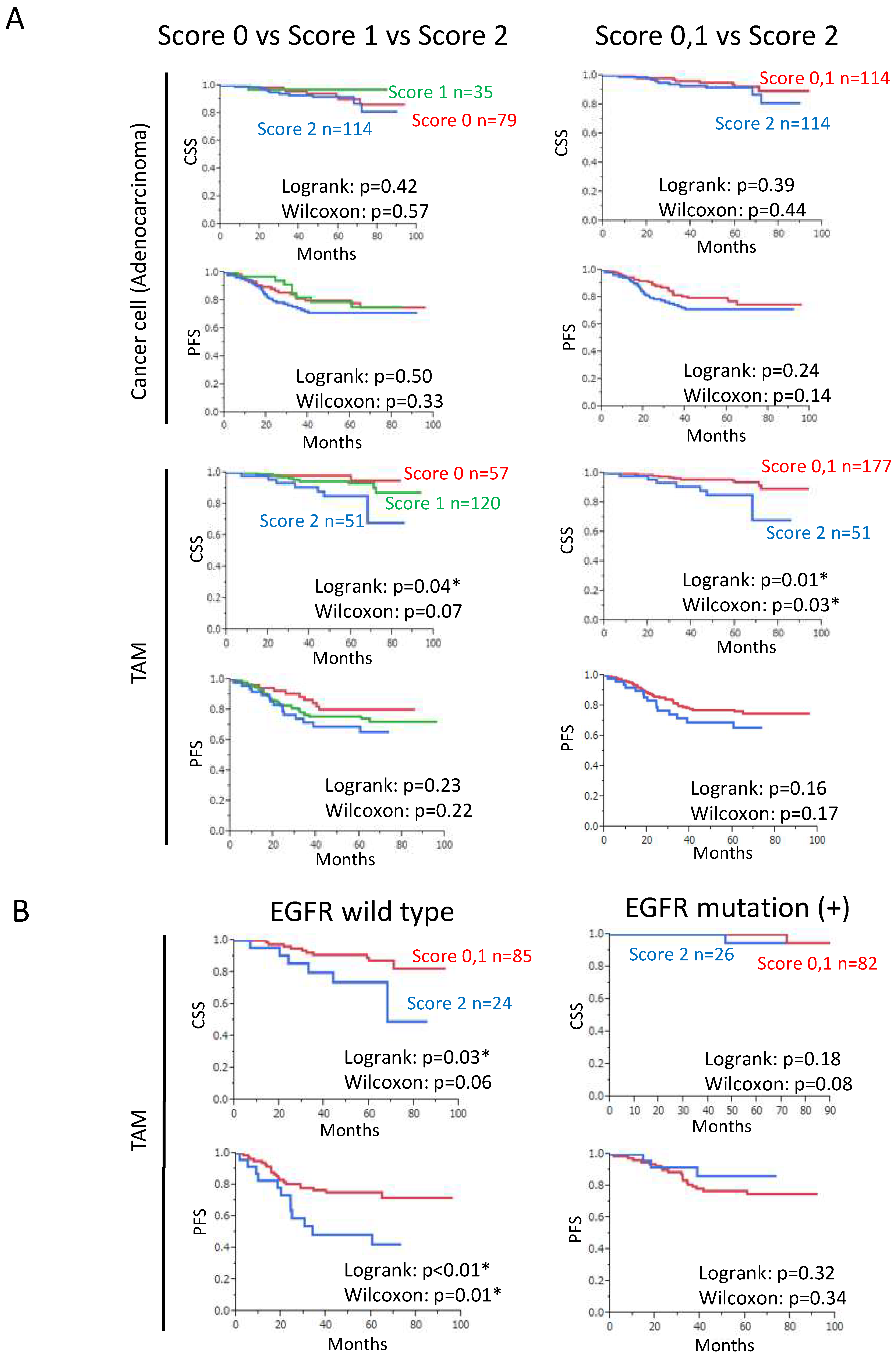
3.2. Increased SPP1 Expression on TAMs Is Associated with a Worse Clinical Course in EGFR-Wild-Type Adenocarcinoma
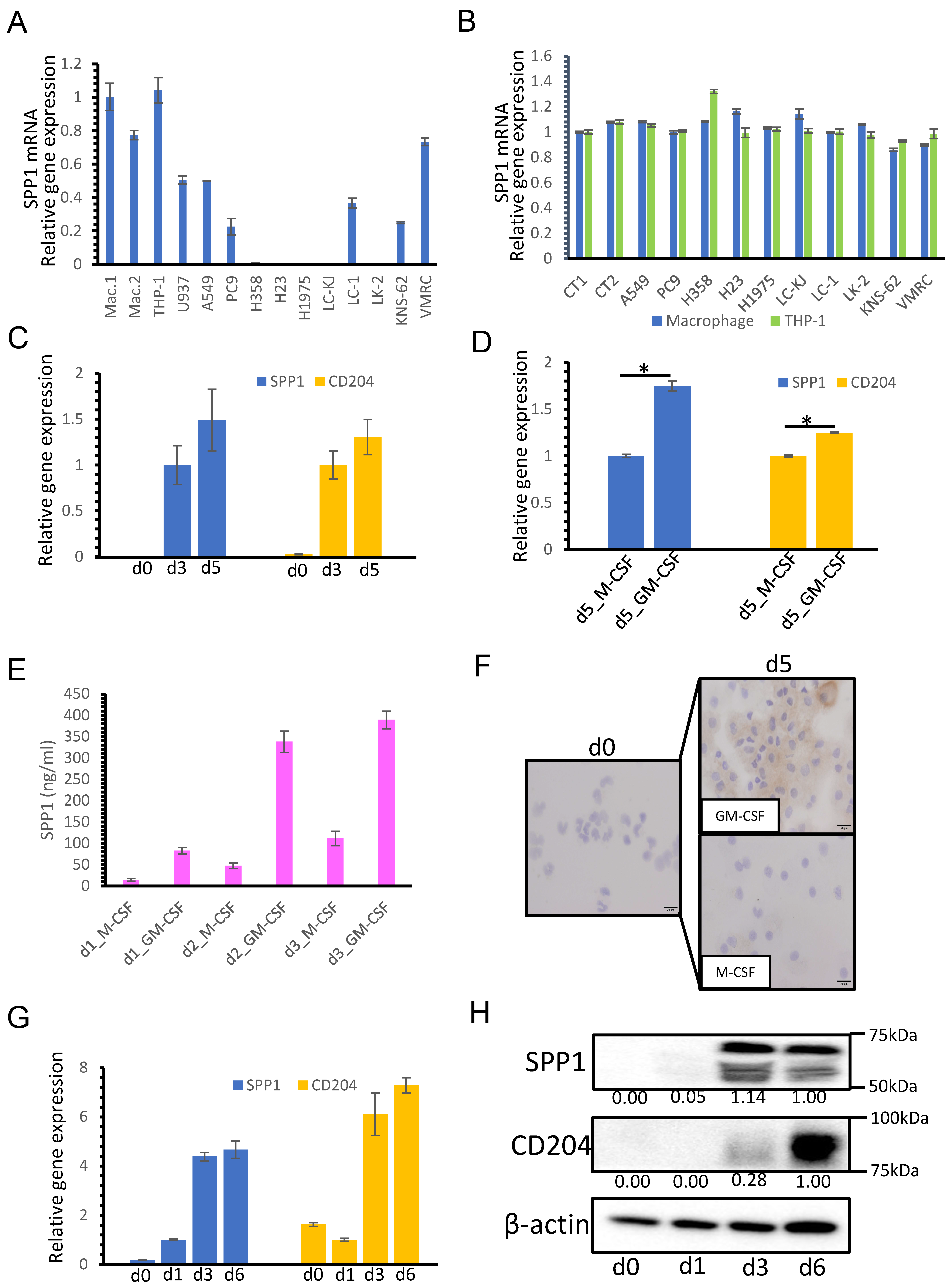
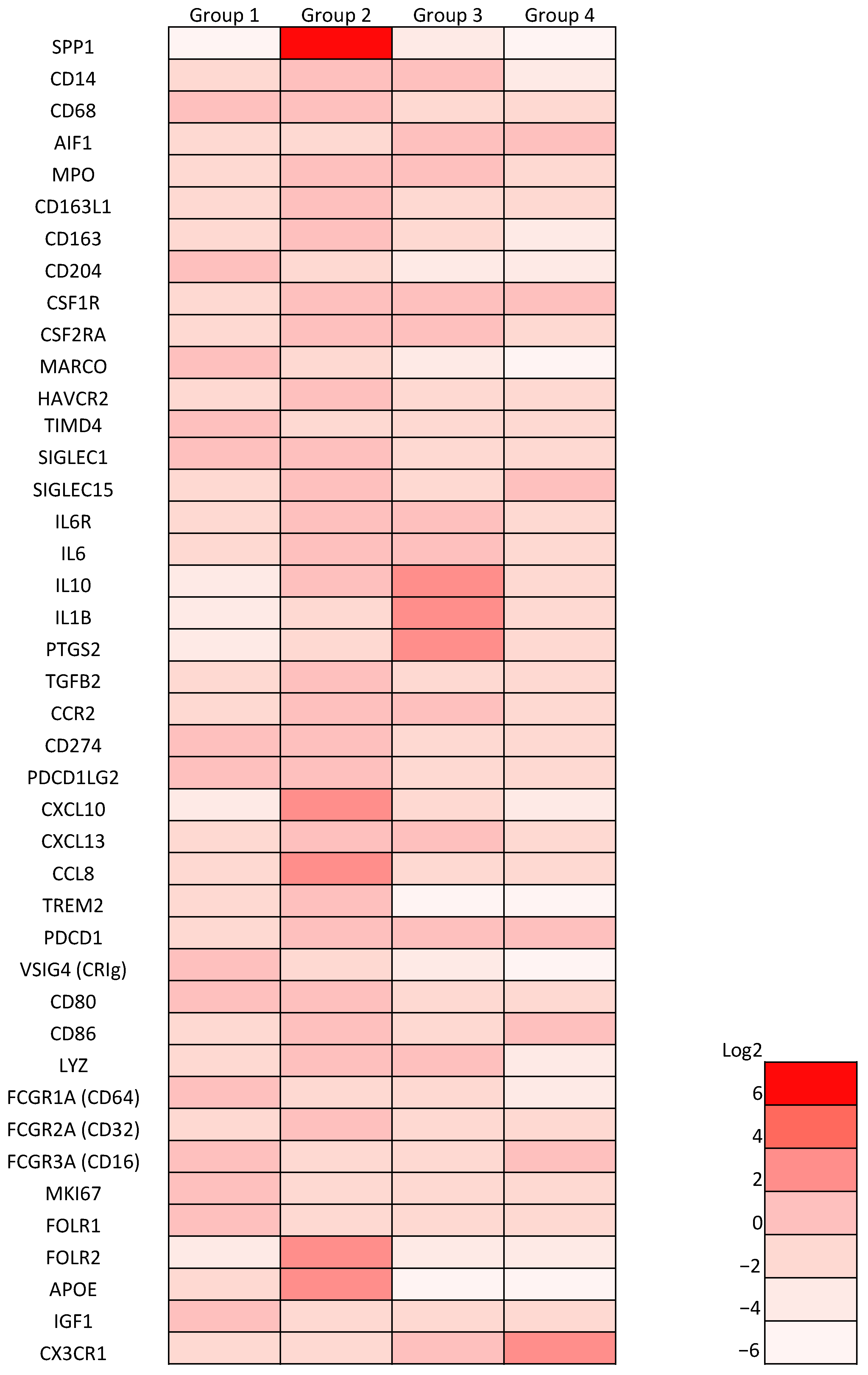
3.3. SPP1 Expression on Macrophages Is Dependent on GM-CSF-Mediated Macrophage Differentiation
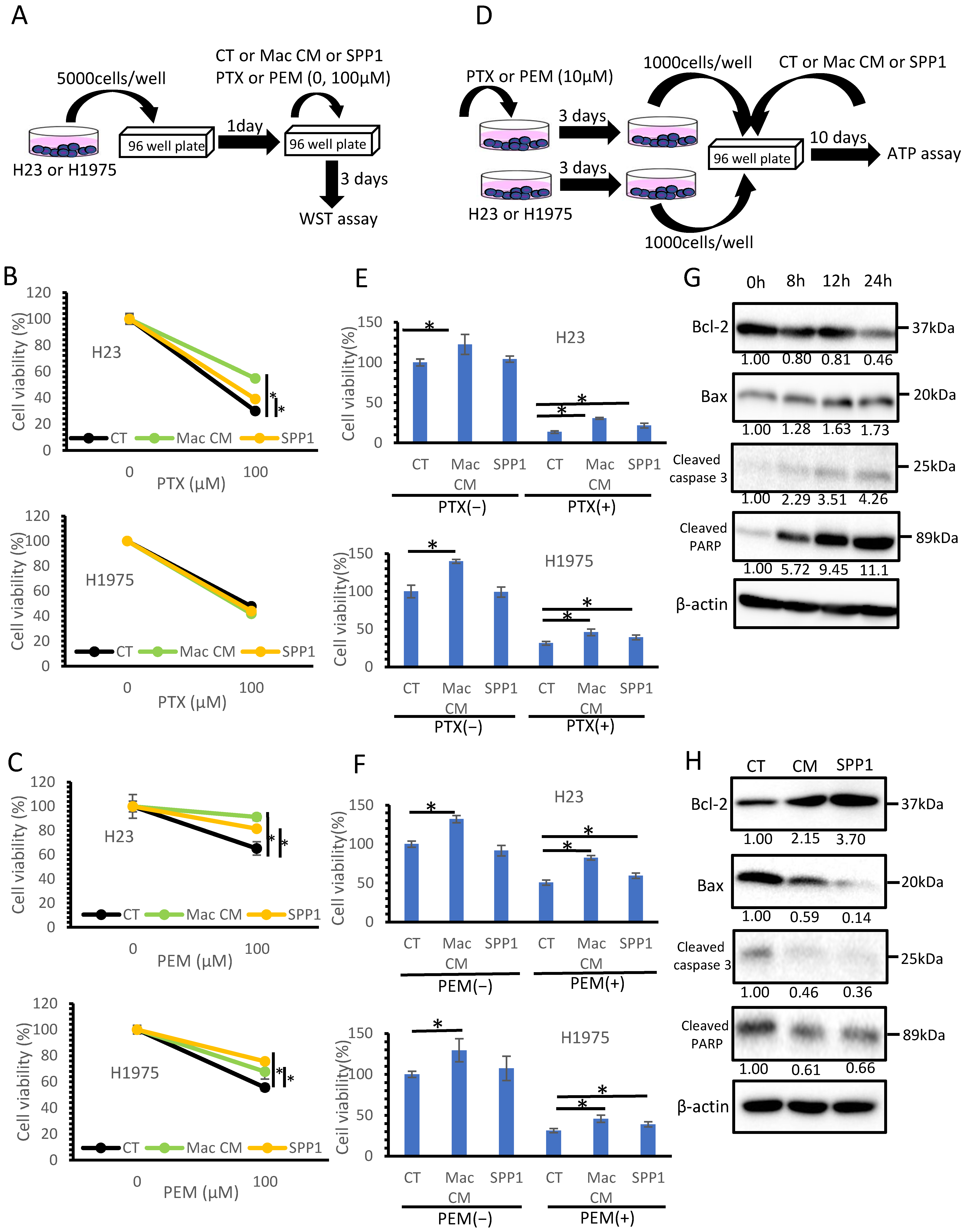
3.4. Macrophage-Derived SPP1 Potentially Contributes to Chemoresistance in Lung Cancer
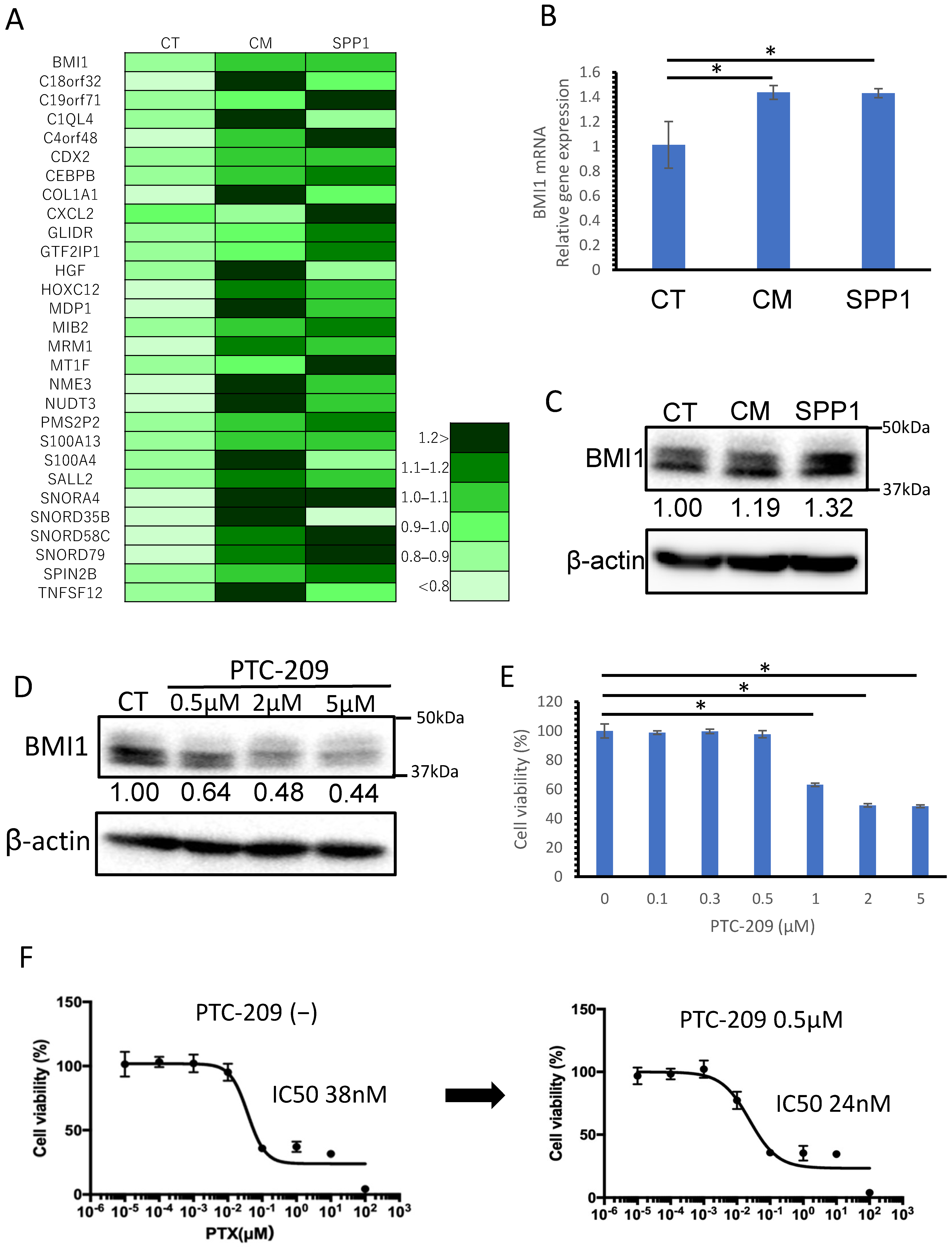
3.5. BMI1, Potentially Induced by TAM-Derived SPP1, Was Considered to Be One of the Factors Involved in Chemoresistance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goya, T.; Asamura, H.; Yoshimura, H.; Kato, H.; Shimokata, K.; Tsuchiya, R.; Sohara, Y.; Miya, T.; Miyaoka, E. Prognosis of 6644 resected non-small cell lung cancers in Japan: A Japanese lung cancer registry study. Lung Cancer 2005, 50, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Asamura, H.; Goya, T.; Koshiishi, Y.; Sohara, Y.; Eguchi, Y.; Mori, M.; Nakanishi, Y.; Tsuchiya, R.; Shimokata, K.; Inoue, H.; et al. A Japanese Lung Cancer Registry study: Prognosis of 13,010 resected lung cancers. J. Thorac. Oncol. 2008, 3, 46–52. [Google Scholar] [CrossRef]
- Hoffman, P.C.; Mauer, A.M.; Vokes, E.E. Lung cancer. Lancet 2000, 355, 479–485. [Google Scholar] [CrossRef]
- Altorki, N.; Kent, M.; Pasmantier, M. Detection of early-stage lung cancer: Computed tomographic scan or chest radiograph? J. Thorac. Cardiovasc. Surg. 2001, 121, 1053–1057. [Google Scholar] [CrossRef] [PubMed]
- Mountain, C.F. Revisions in the International System for Staging Lung Cancer. Chest 1997, 111, 1710–1717. [Google Scholar] [CrossRef]
- Lamort, A.S.; Giopanou, I.; Psallidas, I.; Stathopoulos, G. Osteopontin as a Link between Inflammation and Cancer: The Thorax in the Spotlight. Cells 2019, 8, 815. [Google Scholar] [CrossRef]
- Shi, L.; Wang, X. Role of osteopontin in lung cancer evolution and heterogeneity. Semin. Cell Dev. Biol. 2017, 64, 40–47. [Google Scholar] [CrossRef]
- Wei, J.; Marisetty, A.; Schrand, B.; Gabrusiewicz, K.; Hashimoto, Y.; Ott, M.; Grami, Z.; Kong, L.Y.; Ling, X.; Caruso, H.; et al. Osteopontin mediates glioblastoma-associated macrophage infiltration and is a potential therapeutic target. J. Clin. Investig. 2019, 129, 137–149. [Google Scholar] [CrossRef]
- Kariya, Y.; Kariya, Y. Osteopontin in Cancer: Mechanisms and Therapeutic Targets. Int. J. Transl. Med. 2022, 2, 419–447. [Google Scholar] [CrossRef]
- Zhao, H.; Chen, Q.; Alam, A.; Cui, J.; Suen, K.C.; Soo, A.P.; Eguchi, S.; Gu, J.; Ma, D. The role of osteopontin in the progression of solid organ tumour. Cell Death Dis. 2018, 9, 356. [Google Scholar] [CrossRef]
- Icer, M.A.; Gezmen-Karadag, M. The multiple functions and mechanisms of osteopontin. Clin. Biochem. 2018, 59, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Shinriki, S.; Jono, H.; Ueda, M.; Nagara, M.; Guo, J.; Hayashi, M.; Yoshida, R.; Ota, T.; Ota, K. Osteopontin-integrin αvβ3 axis is crucial for 5-fluorouracil resistance in oral squamous cell carcinoma. FEBS Lett. 2015, 589, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Bai, C.; Guo, Z. The prognostic value of osteopontin in limited-stage small cell lung cancer patients and its mechanism. Oncotarget 2017, 8, 70084–70096. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, J.; Liu, H.; Bi, J.R.; Liu, Y.; Chen, Y.Y.; Cao, J.Y.; Lu, Y.J. The association between osteopontin and survival in non-small-cell lung cancer patients: A meta-analysis of 13 cohorts. OncoTargets Ther. 2015, 8, 3513–3521. [Google Scholar] [CrossRef]
- Peng, B.; Wang, Y.H.; Huang, Z.; Feng, S.J.; Wang, Y.S. Prognostic significance of osteopontin in patients with lung cancer: A meta-analysis. Int. J. Clin. Exp. Med. 2014, 7, 4616–4626. [Google Scholar] [PubMed]
- Shi, S.M.; Su, Z.B.; Zhao, J.J.; Yu, D.J.; Tu, J.W.; Zhu, J.Q.; Zhao, J.P.; Sheng, L.; Wang, S.B.; Sheng, Y.J.; et al. Increased osteopontin protein expression may be correlated with poor prognosis in non-small-cell lung cancer: A meta analysis. J. Cancer Res. Ther. 2016, 12, 277–282. [Google Scholar] [CrossRef]
- Li, Y.; Sun, B.S.; Pei, B.; Li, C.G.; Zhang, Z.F.; Yin, Y.S.; Wang, C.L. Osteopontin-Expressing Macrophages in Non-Small Cell Lung Cancer Predict Survival. Ann. Thorac. Surg. 2015, 99, 1140–1148. [Google Scholar] [CrossRef]
- Zhang, Y.; Du, W.; Chen, Z.; Xiang, C. Upregulation of PD-L1 by SPP1 mediates macrophage polarization and facilitates immune escape in lung adenocarcinoma. Exp. Cell Res. 2017, 359, 449–457. [Google Scholar] [CrossRef]
- Komohara, Y.; Fujiwara, Y.; Ohnishi, K.; Takeya, M. Tumor-associated macrophages: Potential therapeutic targets for anti-cancer therapy. Adv. Drug Deliv. Rev. 2016, 99, 180–185. [Google Scholar] [CrossRef]
- Lin, C.N.; Wang, C.J.; Chao, Y.J.; Lai, M.D.; Shan, Y.S. The significance of the co-existence of osteopontin and tumor-associated macrophages in gastric cancer progression. BMC Cancer 2015, 15, 128. [Google Scholar] [CrossRef] [Green Version]
- Olson, O.; Kim, H.; Quail, D.F.; Foley, E.A.; Joyce, J.A. Tumor-Associated Macrophages Suppress the Cytotoxic Activity of Antimitotic Agents. Cell Rep. 2017, 19, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Goldstraw, P.; Chansky, K.; Crowley, J.; Rami-Porta, R.; Asamura, H.; Eberhardt, W.E.E.; Nicholson, A.G.; Groome, P.; Mitchell, A.; Bolejack, V.; et al. The IASLC Lung Cancer Staging Project: Proposals for Revision of the TNM Stage Groupings in the Forthcoming (Eighth) Edition of the TNM Classification for Lung Cancer. J. Thorac. Oncol. 2016, 11, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Shinchi, Y.; Komohara, Y.; Yonemitsu, K.; Sato, K.; Ohnishi, K.; Saito, Y.; Fujiwara, Y.; Mori, T.; Shiraishi, K.; Ikeda, K.; et al. Accurate expression of PD-L1/L2 in lung adenocarcinoma cells: A retrospective study by double immunohistochemistry. Cancer Sci. 2019, 110, 2711–2721. [Google Scholar] [CrossRef] [PubMed]
- Shinchi, Y.; Ishizuka, S.; Komohara, Y.; Matsubara, E.; Mito, R.; Pan, C.; Yoshii, D.; Yonemitsu, K.; Fujiwara, Y.; Ikeda, K.; et al. The expression of PD-1 ligand1 on macrophages and its clinical impacts and mechanisms in lung adenocarcinoma. CancerImmunol. Immunother. 2022, 1–17. [Google Scholar] [CrossRef]
- Matsubara, E.; Komohara, Y.; Shinchi, Y.; Mito, R.; Fujiwara, Y.; Ikeda, K.; Shima, T.; Shimoda, M.; Kanai, Y.; Sakagami, T.; et al. CD163-positive cancer cells are a predictor of a worse clinical course in lung adenocarcinoma. Pathol. Int. 2021, 71, 666–673. [Google Scholar] [CrossRef] [PubMed]
- Kovaleva, O.V.; Rashidova, M.A.; Samoilova, D.V.; Podlesnaya, P.A.; Mochalnikova, V.V.; Gratchev, A. Immunosuppressive Phenotype of Esophagus Tumors Stroma. Anal. Cell. Pathol. 2020, 2020, 5424780. [Google Scholar] [CrossRef]
- Horlad, H.; Ma, C.; Yano, H.; Pan, C.; Ohnishi, K.; Fujiwara, Y.; Endo, S.; Kikukawa, Y.; Okuno, Y.; Matsuoka, M.; et al. An IL-27/Stat3 axis induces expression of programmed cell death 1 ligands (PD-L1/2) on infiltrating macrophages in lymphoma. Cancer Sci. 2016, 107, 1696–1704. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Kai, K.; Komohara, Y.; Esumi, S.; Fujiwara, Y.; Yamamoto, T.; Uekawa, K.; Ohta, K.; Takezaki, T.; Kuroda, J.; Shinojima, N.; et al. Macrophage/microglia-derived IL-1β induces glioblastoma growth via the STAT3/NF-κB pathway. Hum. Cell 2022, 35, 226–237. [Google Scholar] [CrossRef]
- Kanda, Y. Investigation of the freely available easy-to-use software ‘EZR‘ for medical statistics. Bone Marrow Transpl. 2013, 48, 452–458. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Wu, M.; Li, Y.; Chang, I.; Yuan, Q.; Ekimyan-Salvo, M.; Deng, P.; Yu, B.; Yu, Y.; Dong, J.; et al. Targeting BMI1+ Cancer Stem Cells Overcomes Chemoresistance and Inhibits Metastases in Squamous Cell Carcinoma. Cell Stem Cell 2017, 20, 621–634. [Google Scholar] [CrossRef] [PubMed]
- Paranjape, A.N.; Balaji, S.A.; Mandal, T.; Krushik, E.V.; Nagaraj, P.; Mukherjee, G.; Rangarajan, A. Bmi1 regulates self-renewal and epithelial to mesenchymal transition in breast cancer cells through Nanog. BMC Cancer 2014, 14, 785. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.T.; Chien, P.J.; Chen, S.H.; Sheu, G.T.; Jan, M.S.; Wang, B.Y.; Chang, W.W. BMI1-Mediated Pemetrexed Resistance in Non-Small Cell Lung Cancer Cells Is Associated with Increased SP1 Activation and Cancer Stemness. Cancers 2020, 12, 2069. [Google Scholar] [CrossRef] [PubMed]
- Sugihara, H.; Ishimoto, T.; Watanabe, M.; Sawayama, H.; Iwatsuki, M.; Baba, Y.; Komohara, Y.; Takeya, M.; Baba, H. Identification of miR-30e* regulation of Bmi1 expression mediated by tumor-associated macrophages in gastrointestinal cancer. PLoS ONE 2013, 8, e81839. [Google Scholar] [CrossRef]
- Choi, S.I.; Kim, S.Y.; Lee, J.H.; Kim, J.Y.; Cho, E.W.; Kim, I.G. Osteopontin production by TM4SF4 signaling drives a positive feedback autocrine loop with the STAT3 pathway to maintain cancer stem cell-like properties in lung cancer cells. Oncotarget 2017, 8, 101284–101297. [Google Scholar] [CrossRef]
- Inoue, M.; Shinohara, M. Intracellular osteopontin (iOPN) and immunity. Immunol. Res. 2011, 49, 160–172. [Google Scholar] [CrossRef]
- Shirakawa, K.; Endo, J.; Kataoka, M.; Katsumata, Y.; Yoshida, N.; Yamamoto, T.; Isobe, S.; Moriyama, H.; Goto, S.; Kitakata, H.; et al. IL (Interleukin)-10-STAT3-Galection-3 Axis Is Essential for Osteopontin-Producing Reparative Macrophage Polarization After Myocardial Infarction. Circulation 2018, 138, 2021–2035. [Google Scholar] [CrossRef]
- Shimodaira, T.; Matsuda, K.; Uchibori, T.; Sugano, M.; Uehara, T.; Honda, T. Upregulation of osteopontin expression via the interaction of macrophages and fibroblasts under IL-1b stimulation. Cytokine 2018, 110, 63–69. [Google Scholar] [CrossRef]
- Wang, M.; Han, J.; Marcar, L.; Black, J.; Liu, Q.; Li, X.; Nagulapalli, K.; Sequist, L.V.; Mak, R.H.; Benes, C.; et al. Radiation Resistance in KRAS-Mutated Lung Cancer Is Enabled by Stem-like Properties Mediated by an Osteopontin-EGFR Pathway. Cancer Res. 2017, 77, 2018–2028. [Google Scholar] [CrossRef]
- Li, Y.; Liu, H.; Zhao, Y.; Yue, D.; Chen, C.; Li, C.; Zhang, Z.; Wang, C. Tumor-associated macrophages (TAMs)-derived osteopontin (OPN) upregulates PD-L1 expression and predicts poor prognosis in non-small cell lung cancer (NSCLC). Thorac. Cancer 2021, 12, 2698–2709. [Google Scholar] [CrossRef]
- Casanova-Acebes, M.; Dalla, E.; Leader, A.M.; Leberichel, J.; Nikolic, J.; Morales, B.M.; Brown, M.; Chang, C.; Troncoso, L.; Chen, S.T.; et al. Tissue-resident macrophages provide a pro-tumorigenic niche to early NSCLC cells. Nature 2021, 595, 578–584. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | n (%); Ad | n (%); SCC |
|---|---|---|
| Age | ||
| <65 | 67 (29) | 20 (19) |
| ≧65 | 161 (71) | 83 (81) |
| Gender | ||
| Male | 114 (50) | 93 (91) |
| Female | 114 (50) | 10 (9) |
| B.I. | ||
| <600 | 142 (62) | 14 (13) |
| ≧600 | 86 (38) | 89 (87) |
| pStage (TNM 8th) | ||
| Stage 0 | 11 (5) | 0 (0) |
| Stage IA1 | 41 (18) | 6 (6) |
| Stage IA2 | 58 (26) | 22 (21) |
| Stage IA3 | 34 (15) | 17 (16) |
| Stage IB | 32 (14) | 7 (7) |
| Stage IIA | 3 (1) | 12 (12) |
| Stage IIB | 18 (8) | 19 (18) |
| Stage IIIA | 25 (11) | 14 (14) |
| Stage IIIB | 3 (1) | 6 (6) |
| Stage IV | 3 (1) | 0 (0) |
| Preoperative treatment | ||
| None | 223 (98) | 100 (97) |
| Chemoradiation | 5 (2) | 3 (3) |
| Adjuvant chemotherapy | ||
| None | 167 (73) | 74 (72) |
| UFT | 28 (12) | 10 (10) |
| Platinum based regimen | 33 (15) | 19 (18) |
| EGFR mutation | ||
| negative | 109 (48) | |
| positive | ||
| L858R | 54 (24) | |
| 19del | 46 (20) | |
| other | 8 (3) | |
| not submitted | 11 (5) |
| TAM-SPP1 Expression (Low: Score 0–1, High: Score 2) | ||||
|---|---|---|---|---|
| Low | High | p | ||
| Age | ||||
| <65 | 56 | 11 | 0.22 | |
| ≥65 | 121 | 40 | ||
| Gender | ||||
| Male | 88 | 26 | 1.00 | |
| Female | 89 | 25 | ||
| Smoking | ||||
| B.I. < 600 | 110 | 32 | 1.00 | |
| B.I. ≥ 600 | 67 | 19 | ||
| pStage | ||||
| 0–I | 137 | 39 | 1.00 | |
| II–VI | 40 | 12 | ||
| EGFR mutation | ||||
| negative | 85 | 24 | 0.84 | |
| positive | 82 | 26 | ||
| TAM-SPP1 Expression (Low: Score 0–1, High: Score 2) | ||||
|---|---|---|---|---|
| Low | High | p | ||
| Age | ||||
| <65 | 6 | 14 | 0.01 | |
| ≥65 | 53 | 30 | ||
| Gender | ||||
| Male | 54 | 39 | 0.74 | |
| Female | 5 | 5 | ||
| Smoking | ||||
| B.I. < 600 | 6 | 8 | 0.37 | |
| B.I. ≥ 600 | 53 | 36 | ||
| pStage | ||||
| 0–I | 29 | 23 | 0.90 | |
| II–VI | 30 | 21 | ||
| Univariate | Multivariate | ||||||
|---|---|---|---|---|---|---|---|
| p-Value | HR | 95% CI | p-Value | HR | 95% CI | ||
| Age | <65 vs. 65≦ | 0.6779 | 1.240 | 0.466–3.876 | |||
| Gender | Female vs. Male | 0.0060 | 4.098 | 1.468–14.46 | 0.6636 | 1.339 | 0.379–5.718 |
| Smoking | B.I. < 600 vs. 600 ≦ B.I. | 0.0049 | 3.871 | 1.502–11.12 | 0.7926 | 1.171 | 0.373–4.167 |
| pStage | pStage 0–I vs. pStage II–IV | <0.0001 | 14.59 | 5.222–51.56 | <0.0001 | 10.81 | 3.673–39.99 |
| EGFR | mutation vs. wild type | 0.0002 | 8.735 | 2.485–55.24 | 0.0086 | 5.773 | 1.494–38.31 |
| TAM-SPP1 | Low score vs. High score | 0.0335 | 3.008 | 1.096–7.735 | 0.0172 | 3.599 | 1.273–9.725 |
| Univariate | Multivariate | ||||||
|---|---|---|---|---|---|---|---|
| p-Value | HR | 95% CI | p-Value | HR | 95% CI | ||
| Age | <65 vs. 65≦ | 0.5983 | 1.170 | 0.661–2.188 | |||
| Gender | Female vs. Male | 0.0265 | 1.828 | 1.072–3.179 | 0.9458 | 0.978 | 0.519–1.862 |
| Smoking | B.I. < 600 vs. 600 ≦ B.I. | 0.0016 | 2.362 | 1.389–4.038 | 0.0826 | 1.735 | 0.931–3.284 |
| pStage | pStage0–I vs. pStage II–IV | <0.0001 | 9.886 | 5.712–17.57 | <0.0001 | 9.055 | 5.185–16.23 |
| EGFR | mutation vs. wild type | 0.0584 | 1.673 | 0.981–2.909 | |||
| TAM-SPP1 | Low score vs. High score | 0.1822 | 1.519 | 1.812–2.693 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsubara, E.; Komohara, Y.; Esumi, S.; Shinchi, Y.; Ishizuka, S.; Mito, R.; Pan, C.; Yano, H.; Kobayashi, D.; Fujiwara, Y.; et al. SPP1 Derived from Macrophages Is Associated with a Worse Clinical Course and Chemo-Resistance in Lung Adenocarcinoma. Cancers 2022, 14, 4374. https://doi.org/10.3390/cancers14184374
Matsubara E, Komohara Y, Esumi S, Shinchi Y, Ishizuka S, Mito R, Pan C, Yano H, Kobayashi D, Fujiwara Y, et al. SPP1 Derived from Macrophages Is Associated with a Worse Clinical Course and Chemo-Resistance in Lung Adenocarcinoma. Cancers. 2022; 14(18):4374. https://doi.org/10.3390/cancers14184374
Chicago/Turabian StyleMatsubara, Eri, Yoshihiro Komohara, Shigeyuki Esumi, Yusuke Shinchi, Shiho Ishizuka, Remi Mito, Cheng Pan, Hiromu Yano, Daiki Kobayashi, Yukio Fujiwara, and et al. 2022. "SPP1 Derived from Macrophages Is Associated with a Worse Clinical Course and Chemo-Resistance in Lung Adenocarcinoma" Cancers 14, no. 18: 4374. https://doi.org/10.3390/cancers14184374
APA StyleMatsubara, E., Komohara, Y., Esumi, S., Shinchi, Y., Ishizuka, S., Mito, R., Pan, C., Yano, H., Kobayashi, D., Fujiwara, Y., Ikeda, K., Sakagami, T., & Suzuki, M. (2022). SPP1 Derived from Macrophages Is Associated with a Worse Clinical Course and Chemo-Resistance in Lung Adenocarcinoma. Cancers, 14(18), 4374. https://doi.org/10.3390/cancers14184374